Integrative revision of the Palaearctic owlfly genus Deleproctophylla Lefèbvre (Neuroptera, Myrmeleontidae, Ascalaphinae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
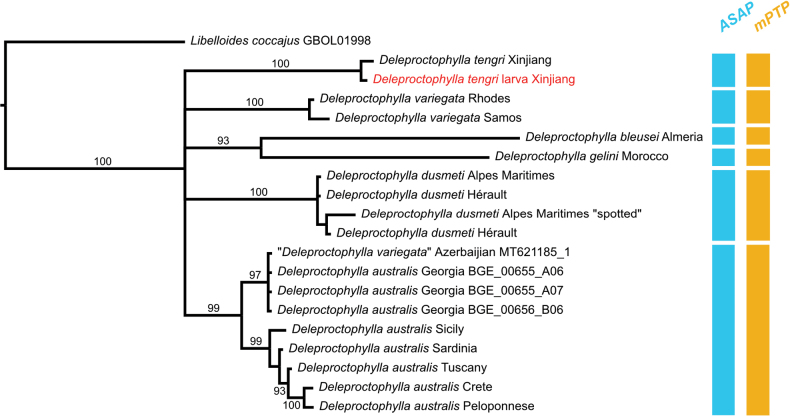 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6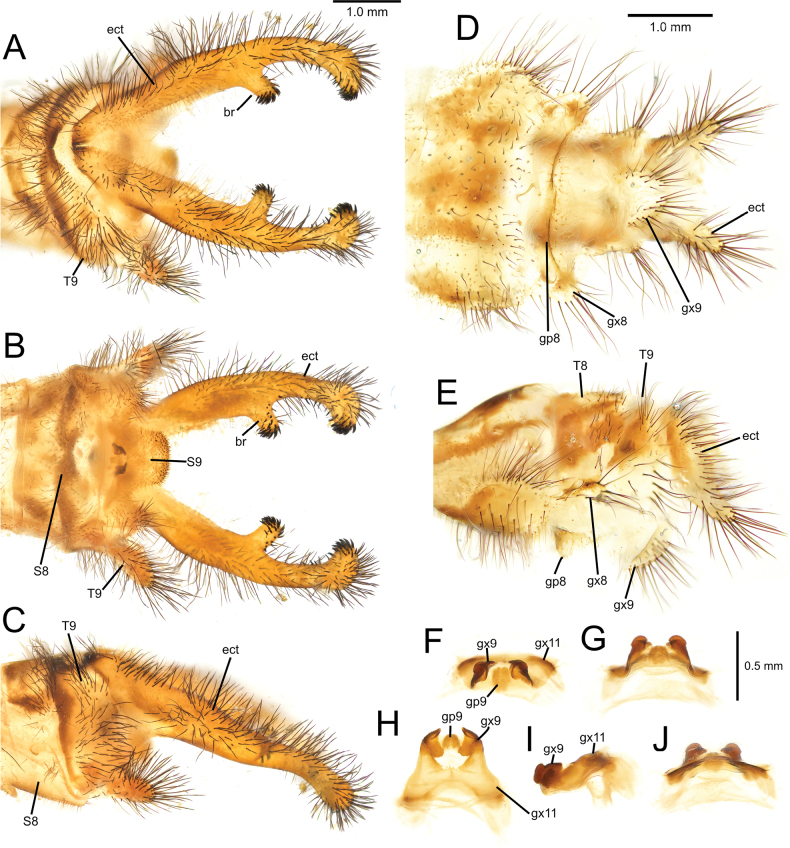 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11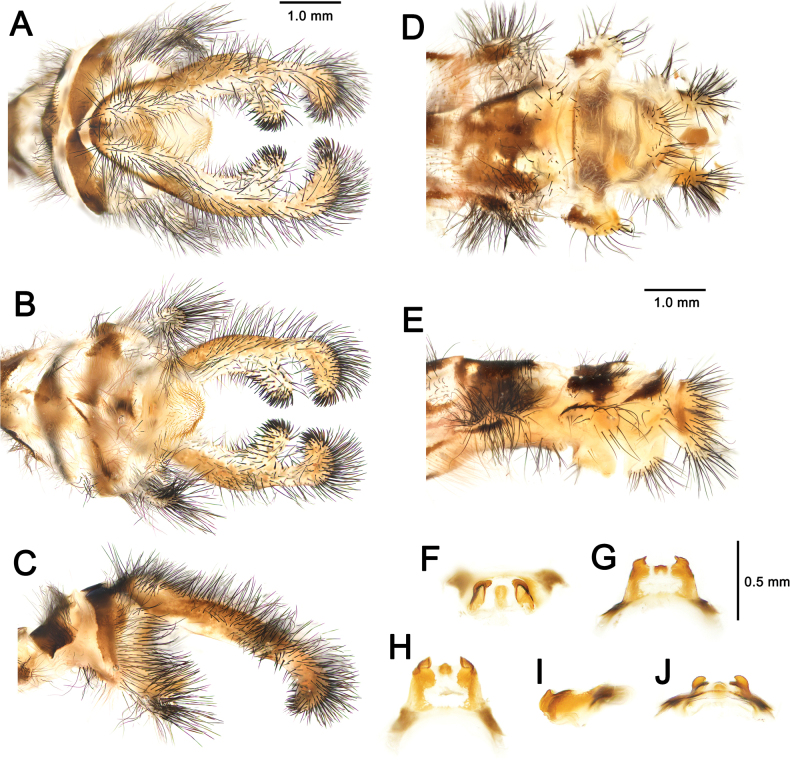 Figure 12
Figure 12 Figure 13
Figure 13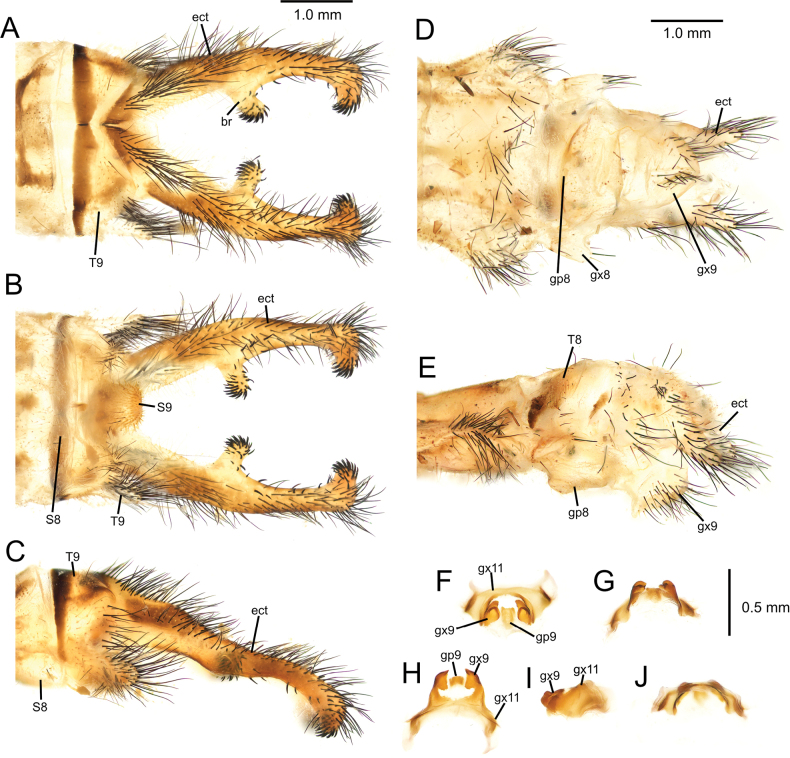 Figure 14
Figure 14 Figure 15
Figure 15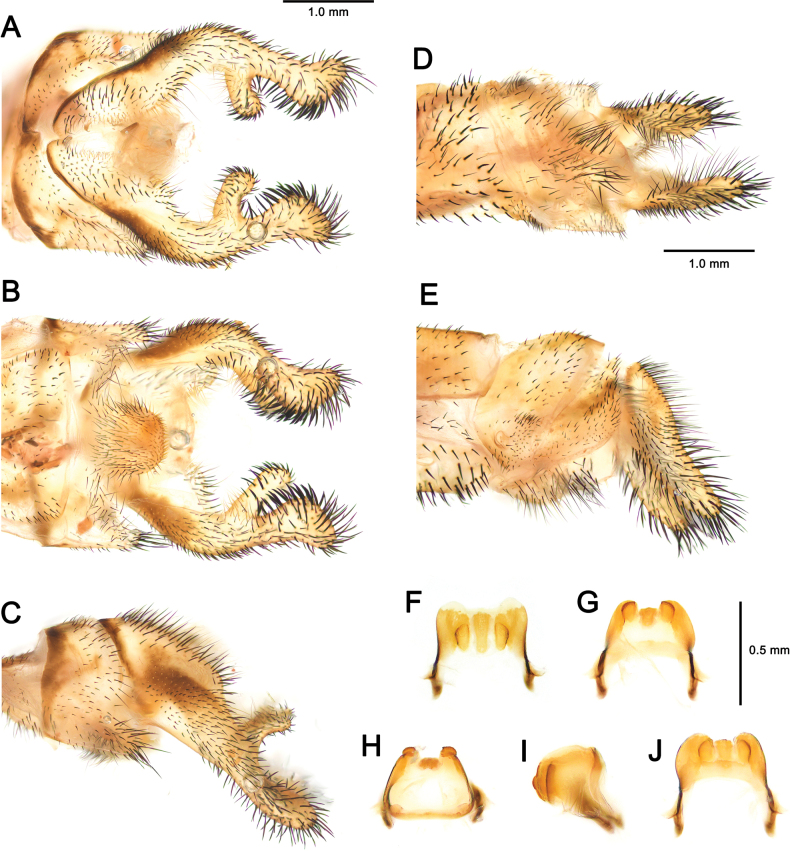 Figure 16
Figure 16 Figure 17
Figure 17 Figure 18
Figure 18 Figure 19
Figure 19 Figure 20
Figure 20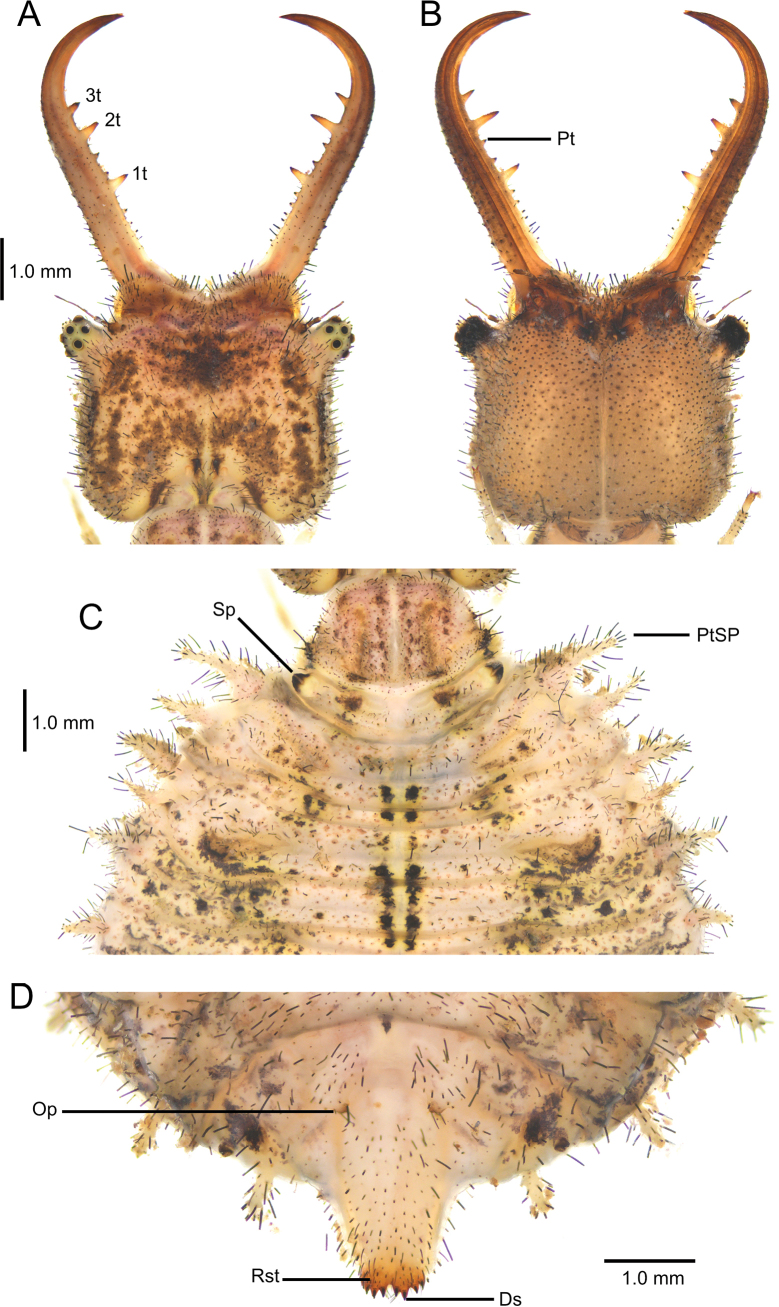 Figure 21
Figure 21 Figure 22
Figure 22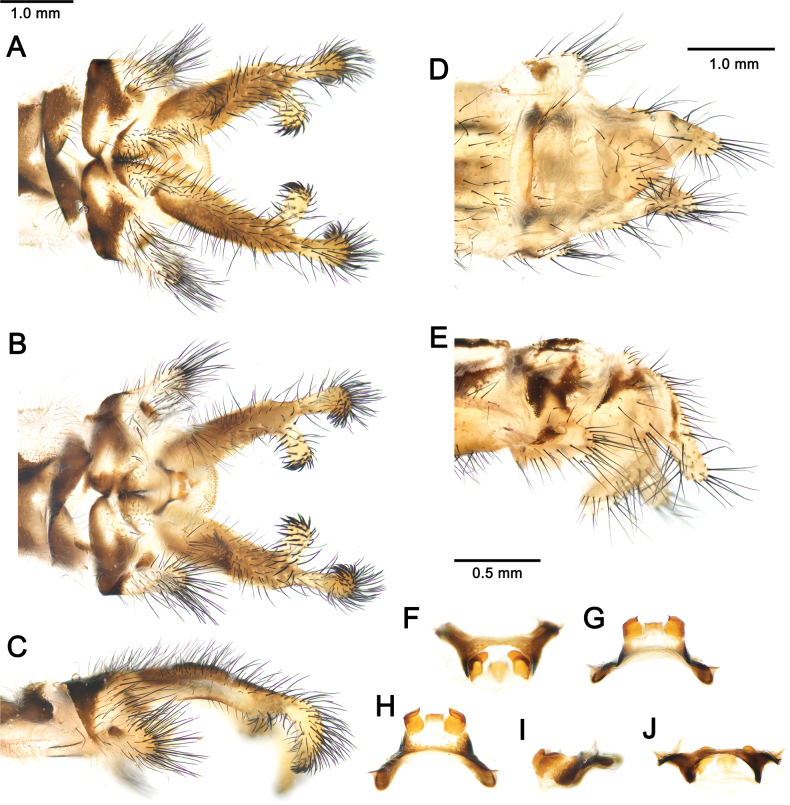 Figure 23
Figure 23 Figure 24
Figure 24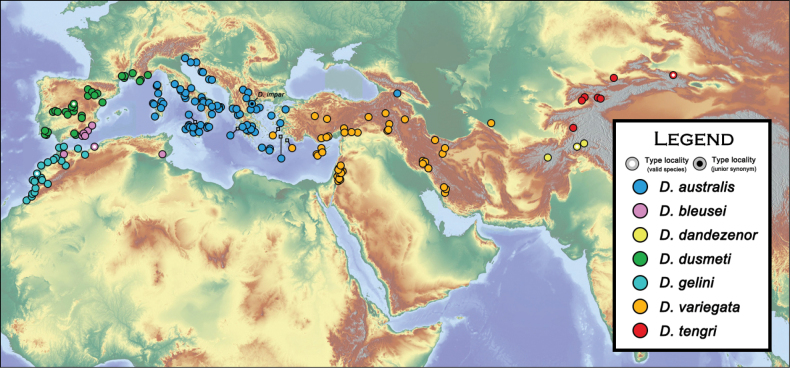 Figure 25
Figure 25| Accession number | Species | Provenance | Reference |
|---|---|---|---|
|
| Germany, Bavaria | (Morinière et al. 2014) | |
|
| Spain, Almería, Cabo de Gata | This study | |
|
| China, Xinjiang, Shihezi | This study | |
|
| China, Xinjiang, Shihezi | This study | |
|
| Greece, Rhodes, Mount Profitis Ilias | This study | |
|
| Greece, Samos | This study | |
|
| France, Alpes Maritimes, Villeneuve Loubet | This study | |
|
| France, Alpes Maritimes, Villeneuve Loubet | This study | |
|
| France, Hérault, St. Paul et Valmalle | This study | |
|
| France, Hérault, St. Paul et Valmalle | This study | |
|
| Italy, Toscana, Nomadelfia | This study | |
|
| Italy, Sardegna, Alghero | This study | |
|
| Italy, Sicily, Agrigento | This study | |
|
| Greece, Crete, Hersonissos | This study | |
|
| Greece, Peloponnese | This study | |
|
| Morocco | This study | |
|
| “ | Azerbaijan | ( |
| BoldSystems: BGE_00655_A06 | Georgia, Tibilisi | BoldSystems | |
| BoldSystems: BGE_00655_A07 | Georgia, Tibilisi | BoldSystems | |
| BoldSystems: BGE_00656_B06 | Georgia, Tibilisi | BoldSystems |
| 1 | Vertex nearly as long as wide (Fig. |
|
| – | Vertex longer than wide (Fig. |
|
| 2 | Hindwing with indistinct marking, as a brown shade bordering crossveins below pterostigma, not extending along |
|
| – | Hindwing with distinct marking, or as shade extending along |
|
| 3 | Metafemur without brown marking (Fig. |
|
| – | Metafemur with brown marking (Fig. |
|
| 4 | Forewing with round apex (Figs |
|
| – | Forewing with acute apex (Figs | |
| 5 | Hindwing with marking extending as band for whole breadth of wing (Figs | |
| – | Hindwing with marking not extending beyond radial field (Fig. |
|
| 6 | Hindwing elongate, hind margin not pronounced (Fig. |
|
| – | Hindwing subtriangular, hind margin angled (Fig. |
|
| 1 | Dorsal side of head capsule dark brown with paler markings, ventral side dark brown with three parallel longitudinal pale stripes (Fig. |
|
| – | Dorsal side of head capsule pale brown mottled dark brown, ventral side uniformly pale brown (Fig. |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFossil Insects in Amber · Hymenoptera taxonomy and phylogeny · Diptera species taxonomy and behavior
Introduction
The lacewing family Myrmeleontidae can be roughly divided into two distinct morphotypes: antlions (i.e., members of subfamilies Dendroleontinae, Nemoleontinae, Myrmeleontinae, and part of Ascalaphinae), and owlflies (i.e., part of Ascalaphinae), which were traditionally considered separate families (Badano et al. 2017; Machado et al. 2019). Phylogenomic and transcriptomic analyses have confirmed that owlflies (formerly Ascalaphidae) are deeply nested within Myrmeleontidae, supporting a revision of their internal classification and subfamily-level divisions (Winterton et al. 2018; Machado et al. 2019; Vasilikopoulos et al. 2020; Lai et al. 2025). Owlflies (Ascalaphinae) are highly distinctive insects with long-clubbed antennae, large compound eyes, and elongated wings. They are powerful fliers, actively preying on small insects in flight, usually at dusk or night (Tjeder 1992; Oswald and Machado 2018). However, a few, like the two Palaearctic genera—Libelloides Schäffer, 1763, and Deleproctophylla Lefèbvre, 1842—are one of the few exceptions, being diurnal and most active during the warmest and sunniest hours of the day (H. Aspöck et al. 1980; Tjeder 1992; Michel and Kral 2008). These genera are not only largely sympatric, but often syntopic, particularly in Mediterranean Europe (H. Aspöck et al. 1980). Despite their overlapping distributions, Libelloides and Deleproctophylla exhibit strikingly different adult morphologies: the former is colorful and conspicuous (with pale yellow or white and black wings), while the latter is buff-colored, with mostly hyaline wings, and well-camouflaged among dry grasses (H. Aspöck et al. 1980). These two genera share very similar male genital structures (H. Aspöck et al. 1980), and their larvae differ only in minor features (Badano and Pantaleoni 2014), despite their phylogenetic affinities has never been tested. The genus Deleproctophylla currently comprises five species, i.e., D. australis (Fabricius, 1787), D. variegata (Klug in Ehrenberg, 1834), D. dusmeti (Navás, 1914), D. gelini (Navás, 1919) and D. bleusei Kimmins, 1949, and is widely distributed from the Mediterranean basin to Central Asia (H. Aspöck et al. 2001). The wing pattern has been traditionally used to distinguish the species of this genus, despite being highly variable, often leading to confusion (Monserrat et al. 2014). This trait has also proven misleading for populations in southern France and Anatolia, where individuals were assigned to D. australis (Colombo et al. 2012) based solely on the presence or absence of a small brown spot below the pterostigma in the forewing.
The entangled taxonomic history of Deleproctophylla
