Fentanyl decreases blood oxygen more than furanylfentanyl despite similar effects on breathing
Catherine Demery, Sierra C. Moore, Kelsey E. Kochan, Mengchu Li, Thomas D. Prince, Benjamin M. Clements, Zhongqi Chen, Jessica R. Whitaker-Fornek, Erica S. Levitt, Jessica P. Anand, John R. Traynor

TL;DR
Fentanyl causes more severe drops in blood oxygen than furanylfentanyl, despite similar effects on breathing, highlighting differences in their risks.
Contribution
The study compares the in vivo effects of fentanyl and furanylfentanyl on respiration and oxygen saturation in mice.
Findings
Furanylfentanyl acts as a partial agonist at MOR with similar affinity and potency as fentanyl.
Fentanyl caused more severe and prolonged oxygen saturation depletion compared to furanylfentanyl.
Fentanyl induced more apneas, explaining the greater drop in blood oxygen.
Abstract
Fatal opioid overdoses involving synthetic opioids have increased >10-fold over the past 8 years. Fentanyl, a potent synthetic opioid acting at the mu-opioid receptor (MOR), has largely replaced heroin as the predominant illicit opioid available. Various analogs have been synthesized from the fentanyl scaffold, including 2-furanylfentanyl, one of the most trafficked analogs from 2019 to 2024. Despite its prevalence, the in vivo effects of furanylfentanyl are not well characterized, especially regarding respiratory depression. To characterize furanylfentanyl relative to fentanyl in vitro and compare the effects of both drugs in mouse models of antinociception and respiratory depression. The affinity and potency of fentanyl and furanylfentanyl were determined in CHO cells overexpressing MOR. Male and female mice were administered (i.p.) fentanyl or furanylfentanyl prior to measuring…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
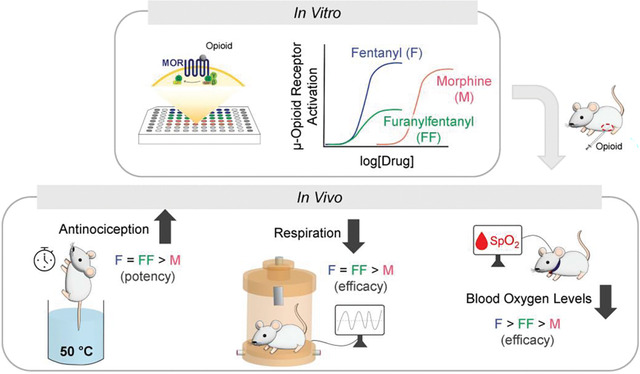 Figure 1
Figure 1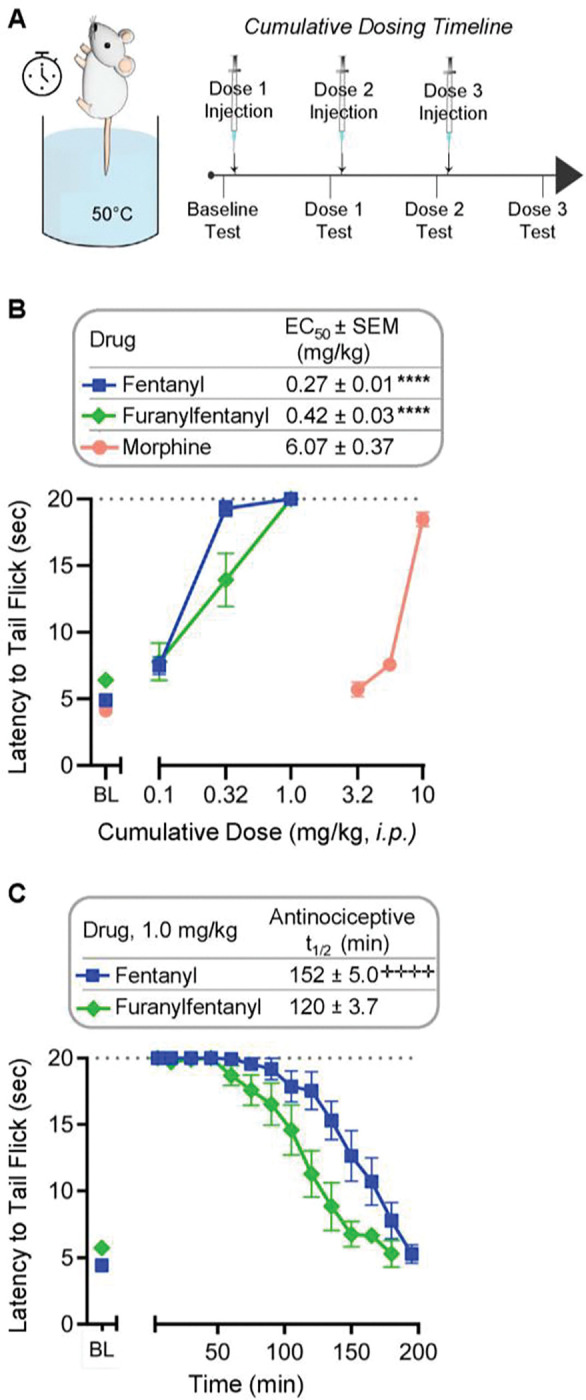 Figure 2
Figure 2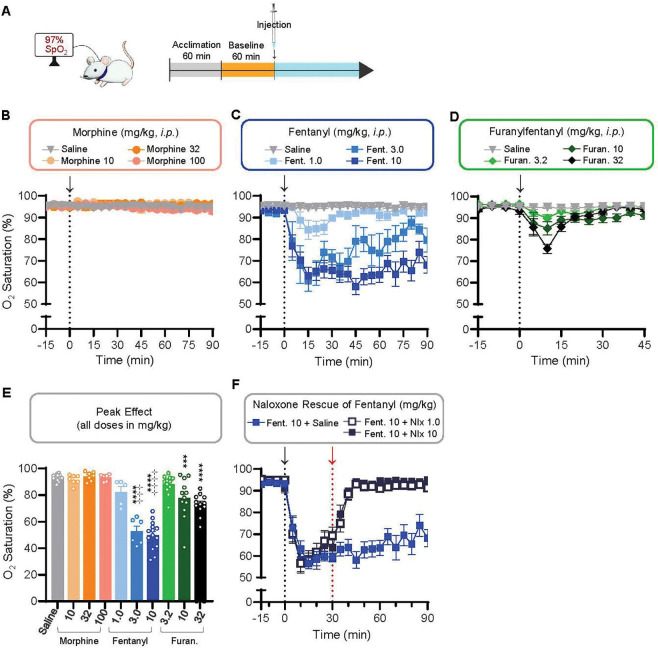 Figure 3
Figure 3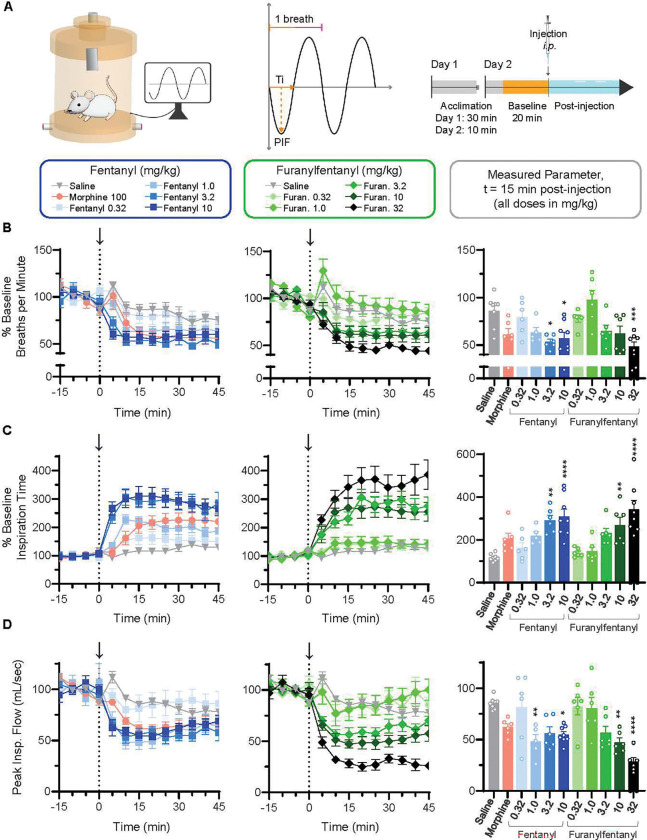 Figure 4
Figure 4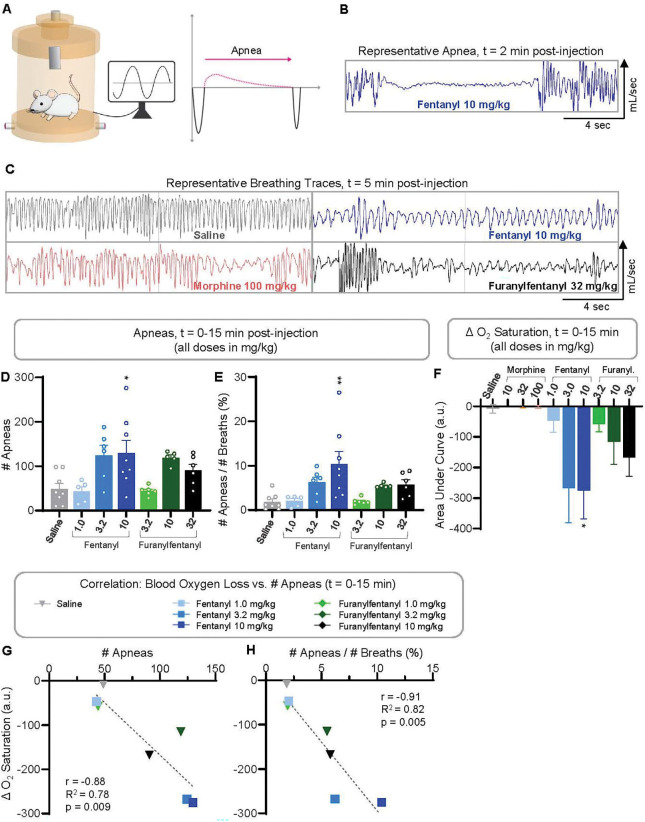 Figure 5
Figure 5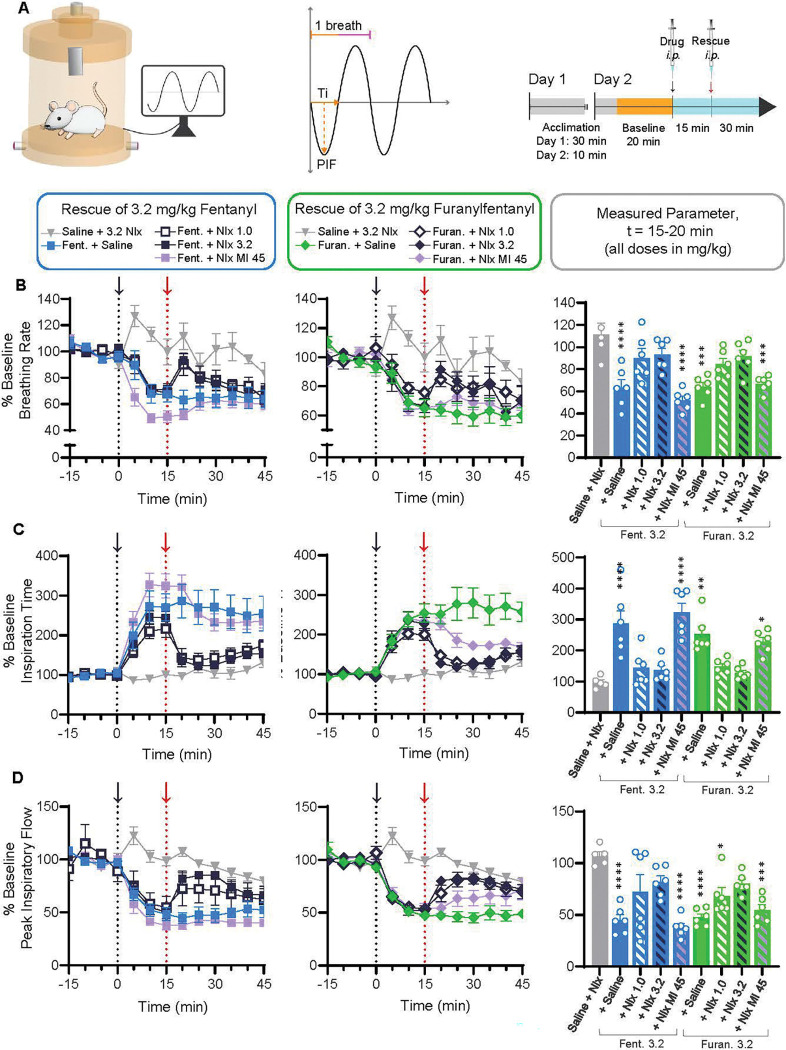 Figure 6
Figure 6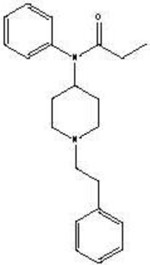 Figure 7
