SOD1 at the Crossroads: Co-Overexpression of Canonical Antioxidant Response and Noncanonical Hydrogen Sulfide Generation Pathways in Down Syndrome, With Immune Cell Implications
Karthik Mouli, Anton V. Liopo, Hongbin Wang, Larry J. Suva, Kenneth R. Olson, Paul J. Derry, Thomas A. Kent

TL;DR
This study explores how Down Syndrome affects immune cells through SOD1 overexpression and links it to oxidative stress and hydrogen sulfide production.
Contribution
The study reveals a novel connection between SOD1 overexpression and noncanonical hydrogen sulfide generation in Down Syndrome immune dysfunction.
Findings
SOD1 mRNA overexpression is strongly linked to inflammatory conditions in Down Syndrome.
SOD1 co-overexpression correlates with altered immune cell profiles and elevated inflammatory markers.
Hydrogen sulfide overproduction in Down Syndrome may occur via thioredoxin-dependent pathways, not CBS.
Abstract
Accelerated immune cell aging is well-recognized feature of Down Syndrome (DS), a condition caused by trisomy of human chromosome 21 (Hsa21). DS predisposes individuals to recurrent infections, autoimmunity, low bone mass and leukemia. To investigate potential connections between immune cell dysfunction or disruption in DS, serum transcriptomic and proteomic datasets from DS and euploid individuals were examined. High DS superoxide dismutase 1 (SOD1) mRNA expression was consistently found and was strongly associated with an increased odds of inflammatory co-occurring conditions such as pharyngitis. SOD1 mRNA overexpression was also associated with decreased M2-polarized macrophages, increased resting-memory CD4+ T cells, elevated serum interleukin-16 levels and interferon-γ protein levels, indicative of pathological pro-inflammatory immune dysregulation. SOD1 mRNA was co-overexpressed…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
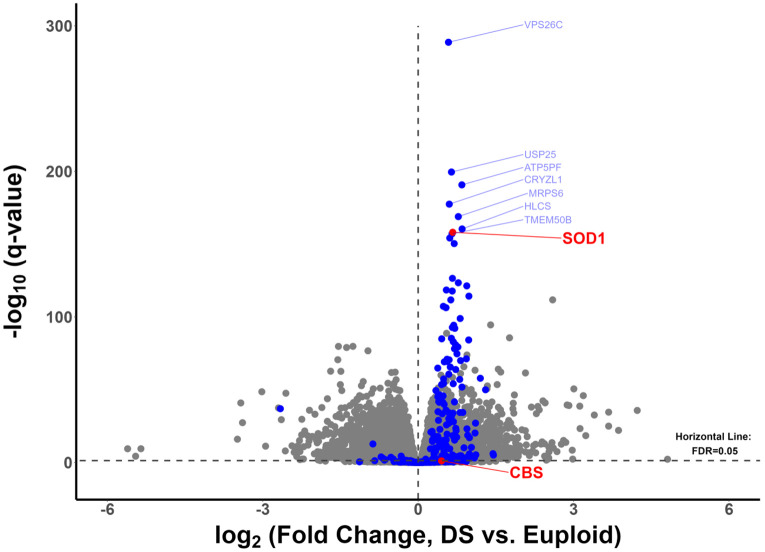 Figure 1
Figure 1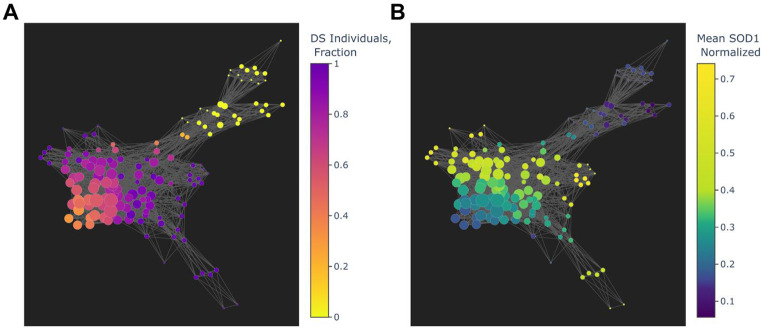 Figure 2
Figure 2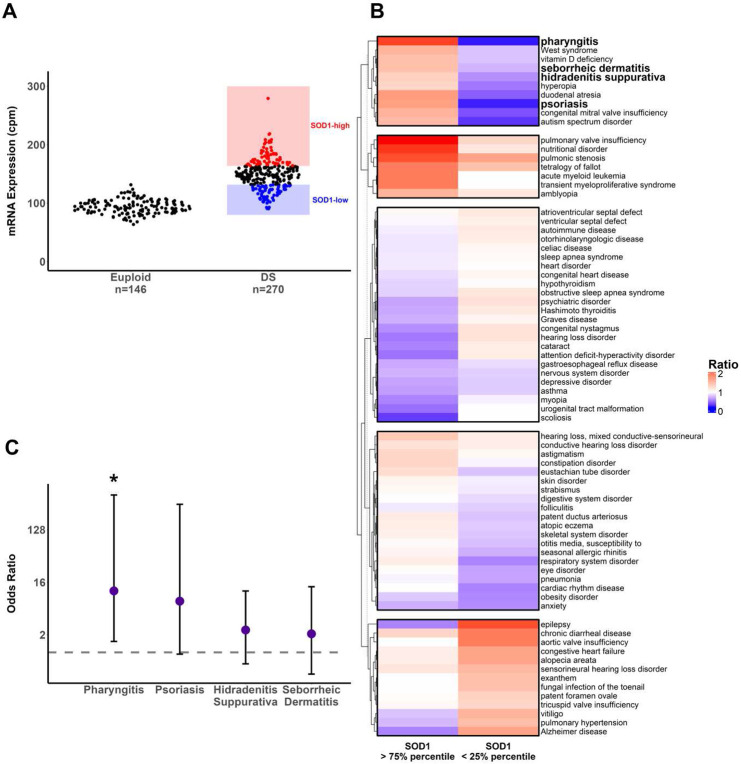 Figure 3
Figure 3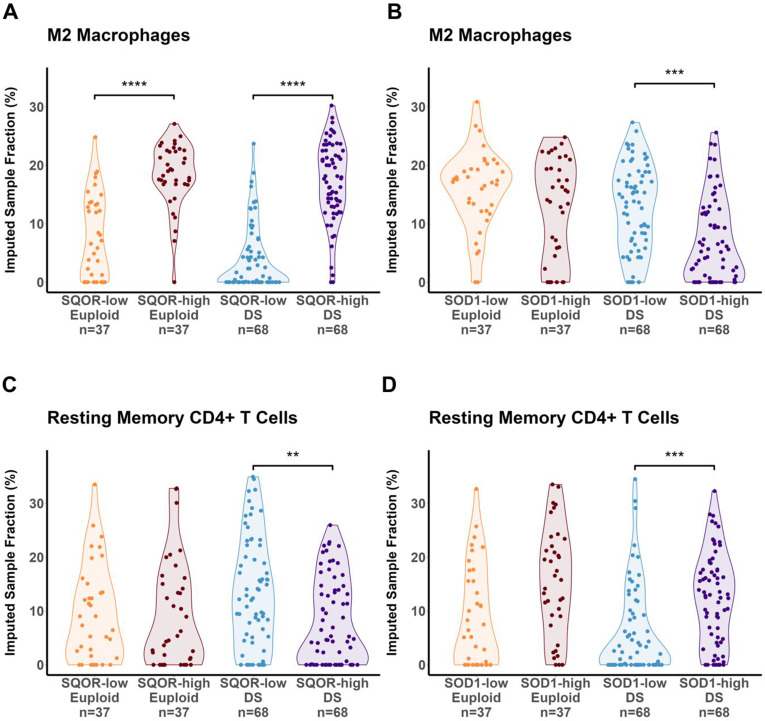 Figure 4
Figure 4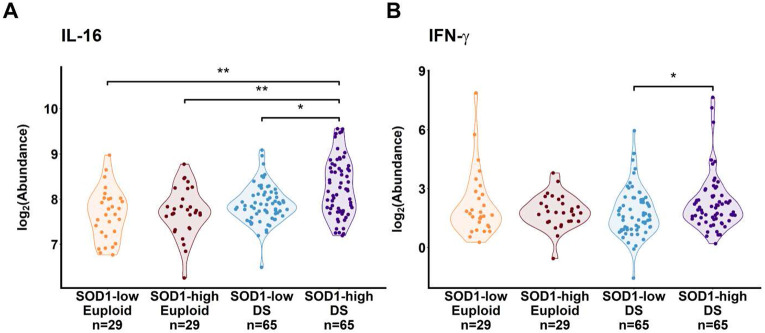 Figure 5
Figure 5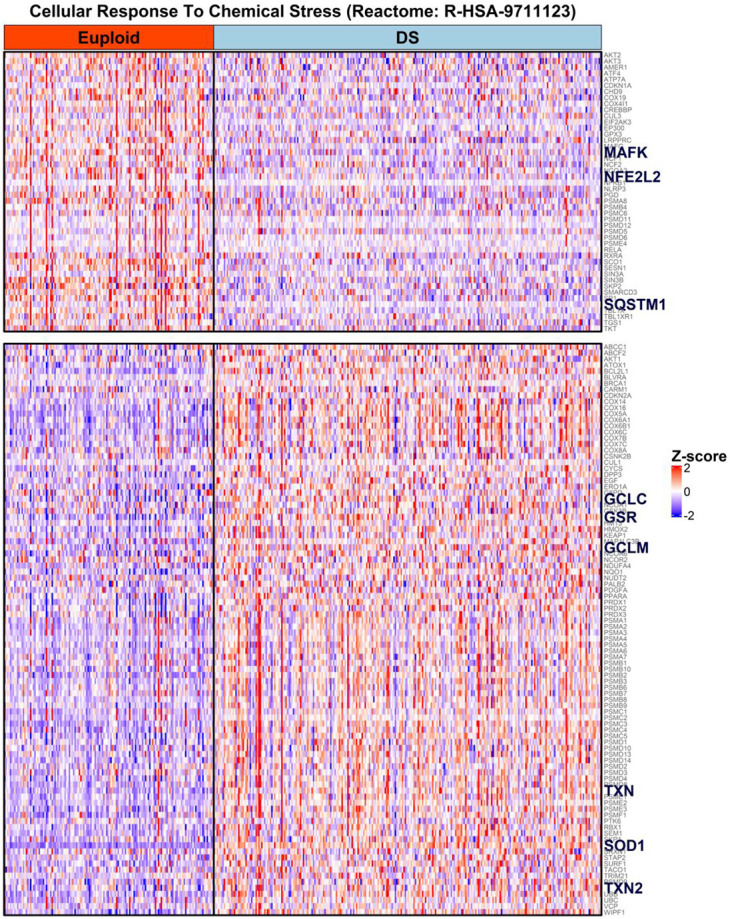 Figure 6
Figure 6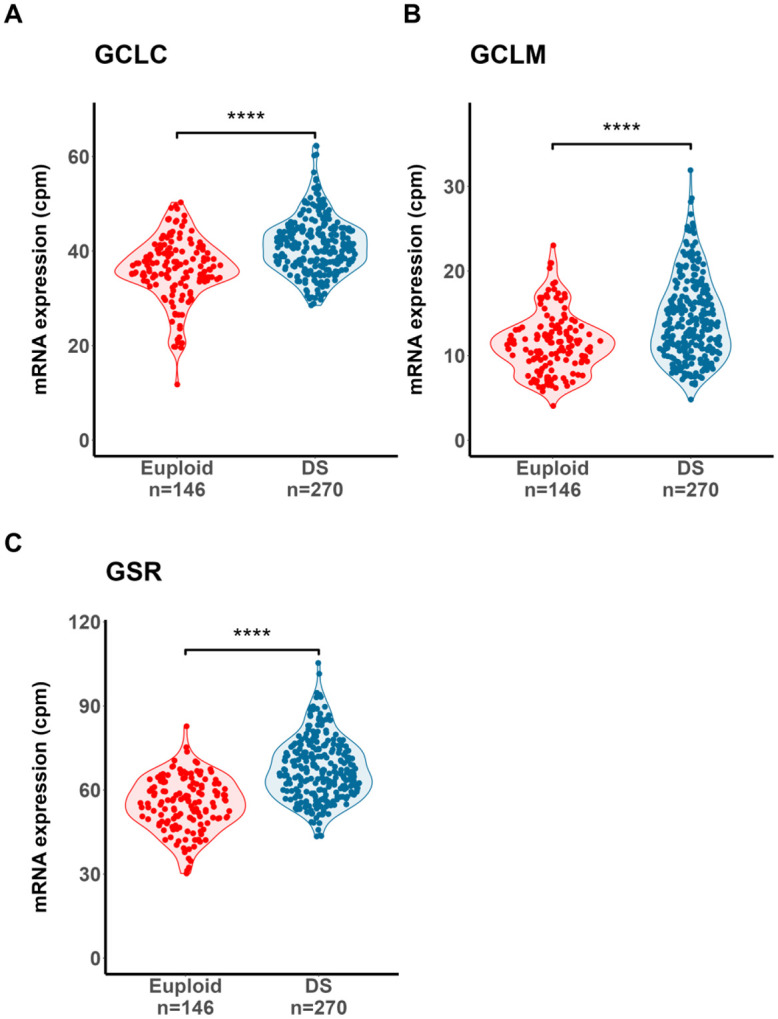 Figure 7
Figure 7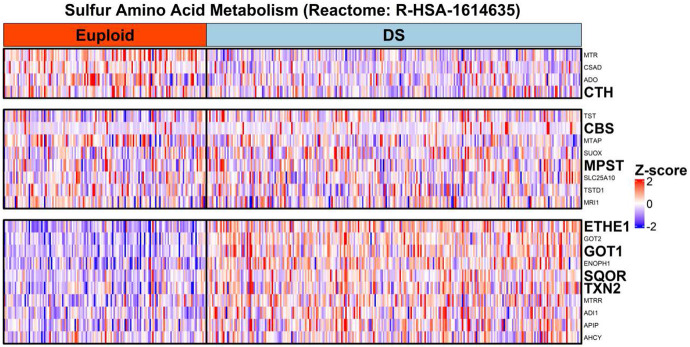 Figure 8
Figure 8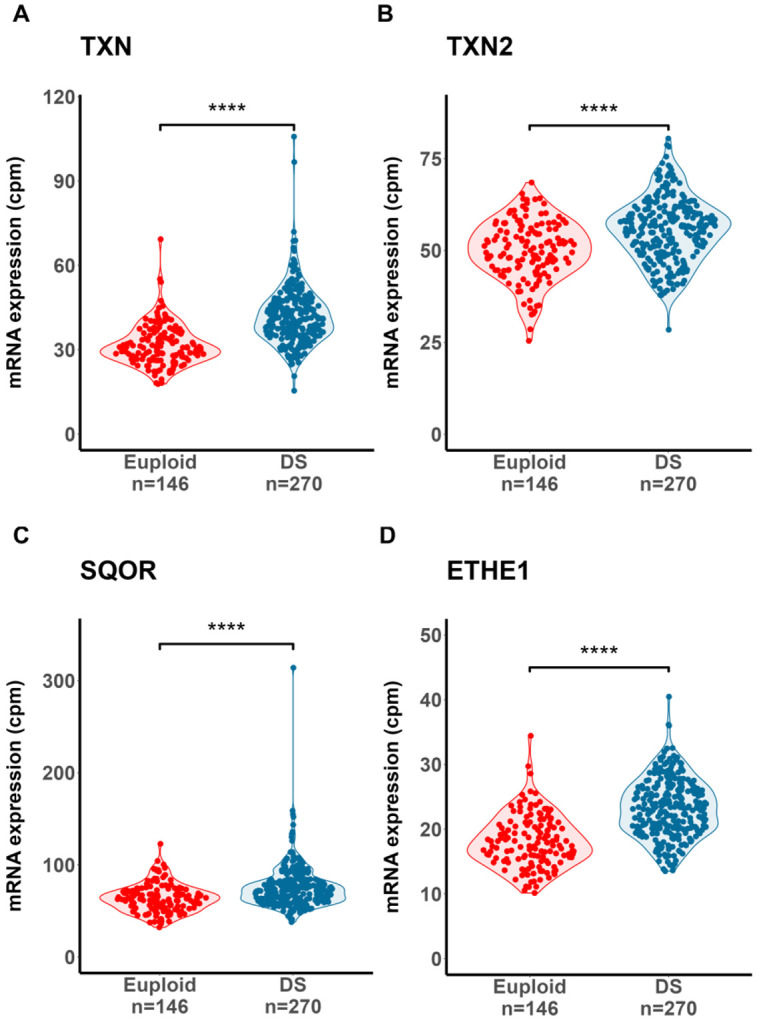 Figure 9