Despite Deleproctophylla being one of the few ascalaphid genera present in Europe, its taxonomic history is marked by a remarkable series of misinterpretations, confusions, and overlooked species. Fabricius (1787) described Ascalaphus australis, the first species of the genus. Later, Klug in Ehrenberg (1834) described Ascalaphus variegatus from the Middle East. Lefèbvre (1842) established the genus name Deleproctophylla, which Rambur (1842) shortly thereafter emended to Theleproctophylla. Hagen (1860) erroneously synonymised Ascalaphus australis and Ascalaphus variegatus with Ascalaphus barbarus (Linnaeus, 1767). This led to the perception of a single Theleproctophylla species, Theleproctophylla barbara, a view perpetuated by McLachlan (1873) and van der Weele (1904). Van der Weele (1908) reinstated the original spelling Deleproctophylla, recognised that A. barbarus as belonging to a distinct genus, and validated D. australis as a distinct species and the genus type species. However, van der Weele (1908) continued to classify Iberian and French populations as D. variegata, primarily based on the absence of a forewing spot, thus considering D. variegata to have a widespread distribution. Notably, most taxonomic issues within Deleproctophylla center on the French and Iberian species. Navás (1914) further refined the taxonomy by distinguishing the Spanish species from D. variegata of Anatolia, describing D. dusmeti. However, he overlooked the presence of two distinct Deleproctophylla species in Spain, mistaking the second for immature D. dusmeti. Later, Kimmins (1949) described D. bleusei from North Africa, but the presence of this species in southern Spain was only recognized much later by Monserrat et al. (2014). H. Aspöck et al. (1980), in their influential work on European Neuroptera, also adhered to the single-species concept for the Iberian Peninsula, leading to their description of D. dusmeti incorporating diagnostic characters of D. bleusei. H. Aspöck et al. (1980) also noted a second morph in Anatolia alongside D. variegata, which they termed the ‘dusmeti-like morph’ (‘dusmeti-ähnliches Phänon’). The status of these Anatolian populations remained unclear, and they were variously cited, sometimes even as D. dusmeti (Dobosz and Ábrahám 2007). Similarly, records of D. australis from southern France, often sympatric with D. dusmeti, remained enigmatic (Colombo et al. 2012).
Remarkably, new field sampling and the study of museum collections have revealed two new species, D. dandizenor sp. nov. and D. tengri sp. nov., considerably expanding the known range of the genus into Asia and reporting it from Afghanistan and China for the first time. The main aims of the present study are: i) to revise the genus Deleproctophylla by integrating morphological and molecular data; ii) to test species boundaries using COI-based species delimitation algorithms; and iii) to describe the two new species from Central Asia.
Materials and methods
Examined collections
The examined specimens are preserved in the following collections.
CAU Entomological Museum, China Agricultural University, Beijing, China
DBC Davide Badano Collection, University of Siena, Siena, Italy
HUAC Collection of Horst and Ulrike Aspöck, Wien (Vienna), Austria
MDC Musée des Confluences, Lyon, France
MfN Museum für Naturkunde, Berlin, Germany
MNHN Musée National d’Histoire Naturelle, Paris, France
MSNG Natural History Museum “Giacomo Doria”, Genova, Italy
NHMD Natural History Museum of Denmark, Copenhagen, Denmark
NHMW Naturhistorisches Museum, Wien (Vienna), Austria
NMPC National Museum of the Czech Republic, Prague, Czech Republic
RMNH Naturalis Biodiversity Center, Rijksmuseum van Natuurlijke Historie, Leiden, the Netherlands
SCM Rippl-Rónai Museum, Kaposvár, Hungary
USMB Upper Silesian Museum, Bytom, Poland
VM Victor Monserrat collection, Madrid, Spain
ZCAU Yuchen Zheng collection, China Agricultural University, Beijing, China
Locality information was transcribed from collection labels, maintaining the original format. To map the distribution of historical specimens or for those without coordinates, we assigned an approximate point corresponding to the centroid of the named locality on the map.
Morphological examination
Genitalia were prepared by clearing the abdominal apex in a 20% KOH solution at 100 °C for seven minutes, followed by rinsing with distilled water. The cleared apex was then transferred to glycerin for further examination.
Habitus photographs were taken using a Nikon® D850 digital camera equipped with an AF-S Micro Nikkor 105 mm f/2.8G ED lens and a Zeiss® Axio Zoom.V16 stereo microscope. The head, thorax, and legs were photographed using a Nikon® D850 digital camera with a Laowa® 25mm f/2.8 2.5–5.0× Ultra Macro lens. Images of the pretarsal claws and genitalia were captured with a Leica® DM2000 microscope fitted with a Nikon® D850 digital camera.
The classification system of Myrmeleontidae follows Machado et al. (2019). Wing venation terminology primarily follows Breitkreuz et al. (2017). Morphological terminology follows Tjeder (1992). Genital terminology is based on U. Aspöck and H. Aspöck (2008), due to the controversy regarding the interpretation of genital segments 10–11 in Myrmeleontidae (Marquez-López et al. 2024).
Morphological abbreviations: C costa, Sc subcosta, RA radius anterior, RP radius posterior, MA media anterior, MP media posterior, CuA cubitus anterior, CuP cubitus posterior, A anal veins, ect ectoproct, br branch of ectoproct, T8 tergite 8, gp8 gonapophysis 8, gx8 gonocoxite 8, T9 tergite 9, S9 sternite 9, gx9 gonocoxite 9, gp9 gonoapophysis 9, gx11 gonocoxite 11.
DNA extraction and amplification
Genomic DNA was extracted from the leg of selected specimens, preserved in ethanol (Table 1). Due to the state of preservation of the type specimens, it was not feasible to extract DNA from D. dandizenor sp. nov. Mitochondrial DNA fragment Cytochrome Oxidase subunit I was amplified with the forward primer LCO1490 (5’- GGTCAACAAATCATAAAGATATTGG -3’) and the reverse primer HCO2198 (5’- TAAACTTCAGGGTGACCAAAAAATCA -3’) (Folmer et al. 1994). PCRs were executed in 25 μl reaction volume with 2.5 μl of DNA from each sample, 1.25 μl of both forward and reverse primers (10 μM), 2.5 μl of MgCl 2 (25 mM), 2.5 μl of deoxynucleotides (dNTPs, 10 mM), 5 μl of Green GoTaq Flexi Buffer (Promega, US), 0.125 μl of GoTaq Flexi DNA polymerase (Promega, US) [5 u/μl], and 9.875 μl of sterile ddH2O. Amplifications were obtained in a T100 Thermal cycler (BioRad, US) Thermal Cycler, using the following conditions: (i) 95 °C for 5 min; (ii) 35 cycles of 95 °C for 1 min, 50 °C for 1 min, 72 °C for 90 s; (iii) 72 °C for 7 min. The kit Wizard ® SV Gel and PCR Clean-Up System (Promega, US) were employed or PCR products purification, and their DNA concentration was assessed through NanoDrop One (Thermo Scientific, US) device. The quality of PCR results and purifications was checked through 1% agarose gel. Samples were sequenced with the same primers and applying Sanger techniques at the core facility of BMR Genomics (Padua, IT) run on by automated DNA sequencer.