Figure 7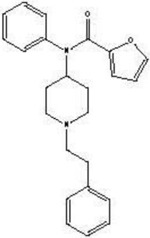 Figure 8
Figure 8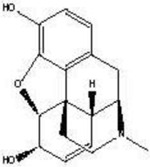 Figure 9
Figure 9- —National Institutes of Health
- —National Institute of General Medical Sciences
- —National Institute on Drug Abuse/NIDA
- —National Heart, Lung, and Blood Institute
- —National Center for Advancing Translational Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroscience of respiration and sleep · Pharmacological Receptor Mechanisms and Effects · Neurotransmitter Receptor Influence on Behavior
INTRODUCTION
Over 100,000 people in the United States died of a drug overdose in 2023 (Ahmad, Cisewski et al. 2025). Deaths associated with synthetic opioids acting at the mu-opioid receptor (MOR), namely fentanyl and its analogs, have increased more than 10-fold over the past decade (Ahmad, Cisewski et al. 2025). Opioids are used clinically to relieve pain, an effect mediated through MOR; yet, activation of MOR also reduces breathing rate and regularity, causing CO_2_ retention and insufficient blood O_2_ levels that can trigger cardiorespiratory arrest (Bateman, Saunders et al. 2023). Fentanyl induces respiratory depression more potently than morphine or heroin (Hill, Santhakumar et al. 2020, Marchette, Carlson et al. 2023). This is hypothesized to result from its greater potency at MOR (Traynor and Nahorski 1995) and greater induction of respiratory muscle rigidity (Cavallo, Kelly et al. 2023). Furthermore, pharmacokinetic attributes of fentanyl enable rapid permeation of the blood-brain barrier (Kiyatkin 2019, Hill, Santhakumar et al. 2020) yielding a rapid onset of action that both hinders the body’s chance to respond to rising CO_2_ levels and decreases the window of opportunity for reversal by administration of naloxone (Narcan^®^).
In addition to being more potent than heroin, fentanyl is easier to synthesize on a large scale (Cicero, Ellis et al. 2015, Anderson 2017, Mars, Rosenblum et al. 2019). As a result, fentanyl has largely replaced heroin as the most common illicit opioid available (Tupper, McCrae et al. 2018, U.S. Drug Enforcement Administration 2025). Besides fentanyl, various fentanyl analogs have been identified in illicit drug seizures and overdose victims. 