Functional characterization of a human epilepsy-associated gene network reveals metabolic regulation as a critical factor underlying seizure susceptibilities
Jingyi Long, Spencer G. Jones, Ana Serna, Boyd van Reijmersdal, Franziska Kampshoff, Sara Aibar, Patrik Verstreken, Martijn A. Huynen, Kevin Lüthy, Mireia Coll-Tané, Annette Schenck

TL;DR
This study uses fruit flies to show that metabolism plays a key role in epilepsy by analyzing a network of genes linked to seizures.
Contribution
The study reveals that metabolic regulation is a critical factor in seizure susceptibility through functional validation of a human epilepsy gene network in Drosophila.
Findings
A conserved module of 26 genes, including 13 epilepsy-associated genes, was identified with synaptic and metabolic functions.
Over one-third of Drosophila knockdown models targeting the module showed altered seizure-like behaviors.
Enhancing AMPK activity increased seizure resistance, highlighting metabolism's role in modifying seizure susceptibility.
Abstract
Epilepsy is a mechanistically complex, incompletely understood neurological disorder. To uncover novel converging mechanisms in epilepsy, we used Drosophila whole-brain single-cell RNA sequencing to refine and characterize a previously proposed human epilepsy-associated gene co-expression network (GCN). We identified a conserved co-expressed module of 26 genes, which comprises fly orthologs of 13 epilepsy-associated genes and integrates synaptic and metabolic functions. Over one-third of the Drosophila pan-neuronal knockdown models targeting this module exhibited altered seizure-like behaviors in response to mechanical or heat stress. These knockdown models recapitulated seizures associated with four epilepsy-associated genes and identified two novel epilepsy candidate genes and three genes for which knockdown conferred seizure protection. Most knockdown models with altered seizure…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
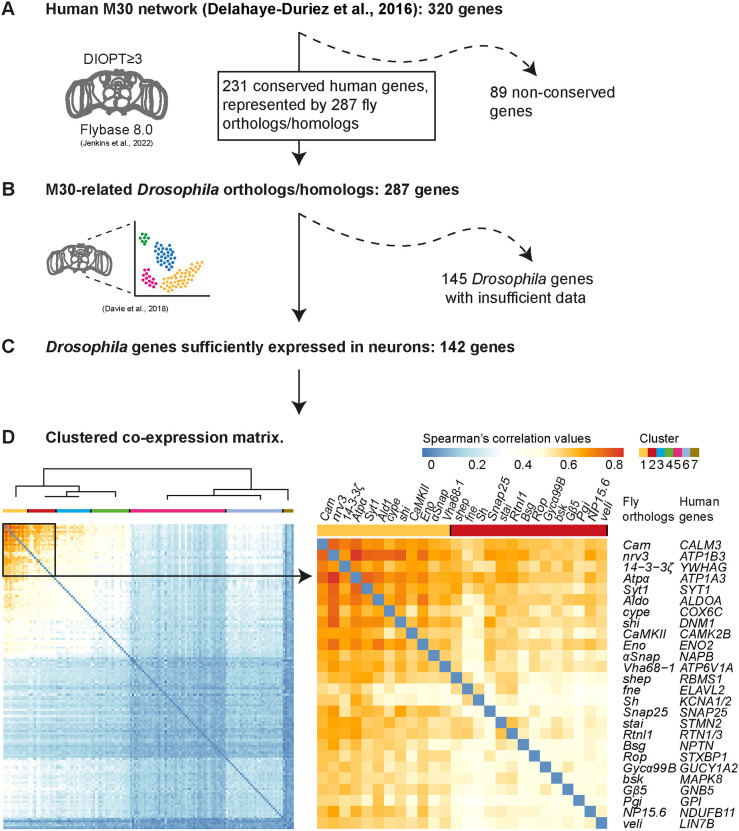 Fig. 1
Fig. 1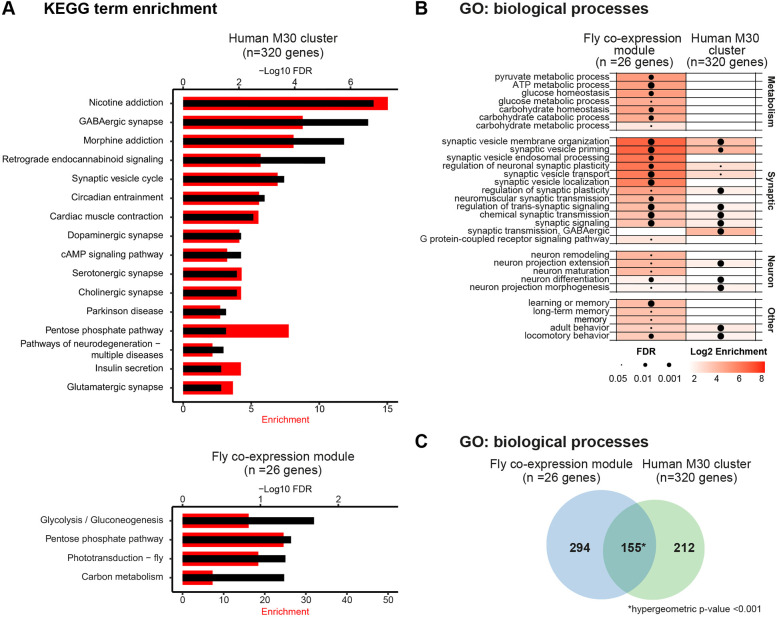 Fig. 2
Fig. 2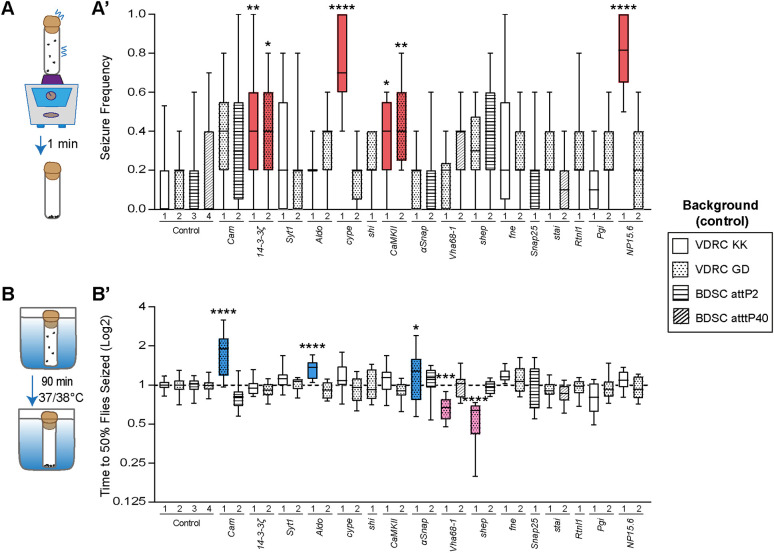 Fig. 3
Fig. 3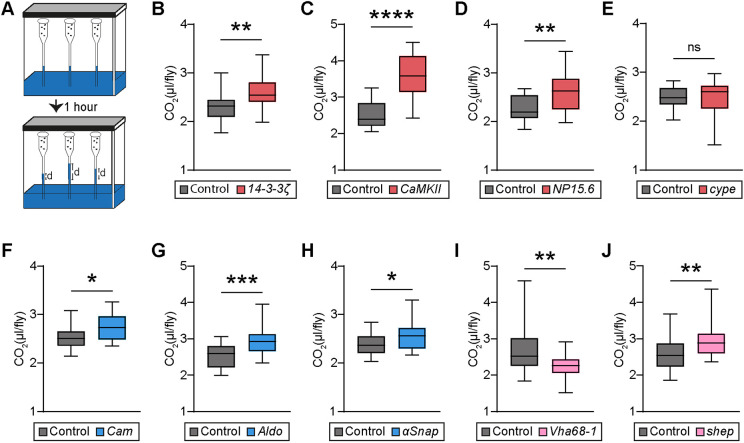 Fig. 4
Fig. 4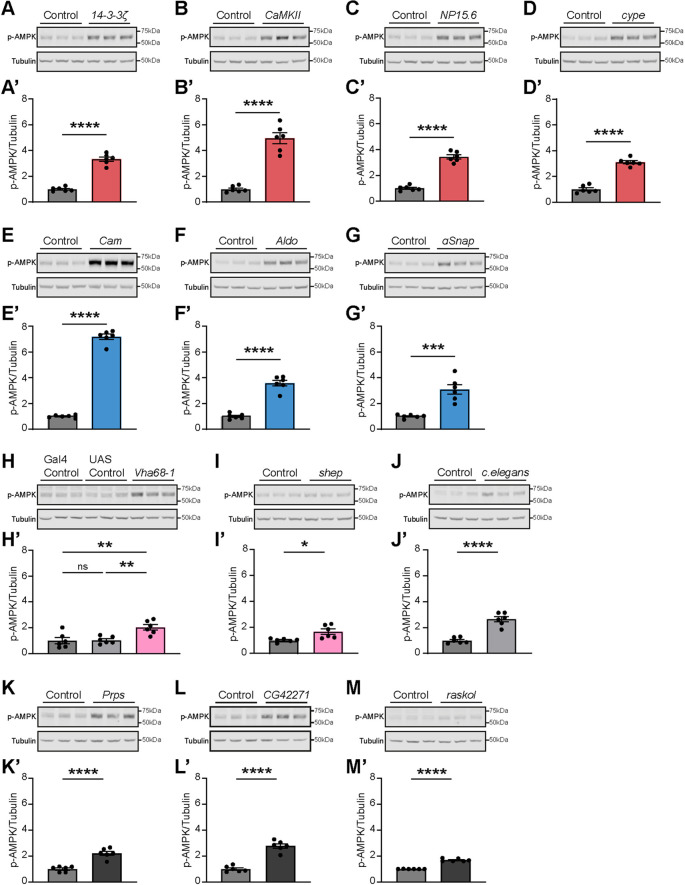 Fig. 5
Fig. 5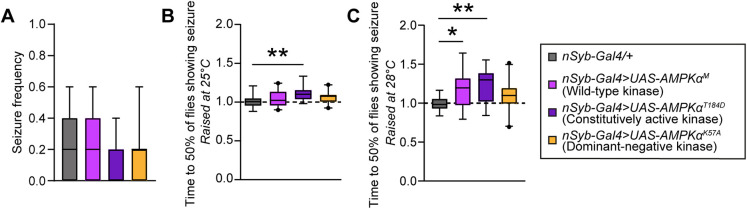 Fig. 6
Fig. 6| Human gene | Established role in epilepsy? | Fly ortholog | |
|---|---|---|---|
| Core cluster (12 genes) |
| No |
|
|
| No |
| |
|
| Yes. Epilepsy gene; OMIM # |
| |
|
| Yes. Epilepsy gene; OMIM # |
| |
|
| Yes. Epilepsy-related gene ( |
| |
|
| No |
| |
|
| No |
| |
|
| Yes. Epilepsy gene; OMIM # |
| |
|
| Yes. Epilepsy-related gene ( |
| |
|
| No |
| |
|
| Yes. Epilepsy gene; OMIM # |
| |
|
| Yes. Epilepsy gene; OMIM # |
| |
| Second subcluster (14 genes) |
| No |
|
|
| No |
| |
|
|
| ||
|
| Yes. Epilepsy-related gene ( |
| |
|
| No |
| |
|
| No |
| |
|
| No |
| |
|
| Yes. Epilepsy gene; OMIM # |
| |
|
| No |
| |
|
| No |
| |
|
| Yes. Epilepsy-related gene ( |
| |
|
| Yes. Epilepsy-related gene ( |
| |
|
| Yes. Epilepsy-related gene ( |
| |
|
| No |
|
- —China Scholarship Councilhttp://dx.doi.org/10.13039/501100004543
- —Radboud Universitair Medisch Centrumhttp://dx.doi.org/10.13039/501100006209
- —ZonMwhttp://dx.doi.org/10.13039/501100001826
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMitochondrial Function and Pathology · CRISPR and Genetic Engineering · RNA Research and Splicing
INTRODUCTION
Epilepsy, characterized by the spontaneous recurrence of unprovoked seizures, impacts over 50 million people globally and manifests in combination with a large number of neurological, cognitive and psychosocial consequences (Fisher et al., 2014; Thijs et al., 2019). Despite recent advances, epilepsy treatment remains challenging, with about one-third of patients showing resistance to current anti-epileptic drugs (Mesraoua et al., 2023). This emphasizes the need to explore the biological underpinnings of epilepsy to enable the development of novel treatment strategies. Next-generation sequencing technologies have markedly accelerated the identification of genetic factors implicated in epilepsy and suggested that genetic predisposition contributes to up to 80% of epilepsy cases (Hildebrand et al., 2013; Thakran et al., 2020). Variants in more than 1500 genes have been linked to epilepsy (Zhang et al., 2023), yet their underlying pathogenic mechanisms and how they contribute to epileptogenesis remain to be determined.
Systems biology and network analyses are potent methods for exploring the molecular processes and pathways involved in diseases (Parikshak et al., 2015). Gene co-expression network (GCN) analysis is useful for identifying clusters of genes that show similar expression patterns under various conditions and thus likely share biological functions (Van Dam et al., 2018). This approach has revealed a first pro-convulsant gene network and proposed sestrin 3 as a regulator among diverse epilepsies (Johnson et al., 2015). Nonetheless, validating the multitude of genes within GCNs remains a significant challenge, as experimentation is both costly and time-intensive (Van Dam et al., 2018). Delahaye-Duriez and colleagues used genome-wide gene expression data from previously published post-mortem healthy human brains (Ramasamy et al., 2014) to perform weighted gene co-expression network analysis and differential co-expression analysis. They identified a co-expression network of 320 genes (termed M30 network) that is enriched in genes associated with monogenic and polygenic epilepsy, providing opportunities for epilepsy treatment exploration (Delahaye-Duriez et al., 2016). They also reported concerted (dys)regulation of the network in several epilepsy-related datasets, but experimental investigation of potential convergent mechanisms was not attempted.