The obtained COI sequence was edited and checked with MEGA XI. The barcode sequence was analyzed through the integrated bioinformatics platform Barcode of Life Data (BOLD) System to test the morphology-based species identification. COI sequences obtained from GenBank were aligned using the ClustalW program implemented in MEGA XI (Table 1).
Phylogenetic and species delimitation analyses
A Bayesian Inference (BI) analysis was performed to reconstruct the phylogenetic relationships among taxa based on aligned COI sequences using MrBayes 3.2.7. Libelloides coccajus (Denis & Schiffermüller, 1775) was selected as an outgroup, based on the potential affinities of these two genera. The analysis was run enforcing the invgamma model. Four Markov chain Monte Carlo (MCMC) chains, of which one was cold and three heated, were run for 1,000,000 generations, setting a burn-in fraction of 50% and sampling the chains every 1000 generations. The convergence of independent runs was assessed through the average standard deviation of split frequencies (<0.01) and potential scale reduction factors (approaching 1).
To test species boundaries among morphologically similar taxa and to support the description of the new species, two single-locus species delimitation approaches were applied: Assemble Species by Automatic Partitioning (ASAP) (Puillandre et al. 2021) and Multi-rate Poisson Tree Processes (mPTP) (Kapli et al. 2017). ASAP is a distance-based method, calculating pairwise genetic distances and assigning a score to different partitions, selecting the most supported one. The analysis was performed on the ASAP web server, using Kimura (K80) transition/transversion (ts/tv) ratio as the nucleotide substitution model. Instead, mPTP is tree-based method, which infers species boundaries by performing Markov Chain Monte Carlo (MCMC) sampling on the supplied phylogenetic tree. The analysis was conducted on the mPTP web server using default parameters with the BI tree as input.
Results
Phylogenetic analyses and species delimitation
The BI analysis of the COI sequences (Table 1) strongly supported the monophyly of the six analyzed species of Deleproctophylla (Bayesian Posterior Probabilities: ≥ 99) (Fig. 1). The results of both species delimitation analyses based on the COI mitochondrial marker were congruent, identifying the same species across the sampled specimens. The best ASAP partition (ASAP score: 1.00, P-value: 2.994012e-01, W: 1) and the mPTP analysis consistently recovered six species within the sample (Fig. 1).
Bayesian phylogenetic tree of COI sequences of the analyzed specimens of Deleproctophylla. Bayesian posterior probabilities > 90 are shown above branches. Vertical bars indicate species delimitation analyses results obtained with ASAP and mPTP.
The phylogenetic analysis strongly supported that the examined larva collected in Xinjiang belongs to D. tengri sp. nov. (Fig. 1). Moreover, both ASAP and mPTP delimited D. tengri as a well distinct taxon.
Both methods confirmed that the population of D. dusmeti with atypical forewing markings belong to this species. Similarly, the analyses supported the monophyly of all examined D. australis specimens, including those collected from Mediterranean islands and the Caucasus. Notably, both methods identified a GenBank sequence from Azerbaijan, originally assigned to D. variegata, as deeply nested within D. australis, suggesting a misidentification. However, the Bayesian analysis recovered two well distinct sister clades within D. australis, one including all the European specimens and the other the Asian specimens.
The species delimitation analyses also supported the distinctiveness of the two species for which only singletons were available, namely D. bleusei and D. variegata.
Taxonomy
Family Myrmeleontidae Latreille, 1807
Subfamily Ascalaphinae Lefèbvre, 1842
Tribe Ascalaphini Lefèbvre, 1842
Deleproctophylla
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
Lefèbvre, 1842
BEA9AA02-FEDA-5955-B349-5403B133D1E3
Figs 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24
Deleproctophylla Lefèbvre, 1842: 6 (emended to “Theleproctophylla” by Rambur, 1842: 351). Type species: Ascalaphus australis Fabricius, 1893, by subsequent designation by van der Weele (1908).
Diagnosis.
Adult. Head breadth at widest point, including compound eyes, slightly larger to width of mesothorax at wing base. Compound eye with deep median furrow, lateral margins slightly indented at furrow. Antenna not reaching pterostigma; club pyriform, wide. Forewing elongate; hindwing broadest at RP fork. Forewing membrane largely hyaline, with brown markings in a few species; hindwing membrane largely hyaline with brown markings or infuscations in apical half. Abdomen without pleuritocavae.
Male genitalia. Ectoproct with prominent “pincer-like” ventral projection, with bifurcating cylindrical branch in distal half, apex bent downward and inward; branch and apex with robust, spiniform setae. Tergite 9 sub-rectangular in lateral view, with prominent ventral thumb-shaped projection. Sternite 9 with caudal paired lateral lobes and a median, prominent sub-rectangular lobe; setae on median lobe spine-like. Complex of gx9+11 proportionally small in comparison to tip of abdomen; gx11 arch-shaped, projecting caudally and slightly curved downward in lateral view; gp11 forming a furrow, with short setae; gx9 widely set apart, with a dorso-apical, upward fold, resembling a smooth hook in lateral view.
Female genitalia. Ectoproct elongate, with short ventral projection. Tergite 9 subrectangular. Gx9 wider than long. Tergite 8 subrectangular. Gx8 distinct, not fused and widely set apart, wider than long, slightly prominent. Gp8 caudally prominent, appearing subtriangular in lateral view (except for D. gelini).
Larva. Head capsule not dilated posteriorly; ocular tubercle cylindrical with small apical protuberance; mandible with three teeth, median tooth largest and positioned closer to apical than to basal tooth; meso- and metathorax each with two pairs of sub-equal cylindrical scolus-like processus; anterior abdominal spiracle lateral; dorsal abdominal series of spiracles composed of eight cylindrical scolus-like processus, ventral series composed of two anterior scolus-like processus and six tubercle-like processus; sternite 8 with short odontoid processus; sternite 9 with short rastra with four digging setae; body dolichasters elongated, stick-shaped.
Comparative notes.
Potential synapomorphies of Deleproctophylla include: i) male ectoproct with caudal projection and median bifurcating branch, ii) male gx9 widely separated from each other and from gp11, with apical fold; iii) female ectoproct with caudal projection. Deleproctophylla is one of the few genera of Ascalaphini characterized by prominent male ectoproct with median branch, such as Bubopsis McLachlan, 1898, and Phalascusa Kolbe, 1897 (Tjeder 1992). However, in Deleproctophylla species, the ectoproct branch develops inward or dorsally (i.e., D. gelini), while in the sympatric species of Bubopsis, the branch extends basally and ventrally. These traits, combined with the characteristic pigmentation pattern of Deleproctophylla, allow easy identification from other ascalaphid genera with elongated ectoproct. In addition, to the obvious coloration differences, Deleproctophylla is immediately set apart from Libelloides in the shape of the male ectoproct, which is proportionally shorter, simpler in shape and lacking branch in the latter. However, these two genera share a similar conformation of the gx9+11 complex, especially in the shape of gx9. The larvae of these two genera are also very similar in body shape, tooth arrangement, and the position, development, and shape of thoracic and abdominal series of scolus-like processus. The larvae of Deleproctophylla can be distinguished by those of Libelloides in the head capsule, which is not dilated posteriorly and the dolichasters covering the body, which are mostly stick-shaped and do not expand apically into a wide rosette (Badano and Pantaleoni 2014).
Remarks.
One of the most unusual features of Deleproctophylla is the presence of the so-called phylla, large, paired, membranous, leaf-shaped structures protruding from the female abdomen (Figs 2F, 3F) (H. Aspöck et al. 1980; Tjeder 1992). The genus name derives from this trait. The phylla consist of the male spermatophore, which remains attached to the female genital opening after mating but is shed when she lays eggs. The evolutionary significance of this prominent structure remains unclear, but it may serve to reduce competition between males by preventing remating with an already fertilized female. In a few other ascalaphid genera, such as Bubopsis McLachlan, females also retain the spermatophore after mating, but in none of them are these structures as large and striking as in Deleproctophylla.
Habitus of Deleproctophylla spp. A, C, E, G. Male; B, D, F, H. Female; A, B. D. australis (Fabricius, 1787), Grosseto (Italy); C, D. D. bleusei Kimmins, 1949, Fes (Morocco); E, F. D. dandizenor Badano, Zheng, U. Aspöck & Dobosz, sp. nov., male holotype and female paratype, Kunar (Afghanistan); G, H. D. dusmeti (Navás, 1914), Saint-Paul-et-Valmalle (France).
Habitus of Deleproctophylla spp. A, C, E. Male; B, D, F. Female; A, B. D. gelini (Navás, 1919), Guigou (Morocco); C, D. D. tengri Zheng, Badano, H. Aspöck & Liu, sp. nov., Xinjiang (China); E, F. D. variegata (Klug, 1834), Cyprus.
Key to the species of Deleproctophylla (adults)
**: **
Key to the known larvae of Deleproctophylla
**: **
Deleproctophylla
australis
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
(Fabricius, 1787)
EE4BE285-C0D1-5529-BB4B-C86382DC91A5
Figs 2A, 2B, 4A–D, 5A, 5B, 6–8
Ascalaphus australis Fabricius, 1787: 250 (type locality: “Europa australiore” = southern Europe; holotype in NHMD). Gmelin 1788: 2645 (Myrmeleon). Villers 1789: 62 (Myrmeleon). Olivier 1790: 245 (Ascalaphus). Fabricius 1793: 96 (Ascalaphus). Turton 1800: 405 (Myrmeleon). Angelini 1827: 468 (Ascalaphus). Lefèbvre 1842: 6 (Deleproctophylla). Rambur 1842: 351 (Theleproctophylla). Schneider 1845: 341 (Theleproctophylla). Walker 1853: 416 (Ascalaphus). Hagen 1860: 53 (Theleproctophylla). Stein 1863: 420 (Ascalaphus). Hagen 1866: 402 (Deleproctophylla). Costa 1866: 14 (Theleproctophylla). McLachlan 1873: 260 (Theleproctophylla). Navás 1905: 547 (Theleproctophylla). Navás 1911: 92 (Theleproctophylla). Navás 1912a: 124 (Theleproctophylla). Navás 1912b: 34 (Theleproctophylla). Lacroix 1913: 100 (Theleproctophylla). Kimmins 1930: 188 (Theleproctophylla). Mosely 1932: 182 (Theleproctophylla). Táborský 1936: 164 (Deleproctophylla). Capra 1936: 213 (Theleproctophylla). Capra 1937: 58 (Theleproctophylla). Zimsen 1964: 612 (Ascalaphus). Zelený 1964: 326 (Theleproctophylla). Popov 1977: 276 (Deleproctophylla). H. Aspöck et al. 1980: 315 (Deleproctophylla). Devetak 1992: 111 (Deleproctophylla). Zakharenko and Krivokhatsky 1993: 73 (Deleproctophylla). Bernardi et al. 1995: 11 (Deleproctophylla). Devetak 1995: 194 (Deleproctophylla). Letardi and Pantaleoni 1996: 299 (Deleproctophylla). H. Aspöck and Hölzel 1996: 78 (Deleproctophylla). Pantaleoni and Letardi 1998: 40 (Deleproctophylla). H. Aspöck et al. 2001: 301 (Deleproctophylla). Whittington 2002: 377 (Deleproctophylla). Popov 2004: 231 (Deleproctophylla). Canbulat 2007: 36 (Deleproctophylla). Dobosz and Ábrahám 2007: 24 (Deleproctophylla). Devetak and Janžekovič 2012: 183 (Deleproctophylla). Colombo et al. 2012: 125 (Deleproctophylla). Badano and Pantaleoni 2014: 301 (larva, Deleproctophylla). Letardi 2016: 194 (Deleproctophylla). Aistleitner 2019: 123 (Deleproctophylla). Jones 2019: 3 (Deleproctophylla). Poggi 2024: 9 (Deleproctophylla).Theleproctophylla australis var. impar Navás, 1923: 154 (type locality: Greece: Thessaloniki; holotype in MNHN but lost at present).
Examined material.