2-furanylfentanyl (furanylfentanyl, Figure 1) is consistently detected in the U.S. illicit drug market (United States Sentencing Commission (USSC) 2021, Tennyson, Ray et al. 2021, USSC 2022, USSC 2023, USSC 2024, USSC 2025) and has been implicated in fatal overdoses worldwide (Helander, Backberg et al. 2016, Guerrieri, Rapp et al. 2017, Daniulaityte, Juhascik et al. 2019, O’Donnell, Gladden et al. 2020). Despite the prevalence of furanylfentanyl, its effects are not well characterized (Nektaria Misailidi, Ioannis Papoutsis et al. 2017). Furanylfentanyl has a similar binding affinity to MOR as fentanyl but only shows partial agonism in vitro (Eshleman, Nagarajan et al. 2020, Bilel, Azevedo Neto et al. 2022, Anand, Moore et al. 2024). Yet, recent research has reported comparable antinociception (Bilel, Azevedo Neto et al. 2022) and abuse liability (Chen, Lai et al. 2025, Obeng, Urquhart et al. 2025) between furanylfentanyl and fentanyl in rodent models. Whether furanylfentanyl induces respiratory depression akin to fentanyl remains unclear.
Herein, we compared the in vitro binding affinity and efficacy of furanylfentanyl, fentanyl, and morphine at MOR, as well as antinociception and respiratory depression. We confirmed that furanylfentanyl behaves as a partial agonist relative to fentanyl and morphine but with similar affinity and potency as fentanyl. In mice, furanylfentanyl significantly reduced blood oxygen levels as determined by pulse oximetry, although transiently and to a lesser extent than a 10-fold lower dose of fentanyl. Using whole-body plethysmography, furanylfentanyl produced similar changes in breathing parameters (rate, inspiration time and peak inspiratory flow) as fentanyl with approximately three-fold lower potency. Thus, doses of furanylfentanyl and fentanyl that induced comparable effects on breathing parameters produced markedly different effects on oxygen saturation. This discrepancy may be explained by a higher frequency of apneas induced by fentanyl (Demery, Moore et al. 2025) compared to its furanyl analog. This suggests another mechanism by which fentanyl produces more lethal effects in humans compared to other opioids.