Drosophila melanogaster is an excellent model to study the etiology of epilepsy owing to its efficiency, cost-effectiveness and ease of genetic manipulation. Assays to induce seizure-like behaviors are well established, and 81% of human epilepsy genes have orthologs in this organism (Fischer et al., 2023). The availability of comprehensive single-cell brain expression datasets in Drosophila (Davie et al., 2018) provided a novel opportunity to functionally characterize GCNs associated with epilepsy. Using the previously identified M30 network, we isolated a highly co-expressed module of 26 genes with conserved neuronal co-expression in this evolutionarily distant organism for in-depth examination. Genetic manipulation of several individual genes in the fly co-expression module induced seizure-like behavior, identifying known epilepsy-associated genes as well as novel candidate genes and modifiers. It also revealed strong functional coherence of the module linked to regulating metabolic rate, implicating a novel converging neuronal mechanism in a specific group of genetic epilepsies.
RESULTS
Identification of a conserved epilepsy-associated gene co-expression module in Drosophila
To provide novel insights into the genetic causes of epilepsy and reveal shared mechanisms, we integrated a previously identified gene co-expression network enriched in epilepsy-associated genes generated based on human post-mortem brain tissue bulk RNA-sequencing (RNA-seq) data (the M30 network, comprising 320 genes) (Delahaye-Duriez et al., 2016) with single-cell gene expression data in Drosophila (Davie et al., 2018) (Fig. 1).
A conserved neuronal co-expression module of 26 Drosophila genes based on the human epilepsy-associated M30 network and whole-brain Drosophila single-cell RNA-sequencing (scRNA-seq) atlas. Workflow diagram representing the steps from the human epilepsy-associated M30 gene network (Delahaye-Duriez et al., 2016) to the identification of the highly co-expressed genes 26 genes. (A) Annotation of M30 Drosophila orthologs identified using FlyBase (Jenkins et al., 2022). (B) Extracting genes with >5000 normalized scRNA-seq counts. Data derived from Davie et al. (2018). (C) Calculations of the pairwise Spearman's ρ values and clustering. (D) The hierarchically clustered heatmap (left) represents the co-expression of 142 Drosophila genes. It highlights a module of 26 highly co-expressed fly genes (indicated with a black square on the left, magnified on the right). Each line or column represents a unique gene. The color indicates co-expression values. To the right of the 26×26 gene matrix, the fly orthologs of the human M30 genes are indicated. The 26-gene module is composed of two clusters: the 12 most co-expressed genes in the core cluster (top left, cluster 1, in yellow) and 14 further genes in cluster 2 (in red).
First, we used the Drosophila RNAi Screening Center Integrative Ortholog Prediction Tool (DIOPT) scores (Hu et al., 2021) to determine the conservation and Drosophila orthologs of the 320 human genes of the M30 network (Table S1; see Materials and Methods). We identified 287 M30-related Drosophila genes, encompassing one-to-one, many-to-one (a sole fly gene representing more than one human gene) and one-to-many (more than one fly gene with a similar DIOPT score representing a human gene) orthologs (Fig. 1A; Table S2). To determine the co-expression of these genes in Drosophila, we retrieved the whole-brain single-cell RNA-sequencing (scRNA-seq) data from Davie et al. (2018) (Fig. 1B). Pairwise Spearman's correlation values were calculated for the 142 Drosophila genes with sufficient expression across most neuronal subtypes present in the dataset (see Materials and Methods) [Fig. 1C; genes listed in Table S3; Spearman's ρ values (r_s_) in Table S4]. To unravel which genes show the highest degree of expression similarity and, hence, are most likely functionally related, hierarchical clustering was performed and represented in a heatmap (Fig. 1D). Visual inspection identified a highly co-expressed module of 26 genes, distinct from the background of lowly correlated genes. These 26 genes belonged to two hierarchical clusters. The first ‘core’ cluster included the 12 genes with the highest co-expression values (0.52<r_s_<0.84). The second cluster of 14 genes was characterized by r_s_ values ranging between 0.36 and 0.61 within this cluster, and r_s_ values up to 0.68 with genes in the top-scoring core cluster.