Figure 9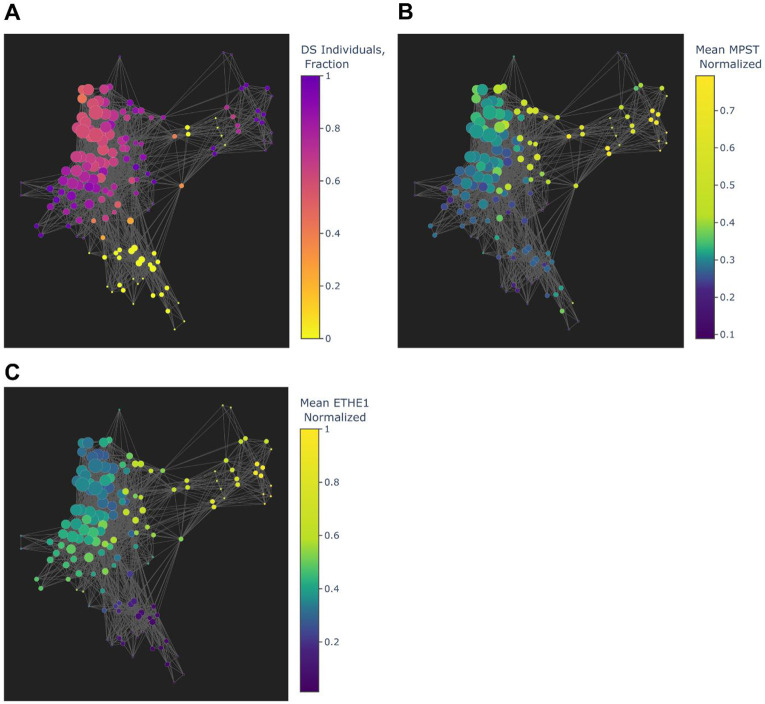 Figure 10
Figure 10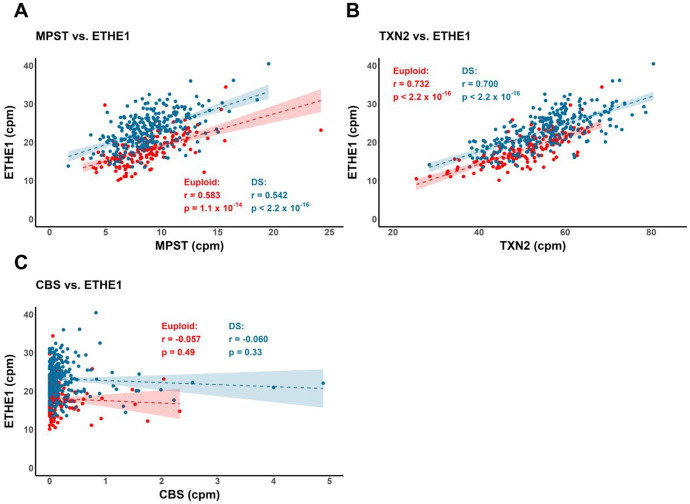 Figure 11
Figure 11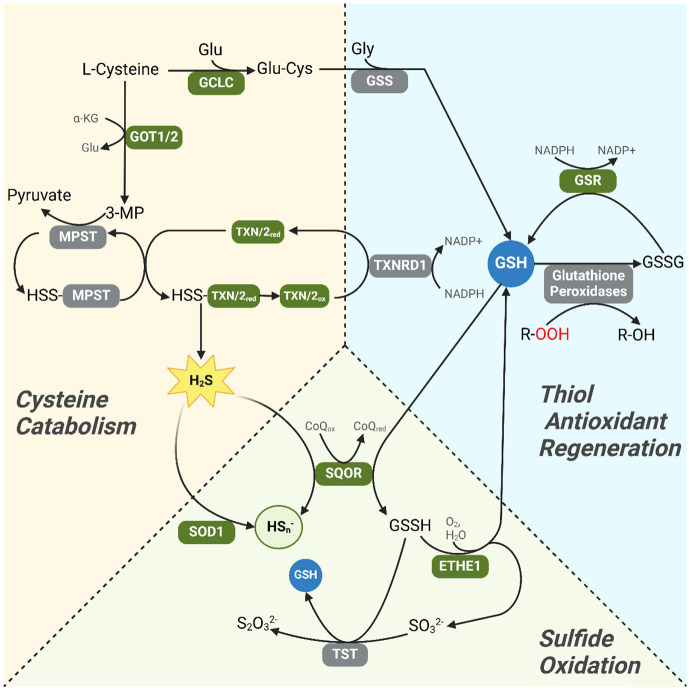 Figure 12
Figure 12- —National Institutes of Health, (TAK) Welch Foundation, TAK
- —National Institutes of Health, LJS
- —National Science Foundation, KRO
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDown syndrome and intellectual disability research · Redox biology and oxidative stress · Cancer-related gene regulation
Introduction:
Down Syndrome (DS), the result of trisomy of human chromosome 21 (Hsa21) trisomy is the most common chromosomal abnormality in humans^1^. While key characteristics of DS include musculoskeletal anomalies, neurocognitive impairment and congenital heart defects, individuals with DS are also at a higher risk of chronic co-occurring conditions including recurrent respiratory infections, autoimmune diseases, hypothyroidism and transient myeloproliferative disorder^1-7^. Increased lifetime risks of immunological and hematological complications in DS have been associated with the pathological accumulation of intracellular oxidative and metabolic stress ^6-8^.