METHODS
Cell Lines
Chinese hamster ovary (CHO) cells stably expressing the human MOR (MOR) were kindly provided by Dr. Lawrence Toll (Spagnolo, Calo et al. 2008) and used for competitive binding and [^35^S]GTPγS binding assays. Cells were incubated at 37°C with 5% CO_2_ in 1:1 Gibco Dulbecco’s Modified Eagle’s Medium:F12 (Thermo Fisher Scientific). All media contained 10% fetal bovine serum (Millipore Sigma) and 5% penicillin/streptomycin (Thermo Fisher Scientific). Cells were grown to confluence before harvesting. PathHunter^®^ CHO cells expressing human MOR (DiscoveRx, 93–0213C2) were used to measure inhibition of adenyl cyclase and β arrestin-2 recruitment.
Preparation of Membrane Fractions
Cells were incubated in warm harvesting buffer (150 mM NaCl, 20 mM HEPES, 0.50 mM EDTA, pH 7.4) for 15 min, then detached using a cell scraper and centrifuged for 3 min at 200 × g. The cell pellet was resuspended in ice-cold 50 mM Tris-HCl buffer (pH 7.4) and homogenized using a BioSpec Products Tissue-Tearor^™^ (Fisher Scientific) for 20 sec on setting 4. The homogenate was then centrifuged for 20 min at 20,000 × g and 4°C. The previous two steps were repeated except using the Tearor for 10 sec on setting 2. The final pellet was resuspended in ice-cold 50 mM Tris-HCl, further homogenized in a glass douncer, then aliquoted and stored at −80°C. A Pierce^™^ BCA Protein Assay Kit (Thermo Fisher Scientific) including bovine serum albumin as the standard was used to determine protein concentration.
Competition Binding
Competitive displacement of [^3^H]diprenorphine (Revvity; 250 μCi, 9.25 MBq) a non-selective opioid ligand, was used to measure drug affinity at MOR. Assays were carried out in 96-well plates with 10–20 μg membrane fraction protein, 0.20 nM [^3^H]diprenorphine, 50 mM Tris-HCl buffer, and various concentrations of test ligand per well. The plate was incubated for 1 hr (to allow binding to reach equilibrium) at room temperature (RT) on a shaker. Next, samples were filtered through Whatman^®^ Grade CF/C glass filters (Millipore Sigma) and washed several times with 50 mM Tris-HCl buffer. Filters were saturated with EcoLume^™^ Liquid Scintillation Cocktail (MP Biomedical) and then radioactivity was quantified using a 2450 MicroBeta^2®^ Microplate Counter (Revvity). Buffer was used to determine total binding and 10 μM naloxone (Fisher Scientific) was used to determine non-specific binding. Results presented are the average ± S.E.M. of three independent assays performed on different days; within each assay, duplicates were performed and averaged. Data were fit to a one-site competition binding curve (non-linear regression) using GraphPad Prism 10.0.0. The Cheng-Prusoff equation was used to convert the resulting IC_50_ values to K_i_ values (Cheng and Prusoff 1973).
[35S]GTPγS Binding
Agonist ability to stimulate [^35^S]guanosine-5’-O-(γ-thio)triphosphate ([^35^S]GTPγS; Revvity; 1250 Ci/mmol) binding at MOR was determined. Assays were carried out in 96-well plates with 10–20 μg membrane fraction protein, 0.10 nM [^35^S]GTPγS, assay buffer (50 mM Tris-HCl, 100 mM NaCl, 10 mM MgCl_2_, 1 mM EDTA, pH 7.4), 30 μM guanosine 5’-diphosphate (Millipore Sigma), and various concentrations of test ligand per well. The plate was incubated for 1 hr at RT on a shaker. Next, samples were filtered through Whatman^®^ Grade CF/C glass filters (Millipore Sigma) and washed several times with assay buffer. Radioactivity was quantified as described earlier. Assay buffer was used to determine baseline stimulation and test agonists were compared to 10 μM D-Ala^2^, N-Me-Phe^4^, Gly^5^-ol)-enkephalin (DAMGO; Millipore Sigma). Results presented are the result of 3–5 independent assays performed on different days; within each assay, duplicates were performed and averaged. Data were fit to a sigmoidal dose-response curve for agonist stimulation (non-linear regression) using GraphPad Prism 10.0.0.
β arrestin-2 Recruitment
β arrestin-2 recruitment to human MOR expressed in PathHunter^®^ CHO cells (DiscoverX 93–0213C2) was determined using the Beta-Glo^®^ β-galactosidase Enzyme-Complementation Assay (Promega). Cells were prepared per manufacturer instructions and then incubated (37°C with 5% CO_2_) with various concentrations of orthosteric agonist for 60 min. Cells were allowed to cool to RT (15 min) then administered chemiluminescent Beta-Glo^®^ Detection Reagent and incubated for 60 min in the dark at RT. β-galactosidase enzyme activity, indicating β arrestin-2 recruitment, was detected as luminescence measured via a GloMax^®^ Explorer Microplate Reader (Promega). Agonist activity was compared to 10 μM DAMGO.
Inhibition of Adenylyl Cyclase
Varying concentrations of orthosteric agonist were used to inhibit cAMP production by adenyl cyclase as described by the cAMP-Glo^™^ Assay Protocol (Promega). All incubations were done at RT. Briefly, PathHunter^®^ CHO cells stably expressing MOR (DiscoverX 93–0213C2) were plated in 96-well plates and incubated in induction buffer for 15 min. To evaluate agonist-induced inhibition of adenyl cyclase, 10 μM forskolin was added and incubated for 15 min. Lysis buffer was then added and the plate was incubated for 30 min on a shaker. Next, cAMP Detection Solution was added and the plate was incubated for 20 min, followed by addition of Kinase-Glo^®^ Reagent and a final 10-min incubation in the dark. Activity was detected by luminescence measured with a GloMax^®^ Explorer Microplate Reader (Promega). Maximum possible effect was considered 10 μM DAMGO with minimum effect as forskolin only (no agonist).
Drugs
Fentanyl, furanylfentanyl, and morphine powders were purchased from Cayman Chemical Company and dissolved in saline. Each animal received one intraperitoneal (i.p.) injection at 10 mL/kg of body weight.