Thirteen genes in both co-expressed clusters are associated with epilepsy (Table 1), as classified in Online Mendelian Inheritance in Man (OMIM) and documented in PubMed. In summary, we identified an epilepsy-associated module comprising 26 genes characterized by evolutionary conserved co-expression.
The epilepsy-associated and highly conserved co-expression module links genes with synaptic and metabolic functions
As a first step toward characterizing the molecular nature of the 26-gene module we identified, we performed Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway and Gene Ontology (GO) enrichment analysis comparing this module against the genome-wide background using g:Profiler (Raudvere et al., 2019). Whereas the human M30 cluster was highly enriched in genes relevant to neural processes, such as synaptic transmission, synaptic vesicle transport and gamma-aminobutyric acid (GABA) signaling (Delahaye-Duriez et al., 2016) [confirmed by our recent query (Fig. 2A, top)], three of the four significantly enriched KEGG pathways of our co-expression module were related to carbohydrate metabolic processes (Fig. 2A, bottom). Similarly, GO-term analysis for biological processes associated with our module ranked carbohydrate-related metabolic processes among the top-scoring ontology terms – e.g. ‘glucose homeostasis’ [false discovery rate (FDR)=1.82×10^−3^, fold enrichment=28.69], ‘ATP metabolic process’ (FDR=4.96×10^−4^, fold enrichment=27.66) and ‘carbohydrate metabolic homeostasis’ (FDR=0.047, fold enrichment=5.41) – in addition to synaptic and other neuronal processes (Fig. 2B). We also identified other enriched functions among the 26-gene module, neither identified by Delahaye-Duriez et al. (2016) nor us when performing GO-term analysis of the human M30 genes. These include G-protein-coupled receptor signaling and cognitive functions such as learning and memory (Fig. 2B; Table S5). In total, 449 significantly enriched GO terms were identified for the co-expressed Drosophila 26-gene module versus 367 for the human M30 cluster. Of these, only 155 overlapped between the human and Drosophila GO terms (Fig. 2C). Whereas this overlap is significant, as expected from a smaller list selected from a larger one, the large divergence is remarkable. In particular, the increased number of significantly enriched GO terms despite the lower number of genes (and hence potential statistical power) illustrates that our approach to identifying evolutionary conserved, biologically coherent clusters within the M30 cluster worked.
*The 26-gene co-expression module links genes with synaptic and metabolic functions. (A) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of the genes in the human M30 network (top) and the Drosophila co-expression module (bottom). The fly co-expression module's enriched KEGG pathways primarily relate to carbohydrate metabolic processes, whereas these are not enriched among the genes of the human M30 network. For each KEGG pathway, the bars show the −log10 false discovery rate (FDR) (black, axis on the top) and the fold enrichment (red, axis on the bottom). (B) Enriched Gene Ontology (GO) biological processes of the Drosophila co-expression module and the human M30 cluster. Only significantly enriched, representative GO biological processes (FDR<0.05) are shown. A full list of significant terms is shown in Table S5. The size of the dots represents the FDR, and the color intensity of each grid represents the log2 enrichment. The carbohydrate metabolism process ranked highly in the GO-term analysis of biological processes. (C) Venn diagram illustrating the overlap of GO biological processes that are enriched in our Drosophila co-expression module compared with the human M30 cluster. P<0.001 (hypergeometric test).
Finally, we evaluated the contribution of epilepsy-associated genes and their gene orthologs to the M30 and our 26-gene module. According to current knowledge (OMIM and PubMed; see Material and Methods), there were 31 epilepsy genes and 22 epilepsy-related genes among the 320 human genes in the M30 cluster (Table S6), together corresponding to 17% (53/320 genes). Among the Drosophila 26 genes are seven epilepsy genes and six epilepsy-related genes, together summing up to 50% (13/26 genes) and a statistically significant threefold enrichment in epilepsy-associated genes compared to the human M30 cluster (P=3.45×10^−5^). This suggests that the functional themes identified in our module are highly relevant to epilepsy.
More than a third of the neuronal knockdown fly models in the co-expression gene module have altered seizure susceptibility
To address whether and which of the identified co-expressed Drosophila genes can be implicated in seizure-like behaviors, we took advantage of the UAS-Gal4 system (Brand and Perrimon, 1993) and the nSyb-Gal4 driver to induce pan-neuronal knockdown models by RNA interference (RNAi) (Dietzl et al., 2007; Perkins et al., 2015). Knockdown appears to be a valid approach, because all 13 epilepsy-associated genes in our 26-gene module (Table 1) have been linked to loss-of-function variants. We systematically targeted the core cluster, encompassing the 12 genes with the highest co-expression and seven genes in the second interrelated cluster, which we selected based on high co-expression values, molecular and biological functions (Tables S4 and S5), and available tools. In total, we used 44 RNAi lines (from four RNAi libraries) to knock down 19 genes, with at least two independent constructs per gene whenever available. In parallel, we crossed the nSyb-Gal4 driver to the corresponding genetic background control lines of each of the RNAi lines. We found that pan-neuronal knockdown of nrv3 (RNAi-1, -2 and -3), Atpα (RNAi-1 and 2), Cam (RNAi-3), Aldo (also known as Ald1) (RNAi-3), Eno (RNAi-1 and 2), shi (RNAi-2), cype (RNAi-3), Vha68-1 (RNAi-3), ɑSnap (RNAi-3), shep (RNAi-3) and Rtnl1 (RNAi-2) led to developmental lethality (Table S7), which precluded further evaluation of seizure-like behavior.
Seizure susceptibility in adult flies can be assessed upon exposure to mechanical (bang-sensitive) or heat stress, which are believed to be caused by different underlying molecular mechanisms (Fischer et al., 2023; Mituzaite et al., 2021). We evaluated both types of seizure-like behaviors in the adult viable RNAi models. In the mechanically induced seizure-like behavior assay, flies are placed in a laboratory vortex at the highest speed to hyperstimulate sensory inputs and assess the resulting seizure-like behavior (Kuebler and Tanouye, 2000) (Fig. 3A). Seizure-like behavior is reflected by uncontrolled wing and leg movements while flies lay on their backs or by paralysis (Kuebler and Tanouye, 2000; Lasko and Lüthy, 2021). Pan-neuronal knockdown of 14-3-3ζ (RNAi-1 and -2), CaMKII (RNAi-1 and -2), cype (RNAi-1) and NP15.6 (RNAi-1) showed significantly increased seizure frequency compared with their isogenic controls (Fig. 3A′). Of these four genes, the human orthologs of all but cype are known to be associated with epilepsy (Morleo and Franco, 2018; Ramocki et al., 2010; Rizzi et al., 2020) (Table 1). The increased seizure-like behavior in Drosophila and its presence in the epilepsy-associated highly co-expression module identify COX6C, the human ortholog of cype, as a potential epilepsy-associated gene.
*Pan-neuronal knockdown models with altered seizure susceptibility upon either mechanical or heat stress induction. (A) Schematic representation of the paradigm used to assess seizure-like behavior upon mechanical stress. (A′) Seizure frequency in all viable screened RNA interference (RNAi) lines and isogenic background controls, crossed with pan-neuronal driver nSyb-Gal4. Red indicates knockdown models that have significantly higher seizure frequency. Pan-neuronal knockdown of 14-3-3ζ (RNAi-1 and 2), cype (RNAi-1), CaMKII (RNAi-1 and 2) and NP15.6 (RNAi-1) leads to a significant increase in seizure frequency compared to that in their genetic background controls (14-3-3ζ RNAi-1, P=0.0025 and RNAi-2, P=0.014; cype RNAi-1, P<0.0001; CaMKII RNAi-1, P=0.014 and RNAi-2, P=0.0012; NP15.6 RNAi-1, P<0.0001). Kruskal–Wallis test with Dunn's correction for multiple testing. N=12 vials per RNAi line, four to six males per vial. (B) Schematic representation of the paradigm used to assess seizure-like behavior upon heat stress. (B′) Normalized average time to which 50% of the flies are seized in all viable screened RNAi lines and isogenic background controls, crossed with pan-neuronal driver nSyb-Gal4. Pink indicates knockdown models that show a significantly shorter time until 50% of the flies show seizure-like behavior at 37°C or 38°C compared to their genetic controls (Vha68-1 RNAi-1, P=0.0007; shep RNAi-1, P<0.0001), indicating that they have higher heat sensitivity and are more prone to seizures. Blue indicates knockdown models that show a significant increase in the time it takes flies to show seizure-like behavior (Cam RNAi-1, P<0.0001; Aldo RNAi-1, P<0.0001; ɑSnap RNAi-1, P=0.018), indicating that they have lower heat sensitivity and are protected against seizure susceptibility. One-way ANOVA with Šídák correction for multiple testing. N=10-12 vials per RNAi line, four to six flies per vial. Data are represented as boxplots that extend from the 25th and 75th percentiles, with the median indicated. Whiskers indicate the fifth and 95th percentiles. *P<0.05, **P<0.01, ***P<0.001, ***P<0.0001. BDSC, Bloomington Drosophila Stock Center; VDRC, Vienna Drosophila Resource Center.