Oxidative and metabolic stress are well characterized features of DS molecular pathophysiology^9^. The overexpression of several Hsa21 genes have been linked to impaired mitochondrial function and electron leakage from the mitochondrial inner membrane electron transport chain, resulting in the generation of toxic reactive oxygen species (ROS)^10^. Notably among Hsa21 resident genes, superoxide dismutase 1 (SOD1) - a key component of the cellular response to ROS - is overexpressed in DS and potentially drives ROS generation. SOD1 catalyzes the dismutation of superoxide radical to hydrogen peroxide (H_2_O_2_)^11^, which is normally decomposed to water and diatomic oxygen by catalase and peroxiredoxins. A SOD1 excess relative to catalase has been observed in DS erythrocytes and was suggested to increase the intracellular accumulation of H_2_O_2_, the precursor to more damaging hydroxyl radicals through reactions with divalent metal ions such as Fe^2+ 6,12-15^. However, the distinct subcellular localizations of SOD1 and catalase – cytoplasmic and peroxisomal, respectively – suggest the SOD1/catalase ratio is insufficient to explain elevated H_2_O_2_ in non-erythrocyte cell types ^16,17^. Moreover, the relationship between the expression of SOD1 and catalysts of cytosolic H_2_O_2_ decomposition remains to be characterized.
In addition to oxidative stress, DS is also characterized by the increased generation of intracellular hydrogen sulfide (H_2_S), a well-documented and potent inhibitor of mitochondrial respiration ^18-20^. Several studies have linked excess H_2_S generation in DS to to the increased expression of another Hsa21 resident gene cystathionine-β-synthase (CBS)^21-23^. CBS catalyzes the conversion of homocysteine to cystathionine as part of the reverse transsulfuration pathway^21-23^. However, we have previously shown an absence of consistently elevated CBS protein in B lymphocytes from DS individuals^18^.Therefore, it seems likely that other metabolic processes catalyzed by non-Hsa21 enzymes contribute to the excess intracellular H_2_S in DS cells.
The increased activity of the enzyme 3-mercaptopyruvate sulfurtransferase (MPST) in DS fibroblasts is one such possibility that has been linked to H_2_S overproduction^20,24^. In addition, H_2_S generation though MPST-mediated cysteine catabolism is dependent on the presence of low molecular thiol peptides such as thioredoxins and glutathione, which are integral components of the cellular thiol antioxidant protein axis and whose activity is upregulated in response to oxidative stress^25,26^.
The potential links between ROS and H_2_S metabolism in DS are strengthened by data demonstrating that in addition to an antioxidant role, SOD1 is also an essential oxidizer of H_2_S ^18,27^. Furthermore, we have shown that the inhibition of SOD1 activity significantly increased H_2_S levels in DS B lymphocytes ^18^. In the current study, the relationship between drivers of increased oxidative stress including SOD1 and the transcription of response effectors was investigated to uncover links between ROS and H_2_S metabolism in DS.
Methods:
Data access:
Patient white blood cell (WBC) bulk messenger RNA (mRNA) sequencing (RNA-seq) and serum proteomics data were collected by the University of Colorado Linda Crnic Institute’s Human Trisome Project, aligned to the human reference genome using RSEM^28^ and made publicly available by the National Institutes of Health Investigation of Co-occurring conditions across the Lifespan to Understand Down syndromE (INCLUDE) data sharing program (https://portal.includedcc.org)^2,3,29^, supported by National Heart, Lung and Blood Institute grant U2CHL156291. All data analyzed in this report were collected under a protocol approved by the Colorado Multiple Institutional Review Board^2^ and de-identified prior to dissemination. Informed consent was obtained from all subjects and/or their legal guardian(s) prior to sample collection. All methods in this study were performed in accordance with the relevant guidelines and institutional regulations.
Analysis of bulk RNA-seq data:
Gene mRNA count data from 270 individuals with Down syndrome (DS) and 146 euploid individuals were aggregated in R 4.4.1 using the dplyr library^30^ (version 1.1.4; https://github.com/tidyverse/dplyr). The age range of participants was between 0.5 and 69.1 years, with 210 female and 206 male individuals. Gene counts were TMM-normalized to counts-per-million (CPM), low-expressed genes (CPM < 1.0) excluded from downstream analyses and differential gene expression in DS versus euploid assessed using edgeR (version 4.0) ^31^. P-values were calculated using the edgeR pipeline’s negative binomial model and likelihood ratio test for DS versus euploid mRNA expression and adjusted for multiple comparisons using the Benjamini-Hochberg procedure ^32^.
Analysis of serum proteomics data:
For individuals with mRNA expression data, serum proteomics data was also available for 260 euploid and 113 DS individuals. Serum proteomics profiles were aggregated, log_2_-normalized and tested for statistical significance between cohorts in R 4.4.1 using the dplyr and stats libraries ^30^.