Animal Husbandry and Care
Adult male and female CD-1 mice (5–12 weeks of age) were used for all experiments unless otherwise specified. Mice were bred in-house at the University of Michigan or purchased from Envigo (Indianapolis, IN). Mice were maintained on a 12-hour light/dark cycle, with experiments occurring during the light phase. Mice were housed in clear polypropylene cages with corncob bedding, in groups of 2–5 by sex, with access to food, water, and enrichment ad libitum. All experiments were conducted in accordance with the Guide for the Care and Use of Laboratory Animals (Committee 2011) and approved by the University of Michigan Institutional Animal Care and Use Committee.
Warm-Water Tail Withdrawal
The antinociceptive effect of various agonists was determined using the warm water tail withdrawal (WWTW) assay. Experimenter was blinded to the drug treatments given. CD-1 mice were restrained in a plastic cylinder with tails submerged 2–3 cm into a 50°C water bath. Latency to tail flick was recorded with a 20 sec cutoff to prevent tissue damage. Dose-response curves were created using a cumulative dosing procedure. Briefly, mice received an i.p. injection of saline, and baseline latency was recorded 15 min later. Next, mice received an i.p. injection of 0.1 mg/kg furanylfentanyl or fentanyl, and latency time was re-measured after 15 min. The previous step was repeated for increasing doses of furanylfentanyl or fentanyl up to a 1.0 mg/kg cumulative dose. For morphine, which has a slower onset and longer duration of action, 30-min dosing intervals were employed. Fentanyl and furanylfentanyl groups, n=12 mice each; morphine and saline groups, n=6 mice each. Excel and GraphPad Prism 10.0.0 were used for all analyses.
MouseOx
The MouseOx^®^ Plus Small Animal Pulse Oximeter (Starr Life Sciences, Oakmont, PA) was used to measure pulse oximetry parameters in awake, freely moving CD-1 mice. (The fur of CD-1 mice is white, so they do not need to be shaved for accurate infrared sensing.) Mice were habituated to enclosures with a practice collar for 1 hr. The practice collar was replaced with the MouseOx^®^ collar and then baseline pulse oximetry measurements, including percent oxygen saturation (SpO_2_), were recorded for 1 hr using MouseOx^®^ software version 1.6.8. At t=0, mice were injected (i.p.) with opioid. Data was recorded for 100 min post-injection and averaged into 5 min bins.
Whole-Body Plethysmography
Respiratory parameters of CD-1 mice were recorded in whole-body plethysmography (WBP) chambers (SCIREQ vivoFlow^®^, Montreal, QC Canada) supplied with 0.5 L/min of medical air USP (Cryogenic Gases). Mice were acclimated to WBP chambers for 30 minutes on day one. On day two, mouse weights and temperatures (Braun digital infrared thermometer) were recorded, and body temperature (35–37°C), chamber temperature (20–22°C) and chamber humidity (10%) were used in volume calculations. Mice were placed in chambers for 30 minutes prior to injection: 10 min of habituation (data not saved) and 20 min of baseline recording. Mice were removed for intraperitoneal injection of opioid or saline and then returned to chambers for 60 min post-injection. For naloxone reversal studies, mice were removed 15 min post-injection of opioid for i.p. naloxone administration, then returned to chambers for 45 min post-naloxone. Data was recorded in Emka Technologies iox 2.10.8.6 software and analyzed in 5-min bins for all data except apneas and EF50 values, which were assessed on a breath-by-breath basis.
Hot Plate
Latency to nociceptive behaviors (i.e., forepaw licking or jumping) in mice on a 55°C hot plate was measured at baseline, 15 min after i.p. injection of 3.2 mg/kg furanylfentanyl, and subsequently every 5 min after i.p. injection of a rescue agent (3.2 mg/kg naloxone, 45 mg/kg naloxone methiodide, or saline as control). Experimenter was blinded to the drug rescue given. A cut-off time of 1 min was implemented to prevent tissue damage.
Data Analysis
For all comparisons, p<0.05 was considered significant. Post-hoc analyses were only considered for significant ANOVAs or mixed effects analyses and were performed to evaluate all possible multiple comparisons. Measures of affinity (Ki), potency (EC_50_), maximum effect, and area under the curve were compared using a one-way ANOVA. Antinociceptive t_1/2_ values were compared via unpaired t-test. For all plethysmography parameters (ex., breathing rate, inspiration time, tidal volume, etc.), data were averaged into 5-min bins for analysis, and groups were compared using a two-way ANOVA or mixed effects model (with treatment and time as independent variables). If significant, then a one-way ANOVA with Tukey’s post-hoc test was used to compare group averages from 10–15 min post-injection; this same method was applied for blood oxygen saturation. For apnea analysis, data were exported as individual breaths. Excel was used to calculate the number of apneas per group from 0–15 min post-injection using the following criteria: , with the average length of Te from t=0–60 min used to define an apnea for each mouse individually. The number of apneas per treatment group from t=0–15 min post-injection were compared via one-way ANOVA with Tukey’s post-hoc test. GraphPad Prism 10.0.0 was used for all statistical analyses except apneic frequency and linear trendlines and S.E.M. for correlation analyses, which were calculated in Excel.
RESULTS
Furanylfentanyl is a potent partial agonist at the mu-opioid receptor
We first examined the properties of fentanyl and furanylfentanyl at MOR in vitro compared to morphine (Table 1). Fentanyl, furanylfentanyl, and morphine all bound with comparable single digit nanomolar affinity to MOR (p=0.102). Fentanyl (p=0.013) and furanylfentanyl (p=0.011) were significantly more potent than morphine in stimulating [^35^S]GTPγS binding; furanylfentanyl shower lower efficacy (20% maximum activation of the standard MOR full agonist, DAMGO) compared to fentanyl and morphine (p<0.0001 for both). In evaluating agonist ability to inhibit adenylyl cyclase, we found that both fentanyl (p<0.0001) and furanylfentanyl (p<0.0001) were significantly more potent than morphine. All three agonists reached maximum effect in this assay, reflecting signal amplification occurring downstream of receptor activation.