Next, we investigated which of our models show seizure-like behavior upon heat stress (Burg and Wu, 2012; Mituzaite et al., 2021) (Fig. 3B). Flies that exhibit increased susceptibility to seizure-like behavior drop to the bottom of the vials faster than wild-type animals and show uncontrolled movements or paralysis. We found that pan-neuronal knockdown of Vha68-1 (RNAi-1) led to a significant increase in heat-sensitive seizure-like behavior compared with their isogenic background controls (Fig. 3B′). Notably, its human ortholog, ATP6V1A, is an established epilepsy gene (Fassio et al., 2018) (Table 1). The Drosophila ortholog of RBMS1, shep (RNAi-1), also showed strikingly increased susceptibility to seizure-like behavior (Fig. 3B′). Interestingly, pan-neuronal knockdown of Cam (RNAi-1), Aldo (RNAi-1) and ɑSnap (RNAi-1) caused decreased heat sensitivity, suggesting a protective role against heat-sensitive seizure-like behavior (Fig. 3B′). Taken together, our experimental approach revealed altered seizure susceptibility in more than a third (11 of 29, 38%) of the tested neuronal knockdown models within the Drosophila co-expression module. It effectively recapitulated the seizures of four epilepsy-associated genes, identified two novel epilepsy candidate genes and proposed seizure-protective effects for another three genes.
A majority of pan-neuronal knockdown models with altered seizure susceptibility have alterations in metabolic rate
The enrichment of KEGG pathways and GO terms related to carbohydrate and ATP metabolism among the identified co-expression module (Fig. 2A,B), along with the fact that epileptic seizures are marked by increased neuronal activity and higher ATP requirements (Rho and Boison, 2022), prompted us to investigate whether changes in the metabolic state occur more widely among the models with altered seizure susceptibility. Carbon dioxide (CO_2_) production can serve as a reliable indicator of substrate oxidation and energy expenditure, thus providing a valuable indication of the metabolic state (Van Voorhies et al., 2004). We measured CO_2_ production using respirometers that measure changes in gas volume when CO_2_ is absorbed, which causes the pressure to drop and the liquid columns in the respirometers' capillaries to rise (Yatsenko et al., 2014) (Fig. 4A). First, we tested the four genes that exhibited increased seizure frequency upon mechanical stress. Pan-neuronal knockdown of 14-3-3ζ, CaMKII and NP15.6 resulted in a significant increase in CO_2_ production compared with that in isogenic controls (Fig. 4B-D), illustrating increased metabolic rates. Pan-neuronal knockdown of cype did not affect the metabolic rate (Fig. 4E).
*Pan-neuronal knockdown models with seizure-like behavior show altered metabolic rate. (A) Schematic representation of the paradigm used to assess metabolic rate by a respirometry system, used to measure CO2 production. Changes in gas volume after CO2 absorption led to a decrease in pressure and an increase in the fluid level inside the respirometer capillaries within a sealed chamber. The distance by which the liquid ascends in the micropipette after 1 h is depicted as distance ‘d’. (B-E) CO2 production in the pan-neuronal knockdown models (nSyb-Gal4>UAS-RNAi) with increased seizure frequency in the mechanically induced seizure assay (in red) and their isogenic background controls (genetic background of the respective RNAi line crossed with the same driver, nSyb-Gal4/+). (B-D) Pan-neuronal knockdown of 14-3-3ζ (B), CaMKII (C) and NP15.6 (D) leads to a significant increase in CO2 production and thus in their metabolic rates (14-3-3ζ, P=0.0041; CaMKII, P<0.0001; NP15.6, P=0.0019). (E) Pan-neuronal knockdown of cype has no effect on metabolic rate (P=0.95). (F-H) CO2 production in the pan-neuronal knockdown models with lower heat sensitivity (in blue) and their isogenic background controls. Pan-neuronal knockdown of Cam (F), Aldo (G) and ɑSnap (H) lead to a significant increase in CO2 production and thus in their metabolic rate (Cam, P=0.017; Aldo, P=0.0003; ɑSnap, P=0.026). (I,J) CO2 production in the pan-neuronal knockdown models with higher heat sensitivity (in pink) and their isogenic background controls. (I) Pan-neuronal knockdown of Vha68-1 leads to lower CO2 production and thus a decrease in its metabolic rate (P=0.0096). (J) Pan-neuronal knockdown of shep leads to an increase in CO2 production and thus in its metabolic rate (P=0.0098). N=18-24 groups per genotype, five flies per group. Data are represented as boxplots that extend from the 25th and 75th percentiles, with the median indicated. Whiskers indicate the minimum and maximum. Two-tailed unpaired t-test or Mann–Whitney test, based on the normality of the distribution. *P<0.05, **P<0.01, ***P<0.001, ***P<0.0001.
Surprisingly, pan-neuronal knockdown of the three genes that caused lower heat sensitivity (an indication of seizure protection)− Cam, Aldo and ɑSnap−also resulted in increased CO_2_ production (Fig. 4F-H). Furthermore, flies with pan-neuronal knockdown of the two genes causing increased heat sensitivity, Vha68-1 and shep (Fig. 4I,J), showed significantly dysregulated metabolic rate as well, albeit in opposite directions, with Vha68-1 knockdown flies being the only one among the tested models exhibiting a decrease in metabolic rate (Fig. 4I). Collectively, our findings show that pan-neuronal knockdown models exhibiting alterations in seizure susceptibility show metabolic dysregulation. Notably, this also applies to genes not associated with the identified metabolic KEGG and GO terms, such as shep and aSnap, which have no previous reported function in energy metabolism/regulation of metabolic rate (Table S8).
Neuronal models with altered seizure susceptibility show increased AMPK phosphorylation
Adenosine monophosphate-activated protein kinase (AMPK) plays a pivotal role in safeguarding cellular energy homeostasis. Cellular energy stress, such as that by glucose deprivation (Zhang et al., 2017) or low ATP levels (Hardie and Carling, 1997), causes AMPK activation by phosphorylation (Lin and Hardie, 2018). This, in turn, increases catabolism and decreases anabolism through the phosphorylation of key proteins in multiple pathways, including mTOR complex 1 (mTORC1), lipid homeostasis, glycolysis and mitochondrial homeostasis (Herzig and Shaw, 2018). Therefore, we asked whether increased metabolic rates in the Drosophila pan-neuronal knockdown models with seizure-like behavior are associated with changes in AMPK phosphorylation. We addressed this question by subjecting head extracts of the above-investigated knockdown models to quantitative western blot analysis using anti-phosphorylated AMPK (p-AMPK) antibodies. This revealed that pan-neuronal knockdown of 14-3-3ζ, CaMKII, NP15.6, Cam, Aldo and ɑSnap, all characterized by increased metabolic rate, led to a significant threefold or higher increase in p-AMPK levels compared to those in their respective isogenic controls (Fig. 5A-C′,E-G′). Pan-neuronal knockdown of Vha68-1, causing heat-sensitive seizure-like behavior and a decrease in metabolic rate, and shep, causing heat-sensitive seizure-like behavior and an increase in metabolic rate, also resulted in a modest, but significant, increase (fold change<2) in AMPK phosphorylation (Fig. 5H-I′). Surprisingly, the pan-neuronal knockdown of cype also manifested a significant rise in AMPK phosphorylation (Fig. 5D,D′), despite maintaining a normal metabolic rate. Together, these results show that alterations in seizure susceptibility in most pan-neuronal knockdown models are paralleled by variable increases in AMPK phosphorylation levels.