Gene set enrichment analysis (GSEA) via active protein interaction subnetworks:
GSEA incorporating protein-protein interaction information was performed on log_2_-transformed mRNA expression fold changes obtained from the edgeR pipeline using the pathfindR library (version 2.4.1)^33^. In brief, protein-protein interaction information was obtained from the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database and used to generate subnetworks of biologically related and differentially expressed genes. Subsequently, GSEA was performed on gene subnetworks and results mapped onto Reactome biological pathway gene sets^34^.
Analysis of clinical conditions:
Clinical diagnosis data from each of the 146 euploid and 270 DS individuals was obtained from the INCLUDE data sharing program and aggregated in R^30^. Cohorts of high and low SOD1-expressing DS individuals were defined as the 1^st^ and 4^th^ quartiles of DS SOD1 expression (n = 68 DS individuals, each), respectively. The prevalence of co-occurring conditions was calculated as the number of incidences of a condition divided by the total number of individuals in each cohort. Odds ratios for each condition in SOD1-high versus SOD1-low DS individuals were calculated as (incidences of condition, SOD1-high / incidences of condition, SOD1-low) / (individuals without condition, SOD1-high / individuals without condition, SOD1-low). The calculated odds ratios were tested for statistical significance in R against a null hypothesis of an odds ratio of 1 using Fisher’s exact tests and resulting P-values were adjusted for multiple comparisons using the Benjamini-Hochberg procedure^32^.
Immune Cell Subpopulation Inference:
Cohorts of high and low SQOR- and SOD1-expressing euploid and DS individuals were defined as the 1^st^ (high) and 4^th^ (low) quartiles of SQOR and SOD1 gene expression within each group separately. Bulk mRNA transcriptomics data for individuals in each of the 4 cohorts were deconvoluted using CIBERSORTx (http://cibersortx.stanford.edu) against the default peripheral blood cell signature matrix to obtain inferred immune cell frequencies ^35^.
Visualization:
All heatmaps were generated and hierarchically clustered using R 4.4.1 and the ComplexHeatmap^36^ library. Visualization of individual gene expression patterns across individuals was achieved using the ggforce library^37^. Linear regression analyses and visualization of gene expression P-values against log_2_-transformed fold changes were generated using the ggplot2 library.
Results:
The mRNA expression of most Hsa21 genes were identified as overexpressed (log_2_(Fold Change, DS vs. euploid) > 0) in DS WBCs relative to euploid, as demonstrated by others ^2,3^ (Figure 1). The mean mRNA log_2_(Fold Change) of significantly overexpressed Hsa21 genes was 0.626, corresponding to a fold change of 1.54 consistent with triplication of a constitutively expressed locus in Hsa21 trisomy. SOD1 was among a cluster of Hsa21 genes consistently overexpressed in DS individuals versus euploid. Similarly, mRNA overexpression of Hsa21 genes – NRIP1, SUMO3, DYRK1A, RCAN1, ETS2, PKNOX1/PREP1 - previously linked to decreased mitochondrial respiration efficiency and increased ROS generation – was also identified ^38,39^ (Supplementary Figure 1). In contrast, CBS mRNA was not significantly overexpressed in DS WBCs (Figure 1), implicating SOD1 and not CBS mRNA overexpression as a salient feature of the DS WBC transcriptome.
To profile the heterogeneity of DS and euploid WBC transcriptomes, topological data analysis was performed to group individuals based on similarities in mRNA expression^40,41^. As a metric of biological dissimilarity due to trisomy, an anomaly score was calculated using the Isolation Forest algorithm^42^ on normalized Hsa21 mRNA expression values from DS and euploid patient transcriptomics data. Hsa21 anomaly scores and the first component from Principal Component Analysis (PCA) were used as filter functions for the Mapper algorithm, which was then used to generate graphical representations that grouped individuals with similar transcriptional profiles into nodes^40,43^. Topological data analysis revealed a distinct separation between nodes predominated by DS and euploid individuals, with observed outward flaring of certain DS-dominant nodes suggesting transcriptional heterogeneity within the DS cohort (Figure 2A). While euploid-dominant nodes exhibited consistently lower mean nodal SOD1 expression, we noted considerable variation in SOD1 expression across DS dominant nodes, suggesting that a potential role for variability in SOD1 expression across DS individuals contributes to phenotypic variation in symptom severity.
Next, we determined whether SOD1 mRNA expression in DS individuals was associated with increased frequencies of co-occurring conditions. Cohorts of SOD1 high- and low-expressing DS individuals were defined as the 1^st^ and 4^th^ quartiles of DS SOD1 expression (Figure 3A). The ratios of prevalences of co-occurring conditions in both cohorts relative to all DS individuals in the dataset were calculated and visualized using hierarchical clustering^44^. A cluster of co-occurring conditions with increased prevalences in SOD1-high but decreased in SOD1-low DS individuals (Figure 3B) was observed, including several co-occurring conditions associated with overactive or dysregulated immune cell function. A significantly higher odds ratio for pharyngitis in SOD1-high DS individuals versus SOD1-low was observed. (Figure 3C). Recurrent respiratory infections (i.e. pharyngitis) and increased risk of autoimmunity (i.e. psoriasis and seborrheic dermatitis) have been previously associated with decreased B and T lymphocyte viability in DS^4,7^. Our results demonstrate that SOD1 overexpression may be associated with dysregulated immune cell function in DS, with adverse consequences to patient health.
Another cluster of co-occurring conditions with increased prevalences in the SOD1-low DS cohort, included the prevalence of acute myeloid leukemia (Figure 3B). Both low and high SOD1 expression contribute to the elevated oxidative stress burden in DS. Low SOD1 expression may contribute via decreased dismutation of excess superoxide radicals, and high SOD1 expression via increased generation of hydrogen peroxide, a precursor to more damaging ROS such as the hydroxyl radical (HO^*−^)^14^. Our observations are entirely consistent with previous cellular studies linking elevated oxidative stress in DS to dysregulated hematopoiesis and an elevated risk for leukemia development^5,8^.
Since SOD1 also catalyzes the protective oxidation of H_2_S, we next compared the associations of high SOD1 expression with the canonical H_2_S-oxidizing enzyme sulfide:quinone oxidoreductase (SQOR) and immune cell frequencies in DS. As above, high and low SOD1 and SQOR expression were defined as the 1^st^ and 4^th^ quartiles respectively, in euploid and DS individuals. The bulk mRNA transcriptomic profile of individuals in each high- and low-expression cohort was deconvoluted using CIBERSORTx to infer immune cell subpopulation frequencies ^35^. In both euploid and DS, SQOR-high individuals had a significantly higher mean sample fraction of M2-polarized macrophages relative to respective SQOR-low (Figure 4A). Functionally different from pro-inflammatory M1 macrophages, M2-polarized macrophages are associated with wound healing and inflammation resolution ^45^. The greater M2-macrophage fraction in SQOR-high individuals is consistent with the idea that oxidized sulfur species increase M2 polarization, as shown previously by treatment with the long-acting H_2_S and polysulfide donor GYY4137. ^46-48 49^. It is possible that increased M2 polarization mitigates any pro-inflammatory macrophage activation in DS ^3^. As such, SQOR overexpression may be a potentially protective adaptation. In contrast, significantly lower M2 macrophage fraction in SOD1-high DS individuals was identified relative to SOD1-low DS (Figure 4B), suggesting a possible loss of resilience to a pro-inflammatory milieu with SOD1 overexpression.