Agonist recruitment of β arrestin-2, a multifunctional protein that typically drives receptor internalization but can act as a scaffold for other signaling molecules (Seyedabadi, Gharghabi et al. 2021), was also determined. We observed that fentanyl (p=0.003) and furanylfentanyl (p=0.002) were also more potent than morphine in this assay. Maximum β arrestin-2 recruitment (relative to DAMGO) differed by agonist: fentanyl (65%; p<0.0001 vs. furanylfentanyl, morphine) > morphine (35%) > furanylfentanyl (17%; p=0.0006 vs. morphine).
Furanylfentanyl induces comparable thermal antinociception to fentanyl
We next evaluated the ability of furanylfentanyl to induce thermal antinociception in mice, as measured by the warm-water tail withdrawal assay. A cumulative dosing procedure was employed (Figure 1A), with 15-min dosing intervals for fentanyl and furanylfentanyl, and 30-min dosing intervals for morphine due to its slower onset (Hill, Santhakumar et al. 2020). As shown in Figure 1B, fentanyl (p<0.0001) and furanylfentanyl (p<0.0001) were significantly more potent in affording antinociception than morphine. There was no difference in antinociceptive potency (EC_50_) between fentanyl and furanylfentanyl (p=0.628). However, the effect of fentanyl persisted for a significantly longer time (Figure 1C; p<0.0001).
Fentanyl decreases blood oxygen saturation in mice to a greater extent than furanylfentanyl
We next considered whether furanylfentanyl would decrease oxygen saturation in vivo. Blood oxygen saturation in the carotid artery was monitored in mice using pulse oximetry (Figure 2A). At all doses tested (10, 32, and 100 mg/kg), morphine did not reduce oxygen saturation compared to saline (Figure 2B). In contrast, doses of 3.0 and 10 mg/kg fentanyl markedly reduced oxygen saturation for an extended period (Figure 2C). Furanylfentanyl transiently decreased blood oxygen saturation (Figure 2D). Comparing the peak effect, doses of 10 (p=0.0010) and 32 mg/kg furanylfentanyl (p<0.0001) significantly reduced blood oxygen saturation relative to saline (Figure 2E). Fentanyl depleted blood oxygen saturation more potently and to a greater extent than furanylfentanyl: doses of 3.0 and 10 mg/kg produced significantly lower blood oxygen levels than 32 mg/kg furanylfentanyl (p=0.0005 vs. 3.0, p<0.0001 vs. 10 mg/kg fentanyl). The effect of 10 mg/kg fentanyl on blood oxygen saturation was fully reversed by the antagonist naloxone, confirming an opioid receptor-mediated action (Figure 2F).
Fentanyl and furanylfentanyl produce similar changes in breathing parameters in mice
We next asked whether the more severe and prolonged decrease in oxygen saturation caused by fentanyl compared to furanylfentanyl could be explained by differences in breathing parameters, as determined using whole-body plethysmography in awake, freely moving animals (Figure 3A). The effects of fentanyl (0.32, 1.0, 3.2, and 10 mg/kg) and furanylfentanyl (0.32, 1.0, 3.2, 10, and 32 mg/kg) administration were examined. As morphine did not reduce oxygen saturation at any dose tested, only the highest dose (100 mg/kg) was included in the plethysmography experiments. The average breathing rate by group at t=15 min post-injection (compared via 1-way ANOVA with post-hoc analyses) showed that doses of 3.2 (p=0.015) and 10 (p=0.024) mg/kg fentanyl, as well as 32 mg/kg furanylfentanyl (p=0.0008), decreased breathing rate relative to saline (Figure 3B, bar graph).
As depicted in Figure 3A, a given breath is comprised of inspiration time (Ti) and expiration time (Te); thus, if breathing rate slows, either Ti and/or Te should lengthen. Ti is the most sensitive marker of opioid-induced respiratory depression (Demery, Moore et al. 2025), and we observed a clear dose-dependent increase in Ti for both fentanyl and furanylfentanyl (Figure 3C). Doses of 3.2 (p=0.001) and 10 (p<0.0001) mg/kg fentanyl, along with 10 (p=0.007), and 32 (p<0.0001) mg/kg furanylfentanyl, significantly increased Ti relative to saline at t=15 min post-injection (Figure 3C, bar graph). The volume of air inhaled (tidal volume) depends on Ti and peak inspiratory flow (PIF), which describes the fastest flow of air through the lungs in each breath. Doses of 1 (p=0.01) and 10 (p=0.021) mg/kg fentanyl, as well as 10 (p=0.004), and 32 (p<0.0001) mg/kg furanylfentanyl decreased peak inspiratory flow relative to saline at t=15 min post-injection (Figure 3D, bar graph). Only 10 mg/kg fentanyl increased tidal volume compared to control (p=0.001), while both 3.2 (p=0.009) and 10 mg/kg fentanyl (p<0.0001) increased tidal volume relative to 32 mg/kg furanylfentanyl (Supplementary Figure 2B). In addition, the effects of 100mg/kg morphine on various breathing parameters, generally similar to those produced by 1.0mg/kg fentanyl, were not different from saline at t=15 min post-injection.
Opioid effects on expiration were minimal (Supplementary Figure 1). Expiration time and end expiratory pause (EEP; a delay between the end of one breath and the beginning of the next) were transiently increased by 10 mg/kg fentanyl at 5 min post-injection (p<0.0001 for both metrics), and 2) EEP was increased by 32 mg/kg furanylfentanyl from t=5–15 min post-injection (p≤0.001 for all timepoints; Supplementary Figure 1B, 1C). Last, 3.2 (p=0.007) and 10 mg/kg fentanyl (p=0.008) increased peak expiratory flow relative to 32 mg/kg furanylfentanyl (Supplementary Figure 1D).