*Pan-neuronal knockdown models with altered seizure susceptibility, but also negative controls with induced non-sense or unrelated RNAi, show different degrees of increased AMPK phosphorylation. (A-M′) Representative western blots (A-M) showing three biological replicates of head homogenates from adult flies containing nSyb-Gal4 and UAS-RNAi genetic elements or their genetic background controls using anti-phosphorylated AMPK (p-AMPK) and anti-tubulin antibodies, and their quantification (A′-M′). The remaining western blots are presented in Fig. S1. p-AMPK levels are normalized against the tubulin loading control. (A-G′) Pan-neuronal knockdown of 14-3-3ζ (A,A′), CaMKII (B,B′), NP15.6 (C,C′), cype (D,D′), Cam (E,E′), Aldo (F,F′) and ɑSnap (G,G′) leads to a significant increase in p-AMPK levels compared to those in their genetic background controls (14-3-3ζ, P<0.0001; CaMKII, P<0.0001; NP15.6, P<0.0001; cype, P<0.0001; Cam, P<0.0001; Aldo, P<0.0001; ɑSnap, P=0.0002). (H,H′) Pan-neuronal knockdown of Vha68-1 leads to a slight but significant increase in p-AMPK levels compared to those in their genetic background controls (Vha68-1, dark gray: P=0.0064; light gray: P=0.0083). Flies containing only the UAS-Vha68-1-RNAi construct show similar levels of p-AMPK than flies containing only nSyb-Gal4 (P=0.9989). (I,I′) Pan-neuronal knockdown of shep leads to a slight but significant increase in p-AMPK levels compared to those in their genetic background controls (shep, P=0.01). (A′-I′) Colors according to behavioral phenotypes in epilepsy assays, as in Figs 3 and 4. (J,J′) Pan-neuronal induction of a UAS-RNAi construct targeting a C. elegans gene leads to a significant increase in p-AMPK levels compared to those in its genetic background control (P<0.0001). (K-M′) Pan-neuronal knockdown of Prps (K,K′), CG42271 (L,L′) and raskol (M,M′) show a significant increase in p-AMPK levels compared to those in their genetic background controls (Prps, P<0.0001; CG42271, P<0.0001; raskol, P<0.0001). Data are shown as mean±s.e.m. (A′-G′,I′-M′) Two-tailed unpaired t-test. (H′) One-way ANOVA with Šídák correction for multiple testing. ns, not significant; *P<0.05, **P<0.01,***P<0.001,***P<0.0001.
To further investigate the increase in AMPK phosphorylation, which we observed in our nSyb-Gal4>UAS-RNAi models without exception, we included additional controls. First, we investigated a non-specific control fly line in which nSyb-Gal4 drives an RNAi hairpin sequence targeting a Caenorhabditis elegans gene with no target in Drosophila. Unexpectedly, pan-neuronal induction of the *C. elegans-*specific RNAi also resulted in increased AMPK phosphorylation (Fig. 5J,J′). To determine whether a UAS-RNAi construct alone could affect AMPK phosphorylation without being induced, we representatively evaluated p-AMPK levels in flies carrying the UAS-Vha68-1-RNAi construct but not the nSyb-Gal4 (UAS control). We found no differences in the levels of p-AMPK between this and our standard Gal4 control (Fig. 5H,H′). Furthermore, we tested three additional RNAi lines targeting orthologs of the human M30 cluster, which are not included in the 26-gene co-expression module. Pan-neuronal knockdown of Prps, CG42271 and raskol all resulted in increased AMPK phosphorylation (Fig. 5K-M′). Combined, these data argue that activation of the RNAi machinery itself appears to affect AMPK phosphorylation, regardless of the targeted gene. However, whereas pan-neuronal knockdown of the M30 orthologs not associated with our module (Prps, CG42271, raskol, the C. elegans-specific RNAi), as well as Vha68.1 and shep knockdown resulted in a modest increase in p-AMPK levels by 1.69- to 2.78-fold, pan-neuronal knockdown of 14-3-3ζ, NP15.6, Aldo and, above all, CaMKII and Cam strongly elevated levels of p-AMPK (3.33- to 7.17-fold change compared to isogenic controls), suggesting that these genes regulate AMPK phosphorylation specifically.
AMPK regulates seizure susceptibility
To address the causality of increased AMPK phosphorylation levels, questioned by at least some degree of unspecific activation upon RNAi, for seizure-like behavior, we aimed to determine whether neuronal manipulation of AMPK in flies alters their seizure susceptibility. We pan-neuronally overexpressed either UAS-AMPKα^M^, UAS-AMPKα^T184D^ or UAS-AMPKα^K57A^, a wild-type, a constitutively active and a dominant-negative form of the kinase, respectively (Johnson et al., 2010; Swick et al., 2013). In the mechanically induced seizure-like behavior assay, the expression of neither UAS-AMPKα^M^, UAS-AMPKα^T184D^ nor UAS-AMPKα^K57A^ had any effect on seizure frequency (Fig. 6A). In contrast, in the heat induced seizure-like behavior assay, the expression of the constitutively active AMPK (UAS-AMPKα^T184D^) resulted in a slight, but significant, reduction in heat sensitivity (longer time to seizure) compared to isogenic controls (Fig. 6B). Because the efficiency of the UAS-Gal4 system is increased at higher temperatures (Brand and Perrimon, 1993), we also evaluated heat-sensitive seizure-like behavior in flies that were reared at 28°C instead of 25°C, and induced seizures at 42°C instead of 38°C. Under these stringent conditions, the reduced heat sensitivity phenotype caused by the overexpression of constitutively active AMPK (UAS-AMPKα^T184D^) was more pronounced, and the overexpression of wild-type AMPK (UAS-AMPKα^M^) also acquired the phenotype (Fig. 6C). Expression of the dominant-negative AMPK allele had no effect on heat-induced seizure susceptibility (UAS-AMPKα^K57A^; Fig. 6B,C). In summary, increasing AMPK levels and/or activity is sufficient to improve resistance to heat induced seizure-like behavior in a dose-dependent manner.
*Overexpression wild-type and constitutively active AMPK alters heat induced seizure susceptibility. (A) Seizure frequency upon mechanical stress is not altered in flies overexpressing either wild-type (nSyb-Gal4>UAS-AMPKɑM), a constitutively active (nSyb-Gal4>UAS-AMPKɑT184D) or a dominant-negative form of AMPK (nSyb-Gal4> UAS-AMPKɑK57A) under the control of pan-neuronal driver, compared to their background control. N=18 vials, five males per vial. Kruskal–Wallis test with Dunn's correction for multiple testing. (B) Pan-neuronal overexpression of a constitutively activate form of AMPK leads to increased average time for 50% of the flies to be showing seizure-like behavior (nSyb-Gal4>UAS-AMPKɑT184D, P=0.0012) compared to their background control, indicating that they have a lower heat sensitivity and thus a protective effect towards seizure susceptibility. N=18-20 vials, four to five males per vial. One-way ANOVA with Dunnett's correction for multiple testing. (C) Pan-neuronal overexpression of wild-type and constitutive active AMPK leads to lower heat sensitivity (nSyb-Gal4>UAS-AMPKɑM, P=0.023; nSyb-Gal4>UAS-AMPKɑT184D, P=0.0011). Flies were raised at 28°C and exposed to a temperature of 42°C to induce seizures. N=16-21 vials, five males per vial. One-way ANOVA with Dunnett's correction for multiple testing. Data are represented as boxplots that extend from the 25th and 75th percentiles, with the median indicated. Whiskers indicate the fifth and 95th percentiles. *P<0.05; *P<0.01.
DISCUSSION
Research on epilepsy has evolved from focusing on single-gene investigations to studying genetic networks and functional processes. Despite significant advancements, the sheer number of genes often identified in bioinformatics approaches, such as genome-wide GCNs, poses challenges for validating hypotheses and further studying experimental phenotypes and mechanisms further. In the current study, we leveraged Drosophila whole-brain scRNA-seq data to refine the previously reported epilepsy-associated M30 network (Delahaye-Duriez et al., 2016). This identified a highly co-expressed cluster of 26 genes, the composition of which suggested a functional connection between synaptic machinery and energy metabolism in epilepsy. Through genetic manipulation and metabolic rate measurements, we confirmed this prediction in several Drosophila models, uncovering a convergence of epilepsy genes operating at the synapse on the regulation of metabolic state. The molecular mechanisms underlying seizure and metabolic modulation in our module appear to still be heterogeneous but include dysregulation of AMPK signaling, activation of which alone was sufficient to confer protection against heat-induced seizure-like behavior.
Relevance of the Drosophila co-expression module to epilepsy
Utilizing Drosophila whole-brain scRNA-seq data, we defined a highly conserved co-expression module of 26 genes, which includes fly orthologs of 13 epilepsy-associated genes (50%). Compared to 17% of epilepsy-associated genes in the human M30 cluster, this highlights increased relevance of the Drosophila module to epilepsy. Moreover, our experimental models successfully recapitulated increased seizure susceptibility linked to four epilepsy-associated genes (14-3-3ζ, CaMKII, NP15.6 and Vha68-1). Both YWHAG (14-3-3ζ in Drosophila) and ATP6V1A (Vha68-1 in Drosophila) are classified as epilepsy genes linked to developmental and epileptic encephalopathy (OMIM #617665 and OMIM #618012, respectively). CAMK2B (CaMKII in Drosophila) is related to intellectual disability that can occur with co-morbid epilepsy (Kury et al., 2017) (OMIM #617799). NDUFB11 (NP15.6 in Drosophila), a component of mitochondrial complex I, is implicated in microphthalmia with linear skin defects syndrome, which includes seizures (Morleo and Franco, 2018) (OMIM #300952). Therefore, our models provide a robust platform for validating and further exploring genes involved in epilepsy.