• Croatia: Dalmatien, Hvar, 17.VII.1962, R. Löberbauer, 1 ♀ (HUAC); • Croatia: Dalmatien-Split, Küste, 22.VI.1953, H. Hamann, 1 ♂ 1 ♀ (HUAC); • Croatia: Ribnica, Otavice 10 km N v. Titov Velea, 28.VII.1965, 1 ♂ 1 ♀ (RMNH); Croatia: • Rovinj, 15.V.1953, 1 ♂ (HUAC); • Croatia: Ninski Stanovi, 40 m, 14.07.2004, L. Ábrahám, 31 ♂♂ 34 ♀♀(SCM); • Croatia: Zadar, Benkovac, 11.07.2002, D. Szalóki, 3 ♀♀ (SCM); • Croatia: Dalmacija, Gor. Poličnik, 12 km E of Zadar, 20.06.2007, leg. Zs. Józan, 1 ♀ (SCM); • Croatia: Dalmacija, Primošten-Bilo, 09.06.2003, Nikola Rahmé 1 ♂ (SCM); • Croatia: Vinodol Obc., Crikvenica, Selce, 180 m, 01.07.2001, K. Székely 1 ♂ (SCM); • Croatia: Ninski Stanovi, 40 m, 14.07.2004, L. Ábrahám, 3 ♂♂ 3 ♀♀ (USMB); • France: Corse, Calvi, 13–27.VII.1971, Fam. V. Oorschot, 1 ♀ (RMNH); • France: Corse, Belgodère, 350 m, 14.VII.1971, Fam. V. Oorschot, 1 ♀ (RMNH); • France: Corse, Belvedere, VII.1973, H.M. Steiner, 3 ♂ 1 ♀ (HUAC); • France: Corsica, Valle de Restino, 02.06.2000, leg. L. Nádai, 1 ♀ (SCM); • France: Haute-Corse, Castifao, 25.06.2002, leg. J. Bard, 1 ♀ (SCM); • Greece: Ahaia, Michas, 1300 m, 38°00'34"N, 21°53'00"E, 7.VII.2008, Dils J. and Faes J., 1 ♀ (RMNH); • Greece: Chalcide, Tristinika, 20.VI.2015, 1♂ (MDC); • Greece: Cos, Tigaki, 3–10.VII.1978, M. C. and G. Kruseman, 2 ♀ (RMNH); • Greece: Georgioupolis, 35°21'N, 24°16'E, H. u. L. Hölzel, 24 ♂ 19 ♀ (NHMW); • Greece: Georgioupolis, 35°21'N, 24°16'E, H. u. L. Hölzel, 1 ♂ 1 ♀ (HUAC); • Greece: Elis, Andritsaina, 1000 m, 29.VII.1978, L. Willemse and J. Tilmans, 1 ♂ (RMNH); • Greece: Ioannina, Mesovouni, 650 m, 39°56'18.8"N, 20°38'12.4"E, 28.VII.2006, T., W. and J. Garrevoet, 2 ♀ (RMNH); • Greece: Lesbos, Sigri, 6–8.VI.1988, J. P. Duffels, 1 ♂ 1 ♀ (RMNH); • Greece: Loutraki-Perachora, 38°N, 23°E, 30.V.1969, Rausch, 1 ♀ (HUAC); • Greece: Pierias, Litochoro, 5.VII.1976, F. and L. Willemse and J. Tilmans, 1 ♀ (RMNH); • Greece, Creta Nord, Hersonissos, 35°18'36.3"N, 25°23'53.9"E, 15.V.2024, J. Vetrovec, 1 ♂ (DB); • Greece: Creta, Lasithi, Xerokampos, 2.VI.1972, M. C. and G. Kruseman, 1 ♂ 2 ♀ (RMNH); • Greece: Kardaras, 800 m, 37°39'22.9"N, 22°18'34.2"E, 7.VII.2008, Dils J. and Faes J., 1 ♂ (RMNH); • Greece: Tripolis-Davia 3–8 km NW of Tripolis, 800 m, 13.VII.1975, J. P. Duffels, 1 ♀ (RMNH); • Greece: Leptokaria, 28.06.1992, Balásházy, 1 ♀ (SCM); • Greece: Sithonia, Sarti, 0–50 m, 02–09.07.1997, D. Szalóki, 1 ♂ 4 ♀♀ (SCM); • Greece: Sithonia, Sarti, 0–50 m, 02–09.07.1997, D. Szalóki, 1 ♀ (USMB); • Greece: Argolída, SE of Néa Epídavros, 37°40'N, 23°09'E, 120 m, 09.06.2007, R. Królik, 1 ♂ (USMB); • Greece: Corfu, Acharavi [CK90], xerothermic meadow, 06.07.2011, W. Żyła, 3 ♂♂ (USMB); • Greece: Corfu, Acharavi [CK90], xerothermic meadow, 07.07.2011, W. Żyła, 1 ♂ (USMB); • Greece: Chalkidiki, Paliouri env., 39°56'31"N, 23°39'29"E, SE of Kassandria, 06–11.06.2013, M. Šnižek, 2 ♂♂ (USMB); • Greece: Chalkidiki, Paliouri env., 39°56'31"N, 23°39'29"E, SE of Kassandria, 12–14.06.2013, M. Šnižek, 3 ♂♂ (USMB); • Greece: Thessaly, Kokkino Nero, 39°49'N, 22°47'E, environs, 17–29.05.2015, A. Skiba, 1 ♂ (USMB); • Greece: Chalkidiki, E of Paliouri, 39°56'N, 23°41'E, Kassandria, 11.06.2022, M. Šnižek, 1 ♂ (USMB); • Greece: Peloponnese, Loutra Killinis, 37°51'12"N, 21°6'37"E, dunes and grass, 19.06.2023, W. Żyła, 1 ♀ (USMB); • Greece: Peloponnese, Messenia, SE of Kalamia, 37°52'13.5"N, 21°07'08.7"E, olive garden, 13.06.2024, W. Żyła, 2 ♀♀ (USMB); • Greece: Peloponnese, Messenia, SE of Kalamia, 37°52'13.5"N, 21°07'08.7"E, olive garden, 14.06.2024, W. Żyła, 1 ♂ 1 ♀ (in absolute alcohol) (USMB); • Italy: Basilicata, 10 km uoru, Potenza, 700 m, 40°35'N, 15°45'E, 1 ♂ (HUAC); • Italy: Calabria, Crotone, Catanzaro, 30–100 m, 38°55'N, 16°55'E, 4.VIII, 1 ♂ (HUAC); • Italy: Prov. Messina, Lipari, Isole, Stromboli, San Vincenzo, 17.VII.1995, Alain Olivier, 1 ♀ (RMNH); • Italy: Tuscany, Grosseto, Nomadelfia (GR), 42°50'23"N, 11°08'50"E, 29.VI.2024, Davide Badano and Yuchen Zheng, 2 ♂ 2 ♀ (ZCAU); • same locality • 1 ♂ 3 ♀ (DB); • Italy: Tuscany, Grosseto, Roselle (GR), 42°49'34"N, 11°10'05"E, 29.VI.2024, Davide Badano and Yuchen Zheng, 1 ♂ 2 ♀ (ZCAU); • same locality data • 1 ♀ (DB); • Italy: Tuscany, Grosseto, Parco Nazionale della Maremma, Marina di Alberese (GR), 42°39'51"N, 11°02'25"E 20.VI.2025, Davide Badano leg., 1 ♂ 2 ♀ (DB); • Italy: Lazio, Roma (RM), Via Appia Antica, 11.VII.1937, L. Barbera, 1 ♀ (MSNG); • Italy: Nomadelfia (GR), 42°50'23.2"N, 11°08'51.4"E, 30.VI.2024, D. Badano and Y. Zheng, (DB); • Italy: Puglia, Bari, Masseria Pellicciari, 450 m, 9.VII.2000, Fontana P., Kleukers R. and Cogo A., 1 ♀ (RMNH); • Italy: Puglia, Castel del Monte, 450 m, 41°05'20.9"N, 16°16'16.3"E, 2.VII.2001, T. and W. Garrevoet, 1 ♀ (RMNH); • Italy: Prov. Salerno, Pioppi, Cost Cilentana, 14.VII.1971, Holzinger, 1 ♀ (HUAC); • Italy: Sardegna, Marrubiu (OR), Sant’Anna, stazione e dintorni, 1.VI.2003, C. Meloni, 1 same locality, ♂ (MSNG); • same locality, 2.VI.2006, 1 ♀ (MSNG); • same locality, 26.VI.2009, C. Meloni, 3 ♂ 12 ♀ (MSNG); • Italy: Sardegna, Alghero (SS), Lu Cantar, 40°32'25.3"N, 8°19'29.7"E, 12.VII.2012, D. Badano, 3 ♂ (DB); • Italy: Sardegna, Alghero (SS), Monte Doglia, 40°36'20.1"N, 8°15'06.0"E, 7.VII.2012, D. Badano, 3 ♀ (DB); • Italy: Sardegna, Monti (OT), 2.VII.2011, D. Badano, 2 ♀ (DB); • Italy: Sicilia, Sud di Castrofilippo (AG), m 450, 4.VI.2015, E. Gallo, 1 ♀ (MSNG); • Italy: Sicilia, Caltanissetta (CL), strada Enna, bivio Gela, 8.VII.1954, Adalberto Parvis, 2 ♂ 3 ♀ (MSNG); • Italy: Sicilia, Peloritani, 10 km SW Barcellona, 100 m, 38.06°N, 15.07°E, 25–26.1974, 2 ♂ 2 ♀ (HUAC); • Italy: Sardinia, Siniscola, Su Tilio, 27.06.2004, V. Feik, 1 ♀ (NMPC); • Italy: Sardinia, “Sardinien, Mti Gennargentu Mte, Spada”, 1250 m, 17–21.06.1968, Ströhle, 2 ♂♂ (SCM); • North Macedonia: Demir Kapija, Barovo, 27.VII.1965, 1 ♀ (RMNH); • North Macedonia: Štip, 23.VII.1965, 1 ♂ (RMNH); • Turkey: Aydin, Kusadasi, 11.VII.1980, H. v. Oorschot, 1 ♀ (RMNH); • Turkey: Balıkesir, nahe Bezirci (zw. Edincik u. Bugdayli), 150 m, 40.15°N, 27.47°E, 22.VII.1978, H.and U. Aspöck, H and R Rausch, P. Ressl, 3 ♂ 4 ♀ (HUAC); • Turkey: Edirne, 08–13.06.[19]47, 1 ♂, Exp. N. Mus. ČRS (NMPC).
Variation of wing patterns in Deleproctophylla spp. A. D. australis (Fabricius, 1787), Crete (Greece); B. D. australis, Alghero, Sardinia (Italy); C. D. australis, Grosseto (Italy); D. D. australis, Balikesir (Turkey); E. D. bleusei Kimmins, 1949, Fes (Morocco); F. D. bleusei, Cabo de Gata (Spain); G. D. dusmeti (Navás, 1914), Saint-Paul-et-Valmalle (France); H. D. dusmeti, Villeneuve-Loubet (France).
Head capsule of the larvae of Deleproctophylla spp. A, B. D. australis (Fabricius, 1787); A. Head, dorsal view; B. Head, ventral view; C, D. D. dusmeti (Navás, 1914); C. Head, dorsal view; D. Head, ventral view.
Diagnostic characters.
Metafemur with brown marking; forewing with brown marking below pterostigma (often absent in eastern populations); hindwing elongate, posterior margin curved; not angled; hindwing with large marking below pterostigma; male ectoproct with branch in apical half of branch, after mid-length; setae on branch only at tip; setae on branch and tip of ectoproct stout and curved.
Redescription.
Size (mm, based on 5 specimens). Head + thorax length 7.7–8.6; antenna length 15.1–2; fore wing length 21.7–22.9; fore wing width 6.4–7.4; hindwing length 17.1–18.7; hindwing width 5.0–5.7.
Head. Vertex yellowish brown; setae dense, long, pale yellow (Fig. 6C). Para-ocular band pale yellow. Frons yellowish brown; setae dense, long, pale yellow. Clypeus and labrum yellowish brown; setae long, pale (Fig. 6B). Mandible yellowish brown, transitioning to dark brown apically. Labium and palp yellowish brown, concolorous with mandible. Antenna with pedicel with dense pale and dark setae, flagellomeres brown, nodes slightly darker; club brown, anterior face paler (Fig. 6A).
Deleproctophylla australis (Fabricius, 1787), adult. A. Habitus, Nomadelfia (Italy); B. Head, frontal view, Balikesir (Turkey); C. Head and thorax, dorsal view, Balikesir (Turkey); D. Head and thorax, lateral view. Abbreviations: C costa Sc subcosta RA radius anterior RP radius posterior MA media anterior MP media posterior CuA cubitus anterior CuP cubitus posterior A anal veins.
Thorax. Cervical sclerite yellowish brown. Pronotum brown; setae long, pale yellow. Mesonotum brown with yellowish brown markings: an anterior rounded pair, a median triangular pair, and a posterior rounded pair; mesoscutellum anteriorly dark brown, posteriorly yellowish brown. Mesonotum with long pale yellow setae. Metanotum largely yellowish brown, medially dark brown; setae pale yellow (Fig. 6C). Pleurae yellowish brown; setae pale yellow (Fig. 6D).
Legs. Pro- and mesothoracic legs yellowish brown, tarsi slightly darker. Metathoracic leg yellowish brown, metafemur with dorsal dark brown marking. Coxae with thin pale yellow setae; femora and tibiae with thin pale yellow setae and ventral thicker blackish setae; tibial spurs narrow, blackish; tarsi with robust blackish setae (Fig. 6D).
Wings. Forewing long, apex rounded. Pterostigma with four to five forked and unforked brown veinlets; membrane brown. Presectoral area with ~6 crossveins. RP with five branches. MA gradually curving toward hind margin in distal portion. MP-CuA area with ~3 irregular rows of cells. Anal area hind margin straight; axillary lobe slightly pronounced. Forewing venation largely dark brown, paler toward wing base; Sc and R light brown; anal veins pale brown. Forewing membrane hyaline (in old specimens with faint brownish), distal section usually with a brown marking in radial area, just below pterostigma (absent in some populations) (Figs 2A, 2B, 4A–D, 6A). Hindwing elongated, broadest at RP fork; hind margin smoothly curved gradually narrowing toward rounded apex. Presectoral area with three crossveins. RP with four branches. MA largely straight, slightly curved toward hind margin distally. MP-CuA area with ~2 irregular rows of cells. Hindwing venation dark brown, paler near wing base. Hindwing membrane hyaline, distal section with a rather large brown marking extending from pterostigma across radial area until first branch of RP (Figs 2A, 2B, 4A–D, 6A).
Abdomen. Tergites pale brown, each with a caudal pair of yellowish markings. Sternites pale brown, paler than tergites. Pleural membrane brown with large paler markings.