Fentanyl induces apneas that correlate with blood oxygen loss
Changes in breathing parameters did not explain the more drastic and prolonged decrease in saturated oxygen induced by 3.2 and 10 mg/kg fentanyl compared to 32 mg/kg furanylfentanyl (Figure 2C, 2D, 2E). Thus, we questioned whether there might be a difference in the number of apneas caused by the two drugs, as apneas have been shown to decrease oxygen saturation in humans (Yadollahi, Giannouli et al. 2010, Wali, Abaalkhail et al. 2020) and we have shown apneas correlate with reductions in blood oxygen levels (Demery, Moore et al. 2025). A representative apnea following 10 mg/kg fentanyl is displayed in Figure 4B. Shown in Figure 4C are representative breathing traces from saline-, 10 mg/kg fentanyl-, 32 mg/kg furanylfentanyl-, and 100 mg/kg morphine-treated mice, As shown in Figure 4D, significantly more apneas were observed in mice administered 10 mg/kg fentanyl (p=0.017) with a similar trend in the 3.2 mg/kg fentanyl group (p=0.057), compared to saline during the first 15 min following opioid administration. Apneic frequency, % apneas out of total breaths, was also significantly increased by 10 mg/kg fentanyl (p=0.0015) during the first 15 min post-injection (Figure 4E). To investigate an association between oxygen depletion (Figure 4F) and frequency of apneas, the Pearson correlation between AUC for blood oxygen saturation loss (t=0–15 min) and the number of apneas (t=0–15 min) was determined (p=0.009; Figure 4G). Likewise, there was a significant correlation between blood oxygen loss and apneic frequency (p=0.005; Figure 4H). These results indicate that there is a correlation between frequency of apneas and blood oxygen depletion following administration of opioids, as we have shown previously (Demery, Moore et al. 2025).
Effects of fentanyl and furanylfentanyl on breathing were readily reversed by naloxone but not naloxone methiodide
The current standard of care for opioid overdose is naloxone, or Narcan^®^. Therefore, we evaluated the ability of two doses of naloxone (1.0 and 3.2 mg/kg, chosen based on previous work (Anand, Moore et al. 2024) to reverse the effects of fentanyl and furanylfentanyl (3.2 mg/kg each) on breathing rate, inspiration time, and peak inspiratory flow. Rescue injections were administered 15 min after opioid (Figure 5A). Either dose of naloxone readily reversed the effects of 3.2 mg/kg fentanyl and furanylfentanyl on breathing rate, inspiration time, and peak inspiratory flow within 5 min of rescue injection (Figure 5B, 5C, 5D), with the exception that 1.0 mg/kg naloxone did not rescue the furanylfentanyl-induced decrease in PIF.
To determine if any of the actions on breathing have a peripheral component, as recently reported (Ruyle, Masud et al. 2025), we examined furanylfentanyl reversal using 45 mg/kg naloxone methiodide, considered to be a peripherally restricted antagonist. Based on previously published literature showing that doses of 30–100 mg/kg naloxone methiodide could reverse the respiratory effects of opioid agonists (Lewanowitsch, Miller et al. 2006), as well as our in vitro data showing that naloxone methiodide has a 15-fold lower affinity for MOR than naloxone (Ki values of 15.24 and 0.72 nM, respectively), the dose of 45 mg/kg was chosen. Naloxone methiodide did not significantly rescue the effects of either opioid on any breathing parameter within the first 5 min post-rescue injection (Figure 5B, 5C, 5D). However, by 10 min post-administration, naloxone methiodide reversed the effects of furanylfentanyl on inspiration time and peak inspiratory flow (bar graphs not shown). To confirm that naloxone methiodide was only acting peripherally, we performed the hot plate test and found that naloxone methiodide at 45 mg/kg reversed furanylfentanyl-induced antinociception (Supplementary Figure 3). Since nociceptive responses in this test involve higher brain centers, this indicates that the dose of naloxone methiodide used likely permeated the central nervous system.
DISCUSSION
Herein, we have shown that furanylfentanyl and fentanyl produce similar effects on breathing rate, inspiration time, and peak inspiratory flow in mice, with furanylfentanyl exhibiting approximately three-fold lower potency than fentanyl. On the other hand, fentanyl markedly decreases blood oxygen saturation relative to the small, transient effect of furanylfentanyl. The greater reduction in blood oxygenation following fentanyl administration may result from the higher frequency of apneas produced by fentanyl.
Both fentanyl and furanylfentanyl were more potent than morphine in G-protein activation and β arrestin-2 recruitment at the mu-opioid receptor. Furanylfentanyl behaved as a lower efficacy agonist relative to fentanyl in both assays, consistent with other literature (Eshleman, Nagarajan et al. 2020, Bilel, Azevedo Neto et al. 2022, Anand, Moore et al. 2024). Yet, the low partial agonist activity of furanylfentanyl in vitro was sufficient to produce substantial effects in vivo. In mice, furanylfentanyl induced thermal antinociception with comparable potency to fentanyl, although the antinociceptive duration of activity was shorter. Fentanyl and furanylfentanyl have previously been shown to produce equipotent mechanical antinociception (Bilel, Azevedo Neto et al. 2022), and comparable abuse liability (Chen, Lai et al. 2025, Obeng, Urquhart et al. 2025),which explains the recreational use of furanylfentanyl in humans. On the other hand, furanylfentanyl administration induces less severe precipitated withdrawal symptoms than fentanyl in mice (Gannon, Shepard et al. 2025, Obeng, Urquhart et al. 2025), revealing the nuanced characteristics of this potent partial agonist in vivo.
At higher doses than those required to achieve antinociception, furanylfentanyl transiently decreased blood oxygen saturation, though the effect was of much lower magnitude than fentanyl, even at a 10x greater dose. This agrees with a previous study reporting that furanylfentanyl did not reduce oxygen saturation at doses up to 6 mg/kg (Bilel, Azevedo Neto et al. 2022), measured as average drug effect over a 2-hour period. Given the reports of human fatalities involving furanylfentanyl (Helander, Backberg et al. 2016, Guerrieri, Rapp et al. 2017, Daniulaityte, Juhascik et al. 2019, O’Donnell, Gladden et al. 2020), the transient decrease in blood oxygen levels we observed in mice may translate to a more severe effect in humans, as mice can better adapt to changes in blood oxygen levels due to their higher specific metabolic rate (Demetrius 2005, Perlman 2016).