In addition, our findings suggest a seizure-modulatory, protective effect upon the knockdown of three genes, Aldo, Cam and ɑSnap. ALDOA, the human ortholog of Aldo, is a key enzyme of glycolysis (Morais et al., 2017). Interestingly, although neuronal excitability has been attributed to ion channels and synaptic transmission, glucose metabolism also plays a crucial role in its regulation. Glycolysis inhibition has been shown to reduce neuronal excitability (Shao et al., 2018), highlighting the importance of metabolic pathways in modulating seizure susceptibility. Calmodulin (Cam in Drosophila) plays a crucial role in calcium signaling by binding to Ca^2+^, which activates CaMKII (Junho et al., 2020). Several anticonvulsants exert calcium-antagonistic effects, suggesting that calmodulin inhibition might also reduce seizures (Chaisewikul et al., 2001; Walden et al., 1992). Drosophila ɑSnap and its human orthologs NAPA and NAPB are members of the evolutionarily conserved soluble N-ethylmaleimide-sensitive fusion attachment protein (SNAP) family involved in vesicle docking and fusion (Andreeva et al., 2006). Our data and previous studies have shown that loss of α/βSNAP alters seizure susceptibility (Burgalossi et al., 2010; Mignon-Ravix et al., 2023).
Lastly, our study also identified two novel candidate epilepsy genes, based on ‘guilt by association’ (Piovesan et al., 2015), through our co-expressed cluster and the obtained seizure-like phenotypes in Drosophila: RBMS1 (shep) and COX6C (cype). RBMS1 encodes an RNA-binding protein that may contribute to epilepsy through the misregulation of its mRNA targets, while COX6C encodes a subunit of complex IV in the mitochondrial respiratory chain, crucial for oxidative phosphorylation (OXPHOS) (Wang et al., 2022). The connection between epilepsy and OXPHOS function is further supported by NDUFB11 (NP15.6), a gene also in our cluster, associated with seizure-like behavior in our model as well as with seizures in patients (Morleo and Franco, 2018). Because mitochondrial OXPHOS deficiency accelerates glycolysis (Zheng, 2012), this aligns with our observed protective effect of ALDOA knockdown – which would reduce glycolytic flux – and highlights the role of metabolic regulation within our co-expression module and its relevance to epilepsy. Evidence for disease association of RBMS1 (shep) and COX6C (cype) is, however, missing at this point in time.
Deregulated energy metabolism as a common denominator of altered seizure susceptibility in the Drosophila co-expression module
Unbiased KEGG pathway and GO-term enrichment analyses revealed a contribution of carbohydrate metabolic processes to the fly co-expression module. Strikingly, we experimentally determined that altered energy metabolism (i.e. metabolic rate) was not at all limited to Aldo (the only viable condition of the three genes – Aldo, Eno and Pgi – operating in carbohydrate metabolic processes) but was a highly penetrant feature among the pan-neuronal knockdown models with altered susceptibility to seizures. Of note, this included models for which the underlying gene has been previously connected to energy metabolism (14-3-3ζ, CaMKII, *NP15.*6, Cam and Vha68-1; see Table S8) as well as genes with no previous indication for a regulatory function in energy metabolism (shep and aSnap; Table S8).
There is accumulating evidence for brain metabolic dysfunction, particularly hypermetabolism, contributing to epilepsy (Rho and Boison, 2022). Hypermetabolism is frequently considered a compensatory response, especially in situations in which increased neuronal activity drives higher energy consumption (Ashraf et al., 2015; Liotta et al., 2024). A direct contribution of hypermetabolism to epilepsy is further supported by the clinical efficacy of ketogenic diet (D'andrea Meira et al., 2019) and anti-epileptic drugs such as the lactate dehydrogenase-inhibitor stiripentol (Sada et al., 2015). Most of our knockdown models also exhibited increased metabolic rates, leading us to propose that hypermetabolism, also a common feature of OXPHOS deficiency (Sturm et al., 2023), plays a significant role in regulating seizure susceptibility.
However, there is also a body of literature reporting hypometabolism in epilepsy as well as pro-epileptic effects of glycolytic inhibition. The reasons for the observed discrepancies have been proposed to include acute versus chronic effects (Liotta et al., 2024; Samokhina et al., 2017; Stafstrom et al., 2009), different types of metabolic disorders (Bartolini et al., 2023; Klepper et al., 2020), variations in epileptogenic zones (Carvalho et al., 2022; Schur et al., 2018), differences between excitatory and inhibitory deficits (Bowser and Khakh, 2004; Chassoux et al., 2016), and positive feedback or compensatory effect of dynamic changes (Koenig and Dulla, 2018; Otahal et al., 2014), all of which can, in part, contribute to opposite effects. Thus, the relationships between brain metabolism and epileptic seizures are complex (Rho and Boison, 2022). This complexity may also be underlying our finding of hypermetabolism in both subgroups of our models, those with increased as well as with decreased susceptibility for seizure-like behavior. Further investigating the metabolic substrates that fuel increased metabolic rate in our models could hold some answers. Regardless of this knowledge gap, our study revealed a group of disorders that represent highly feasible candidates for metabo-therapeutic interventions.
AMPK signaling in Drosophila models with altered seizure-like behavior
To ask whether the identified role of the co-expression module in the control of metabolic rate can be explained by convergence onto a specific molecular pathway, we turned to AMPK, an energy and glucose sensor (Lin and Hardie, 2018). Our pan-neuronal knockdown models with altered seizure susceptibility showed increased AMPK phosphorylation, although regardless of their metabolic rate (with p-AMPK also being increased in cype with unaltered, and Vha68-1 with decreased, metabolic rate). To further investigate this surprising finding, we assessed additional controls. These revealed a modest but significant increase in AMPK phosphorylation upon RNAi targeting genes not associated with our 26-gene co-expression module as well as a control with RNAi against a non-Drosophila gene. Of note, this finding was specific to induced RNAi; we neither observe changes in p-AMPK levels in non-induced UAS-RNAi models or Gal4-only controls. This finding is alarming as it suggests a target-independent effect of the activated RNAi machinery on AMPK signaling, making it challenging to interpret some of our findings. However, p-AMPK levels in controls were considerably lower than in some of our pan-neuronal knockdown models (14-3-3ζ, NP15.6, Aldo and, particularly, Cam and CaMKII). Although these findings alone are not sufficient to explain the absence versus presence of seizures, or the directionality of changes in metabolic rate, inducing expression of either wild-type or constitutively active AMPK construct conferred a dose-dependent protective effect against heat-induced seizures. This aligns with studies reporting that time-restricted feeding had an anticonvulsant effect and resulted in increased p-AMPK levels, as well as with findings that metformin, an AMPK activator, shows potential anti-epileptic effects (Alnaaim et al., 2023; Landgrave-Gomez et al., 2016). Altogether, we propose that increased AMPK activation underlies accelerated metabolism to alleviate seizure susceptibility in some of our models yet conclude that other mechanisms must contribute. Hence, our co-expression module appears to converge on altered metabolic rate, but not on a uniform underlying molecular mechanism.
Limitations and future directions
Despite the discovery of metabolic dysregulation in a number of rare epilepsy disorder models linking synaptic and metabolic mechanisms, our study has some limitations. In addition to the complex relationship between metabolic rate, p-AMPK levels and altered seizure susceptibility, further limitations are based on decisions that have guided our bioinformatics approach. Specifically, we refined the highly epilepsy-associated human M30 gene network using data from the first Drosophila scRNA-seq brain dataset (Davie et al., 2018), which, although pioneering, had less sequencing depth compared to more recent studies (Jovic et al., 2022; Li et al., 2022). We also decided to focus our study on co-expression in neurons, given that the core physiological feature of epileptic seizures is neuronal hyperexcitability (Holmes and Ben-Ari, 2001). However, an increasing number of studies provide compelling evidence for glial involvement in the pathophysiology of epilepsy (Patel et al., 2019). Further investigations into whole-brain or perhaps even glia-specific co-expression can be expected to contribute additional converging themes in epilepsy.
In conclusion, we have shown that single-cell expression data and experimental follow-up in Drosophila can refine human disease-relevant GCNs, highlighting the potential of integrating cross-species data to enhance our understanding of human diseases. Our work identified a co-expressed gene module, conserved across evolution and operating at the interface of synaptic and metabolic function, as a converging mechanism in epilepsy. We consider this particularly interesting considering emerging evidence for local glycolytic and mitochondrial mechanisms in nerve terminals/at synapses (Faria-Pereira and Morais, 2022; Li and Sheng, 2022; Myeong et al., 2023 preprint; Pulido and Ryan, 2021). Our findings that genetically disturbing synaptic key players (CaMKII and Vha68-1), reversely, lead to changes in metabolic rate indicates previously unappreciated mechanisms, crosstalk or feedback mechanisms. To further understand these will provide fundamental insights that may open novel opportunities to interfere with intertwined synaptic and metabolic epileptic pathologies.