Male genitalia. Ectoproct narrow, with branch in apical half, after mid-length. Setae of ectoproct long, dark brown; setae on branch limited to tip, dense, black, stout, curved, and spine-like; setae only on tip of ectoproct, black, stout, curved, and spine-like (Fig. 7A–C). Tergite 9 sub-rectangular in lateral view, with ventral projection; setae long and black, longer and denser on projection. Sternite 9 with three lobes, of which median largest; setae thin, spine-like on median lobe (Fig. 7B, C). Gx11 arch-shaped, slightly curved downward in lateral view; gx9 strongly sclerotized, with upward fold; gp9 lightly sclerotized forming a shallow median furrow, with short setae (Fig. 7F–J).
Deleproctophylla australis (Fabricius, 1787), adult, Nomadelfia (Italy). Male terminalia: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia: D. Ventral view; E. Lateral view. Male genitalia: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view. Abbreviations: ect ectoproct br branch of ectoproct T8 tergite 8 S8 sternite 8 gx8 gonocoxite 8 gp8 gonapophysis 8 T9 tergite 9 S9 sternite 9 gx9 gonocoxite 9 gp9 gonoapophysis 9 gx11 gonocoxite 11.
Female genitalia. Ectoproct elongated; setae long, black (Fig. 7D, E). Tergite 9 sub-rectangular. Gx9 wider than long ventrally; setae long. Tergite 8 subrectangular. Gx8 wider than long, with long setae. Gp8 prominent; setae thin (Fig. 7D, E).
Larva. See Badano and Pantaleoni (2014) (Fig. 5A, B).
Distribution.
Asia: Azerbaijan (see remarks for D. variegata), Georgia, Turkey (see remarks). Europe: Albania, Bulgaria, Croatia, France (Corsica), Greece, Italy (including Sardinia and Sicily), North Macedonia, Turkey (Fig. 25) (H. Aspöck et al. 1980; Devetak 1995; H. Aspöck et al. 2001; Popov 2004; Devetak and Janžekovič 2012; Kerimova et al. 2022; Japaridze et al. 2024; Oswald 2025).
Life history notes.
Deleproctophylla australis is typically associated with arid grasslands, meadows, and clearings (H. Aspöck et al. 1980; Devetak 1995; Popov 2004) (Fig. 8). Deleproctophylla australis is usually active at daylight, despite flights at dusk with hot weather. The female seems to prefer ovipositing on short grass stems (Fig. 8A, B), the eggs are placed in rows, as typical for the tribe (Fig. 8D).
Deleproctophylla australis (Fabricius, 1787), live specimens, Nomadelfia (Italy). A, B. Female adult, laying eggs; C. Male adult; D. Eggs on a branch. Photos by YZ.
Remarks.
Deleproctophylla australis is the type and first described species of Deleproctophylla. Among all congeners, D. australis is also the species with the widest distribution, ranging from the Tyrrhenian islands to western Asia (Fig. 25). However, this owlfly is usually localized and not particularly common (H. Aspöck et al. 1980, 2001).
The reliability of the forewing marks as a diagnostic character of D. australis is challenged by its absence in some specimens both due to individual variation and geographic differences (Fig. 4A–D). Deleproctophylla australis is distinguished from congeners, particularly the highly similar D. dusmeti, by a combination of wing and genital characters; D. australis is characterized by the elongated hindwing without a prominent posterior angle (present in D. dusmeti) and male ectoproct with branch arising after mid-length (at mid-length in D. dusmeti). Western populations of D. australis are typically characterized by the presence of a forewing spot below pterostigma, a trait often considered diagnostic in distinguishing this species from its European congeners (e.g., H. Aspöck et al. 1980) (Figs 2A, 2B, 4A–D, 6A). However, the frequency of this forewing marking diminishes eastward, with the populations of the Balkan Peninsula including spotted and unspotted specimens and those of Anatolia lacking the spot (Fig. 4D). These Asian unmarked specimens were assigned to a “dusmeti-like morph”, due to the shared absence of forewing marking and presence of spot on metafemur (H. Aspöck et al. 1980). The re-examination of specimens from these populations suggests that these enigmatic individuals from Anatolia belong to D. australis, as supported by wing shape and characters of male genitalia. Nevertheless, specimens of D. australis without spotted fore wings can be present in all populations, including Italy (Fig. 4B).
Deleproctophylla
bleusei
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
Kimmins, 1949
81ED67A9-402A-5FD4-A0B8-B49545A71696
Figs 2C, 2D, 4E, 4F, 9, 10, 24A, 24B
Deleproctophylla bleusei Kimmins, 1949: 28 (type locality: Algeria: Naama, Mécheria; holotype in NHMUK). H. Aspöck and Hölzel 1996: 78 (Deleproctophylla). H. Aspöck et al. 2001: 302 (Deleproctophylla). Güsten 2003: 133 (Deleproctophylla). Monserrat et al. 2014: 157 (Deleproctophylla). Monserrat 2015: 478 (Deleproctophylla). Michel and François 2019: 183 (Deleproctophylla). Monserrat et al. 2022: 81 (Deleproctophylla).
Material examined.
Type specimen: Holotype. Images available at https://data.nhm.ac.uk/object/73295e3e-26cd-4bf4-8c10-0879722377ff.
Additional material.
• Morocco: Fes-el-Bali (60 km SE Ouezzane), 34.38°N, 5.16°W, 300 m, 8.VII.1977, 4 ♂, 6 ♀ (HUAC); • Spain: Alicante, Cox, VIII.1911, Andréu, 1 ♂ (MCNM); • Morocco: Middle Atlas, Parc National de Tazekka, env. of Barrage Bab Louta Lake, 34°0'5.90"N, 4°18'28.70"W, 590 m a.s.l., dry, ruderal meadow, 15.06.2022, 2 ♂, W. Szczepański (USMB); • some locality, date, 1 ♂, W. Szczepański (SCM); • Spain: Almería, Cabo de Gata, 30SWF67, 27.V.2010, F. Rodríguez and J.R. Correas, 1 ♂ (VM); • same locality 16.VI.2013, 2 ♂, 1 ♀ (VM); • same locality 6.VII.2013, 1 ♂, 2 ♀ (VM), 1 ♀ (DB); • Almería, Cabo de Gata, Salinas, Las Amoladeras, 30SWF67, 14.VI.2012, 1 ♀ (VM); • same locality, 6.VII.2013, 2 ♀ (VM); • same locality, 30.VII.2013, 1 ♀ 1 ♂ (VM); • Spain: Almería, San José, Playa de los Genoveses, 14.VI.2012, 3 ♂ 2 ♀ (VM); • Spain: Almería, San José, Playa de Mónsul, 10 m, 14.VI.2012, 1 ♀ (VM); • Spain: Murcia, Lorca, Purias, 30SXG25, 550 m, 12.VI.2010, José Carrillo, 1 ♀(VM); • same locality, 19.VI.2010, José Carrillo, 1 ♂ (VM).
Diagnostic characters.
Metafemur unmarked; forewing unmarked; hindwing posterior margin markedly curved, convex; hindwing with marking below pterostigma; male ectoproct with branch long, in apical half of branch, just after mid-length; setae on branch only at tip; setae on branch and tip of ectoproct stout and curved.
Redescription.
Size (mm, based on 4 specimens) Head + thorax length 6.7–7.1; antenna length 11.6–14.5; fore wing length 15.7–22.4; fore wing width 6.5–6.7; hindwing length 19.2–22.7; hindwing width 4.2–5.5.
Head. Occiput yellowish brown; setae dense, long, pale yellow (Fig. 9C). Vertex yellowish brown; setae dense, long, pale yellow (Fig. 9B). Para-ocular band pale yellow. Frons yellowish brown; setae dense, long, pale yellow. Clypeus and labrum yellowish brown; setae long, pale. Mandible yellowish brown, darker apically. Labium and palp yellowish brown. Antenna with pedicel with dense pale and dark setae, flagellomeres brown, nodes slightly darker; club brown, paler anteriorly (Fig. 9A).
Deleproctophylla bleusei Kimmins, 1949, Fes (Morocco). A. Habitus; B. Head, frontal view; C. Head and thorax, dorsal view; D. Head and thorax, lateral view.
Thorax. Cervical sclerite yellowish brown. Pronotum brown; setae long, pale yellow. Mesonotum yellowish brown; internal margins of sclerites with darker brown shades; mesoscutellum yellowish brown with faint median brown stripe; setae long, pale yellow. Metanotum yellowish brown, with faint median marking; setae pale yellow (Fig. 9C). Pleurae yellowish brown; setae pale yellow (Fig. 9D).
Legs. Pro- and mesothoracic legs entirely yellowish brown. Metathoracic leg yellowish brown, unmarked. Setae of coxae pale yellow; setae of femora and tibiae thin, pale yellow, thicker blackish setae ventrally; tibial spurs narrow; setae of tarsi blackish (Fig. 9D).
Wings. Forewing long, apex rounded. Pterostigma with ~4 forked and unforked brown veinlets; membrane very pale brown, almost whitish in young (but not freshly emerged) individuals. Presectoral area with ~5 crossveins. RP with five branches. MA progressively curving toward hind margin in distal portion. MP-CuA area with ~3 irregular rows of cells. Axillary lobe slightly prominent. Forewing venation largely brown, paler toward wing base; Sc and R yellowish, very pale in young individuals; anal veins yellowish. Forewing membrane hyaline (in old specimens faint brownish), unmarked (Figs 2C, 2D, 4E, 4F, 9A). Hindwing broad, wider at RP fork; hind margin markedly convex, with a pronounced curve. Presectoral area with three crossveins. RP with four branches. MA largely straight, slightly curved toward hind margin distally. MP-CuA area with ~2 irregular rows of cells. Hindwing venation brown, paler near wing base; Sc yellowish. Hindwing membrane hyaline, distal section with a long narrow brown marking, sometimes stripe-like, extending from pterostigma to first branch of RP (Figs 2C, 2D, 4E, 4F, 9A).
Abdomen. Tergites pale brown, each with a caudal pair of yellowish markings. Sternites pale brown, paler than tergites. Pleural membrane pale brown.
Male genitalia. Ectoproct narrow, with long and thin branch bifurcating beyond mid-length, in apical half. Setae of ectoproct long, dark brown; setae on branch only at apex, dense, black, stout, curved, and spine-like; setae on tip of ectoproct black, stout, curved, and spine-like (Fig. 10A–C). Tergite 9 sub-rectangular in lateral view, with prominent ventral projection; setae long and black, longer and denser on projection. Sternite 9 with three lobes, of which median largest; setae thin, spine-like on median lobe (Fig. 10B, C). Gx11 arch-shaped, projecting caudally and curved downward laterally; gx9 sclerotized, dorsally folded; gp9 lightly sclerotized forming a shallow median furrow (Fig. 10F–J).
Deleproctophylla bleusei Kimmins, 1949, Fes (Morocco). Male terminalia: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia: D. Ventral view; E. Lateral view. Male genitalia: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view.
Female genitalia. Ectoproct elongated; setae long, black (Fig. 10D, E). Tergite 9 sub-rectangular. Gx9 wider than long; setae long. Tergite 8 subrectangular. Gx8 wider than long, with long setae. Gp8 projecting ventrally; setae thin (Fig. 10D, E).
Comparative notes.
Deleproctophylla bleusei has a long history of taxonomic confusion with D. dusmeti, to which it is sympatric in the Iberian Peninsula and closely resembles in wing pattern (Monserrat et al. 2014). However, D. bleusei can be distinguished by the unmarked metafemur (Fig. 9D) and characters of male ectoproct, such as long and thin branch arising after mid-length (Fig. 10A). The shape of the hindwing provides a reliable diagnostic trait: D. bleusei has smoothly curved posterior margin in D. bleusei, whereas D. dusmeti is characterized by a distinctly triangular shape. Additionally, young (but not newly hatched) specimens of D. bleusei are characterized by a pale C, Sc and R veins, as well as an almost whitish pterostigma. This distinctive venation coloration is also evident in the holotype.
Distribution.
Africa: Algeria, Morocco, Tunisia. Europe: Spain (H. Aspöck et al. 2001; Monserrat et al. 2014; Oswald 2025) (Fig. 25).
Deleproctophylla
dandizenor
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
Badano, Zheng, U. Aspöck & Dobosz sp. nov.
40445BE1-F3D7-56FD-8718-58C579950214
https://zoobank.org/D2497E1D-FABE-4B44-9F61-93B1E542BC5F
Type material.
Holotype • Afghanistan: Pr. Kunar, Nuristan ob. LindaiSin-Tal vic. Barg e Matal, Dandizenor Mts., 2600 m, 13.VII.1970, Naumann 1 ♂ (NHMW); paratype, same information as holotype, 1 ♀ (NHMW). Paratypes • Afghanistan: Province Bamiyan, Yakawlang District, SW Kotak village, ca 2800 m, 03.07.2016, I. Pljushtch, 1 ♂ (SCM); • Afghanistan: Province Bamiyan, Bande-Amir National Park, 2950 m, 26.06.2016, I. Pljushtch, 1 ♂ (USMB); • Pakistan: Hindukush Mts., Booni, 2000 m, 11.07.1994, B. Herczig, Gy. M. László and G. Ronkay, 1 ♂ (SCM).
Diagnosis.
Metafemur with large brown marking; pterostigma of both wings dark brown; wing venation rather dense, with many crossveins; forewing hyaline; hindwing broad; hindwing with large band-shaped marking from pterostigma to posterior margin of wing; male ectoproct robust, with branch in apical two-thirds of branch; ectoproct, including branch and apex densely covered with setae; setae on branch and apex spine-like. Male tergite 9 with caudal margin with long hair-like setae, ventral projection stout, hairy. Female ectoproct with short ventral projection. Female gx9 and gp8 relatively prominent.
Description.
Size (mm, based on 2 specimens) Head + thorax length 8.1–8.2; antenna length 15.4–15.5; fore wing length 20.3–21.9; fore wing width 6.5–6.8; hindwing length 15.5–17.7; hindwing width 5.4–5.5.