Furthermore, significantly lower resting memory CD4+ T cells were also identified in SQOR-high versus SQOR-low DS (but not euploid) cohorts (Figure 4C). In addition, a significantly higher memory CD4+ T cells fraction was also observed in SOD1-high versus SOD1-low DS individuals (Figure 4D). Serum proteomic analysis revealed significantly higher levels of pro-inflammatory cytokines such as interleukin-16 (IL-16) and interferon gamma (IFN-γ) in SOD1-high versus SOD1-low DS (Figure 5A-B). While memory CD4+ T cells are essential for the coordination of innate and adaptive immune responses in healthy individuals ^50^, lower stimulus threshold for activation relative to non-memory cells has implicated them as drivers of autoimmunity ^51^. IL-16 is a crucial chemoattractant and activator of CD4+ T cells, which release IFN-γ to polarize macrophages into the M1 paradigm, thereby potentiating inflammation^52,53^. Indeed, an increased frequency of memory CD4+ T cells and elevated levels of IL-16 and IFN-γ are recognized as components of a hyperactive inflammatory state and correspondingly high risk of autoimmunity in DS ^3,54^. A greater fraction of memory CD4+ T cells is consistent with observations of higher inflammatory disease risk in SOD1-high DS individuals, and merits future investigation using single-cell transcriptomics and in vivo DS mouse models.
Next, to understand whether SOD1 overexpression is associated with altered patterns of DS gene expression biological pathways co-overexpressed with SOD1 mRNA were examined using gene set enrichment analysis (GSEA). GSEA results were correlated with underlying biological mechanisms using active subnetworks of connections between biologically related genes. Individual genes that were significantly over- or under-expressed in DS were identified using the R pathfindR library, with protein-protein interaction information supplied from the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database^33,55^. Subsequently, GSEA was performed on identified gene subnetworks and results mapped onto Reactome biological pathway gene sets^56^, identifying 484 significantly over- and underexpressed gene pathways in DS. Of the 484 pathway gene sets, only 1 pathway included SOD1 (“Cellular Response to Chemical Stress”, Reactome ID # R-HSA-9711123), which contains genes involved in the detoxification of reactive oxygen and nitrogen species (ROS and RNS, respectively). DS-overexpressed genes in this set included those involved in the synthesis (GCLC, GCLM) and regeneration (GSR) of reduced glutathione (GSH), an essential peptide co-reactant for ROS detoxification and the reversal of ROS-mediated injuries^25^ (Figure 6, 7A-C). The co-overexpression of SOD1 with genes involved in glutathione and thioredoxin/peroxiredoxin-mediated reversal of intracellular oxidative damage in DS suggests that oxidative consequences of SOD1 overexpression are linked with a reflexive cellular drive to mitigate ROS-mediated damage.
Also overexpressed in DS were thioredoxins (TXN, TXN2) and thioredoxin-dependent peroxiredoxins (PRDX1, PRDX2, PRDX3). Peroxiredoxins are essential, cysteine-containing enzymes that reduce H_2_O_2_ and organic (i.e. lipid, DNA) peroxides^16^, levels of which are pathologically elevated in cells from DS mouse^15^ and human models^13,18,57^. PRDX2 is the dominantly expressed cytosolic peroxiredoxin in WBCs and neurons ^58^ and was recently shown elevated in DS iPSC-derived neural progenitor cells ^59^. We report a significant positive correlation between SOD1 and PRDX2 mRNA expression in euploid and DS WBCs (Supplementary Figure 2). Also, SOD1/PRDX2 mRNA expression ratios were 23.5% higher in the DS cohort relative to euploid (Supplementary Figure 3). These findings suggest that DS WBCs exhibit excess SOD1-mediated H_2_O_2_ generation relative to cytosolic decomposition versus euploid cells, and that the overexpression of antioxidant effectors may not be sufficient to counteract ROS generation in DS.
Intriguingly, a subset of genes within the same “Cellular Response to Chemical Stress” gene set were significantly underexpressed in DS relative to euploid (NFE2L2, MAFK and SQSTM1) (Figure 6). NFE2L2 encodes the nuclear factor erythroid 2-related factor 2 (Nrf2) transcription factor which is co-activated by transcription factors of the small Maf family (i.e. MAFK) to stimulate the expression of antioxidative response genes^60^. Underexpression of NFE2L2 and MAFK mRNA in DS may therefore suggest an impaired capacity to activate Nrf2-regulated oxidative damage response effectors, potentially indicating decreased resilience to oxidative stress. Furthermore, the under expression of the mitophagy regulator sequestosome 1 (SQSTM1) has been linked to impaired mitophagy and accumulation of damaged mitochondria in DS cells, which exacerbates the oxidative stress burden via increased superoxide generation^61^. Our results are consistent with prior experimental data indicating decreased resilience to, and increased generation of ROS in DS cells ^14,61^ and support compensatory reliance on intracellular thiol antioxidant (glutathione and thioredoxin-mediated) systems.
The observation that SOD1 co-overexpression with thiol antioxidant system effectors has important implications for hydrogen sulfide (H_2_S) generation in DS cells. In addition to their essential role as co-reactants in cellular antioxidant pathways, glutathione and thioredoxin are important components of intracellular H_2_S detoxification and generation pathways respectively ^26,49^. H_2_S overproduction is a recognized feature of DS molecular pathophysiology ^19,62^ and is associated with decreased oxidative phosphorylation ^18,19^. Endogenous H_2_S generation can occur through cystathionine-β-synthase (CBS)- and cystathionine-γ-lyase (CTH)-dependent cysteine synthesis from homocysteine (reverse transsulfuration) and cysteine catabolism via 3-mercaptopyruvate sulfurtransferase (MPST)^26^. In the latter pathway, MPST reacts with the cysteine transamination catabolite 3-mercaptopyruvate, generating a persulfidated enzyme intermediate that can react with thioredoxins to liberate H_2_S^26,63^. Heatmap analysis of genes participating in sulfur amino acid metabolism (Reactome ID #R-HSA-1614635) indicated the overexpression of key genes related to thioredoxin/MPST-mediated H_2_S generation, including thioredoxin 2 (TXN2) and cytosolic (GOT1) as well as mitochondrial aspartate aminotransferase (GOT2), the latter 2 enzymes being the principal source of intracellular 3-mercaptopyruvate generation via cysteine transamination^26,64^ (Figure 8, 9A-B). In contrast, CTH mRNA was significantly under-expressed in DS individuals relative to euploid, and no significant difference in CBS expression was detected between DS and euploid. These results suggest that a cysteine transamination, MPST and thioredoxin-dependent axis drives the generation of excess H_2_S in DS WBCs.