Surprisingly, the difference in blood oxygen levels following fentanyl and furanylfentanyl administration was not explained by differences in the suppression of breathing rate or inspiration. As the two fentanyls produced similar effects on breathing metrics derived from plethysmography, albeit fentanyl 3–10-times more potently, these observations do not explain the greater effect on blood oxygen levels induced by fentanyl. Rather, we found that the number of apneas was associated with blood O_2_ levels, consistent with our previous work (Demery, Moore et al. 2025) and in line with findings that apneas reduce oxygen saturation in humans (Yadollahi, Giannouli et al. 2010, Wali, Abaalkhail et al. 2020). Morphine was even less effective at reducing blood oxygen than furanylfentanyl despite its higher in vitro efficacy. This is likely a reflection of morphine’s slower onset, which allows central chemoreceptors to compensate for rising CO_2_ levels in the blood to better stabilize blood gases (Bateman, Saunders et al. 2023) – a reflex that would be hindered by the more rapidly acting fentanyl and furanylfentanyl. Together, these results highlight the complexity of the mechanisms underlying fentanyl’s lethality, which is not clearly delineated based on plethysmography parameters alone.
We found that naloxone at 3.2 mg/kg was readily able to rescue changes in breathing parameters caused by 3.2 mg/kg fentanyl or furanylfentanyl. This contrasts with a previous report in mice under hypercapnic conditions, which found that 3.0 mg/kg naloxone was necessary to rescue the effects of 0.15 mg/kg fentanyl on breathing (Hill, Santhakumar et al. 2020), suggesting a much higher naloxone-to-fentanyl dosage requirement. However, the effects of opioids are exacerbated under the somewhat artificial conditions of hypercapnia, potentially explaining the discrepancy between findings. In light of recent research demonstrating a peripheral opioid receptor contribution to opioid-induced respiratory depression (Ruyle, Masud et al. 2025), we investigated the rescue potential of naloxone methiodide, a peripherally restricted opioid receptor antagonist, and found it was able to attenuate respiratory depression induced by furanylfentanyl with a slower onset of effect than naloxone. However, naloxone methiodide, at the same dose and on the same time course, also blocked supraspinal antinociception, leading us to conclude that the observed reversal of furanylfentanyl-induced changes in breathing was due to naloxone methiodide acting centrally.
In conclusion, we have shown that furanylfentanyl reduces oxygen saturation much less than fentanyl at doses that produce comparable reductions in breathing rate and inspiration. Since both drugs have similar effects on breathing, albeit that fentanyl is 3x more potent, the differential effects could be explained by the greater number of apneas induced by fentanyl in the first 15 minutes post-injection, during which time oxygen levels in furanylfentanyl- but not fentanyl-treated mice recover. It is possible that inducing apneas requires higher opioid efficacy than depressing breathing rate or inspiration, reflecting the lower efficacy of furanylfentanyl relative to fentanyl observed in vitro. The findings underscore the complexities of fentanyl-induced respiratory depression while also contributing to our understanding of opioid-induced respiratory phenotypes. Overall, our results indicate that the recreational use of 2-furanylfentanyl in humans poses a significant overdose risk that is greater than that of morphine but lesser than that of fentanyl.
Supplementary Material
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmad F. B., Cisewski J. A., Rossen L. M. and Sutton P. (2025). Provisional drug overdose death counts. National Center for Health Statistics.
- 2Anand J. P., Moore S. C., Dixon E. E., Perrien Naccarato C. M., West J. L., Delong L. J., Burgess E., Twarozynski J. J. and Traynor J. R. (2024). “Structure-Activity Relationships of the Fentanyl Scaffold: Identification of Antagonists as Potential Opioid Overdose Reversal Agents.” ACS Chem Neurosci 15(15): 2830–2841.38994846 10.1021/acschemneuro.4c 00203 PMC 11459602 · doi ↗ · pubmed ↗
- 3Anderson T. (2017). “Curbing prescription opioid dependency.” Bull World Health Organ 95(5): 318–319.28479631 10.2471/BLT.17.020517 PMC 5418819 · doi ↗ · pubmed ↗
- 4Bateman J. T., Saunders S. E. and Levitt E. S. (2023). “Understanding and countering opioid-induced respiratory depression.” Br J Pharmacol 180(7): 813–828.34089181 10.1111/bph.15580 PMC 8997313 · doi ↗ · pubmed ↗
- 5Bilel S., Azevedo Neto J., Arfe R., Tirri M., Gaudio R. M., Fantinati A., Bernardi T., Boccuto F., Marchetti B., Corli G., Serpelloni G., De-Giorgio F., Malfacini D., Trapella C., Calo G. and Marti M. (2022). “In vitro and in vivo pharmaco-dynamic study of the novel fentanyl derivatives: Acrylfentanyl, Ocfentanyl and Furanylfentanyl.” Neuropharmacology 209: 109020.35247453 10.1016/j.neuropharm.2022.109020 · doi ↗ · pubmed ↗
- 6Cavallo D., Kelly E., Henderson G. and Abdala Sheikh A. P. (2023). “Comparison of the effects of fentanyls and other mu opioid receptor agonists on the electrical activity of respiratory muscles in the rat.” Front Pharmacol 14: 1277248.38074147 10.3389/fphar.2023.1277248 PMC 10710149 · doi ↗ · pubmed ↗
- 7Chen Y., Lai M., Li X., Qiao Y., Xu D., Fu D., Di B. and Xu P. (2025). “Assessment of abuse potential of furanylfentanyl and tetrahydrofuranylfentanyl.” Psychopharmacology (Berl) 242(9): 2123–2133.40244334 10.1007/s 00213-025-06790-2 · doi ↗ · pubmed ↗
- 8Cheng Y. and Prusoff W. H. (1973). “Relationship between the inhibition constant (K 1) and the concentration of inhibitor which causes 50 per cent inhibition (I 50) of an enzymatic reaction.” Biochem Pharmacol 22(23): 3099–3108.4202581 10.1016/0006-2952(73)90196-2 · doi ↗ · pubmed ↗