Head. Vertex yellowish brown; setae dense, long, pale yellow (Fig. 11C). Para-ocular band yellowish brown, darker anteriorly. Frons yellowish brown; setae dense, long, pale yellow. Clypeus and labrum yellowish brown; setae long, pale (Fig. 11B). Mandible yellowish brown, transitioning to brown apically. Labium and palp yellowish brown, concolorous with mandible. Antenna with pedicel with pale and dark setae, flagellomeres brown; club brown (Fig. 11A).
Deleproctophylla dandizenor Badano, Zheng, U. Aspöck & Dobosz, sp. nov., Kunar (Afghanistan). A. Habitus; B. Head, frontal view; C. Head and thorax, dorsal view; D. Head and thorax, lateral view.
Thorax. Cervical sclerite yellowish brown. Pronotum brown; setae long, pale yellow. Mesonotum brown with yellowish brown markings: an anterior small, rounded pair, a median subtriangular pair, and a posterior rounded pair; mesoscutellum dark brown, caudal margin yellowish brown. Mesonotum with long pale-yellow setae. Metanotum brown, medially dark brown; setae pale yellow (Fig. 11C). Pleurae yellowish brown; setae pale yellow (Fig. 11D).
Legs. Pro- and mesothoracic legs yellowish brown, tarsi dark brown. Metathoracic leg yellowish brown, metafemur with large dark brown marking. Coxae with thin pale yellow setae; femora and tibiae with thin pale yellow setae and ventral thicker blackish setae; tibial spurs narrow, blackish; tarsi with robust blackish setae (Fig. 11D).
Wings. Forewing long, broad, apex rounded. Venation dense, with many crossveins. Pterostigma with four or five forked and unforked brown veinlets; membrane dark brown. Presectoral area with ~4 crossveins. RP with five branches. MA gradually curving towards hind margin in distal portion. MP-CuA area with ~3 irregular rows of cells. Anal area hind margin straight; axillary lobe inconspicuous. Forewing venation dark brown, Sc and R paler; anal veins pale brown. Forewing membrane hyaline. Hindwing broad, widest after RP fork; hind margin smoothly curved toward rounded apex. Presectoral area with three crossveins. RP with four branches. MA straight, only slightly curved. MP-CuA area with ~2 irregular rows of cells (Figs 2E, 2F, 11A). Hindwing venation dark brown, paler near wing base. Hindwing membrane hyaline, distal section with a large brown marking extending from pterostigma to cubital area, shaped as curved band gradually narrowing posteriorly, pigmentation bordering crossveins, central part of cells paler; marking fainter in males than in females (Figs 2E, 2F, 11A).
Abdomen. Tergites brown. Sternites brown, paler than tergites. Pleural membrane brown with large paler markings.
Male genitalia. Ectoproct robust, with branch at two-thirds of length and apex stout. Setae of ectoproct dense, long, dark brown, gradually longer caudally; setae on whole branch, denser, and spine-like at apex; setae on apex of ectoproct spine-like (Fig. 12A–C). Tergite 9 sub-rectangular in lateral view, with robust ventral projection; setae long and black, longer and denser on caudal margin of the sclerite and on projection. Sternite 9 with lateral lobes inconspicuous, rather small, median lobe prominent, apically rounded (Fig. 12B, C). Gx11 arch-shaped, slightly curved; gx9 strongly sclerotized, with prominent dorso-apical fold, giving to apex a hooked appearance, gp9 relatively small, lightly sclerotized forming a shallow median furrow, with short setae (Fig. 12F–J).
Deleproctophylla dandizenor Badano, Zheng, U. Aspöck & Dobosz, sp. nov., Kunar (Afghanistan). Male terminalia of holotype: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia of paratype: D. Ventral view; E. Lateral view. Male genitalia of holotype: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view.
Female genitalia. Ectoproct elongated, relatively short and stout ventral projection; setae long, black (Fig. 12D, E). Tergite 9 sub-rectangular. Gx9 wider than long, prominent; setae long. Tergite 8 subrectangular. Gx8 prominent, as long as wide, with long setae. Gp8 projecting ventrally, with prominent caudal margin; setae thin (Fig. 12D, E).
Distribution.
Asia: Afghanistan, Pakistan (Fig. 25).
Etymology.
The species epitheton is derived from the name of a mountain range in the east of Afghanistan. It is a substantive and an apposition to the genus.
Comparative notes.
Deleproctophylla dandizenor is a broad-winged species, easily distinguishable from all its congeners by the narrow, curved, hindwing marking that extends from pterostigma almost to the posterior wing margin. The hindwing appears more pigmented in female in comparison to male, in which the marking is reduced to an infuscation (Fig. 2E, F). A similar difference in wing pattern is also observed in D. tengri sp. nov., despite the shape of markings is very different in these two species. However, due to the limited number of known specimens, it remains unclear whether these differences are attributable to sexual dimorphism or individual variation. The male of D. dandizenor is characterized by the robust hairy ectoproct with branch that bifurcates after mid-length, and stout apex. Moreover, the spine-like setae on the branch and on the apex of the ectoproct are rather long and slightly curved and they extend for the whole length of both structures (Fig. 12A–C). The hair-like setae on the caudal margin of tergite 9 are also diagnostic of this species. The female of D. dandizenor differs from the other species of the genus by the relatively stout ectoproct, with very short ventral projection and the prominent gx8 and gp8 (Fig. 12D, E).
Deleproctophylla
dusmeti
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
(Navás, 1914)
B7B52E33-475C-580D-9865-317C008BD42E
Figs 2G, 2H, 4G, 4H, 5C, 5D, 13, 14
Theleproctophylla dusmeti Navás, 1914: 57 (type locality: Spain: Madrid, Montarco; lectotype in MNCN). Navás 1915a: 68 (Theleproctophylla). Navás 1915b: 461 (Theleproctophylla). Bohigas and Sánchez 1917: 311 (Theleproctophylla). Navás 1920: 7 (Deleproctophylla). Escribano 1921: 365 (Theleproctophylla). Monserrat 1978: 182 (Deleproctophylla). H. Aspöck et al. 1980: 316 (with some questionable distribution, Deleproctophylla). Monserrat 1982: 70 (Deleproctophylla). H. Aspöck and Hölzel 1996: 78 (with some questionable distribution, Deleproctophylla). Sziráki 1998: 61 (with some questionable distribution, Deleproctophylla). H. Aspöck et al. 2001:302 (Deleproctophylla); Whittington 2002: 377 (with some questionable distribution, Deleproctophylla). Canbulat and Kiyak 2004: 114 (probably misidentification, Deleproctophylla). Canbulat 2007: 36 (probably misidentification, Deleproctophylla). Dobosz and Ábrahám 2007: 24 (probably misidentification, Deleproctophylla). Koçak and Kemal 2012: 17 (probably misidentification, Deleproctophylla). Monserrat et al. 2012: 38 (Deleproctophylla). Monserrat 2013: 298 (Deleproctophylla). Monserrat et al. 2014: 153 (Deleproctophylla). Badano and Pantaleoni 2014: 301 (Deleproctophylla). Aistleitner 2019: 123 (with some questionable distribution, Deleproctophylla). Hassan et al. 2019: 513 (questionable distribution, Deleproctophylla). Aistleitner and Lencina 2020: 102 (Deleproctophylla). Monnerat and Ábrahám 2020: 144 (questionable distribution, Deleproctophylla). Hassan and Liu 2021: 416 (questionable distribution, Deleproctophylla). Poggi 2024: 9 (Deleproctophylla).
Examined material.
Type specimen. Paralectotype: • Granada, 8.VII.01, Typus, Theleproctophylla dusmeti Navás SJ det., Theleproctophylla dusmeti Navás, 1914, Paralectotype, J. Legrand det, 1 ♀ (MNHN) (Fig. 24A, B).
Other specimens.
• France: Alpes Maritimes, Villeneuve Loubet, 43°39'44.08"N, 3°40'36.14"E, 16.VII.2011, D. Badano, 4 ♂ 5 ♀ (DB); • France: Bouches-du-Rhône, Eygalières, Le Mas de Montfort, m 170, 43°44'48.7"N, 04°55'17.7"E, 26.VI.2019, 2 ♂ (DB); • France: Besse/Var, 250 m, 18.VII.1964, Bilek, 1 ♂ 1 ♀ (HUAC); • France: Hérault, Gigean, 26–29.VI.2014, A. Prost, 2 ♂ 1 ♀ (MDC); • France: Hérault, Prades Le Lez, 43°40'58.78"N, 3°52'33.30"E, 11.VII.2011, D. Badano, 3 ♀ (DB); • France: Hérault, St. Paul et Valmalle, 43°36'36.14"N, 3°40'36.14"E, 26.VI.2011, D. Badano, 2 ♂ 1 ♀ (DB); • same locality, 29.VI.2011, D. Badano, 2 ♀ (DB); • same locality, 5.VII.2011, D. Badano, 4 ♂ 1 ♀ (DB); • same locality, 10.VII.2011, D. Badano, 3 ♂ 3 ♀ (DB); • France: Rochefort-du-Gard, 18.07.2002, J. Bard, 1 ♂ (SCM); • Portugal: Algarve, Odelouca, 14.VI.1970, C. and A. Jeekel, 1 ♀ (RMNH); • Spain: Aragona, Sena, 17.VII.1923, L. Navás, 1 ♂ 1 ♀ (MSNG); • Spain: Granada, 8.VII.1901, 1 ♀ (MNHN); • Spain: Madrid, Coll. Lacroix, 1 ♂ 2 ♀ (MNHN); • Spain: Zaragoza, Pina de Ebro, 05.07.1991, V. Redondo, 1 ♂ (SCM).
Diagnostic characters.
Metafemur with brown marking; forewing unmarked; hindwing subtriangular, posterior margin angled; hindwing with marking below pterostigma; male ectoproct with branch at mid-length; setae on branch only at tip; setae on branch and tip of ectoproct stout and curved.
Redescription.
Size (mm, based on 5 specimens). Head + thorax length 7.8–8.3; antenna length 14.1–16.5; fore wing length 19.7–23.8; fore wing width 5.6–6.4; hindwing length 15.3–18.4; hindwing width 4.7–5.8.
Head. Occiput yellowish brown; setae dense, long, pale yellow. Vertex yellowish brown; setae dense, long, pale yellow (Fig. 13C). Para-ocular band pale yellow. Frons yellowish brown; setae dense, long, pale yellow. Clypeus and labrum yellowish brown; setae long, pale (Fig. 13B). Mandible yellowish brown, dark brown apically. Labium and palp yellowish brown. Antenna with pedicel with dense pale and dark setae, flagellomeres brown, nodes slightly darker; club brown, paler anteriorly (Fig. 13A).
Deleproctophylla dusmeti (Navás, 1914). A. Habitus, Saint-Paul-et-Valmalle (France); B. Head, frontal view, Toulon (France); C. Head and thorax, dorsal view, Toulon (France); D. Head and thorax, lateral view, Besse-sur-Issole (France). Abbreviations: C costa Sc subcosta RA radius anterior RP radius posterior MA media anterior MP media posterior CuA cubitus anterior CuP cubitus posterior A anal veins.
Thorax. Cervical sclerite yellowish brown. Pronotum brown; setae long, pale yellow. Mesonotum brown with yellowish brown markings: an anterior stripe-like pair, a median triangular pair, and a posterior rounded pair; mesoscutellum dark brown with a pair of yellowish rounded markings and posterior margin; setae pale yellow. Metanotum largely yellowish brown, medially dark brown; setae pale yellow (Fig. 13C). Pleurae yellowish brown; setae pale yellow (Fig. 13D).
Legs. Pro- and mesothoracic legs yellowish brown, tarsi slightly darker. Metathoracic leg yellowish brown, dorsal side of metafemur with dark brown marking. Setae of coxae pale yellow setae; setae of femora and tibiae thin and pale yellow dorsally, thicker and blackish ventrally; tibial spurs narrow; setae of tarsi blackish (Fig. 13D).
Wings. Forewing long, apex rounded. Pterostigma with four to five forked and unforked brown veinlets; membrane brown. Presectoral area with five crossveins. RP with five branches. MA gently toward hind margin. MP-CuA area with ~3 irregular rows of cells. Anal area hind margin straight; axillary lobe slightly pronounced. Forewing venation largely dark brown, paler toward wing base; Sc and R pale brown; anal veins pale brown. Forewing membrane hyaline (Figs 2G, 2H, 4G, 4H, 13A). Hindwing broad, subtriangular, widest after RP fork; hind margin angled, gradually narrowing toward rounded apex. Presectoral area with three crossveins. RP with four branches. MA largely straight, slightly curved distally. MP-CuA area with ~2 irregular rows of cells. Hindwing venation dark brown, paler near wing base. Hindwing membrane hyaline, distal section with a rather large brown marking extending from pterostigma across to second branch of RP (Figs 2G, 2H, 4G, 4H, 13A).
Abdomen. Tergites pale brown, with caudal pairs of yellowish markings. Sternites pale brown, paler than tergites. Pleural membrane brown with large paler markings.
Male genitalia. Ectoproct narrow, with branch at mid-length. Setae of ectoproct long, dark brown; setae on branch limited to tip, dense, black, stout, curved, and spine-like; setae only on tip of ectoproct, black, stout, curved, and spine-like (Fig. 14A–C). Tergite 9 sub-rectangular in lateral view, with ventral projection; setae long and black, longer and denser on projection. Sternite 9 with paired lateral and a single median lobe; setae thin, spine-like on median lobe (Fig. 14B, C). Gx11 arch-shaped, projecting caudally and slightly curved downward in lateral view; gp11 lightly sclerotized, forming a median furrow, with minute setae; gx9 widely set apart, strongly sclerotized, dorsally folded (Fig. 14F–J).