In addition to the CBS- and MPST-catalyzed routes, H_2_S can also be generated by cysteinyl-tRNA synthetases, which were not included in the Reactome list of genes participating in sulfur amino acid metabolism at the time of our analysis. While the expression of the cytosolic cysteinyl-tRNA synthetase 1 (CARS1) was not different between the two cohorts, the expression of the mitochondrial CARS2 was significantly lower in DS (Supplementary Figure 4). Together, these findings support a decreased role of CARS1/2-catalyzed generation toward H_2_S overproduction in DS WBCs.
Overexpression of sulfide:quinone oxidoreductase (SQOR) and persulfide dioxygenase (ETHE1) was also observed in DS versus euploid (Figure 8, 9C-D). SQOR and ETHE1 catalyze the canonical, glutathione-dependent detoxification of H_2_S to polysulfides and sulfite (SO_3_^2−^) respectively, and their overexpression suggests a greater intracellular demand for H_2_S detoxification in DS^49^. To characterize the association, if any, between cysteine catabolism-linked H_2_S generation and H_2_S oxidation pathways in the context of DS, we used topological data analysis to visualize group dissimilarities in gene expression by calculating an anomaly score using the mRNA expression levels of MPST, SQOR, ETHE1, TXN2, TST, CTH, GOT1 and GOT2. We observed a notable separation between DS- and euploid-predominant nodes (Figure 10A), indicating that the expression of non-Hsa21 genes linked to sulfur metabolism can distinguish clusters of DS individuals from euploid independent of CBS expression. The concurrent overexpression of MPST (Figure 10B) and ETHE1 (Figure 10C) in DS-dominant nodes was also observed, suggesting elevated cysteine catabolism-linked H_2_S generation and sulfide oxidation. Additionally, significant positive correlations between MPST, TXN2 and ETHE1 mRNA – but not with CBS – in DS and euploid WBCs were observed, potentially indicating a stronger contributing role of an MPST-mediated H_2_S generation pathway to the sulfide burden in DS cells (Figure 11A-C).
Discussion:
In this report, a novel transcriptomic and proteomic characterization uncovered an increased glutathione- and thioredoxin-dependent intracellular drive correlated with SOD1 overexpression and H_2_S overproduction in DS WBCs. An important limitation of transcriptomic investigations is that despite highlighting networks of gene regulation, mRNA levels do not imply protein levels inside cells. Similarly, serum proteomics does not convey information regarding the tissue-specific distribution and dynamics of inflammation in DS. Furthermore, our analyses of co-occurring clinical conditions are limited by variability in the presentation and reporting of clinical symptoms. Despite these limitations, however,our observations are consistent with a cellular response to reverse oxidative stress-mediated injuries, previously related empirically elevated ROS production and catabolismin multiple cell and animal models of DS^6,8,12,14^. Increased imputed fractions of pro-inflammatory CD4+ T cells and the inflammation-associated cytokines IL-16 and IFN-γ with SOD1 overexpression further implicate it as a contributor to immune dysregulation in DS^3^. Furthermore, the pro-inflammatory effects of SOD1 overexpression contrast with the anti-inflammatory M2 macrophage polarization with that of SQOR. Despite the protective H_2_S oxidizing function, excess SOD1 may negatively alter immune cell homeostasis in DS – a role that merits further investigation.
Others have identified mechanisms leading to the generation of excess oxidative stress in DS cells, such as SOD1 triplication^12,65^, altered proteostasis ^66^ and dysregulated clearance of damaged mitochondria^61^. However, the underlying mechanism(s) supporting resilience to oxidative damage in DS cells remain unclear. Prior clinical studies have shown increased activities of glutathione peroxidases in DS erythrocytes^67,68^ and protein levels of peroxiredoxins 1 and 2 in specific brain regions from DS patients ^69^.
Peroxiredoxins and glutathione peroxidases are enzyme families that reduce the inorganic (H_2_O_2_) and organic peroxides that arise due to ROS-mediated damage ^16^. Both classes depend on thiol-containing peptide co-reactants, thioredoxins and glutathione respectively, as essential reducing agents. Thioredoxins are a family of proteins (~12 kDa) that contain a conserved cysteine-glycine-proline-cysteine (CGPC) motif, allowing intramolecular disulfide bonds to form upon oxidation and constituting an integral aspect of their antioxidant function ^70^. Our findings of increased thioredoxin (TXN, TXN2) gene expression in DS WBCs suggest an increased intracellular demand for thiol antioxidant co-reactants to reverse oxidative damage. Increased transcription also suggests that normal NADPH-dependent regeneration of thioredoxins ^70^ is insufficient to meet heightened demand in DS cells. This idea is consistent with prior studies showing that DS neurons are more susceptible to cell death following inhibition of thioredoxin redox cycling with auranofin and indicate an increased sensitivity to thiol antioxidant depletion ^71^.
In addition to the overexpression of thioredoxins, our analyses uncover evidence for the increased regeneration and de novo synthesis of glutathione in DS. Glutathione is a tripeptide composed of glutamate, cysteine and glycine and is the most abundant endogenous antioxidant in humans^72,73^. As a co-reactant for antioxidant enzymes, reduced glutathione (GSH) donates electrons toward the reduction of organic peroxides, forming oxidized glutathione (GSSG). GSSG is a disulfide derived from two GSH molecules and can be reduced to regenerate GSH upon NADPH-dependent reduction in a reaction catalyzed by the enzyme glutathione reductase (GSR) ^25^. GSH can also be synthesized de novo, and the rate limiting step of GSH synthesis is the conjugation of cysteine to glutamate catalyzed by glutamate cysteine ligase (GCL), encoded by the GCLC and GCLM genes^73^.
The increased GSR gene expression reported here is a potential mechanism to explain reports of higher GSR activities in whole blood from DS individuals relative to euploid^65^, suggesting accelerated glutathione turnover due to increased oxidation. Garlet et al. ^65^ also reported that serum GSH levels were lower in DS individuals relative to euploid^65^. As such, a combination of decreased blood GSH together with elevated glutathione peroxidase activity may be indicative of an increased reliance on GSH-dependent antioxidant pathways leading to accelerated GSH depletion. Increased de novo synthesis of GSH - supported by GCLC and GCLM overexpression– provides a compensatory mechanism to replenish GSH stores within DS cells. Furthermore, GCLC and GCLM overexpression is concordant with increased serum cysteine levels in DS individuals relative to euploid controls ^74^, indicating increased cysteine import linked to de novo GSH synthesis. Our findings motivate further investigations into the post-transcriptional regulation of GCL activity and its contribution to inadequate matching of GSH supply and demand in DS cells.