MATERIALS AND METHODS
Gene orthology and scRNA-seq co-expression analysis
The orthologs of human genes from the epilepsy-associated M30 network (Delahaye-Duriez et al., 2016) were identified using FlyBase (release FB2020_02) (Gramates et al., 2022; Jenkins et al., 2022) with DIOPT scores more than or equal to three. Of these, 163 M30 genes were identified as one-to-one fly orthologs. In addition, 37 M30 genes represented members of shared, conserved gene families with reduced redundancy in the fly, leading to the inclusion of 17 many-to-one Drosophila orthologs. Lastly, 31 M30 genes had more than one single Drosophila homolog with high and similar DIOPT scores, leading to the inclusion of 107 one-to-many fly genes, together resulting in a catalog of 287 unique M30-related Drosophila genes. scRNA-seq data from whole-brain Drosophila neurons were obtained from a previously published dataset (Davie et al., 2018; Hwang et al., 2018). The dataset ‘Aerts_Fly_AdultBrain_Filtered_57k.loom’ was downloaded from SCope, under ‘Davie_et_al_Cell_2018’, and the expression matrix and the cell type annotations were extracted using the SCopeLoomR package. Of the 287 M30-related Drosophila genes, 142 were retained after requiring a minimum expression of 5000 reads across all cells in the dataset. To focus on relevant neuronal populations, a coherent dataset of defined neuronal cell types representing diverse neuronal functions was selected, including populations of serotonergic, tyraminergic, peptidergic, dopaminergic and octopaminergic neurons (adPN, adPN/C15, adPN/C15&kn, adPN/kn, adPN/kn&CG31676, AstA/NPF, AstA/Nplp1, Capa, CCAP, CCHa1, Clock, Crz, DCN, DN1, Dopaminergic, dorsal_Fan-shaped_Body, FMRFa, Gr43a, Hsp, Hug, ITP, L1, L2, L3, L4/L5, Lamina_monopolar, Lawf1, Lawf2, LNv, lPN, lPN/CG31676, lPN/unpg, MBON, Mip, Mip/ITP, Mip/OCT, Octopaminergic, Olfactory_projection_neurons, PAM, Peptidergic, Poxn, Proc/Gpb5, Proc/Ms, Serotonergic, Tyraminergic). This selection excluded cell types that had previously been reported to possess strongly distinct transcriptomics profiles as they could bias the correlation analysis [glial cells, optic lobe neurons, photoreceptors and Kenyon cells (Davie et al., 2018; Hwang et al., 2018)]. Gene co-expression relationships were evaluated by calculating pairwise Spearman's correlation coefficients across all selected neuronal cells (i.e. correlation on the 142 genes×5980 cells log2-transformed matrix). Correlation values were assembled into a gene-gene correlation matrix. Genes were hierarchically clustered based on pairwise correlation distances to identify co-expression modules, hclust[as.dist(1-geneCor)], and visualized using the NMF::pheatmap function.
Classification of epilepsy-associated genes and statistical analysis
We divided human orthologs of genes in the fly co-expression module into three categories: epilepsy genes, epilepsy-related genes and genes not associated with epilepsy to date. For this, human orthologs were first searched in the OMIM database. Genes with mentioning of epilepsy or an epilepsy-related term (such as epileptic encephalopathy) in the main phenotype description field of the Gene-Phenotype Relationships table were categorized as ‘epilepsy gene’. Genes for which epilepsy or seizures were listed in OMIM's clinical features section or for which we found case reports in the literature, retrieved by search terms ‘epilepsy’ or ‘seizure’, were categorized as ‘epilepsy-related gene’. Genes that did not fall into the epilepsy or epilepsy-related categories were classified as not associated with epilepsy. Statistical analysis of enrichment for epilepsy-associated genes (comprising epilepsy and epilepsy-related genes) was performed using cumulative distribution function of hypergeometric distribution with the Hypergeometric P-value calculator.
GO and KEGG pathway analyses
GO for biological processes (GO: BP) and KEGG analysis was performed for the bipartite fly co-expression module (n=26) with all Drosophila genes used as the background, as well as for the human M30 cluster (n=320) with all human genes as the background, using g:Profiler (Raudvere et al., 2019). Terms were defined as significant if the Benjamini–Hochberg FDR was <0.05. A selection of significant GO: BP terms were visualized using a dot-heatmap generated using ggplot2 to display both FDR significance and enrichment (Wickham, 2016). The complete list of significantly enriched GO: BP and KEGG terms is provided in Table S5. Hypergeometric test statistics and overlap between significant GO terms in the fly co-expression module and human M30 cluster was determined in R using a population size of all potential GO: BP terms (27,047) and visualized using a Venn diagram generated using BioVenn (Hulsen et al., 2008).
Drosophila stocks and maintenance
Flies were cultured in a standard medium containing cornmeal, yeast, sugar and agar. The rearing conditions were maintained at a constant temperature of 25°C and 60% relative humidity, under a 12:12-h light-dark (LD) cycle. The Gal4/UAS system was used to study the effect of neuronal-specific knockdown of different genes (Brand and Perrimon, 1993). For this purpose, the pan-neuronal driver nSyb-Gal4 [Bloomington Drosophila Stock Center (BDSC) #51635] was crossed to UAS-RNAi lines. UAS-RNAi lines were obtained from the Vienna Drosophila Resource Center (VDRC) or BDSC and are summarized in Table S7. Further Drosophila stocks that were used in this study are as follows: the different forms of AMPK [UAS-AMPKαM (BDSC #32110), UAS-AMPKαK57A (BDSC #32112) and UAS-AMPKαT184D (BDSC #32110)]; the genetic background controls of the KK library (VDRC #60100), of the GD library (VDRC #60000), and of the TRiP libraries attP2 (BDSC #36303) and attP40 (BDSC #36304). To generate the experimental control animals, lines containing the corresponding genetic background to the RNAi constructs (VDRC #60000, VDRC #60100, BDSC #36303, BDSC #36304) were crossed to the driver line.
Mechanical induced seizure-like behavior
The mechanically induced seizure-like behavior assay was adapted from Kuebler and Tanouye (2000). At day of eclosion (day 0), five male flies per genotype were collected under CO_2_ anesthesia and housed in vials with standard food. Animals were transferred to fresh food on day 3. At 6 days old, flies were allowed to acclimate for 5 min in new, transparent vials and were sequentially mechanically stimulated in a vortex mixer at the maximum setting (2400 rpm) for 1 min, and videos were recorded after the stimuli. The number of flies that showed seizure-like behavior [either lying on their backs with uncontrolled wing flapping and/or leg twisting (Kuebler and Tanouye, 2000) within 5 s or paralysis for more than 5 s post-stimuli] was scored. The entire procedure was conducted under double-masked conditions to ensure unbiased scoring, and the genotype identities were only decoded post-analysis. Statistical analyses were performed using GraphPad Prism version 10.1.2 for Windows (GraphPad Software, San Diego, CA, USA). Because the dataset did not follow a Gaussian distribution, it was analyzed using Kruskal–Wallis test with Dunn's multiple comparisons test. Significance in the figures correspond to adjusted P-values, reflecting only those results meeting the corrected significance threshold. All data presented were derived from two (Fig. 3) or three (Fig. 6) independent experiments.
Heat induced seizure-like behavior
The heat induced seizure-like behavior assay was adapted from Fischer et al. (2016). Flies were reared at 25°C or 28°C as indicated. For quantifying heat induced seizure-like behavior, aliquots of five males per genotype were collected at day 0 post-eclosion under CO_2_ anesthesia, and housed in vials with standard food, with a transfer to fresh vials on day 3. At 6 days old, flies were acclimated for 5 min in new, transparent vials. Vials were individually submerged in a temperature-controlled water bath and adjusted to the target temperature with a tolerance of ±0.2°C. Flies in the KK and TRiP genetic backgrounds were immersed at a temperature of 37°C, whereas those in the GD background were exposed to 38°C, based on their different temperature sensitivity. The number of flies showing seizure-like behavior was recorded at 5-min intervals over a duration of 80 min. For testing the ability of AMPK overexpression under stringent conditions, flies were immersed into a 42°C water bath. The number of flies showing seizure-like behavior was recorded at 10 s intervals over a duration of 10 min. To calculate the time at which 50% of individuals showed seizures, a sigmoidal curve with a variable slope was applied to the raw data. This calculation of the time per genotype, referred to as logEC50, was performed using GraphPad Prism. This timepoint served as an indicator of seizure susceptibility and was utilized for subsequent statistical analyses. The values for the RNAi and overexpression lines were normalized against their respective background controls, owing to day-to-day variation. The entire procedure was conducted under double-masked conditions to ensure unbiased scoring, and the genotype identities were only decoded post-analysis. Statistical analyses were performed using GraphPad Prism. Because the dataset was normally distributed, it was analyzed using ordinary one-way ANOVA tests with Šídák's multiple comparisons test. Significance in the figures is denoted as adjusted P-values, reflecting only those results meeting the corrected significance threshold. All data presented were derived from two (Fig. 3) or three (Fig. 6) independent experiments.