Deleproctophylla dusmeti (Navás, 1914), Madrid (Spain). Male terminalia: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia: D. Ventral view; E. Lateral view. Male genitalia: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view. Abbreviations: ect ectoproct br branch of ectoproct T8 tergite 8 S8 sternite 8 gx8 gonocoxite 8 gp8 gonapophysis 8 T9 tergite 9 S9 sternite 9 gx9 gonocoxite 9 gp9 gonapophysis 9 gx11 gonocoxite 11.
Female genitalia. Ectoproct elongated, projecting ventrally; setae long, black. Tergite 9 sub-rectangular. Gx9 wider than long; setae long (Fig. 14D, E). Tergite 8 subrectangular. Gx8 wider than long, with long setae. Gp8 projecting ventrally; setae thin (Fig. 14D, E).
Larva. See Badano and Pantaleoni (2014) (Fig. 5C, D).
Distribution.
Europe: France, Portugal, Spain (H. Aspöck et al. 1980; Rehfeldt 1989; H. Aspöck et al. 2001; Monserrat et al. 2014; Oswald 2025) (Fig. 25).
Comparative notes.
Deleproctophylla dusmeti may be difficult to distinguish from its two congeners in western Europe, D. australis and D. bleusei. The wing patterns traditionally used to differentiate these species are unreliable and exhibit high individual variation (Fig. 4). The diagnostic characters of D. dusmeti include a triangular-shaped hindwing (in contrast to the elongated hindwing of D. australis) and a male ectoproct branch that protrudes at mid-length (whereas in D. australis and D. bleusei the branch arises after mid-length) (Figs 13A, 14A).
Remarks.
D. dusmeti has been reported from several Asian countries, including Azerbaijan, Turkey, and Pakistan (H. Aspöck et al. 2001; Canbulat and Kiyak 2004; Canbulat 2007; Dobosz and Ábrahám 2007; Koçak and Kemal 2012; Hassan et al. 2019; Monnerat and Ábrahám 2020; Hassan and Liu 2021; Oswald 2025). However, the examination of the specimens assigned to the “dusmeti-like form” (see H. Aspöck et al. 1980) suggest that these instead belong to D. australis, representing individuals without forewing marking, a common trait in eastern populations of this species. The vague report from Pakistan (Hassan et al. 2019; Hassan and Liu 2021; Oswald 2025), which lacks any locality information cannot be verified at present. It may actually refer to D. dandizenor sp. nov., another species with unmarked forewing. The current observations suggest that D. dusmeti is exclusively a western European species. Deleproctophylla dusmeti is typically distinguished from D. australis by the absence of forewing spots. However, some populations along the southeastern coast of France include individuals with marked forewings mixed with unmarked ones (Fig. 4H). The study of genital morphology and species delimitation analyses confirm that these specimens belong to D. dusmeti (Fig. 1). These spotted individuals are likely the source of records of D. australis in southern France, despite this species is absent from the French mainland (though it is present in Corsica). The distributions of D. dusmeti and D. australis do not overlap, being separated by a relatively small geographic gap along the northern Tyrrhenian coast of Italy (Fig. 25).
Deleproctophylla
gelini
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
(Navás, 1919)
F401A582-E6BF-5788-88B5-DC172056FE51
Theleproctophylla gelini Navás, 1919: 21 (type locality: Morocco: Safi; holotype in MNHN but lost at present). Navás 1930: 121 (Theleproctophylla). H. Aspöck and Hölzel 1996: 78 (Deleproctophylla). H. Aspöck et al. 2001:302 (Deleproctophylla). Whittington 2002: 377 (Deleproctophylla). Ábrahám and Mészáros 2004: 333 (Deleproctophylla). Faucheux 2006: 204 (Deleproctophylla). Faucheux et al. 2012: 171 (Deleproctophylla). Ábrahám 2017: 126 (Deleproctophylla).
Examined material.
• Morocco: Agadir, 27.V–6.VI.1973, C. and A. Jeekel, 2 ♀ (RMNH); • Morocco: Bou Tazzert, Pres Mogadoa, 1928, R. Benoist. 2 ♂, 1 ♀, 2 ex. (MNHN); • Morocco: Tamri, 30°42'31.9"N, 09°51'26.1"W, 15 m, 30.06.2008, L. Ábrahám, L. Bognár and L. Nagy, 5 ♂♂ 1 ♀ (SCM); • Morocco: Tamri, 30°42'31.9"N, 09°51'26.1"W, 15 m, 30.06.2008, L. Ábrahám, L. Bognár and L. Nagy, 1 ♂ 1 ♀ (USMB); • Morocco: Atlantic Plain, 4 km W Agoudi n’Aït Amar 30°42'31.3"N, 9°51'26.2"W, 10 m, 27.06.2024, W. Szczepański, 4 ♂♂ 2 ♀♀ (USMB);
Diagnostic characters.
Vertex as long as wide; metafemur without brown marking; forewing extensively marked; hindwing broad, posterior margin curved; forewing with complex pattern: brown marking at base, subcostal area brown, brown marking at RP bifurcation, and large brown marking from pterostigma to first branch of RP; hindwing with a similar pattern: brown marking at base, subcostal area brown, brown marking at RP bifurcation, and large brown marking from pterostigma to posterior margin; male ectoproct bent inward, with branch at mid-length and protruding dorsally; setae on branch only at tip; setae on tip of ectoproct robust and relatively long; male with gx11 straight, broad, incapsulating gx9; gx9 without apical fold; female ectoproct long; gx8 and gp8 not prominent.
Redescription.
Size (mm, based on 3 specimens). Head + thorax 7.4–9; antenna length 14.9–16.9; fore wing length 21–24.9; fore wing 6.2–7.3; hindwing 16.1–20.5; hindwing width 5.2–6.1.
Head. Vertex nearly as long as wide, yellowish brown; setae dense, long, pale yellow (Fig. 15C). Para-ocular band pale yellow. Frons yellowish brown; setae dense, long, pale yellow. Clypeus and labrum yellowish brown; setae long, pale (Fig. 15B). Mandible yellowish brown, dark brown apically. Labium and palp yellowish brown. Antenna with pedicel with dense pale and dark setae, flagellomeres pale brown, nodes dark brown; club dark brown, paler anteriorly (Fig. 15A).
Deleproctophylla gelini (Navás, 1919), Guigou (Morocco). A. Habitus; B. Head, frontal view; C. Head and thorax, dorsal view; D. Head and thorax, lateral view.
Thorax. Cervical sclerite yellowish brown. Pronotum brown; setae long, pale yellow. Mesonotum yellowish brown gradually darker medially and along sclerites margins. Metanotum yellowish brown, medially darker; setae pale yellow (Fig. 15C). Pleurae yellowish brown; setae pale yellow (Fig. 15D).
Legs. Pro- and mesothoracic legs yellowish brown, including tarsi. Metathoracic leg yellowish brown, unmarked. Setae of coxae pale yellow setae; setae of femora and tibiae thin and pale yellow dorsally, thicker and blackish ventrally; tibial spurs narrow; setae of tarsi blackish (Fig. 15D).
Wings. Forewing broad, apex rounded. Pterostigma with ~5 forked and unforked brown veinlets; membrane brown. Presectoral area with five crossveins. RP with ~5 branches. MA gently toward hind margin. MP-CuA area with ~3 irregular rows of cells. Anal area hind margin straight; axillary lobe slightly pronounced. Forewing venation largely pale brown. Forewing membrane largely hyaline with a complex brown pattern: base of wing brown, apical costal crossveins with brown shade at base, subcostal area brown, brown marking at RP branching, apical portion of wing with a large brown marking extending from subcostal area and pterostigma to first branch of RP (Figs 3A, 3B, 15A). Hindwing broad, widest at RP fork; hind margin rounded, smoothly curved toward rounded apex. Presectoral area with three crossveins. RP with five branches. MA largely straight, slightly curved distally. MP-CuA area with ~2 irregular rows of cells. Hindwing venation pale brown, Hindwing membrane hyaline, with a complex brown pattern: base of wing brown, apical costal crossveins with brown shade at base, subcostal area brown, brown marking at RP branching, apical portion of wing with a large brown marking extending from subcostal area and pterostigma to posterior margin of wing (Figs 3A, 3B, 15A).
Abdomen. Tergites pale brown. Sternites pale brown, paler than tergites. Pleural membrane brown.
Male genitalia. Ectoproct robust, strongly bent inward, with branch slightly beyond mid-length. Branch projecting dorsally, broad, and relatively thick, progressively widening at apex. Apex of ectoproct club-shaped expanding internally. Setae of ectoproct short, dark brown; setae on branch sparse, those on apical section, black, stout, and curved; setae on tip of ectoproct progressively denser caudally, black, stout, curved, and spine-like on the internal side (Fig. 16A–C). Tergite 9 trapezoid in lateral view, with broad ventral projection; setae long and black, longer and denser on projection (Fig. 16B, C). Sternite 9 without paired lateral lobes, being composed only by the median lobe; setae thin, spine-like lobe. Gx11 arch-shaped, projecting caudally, broad and incapsulating gx9, straight in lateral view; gp11 lightly sclerotized, elongated, with a shallow median furrow, with minute setae; gx9 widely set apart, strongly sclerotized, flattened, without dorsal fold (Fig. 16F–J).
Deleproctophylla gelini (Navás, 1919), Guigou (Morocco). Male terminalia: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia: D. Ventral view; E. Lateral view. Male genitalia: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view.
Female genitalia. Ectoproct strongly elongated, projecting ventrally; setae long, black (Fig. 16D, E). Tergite 9 sub-rectangular. Gx9 wider than long; setae long. Tergite 8 subrectangular. Gx8 not prominent, with long setae. Gp8 not prominent; setae thin. Sternite 7 with sparse stout setae (Fig. 16D, E).
Distribution.
Africa: Morocco (H. Aspöck et al. 2001; Ábrahám and Mészáros 2004; Oswald 2025).
Comparative notes.
Among all Deleproctophylla species, D. gelini is the easiest to identify due to its unique fore- and hindwing pattern (Ábrahám and Mészáros 2004) (Fig. 3A, B). Additionally, this species differs from all congeners in both male and female genitalia, which are characterized by several distinctive characters (Ábrahám and Mészáros 2004) (Fig. 16). The male ectoproct is stout and distinctly bent inward and the branch arises dorso-caudally at mid-length. The branch stacks on the ectoproct, instead of being directed inward, as seen in all other species. Furthermore, the sternite 9 of the male lacks the lateral lobes found in other species (Fig. 16A–C). The gx9+11 complex is also different being, shaped like a capsule. The Gx11 is broad, not curved downward, and encases the gx9, which lacks the apical dorsal fold present in all other species (Fig. 16F–J). The female of D. gelini is characterized by an elongated ectoproct projecting ventrally and reduced gx8 (Fig. 16D, E).
Deleproctophylla
tengri
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
Zheng, Badano, H. Aspöck & Liu sp. nov.
67F1EA64-EACE-5BEF-8317-C0FA5BB2A1AA
https://zoobank.org/885D76FC-EBFA-4850-B43A-78355B359BCA
Type material.
Holotype • China: Xinjiang, Shihezi [石河子], Maangou Village [马鞍沟村], Mt. Jiangjunshan [将军山], 650 m, 6.VI.2020, Haitian Song, 1 ♂ (IZCAS). Paratypes • Same locality as holotype, 18.VII.2022, Rui Wang, 1 ♀ (IZCAS); • same locality as holotype, 14.VII.2025, Yuchen Zheng and Rui Wang, 1 ♂ 1 ♀ (CAU); • Kazakhstan: Alma-Ata (present Almaty), 1 ♀ (NHMW); • Kyrgyzstan: Narinskaya Obl. NE Naryn, Aral, 1500 m, 10.VII.2000, Vladimir Dolin, 1♂ (HUAC); • Kyrgyzstan: 5 km NE of Ugut, 41°25.561'N, 74°52.358'E, 1616 m, 09.07.2013, L. Ábrahám and S. Ilniczky, 1 ♂ 2 ♀♀ (SCM); • Kyrgyzstan: 2 km S of Kala, 41°26.669'N, 72°12.873'E, 789 m, 03.07.2013, L. Ábrahám and S. Ilniczky, 6 ♀♀ (SCM); • Kyrgyzstan: Jalal-Abad Region, ca 7 km N of Tash-Kömür, 41°25'N, 72°15'E, 660 m, 20–21.06.2003, R. Dobosz, 1 ♀ (Phot. 1) (USMB); • Tajikistan: Gorno-Badakhshan, S. side Saghirdasht Pass 10 km N. of Kala-i Khumb, 38°32'51.7"N, 70°48'1.6"E, 1850 m, T. and W. Garrevoet, 1 ♀ (RMNH).
Larval material.
China: Xinjiang, Shihezi, 5 km south of the Shihezi City, 686 m, 20.V.2025, Ziyang Ni (ZCAU).
Diagnostic characters.
Metafemur unmarked; forewing with acute apex, hyaline, without marking; hindwing subtriangular, posterior margin angled; hindwing marking generally as shade along crossveins, extending on RP; male ectoproct median branch basally wide, gradually bifurcating, subtriangular shape.
Description.
Size (mm, based on 2 specimens). Head + thorax length 6.9; antenna length 11.22–14.15; fore wing length 20.1–21.2; fore wing width 6.1–6.5; hindwing length 15–16; hindwing width 5.7–6.2.
Head. Vertex dark brown; setae dense, long, pale (Fig. 17C). Para-ocular band pale yellow. Frons yellow, medially with a dark brown spot; setae dense, long, pale. Clypeus and labrum yellow; setae long, pale (Fig. 17B). Mandible yellow, transitioning to dark brown apically. Labium and palp yellow. Antenna pedicel with dense pale and dark setae, flagellomeres generally dark brown; club basally dark brown, gradually transitioned to pale yellow to the tip (Fig. 17A).