In addition to its role as an essential antioxidant, GSH is also a co-reactant in mitochondrial sulfur oxidation pathways, an essential mechanism for intracellular H_2_S detoxification ^26,49,75^. In the presence of GSH, sulfide:quinone oxidoreductase (SQOR) acts in mitochondria to oxidize hydrogen sulfide ^49^ SQOR facilitates the transfer of sulfur atoms from H_2_S to form glutathione persulfides (GSSH), which are oxidized by ETHE1 (persulfide dioxygenase) and TST (thiosulfate sulfurtransferase) to yield sulfite and thiosulfate oxidation products respectively^49^. It is possible that H_2_S overproduction in DS contributes to increased GSH production as part of an increased need for H_2_S detoxification, an idea supported by the present findings of increased SQOR and ETHE1 gene expression in DS WBCs. An important caveat is that GSH and thioredoxin-dependent antioxidant effectors can also metabolize H_2_S and other reactive sulfur species (RSS) ^76^. The overlapping cellular responses to RSS and ROS make it difficult to distinguish the effects of one versus the other on the WBC transcriptome. Future experiments are therefore needed to determine the effects of glutathione and cysteine metabolism in DS on cellular responses to ROS and RSS.
Together with increased thiol antioxidant metabolism and regeneration, we report a correlation between increased cysteine catabolism and H_2_S generation in DS WBCs (Figure 12), but not with CBS. A lack of correlation between H_2_S overproduction and CBS levels is supported by our report of elevated H_2_S and polysulfide levels in DS B lymphocytes relative to euploid cells, despite no significant differences in CBS protein levels. ^18^. Differences in cell lineage may account for the notable variability in CBS overexpression across multiple systems in humans and animal models^19^ and thus, future experiments utilizing single-cell RNA sequencing to identify the effect of cell type on CBS expression will be useful. As mRNA expression and protein levels may not reflect protein activity, additional experiments are needed to evaluate post-translational and metabolic control of CBS activity in DS WBCs to better assess the contribution to H_2_S overproduction.
In contrast, our findings of increased aminotransferase and thioredoxin expression implicate MPST as a potential driver of H_2_S overproduction in DS WBCs. MPST catalyzes the decomposition of 3-mercaptopyruvate to pyruvate and H_2_S in the presence of thiol-containing species such as lipoic acid, cysteine and thioredoxins^26^. 3-mercaptopyruvate, in turn, is generated from the transamination of cysteine by aspartate and glutamate aminotransferases (GOT1/2)^75^. GOT1/2 and TXN overexpression in DS. Therefore, these data suggest an enhanced capacity for MPST-mediated H_2_S generation through the excess generation and decomposition of 3-mercaptopyruvate, respectively. While a prior study by Panagaki et al. reported higher levels of MPST in DS dermal fibroblasts relative to euploid controls^24^, we did not observe significantly elevated MPST mRNA expression in DS WBCs^26,75^. It must be noted that MPST-mediated H_2_S generation is sensitive to the concentration of specific sulfide acceptors and increases together with thioredoxin concentrations at physiological levels ^77^. Increased thioredoxin expression in DS cells likely synergizes with MPST activity to drive H_2_S overproduction in the oxidatively stressed DS cell. ^77^
Our findings therefore suggest that H_2_S overproduction is a response to the oxidative consequences of Hsa21 trisomy. One such consequence is increased per-mitochondrion superoxide generation, previously attributed to dysregulated mitochondrial biogenesis and autophagy dynamics ^14,61,78^. As H_2_S directly reacts with superoxide ^79^, reflexive upregulation of H_2_S production may serve as a protective buffer against increased trisomy 21-related mitochondrial ROS generation. H_2_S also serves as the precursor for polysulfides, which readily react with superoxide ^62,80^ and modify cysteine residues by persulfidation. Cysteine persulfidation is protective modification which shields its side chain sulfur from irreversible ROS-mediated oxidation, thus preserving its essential catalytic function in various proteins ^81^. Particularly, the persulfidation of active site cysteines in PRDX2 protect it against H_2_O_2_-mediated overoxidation, which would otherwise lead to the formation of catalytically inactive aggregates ^58,82^. A larger “polysulfide pool” because of increased H_2_S generation may thus promote cellular resilience to ROS in DS, meriting further study. Despite its potential protective effects, it is still possible that H_2_S overproduction represents a “metabolic overcorrection” to increased ROS generation in DS, and future investigations are needed to determine the mechanisms that sustain overproduction in the absence of CBS overproduction.
In summary, we report a series of novel mechanisms underlying a thiol antioxidant axis-driven response to oxidative stress in DS WBCs that provides a possible explanation for H_2_S overproduction. Future studies targeting the crosstalk between ROS and H_2_S metabolism are warranted to improve WBC health and their effects on immune function in DS.
Supplementary Material
Supplement 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Antonarakis S. E. Down syndrome. Nat Rev Dis Primers 6, 9, doi:10.1038/s 41572-019-0143-7 (2020).32029743 PMC 8428796 · doi ↗ · pubmed ↗
- 2Donovan M. G. Variegated overexpression of chromosome 21 genes reveals molecular and immune subtypes of Down syndrome. Nature Communications 15, 5473, doi:10.1038/s 41467-024-49781-1 (2024). · doi ↗
- 3Galbraith M. D. Multidimensional definition of the interferonopathy of Down syndrome and its response to JAK inhibition. Science Advances 9, eadg 6218, doi:10.1126/sciadv.adg 6218. · doi ↗
- 4Ghezzi M. Recurrent Respiratory Infections in Children with Down Syndrome: A Review. Children (Basel) 11, doi:10.3390/children 11020246 (2024). · doi ↗
- 5Hasaart K. A. L., Bertrums E. J. M., Manders F., Goemans B. F. & van Boxtel R. Increased risk of leukaemia in children with Down syndrome: a somatic evolutionary view. Expert Rev Mol Med 23, e 5, doi:10.1017/erm.2021.6 (2021).33902785 PMC 8086399 · doi ↗ · pubmed ↗
- 6Marcovecchio G. E. Premature Senescence and Increased Oxidative Stress in the Thymus of Down Syndrome Patients. Front Immunol 12, 669893, doi:10.3389/fimmu.2021.669893 (2021).34140950 PMC 8204718 · doi ↗ · pubmed ↗
- 7Ram G. & Chinen J. Infections and immunodeficiency in Down syndrome. Clin Exp Immunol 164, 9–16, doi:10.1111/j.1365-2249.2011.04335.x (2011).21352207 PMC 3074212 · doi ↗ · pubmed ↗
- 8Marderstein A. R. Single-cell multi-omics map of human fetal blood in Down syndrome. Nature 634, 104–112, doi:10.1038/s 41586-024-07946-4 (2024).39322663 PMC 11446839 · doi ↗ · pubmed ↗