Metabolic rate
The metabolic rate assay was adapted from Yatsenko et al. (2014). In brief, to measure CO_2_ production in Drosophila, aliquots of 25 males per genotype were collected after eclosion (day 0-1) under CO_2_ anesthesia, gently transferred into vials containing standard food and flipped to new vials on day 3. For behavioral testing, 6- to 7-day old flies were transferred without CO_2_ anesthesia to homemade respirometers in groups of five and allowed to acclimate for 15 min. The respirometers, crafted using pipette tips and capillary micropipettes with soda lime as a CO_2_ absorbent, were hung vertically to immerse the tip into a Bromophenol Blue solution, and the chambers were sealed. Following a 75-min acclimation period within the sealed chamber, photos were taken documenting the liquid level at the start and end of a 1-h period. Photos were analyzed using Fiji 1.53 to measure the liquid's travel distance. The resulting data were presented as microliters of CO_2_ produced per fly. Statistical analyses were performed using GraphPad Prism. Datasets following a Gaussian distribution were analyzed with two-tailed unpaired t-tests. Datasets that did not follow a Gaussian distribution were analyzed using Mann–Whitney tests. All data presented were derived from three independent experiments.
Novelty of identified links between genes with altered seizure susceptibility, energy metabolism and AMPK activity
To provide an overview on the literature implicating genes of the evolutionarily conserved and co-expressed 26-gene module with altered seizure susceptibility in energy metabolism and/or AMPK activity, a comprehensive search of the PubMed database was conducted for each gene using terms ‘metabolic rate’, ‘AMPK’, ‘CO_2_ production’, ‘O_2_ consumption’, ‘energy metabolism’ and ‘glycolytic flux’. Relevant matches are listed in Table S8.
Western blotting
To detect AMPK phosphorylation levels aliquots of 25 males per genotype were collected after eclosion (day 0-1) under CO_2_ anesthesia. They were gently transferred into vials containing standard food and subsequently flipped to new vials on day 3. After 6 days, flies were CO_2_ anesthetized, transferred to Eppendorf tubes and snap frozen in liquid nitrogen. Heads were separated from bodies by flicking the tubes and were collected with a brush at 4°C. Twenty heads per sample were homogenized in 80 μl RIPA buffer with phosphatase inhibitors. Samples were mixed with a 1:1 volume of NuPAGE LDS Sample Buffer 4× (NP0007, Life Technologies, Inchinnan, UK) with DL-Dithiothreitol solution (43816-10ML, Sigma-Aldrich, Saint Louis, MO, USA), and boiled for 10 min. Then, 12 μl of each sample was loaded onto a 15-well NuPAGE 4-12% Bis-Tris gel (NP0323BOX, Life Technologies) and electrophoresed for 15 min at 80 V, followed by 120 min at 120 V. Proteins were transferred to nitrocellulose membranes (Trans-Blot Turbo Transfer Pack, 1704158, Bio-Rad, Hercules, CA, USA) using the Trans-Blot Turbo Transfer System (1704150, Bio-Rad) at 25 V for 7 min. Membranes were blocked for 1 h in 10% bovine serum albumin (BSA; A8806-1G, Sigma-Aldrich), and then incubated in primary antibody diluted in a 1:1 volume of 10% BSA and Tris-buffered saline buffer with 0.2% Tween 20 (TBST) overnight at 4°C. The antibody against the phosphorylated form of α-AMPK (2535S, Cell Signaling Technology, Danvers, MA, USA; RRID:AB_331250) was diluted 1:500; anti-β-Tubulin (E7, Developmental Studies Hybridoma Bank, Iowa City, IA, USA; RRID:AB_528499) was diluted 1:6000. The membranes were then washed three times for 10 min each in TBST. Membranes were incubated in a goat anti-mouse IgG (H+L) secondary antibody (dilution 1:5000; A-21057, Thermo Fisher Scientific, Waltham, MA, USA; RRID: AB_2535723) and a goat anti-rabbit secondary antibody (dilution 1:5000; 926-32211, Li-cor, Lincoln, NE, USA; RRID:AB_621843), diluted in a 1:1 volume of 10% BSA and TBST buffer at room temperature, for 2 h in the dark. Three 10-min washes with TBST buffer were done, and the membranes were immersed in Tris-buffered saline in the dark. Membranes were imaged on an Odyssey Infrared Scanner (Odyssey^®^ DLx Imaging System, 9142-00, Li-cor). Quantification was conducted using Image Studio software (version 4.0.21). All direct comparisons were only done on samples run on the same gel, and the Odyssey Integrated Intensity was used to estimate the relative strength of signals across different blots. Each experimental group comprised three biological replicates from two independent experiments (six samples), and each sample was run once on a gel for quantification. Statistical analyses were performed using GraphPad Prism. Datasets following a Gaussian distribution were analyzed with two-tailed unpaired t-tests. All data presented were derived from two independent experiments.
Supplementary Material
10.1242/dmm.052307_sup1Supplementary information
Table S1. The Drosophila orthologs of 320 human genes in M30 cluster
Table S2. 287 selected Drosophila orthologs of M30 cluster
Table S3. 142 Drosophila genes for clustering analysis
Table S4. Hierarchically clustered coexpression matrix of 142 x 142 genes with pairwise Spearman correlation values
Table S5. Complete KEGG pathway and GO biological process terms list
Table S6. Epilepsy-associated gene list of M30 network
Table S7. Crosses and genotypes in co-expression gene modules and associated lethality in pan-neuronal
Table S8. Novel versus established links between genes of the evolutionarily conserved and co-expressed 26-gene module with altered seizure susceptibility, energy metabolism and AMPK activity. Abbreviations: AMPK - Adenosine monophosphate-activated protein kinase.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Almeida, A. M., Murakami, Y., Baker, A., Maeda, Y., Roberts, I. A. G., Kinoshita, T., Layton, D. M. and Karadimitris, A. (2007). Targeted therapy for inherited GPI deficiency. N. Engl. J. Med. 356, 1641-1647. 10.1056/NEJ Moa 06336917442906 · doi ↗ · pubmed ↗
- 2Alnaaim, S. A., Al-Kuraishy, H. M., Al-Gareeb, A. I., Ali, N. H., Alexiou, A., Papadakis, M., Saad, H. M. and Batiha, G. E. S. (2023). New insights on the potential anti-epileptic effect of metformin: mechanistic pathway. J. Cell. Mol. Med. 27, 3953-3965. 10.1111/jcmm.1796537737447 PMC 10747420 · doi ↗ · pubmed ↗
- 3Andreeva, A. V., Kutuzov, M. A. and Voyno-Yasenetskaya, T. A. (2006). A ubiquitous membrane fusion protein alpha SNAP: a potential therapeutic target for cancer, diabetes and neurological disorders? Expert Opin Ther. Targets 10, 723-733. 10.1517/14728222.10.5.72316981829 · doi ↗ · pubmed ↗
- 4Ashraf, A., Fan, Z., Brooks, D. J. and Edison, P. (2015). Cortical hypermetabolism in MCI subjects: a compensatory mechanism? Eur. J. Nucl. Med. Mol. Imaging 42, 447-458. 10.1007/s 00259-014-2919-z 25267349 · doi ↗ · pubmed ↗
- 5Baker, K., Gordon, S. L., Melland, H., Bumbak, F., Scott, D. J., Jiang, T. J., Owen, D., Turner, B. J., Boyd, S. G., Rossi, M. et al. (2018). SYT 1-associated neurodevelopmental disorder: a case series. Brain 141, 2576-2591. 10.1093/brain/awy 20930107533 PMC 6113648 · doi ↗ · pubmed ↗
- 6Bartolini, E., Ferrari, A. R., Fiori, S. and Della Vecchia, S. (2023). Glycaemic imbalances in seizures and epilepsy of paediatric age: a literature review. J. Clin. Med. 12, 2580. 10.3390/jcm 1207258037048663 PMC 10095009 · doi ↗ · pubmed ↗
- 7Bowser, D. N. and Khakh, B. S. (2004). ATP excites interneurons and astrocytes to increase synaptic inhibition in neuronal networks. J. Neurosci. 24, 8606-8620. 10.1523/JNEUROSCI.2660-04.200415456834 PMC 6729897 · doi ↗ · pubmed ↗
- 8Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118, 401-415. 10.1242/dev.118.2.4018223268 · doi ↗ · pubmed ↗