Deleproctophylla tengri Zheng, Badano, H. Aspöck & Liu, sp. nov., adult, Xinjiang (China). A. Habitus, paratype female; B. Head, frontal view, holotype male; C. Head and thorax, dorsal view, holotype male; D. Head and thorax, lateral view, holotype male.
Thorax. Cervical sclerite yellow, with a pair of dark markings. Pronotum black, lateral margin yellow; setae long, pale. Mesonotum black with yellow markings: an anterior transversal pair, a median triangular pair and two lateral pairs; mesoscutellum mostly black, medially with a pair of yellowish markings, posterior margin yellow (Fig. 17C). Pleurae pale yellow (Fig. 17D).
Legs. Pro- and mesothoracic legs pale yellow, tarsi each generally pale yellow and distally dark brown; hind tarsus mostly dark brown. Coxae with thin pale setae; femora and tibiae with thin pale setae and ventral thicker blackish setae; tibial spurs narrow, blackish; tarsi with robust blackish setae (Fig. 17D).
Wings. Forewing long, apex acute. Pterostigma with three to four forked and unforked brown veinlets; membrane brown. Presectoral area with five to six crossveins. RP with four branches. MA gradually curving toward hind margin in distal portion. MP-CuA area with ~3 irregular rows of cells. Anal area hind margin straight; axillary lobe slightly pronounced. Forewing venation largely brown, paler toward wing base; Sc pale yellow, paler toward distal part; R pale yellow, darker toward distal part. Forewing membrane hyaline (Figs 3C, 3D, 17A). Hindwing subtriangular, broadest at RP fork; hind margin angular curved narrowing toward rounded apex. Presectoral area with two crossveins. RP with three branches. MA largely straight, slightly curved toward hind margin distally. MP-CuA area with ~2 irregular rows of cells. Hindwing venation brown, paler near wing base and distally. Hindwing membrane hyaline, hindwing marking generally as shade along crossveins, extending on RP (Figs 3C, 3D, 17A).
Abdomen. Tergites black, each with a caudal pair of yellowish markings. Sternites yellow. Pleural membrane brown with large paler markings.
Male genitalia. Ectoproct narrow, with branch in apical half, basally wide, gradually bifurcating, subtriangular shape. Setae of ectoproct long, dark brown; setae on branch limited to tip, dense, black, stout, curved, and spine-like; setae only on tip of ectoproct, black, stout, curved, and spine-like (Fig. 18A–C). Tergite 9 sub-rectangular in lateral view, with ventral projection; setae long and black, longer and denser on projection. Sternite 9 wide trapezoid in ventral view; setae thin, spine-like on median part (Fig. 18B, C). Gx11 arch-shaped, external tip rounded, slightly curved downward in lateral view; gp11 lightly sclerotized forming a shallow median furrow, with short setae; gx9 strongly sclerotized, with upward fold, widely hook-like in lateral view (Fig. 18F–J).
Deleproctophylla tengri Zheng, Badano, H. Aspöck & Liu, sp. nov., adult, Xinjiang (China). Male terminalia of holotype: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia of paratype: D. Ventral view; E. Lateral view. Male genitalia of holotype: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view.
Female genitalia. Ectoproct elongated; setae long, black. Tergite 9 sub-rectangular. Gx9 longer than wide; setae long. Tergite 8 subrectangular. Gx8 wider than long, with long setae. Gp8 flatted; setae thin (Fig. 18D, E).
Diagnosis of 3rd instar larva.
Dorsal head yellowish brown, with many dark brown spots, spots on the middle part relatively sparse, anteromedially with a dark brown marking. Dorsal segments 1–7 medially with a pair of short dark brown stripes, laterally scattered with irregular dense dark brown markings (Fig. 20).
Description of 3rd instar larva.
Size (mm). Body length (excluding mandible): 15.74; head length: 3.83; head width: 4.07; mandible length: 4.06.
Head. Wider then long, laterally with short setae (Fig. 21A, B). Clypeo-labrum generally brown, concave internally; anterior margin covered with some dolichasters. Dorsal side of head capsule yellowish brown, with many dark brown spots, spots on the middle part relatively sparse, anteromedially with a dark brown marking; head ventrally yellowish brown, with many short dolichasters, pore developed. Ocular tubercle prominent and large, apical part with a tiny projection. Antennae brown (Fig. 21A, B). Mandible nearly as long as head width; dorsal mandible basally pale brown, gradually darkening to the distal part; ventral mandible generally dark brown. Mandible basally with three or four pseudo-teeth and three interdental mandibular setae; second tooth largest and close to the third tooth; first tooth longer than third tooth; two pseudo-teeth between first and second teeth (Fig. 21A, B).
Thorax. Laterally with short setae. Pronotum generally pale pink, laterally pale yellow, scattered with some dark dots, posterolaterally with a pair of black markings, covered with tiny dark dolichaster. Mesothorax generally pale yellow, with spiracles present on short stout black sclerotized tubercle, lateral margin dark brown. Meso- and metathoracic setiferous processus scolus-like, distally constrict. Anterior mesothoracic setiferous processus longer than the posterior pair (Fig. 21C).
Legs. Pale yellowish brown, densely covered by short setae.
Abdomen. Pale yellow. Dorsal segments 1–7 medially with a pair of short dark brown stripes, laterally scattered with irregular dense dark brown markings. Each ventral segments 1–7 pale, laterally scattered with many dark spots, ventral segments 2–7 medially with a dark spot. Each segment 1–7 with a pair of dorsal scolus-like processus and a pair of ventral short scolus-like processus. Abdominal segment 9 pale yellow, long trapezoid, twice as long as wide, developed rastra with four pairs of digging setae (Fig. 21D).
Distribution.
Asia: China (Xinjiang); Kazakhstan (Almaty Region); Kyrgyzstan (Jalalabad and Narinsky Region); Tajikistan (Gorno-Badakhshan Autonomous Region) (Fig. 25).
Etymology.
The species name tengri is derived from the Turkic word “tengri,” meaning sky or heaven. The species was found in the surrounding area of the Tianshan Mountains (also known as the Tengri Tagh Mountains), a mountain range whose name, across various language families, signifies “Mountains of sky or heaven”. Thus, the name tengri reflects both the geographical origin of the species and the celestial meaning of the term in Turkic languages. The species epitheton is a substantive and an apposition to the genus.
Comparative notes.
Deleproctophylla tengri can be readily distinguished from the congeners by the median branch of the male ectoproct is subtriangular and distinctly broad at the base (Fig. 18A–C). The larva of D. tengri sp. nov. exhibits all the diagnostic larval features of the genus and differs only slightly from the known congeners D. australis and D. dusmeti (Figs 5, 20, 21). The main distinguishing feature of D. tengri compared to its western counterparts is the paler head capsule, which is distinctly mottled dark brown dorsally and paler ventrally (Fig. 21A, B). In contrast, the larvae of D. australis and D. dusmeti, which are virtually indistinguishable from each other, are characterized by a uniformly dark head capsule (Fig. 5).
Biology.
Deleproctophylla tengri inhabits arid hills (Fig. 19A, B). The larva of this new species was observed preying on a Parnassius apollonius Eversmann, 1847 (Lepidoptera: Papilionidae) on a hillside meadow (Fig. 19C).
Deleproctophylla tengri Zheng, Badano, H. Aspöck & Liu, sp. nov., live specimens, Xinjiang (China). A. Habitat, Mount Jiangjunshan (Shihezi); B. Female adult, paratype, Mount Jiangjunshan (Shihezi); C. Larva, feeding on a Parnassius apollonius (Eversmann, 1847). A, B. Photos by YZ; C. Photo by Ziyang Ni.
Deleproctophylla tengri Zheng, Badano, H. Aspöck & Liu, sp. nov., 3rd instar larva, paratype Xinjiang (China). A. Dorsal view; B. Ventral view; C. Lateral view.
Deleproctophylla tengri Zheng, Badano, H. Aspöck & Liu, sp. nov., 3rd instar larva, paratype Xinjiang (China). A. Head, dorsal view; B. Head, ventral view; C. Thorax, dorsal view; D. Apex of abdomen, ventral view. Abbreviations: 1t first tooth 2t second tooth 3t third tooth Pt pseudotooth Sp spiracle PtSP Prothoracic setiferous process Op odontoid process Rst rastra Ds digging setae.
Remarks.
This new species is endemic to the surrounding area of the Tianshan Mountains. We suspect that the specimens of D. variegata from Kyrgyzstan examined by Krivokhatsky et al. (2015) (without figure) are probably conspecific with this new species, considering that the record from Kyrgyzstan belongs to the same fauna as this new species (we also designated a paratype from Kyrgyzstan). Furthermore, Krivokhatsky et al. (2015) and Kerimova et al. (2023) also mentioned that D. variegata occurs in Uzbekistan and Kazakhstan, which might represent the same situation mentioned above. Nikolajev (2006) wrongly reported D. variegata from Takuma sands from the Ili River Valley (Almaty Region). The picture included in the publication confirms that the record most likely refers to D. tengri.
Deleproctophylla
variegata
Taxon classificationAnimaliaNeuropteraMyrmeleontidae
(Klug in Ehrenberg, 1834)
0BD298CE-074B-5AAE-8E44-58002D7EDDB0
Ascalaphus variegatus Klug in Ehrenberg, 1834: 36 (type locality: northern Lebanon; holotype in MfN). Hagen 1860: 53 (Ascalaphus). Hagen 1866: 387 (Ascalaphus). McLachlan 1873: 260 (Ascalaphus). van der Weele 1904: 98 (Theleproctophylla). van der Weele 1908: 287 (Deleproctophylla). Navás 1909: 54 (misidentification, Theleproctophylla). Navás 1911: 92 (misidentification, Theleproctophylla). Navás 1912a: 124 (misidentification, Theleproctophylla). Navás 1925: 31 (misidentification, Theleproctophylla). Alexandrov-Martynov 1926: 199 (probably misidentification of D. tengri, Ascalaphus). Scott 1929: 80 (Deleproctophylla). Navás 1929: 14 (misidentification, Theleproctophylla). H. Aspöck et al. 1980: 317 (Deleproctophylla). H. Aspöck and Hölzel 1996: 78 (Deleproctophylla). Sziráki 1998: 61 (Deleproctophylla). Sziráki 2000: 91 (Deleproctophylla). Santos 2000: 282 (Theleproctophylla). H. Aspöck et al. 2001: 302 (Deleproctophylla). Whittington 2002: 377 (Deleproctophylla). Koçak and Kemal 2002: 7 (Deleproctophylla). Canbulat 2007: 36 (Deleproctophylla). Dobosz and Ábrahám 2007: 19 (Deleproctophylla). Koçak and Kemal 2012: 17 (Deleproctophylla). Ilyina et al. 2013: 35 (Deleproctophylla). Krivokhatsky et al. 2015: 814 (probably misidentification of D. tengri, Deleproctophylla). Aistleitner 2019: 124 (Deleproctophylla). Monnerat and Ábrahám 2020: 144 (Deleproctophylla). Kerimova et al. 2023: 132 (Deleproctophylla). Poggi 2024: 10 (Deleproctophylla).
Examined material.
Type material. Lectotype • Smyrna Leonis col., 129, Type, Syria Ehren., variegatus Klug, variegatus Kl, Theleproctophylla variegata Kl. Soldaniky det. (MfN) (Fig. 24C, D).
Other specimens.
• Cyprus: Umg. v. Paphos ca 350 m, 1980 (27.V–8.VI), H. u. L. Hölzel, 27 ♂ 27 ♀ (NHMW); • Cyprus: 6 km Z v. Kyrenia, 300 m, 15.VI.1971, M. J. and J. P. Duffels, 1 ♀ (RMNH); • Cyprus: Larnika Salt Lake, 1–16.VI.1982, M. C. and G. Kruseman, 2 ex (RMNH); • Cyprus: Mamonia, N of Kouklia, 7–14 and 18.VI.1982, M. C. and G. Kruseman, 1 ♀ (RMNH); • Greece: Rhodes, Mount Profitis Ilias, 22.VII.2009, 1 ♂ 1 ♀ D. Badano, (DB); • Iran: Lorestan, Zagros Mts. near Borujerd, Vil. Vaenna’Ialt. 2400–3000 m, 29.VII.2014, E. Rutjan, 1 ♀ (DB); • Iran: Tehran, Qolhak, 1700 m, 1961.VI.1923, J. Klapperich, 1 ♂ 1 ♀ (HUAC); • Iran: N Iran, Abyek, Loc. No. 30, 24.06.[19]70, Exp. Nat. Mus. Praha, 1 ♂ (NMPC); • Iran: S Iran, 30 km E Kazerun, 1300 m, Loc. No. 229, 08–10.06.1973, Exp. Nat. Mus. Praha, 1 ♀ (NMPC); • Iran: S Iran, 28 km N Masiri, 1650 m, Loc. No. 236, 12.06.1973, Exp. Nat. Mus. Praha, 2 ♀♀ (NMPC); • Iran: Qolhak, B. Tehran, 1700 m, 09.06.1961, J. Klapperich, 1 ♂ (SCM); • Iran: Qolhak, B. Tehran, 1700 m, 23.06.1961, J. Klapperich, 1 ♀ (SCM); • Iran: Province Fars, Persepolis, Zagros Mts., 1200 m, 25–26.05.1999, Hácz-Kőszegi, 1 ♂ (SCM); • Iran: Province Lorestan, Ostoran Mt., 17 km E from Dorud, 2300–2700 m, 22.06.2000, Gy. Fábián, L. Szécsényi and K. Székely, 1 ♀ (SCM); • Iran: Province Hamadan, Nahavand 34°02.756'N, 48°22.614'E, 1851 m, 26.06.2005, L. Ábrahám, 4 ♂♂ 14 ♀♀ (SCM); • Iran: Province Fars, Kuh-e Kum Mts., Hasanabad vill. env., 2100 m, 27.05.2000, M. Kalabza, 1 ♀ (SCM); • Iran: E Téhrán, 2200 m, 18.05.2000, ?, 1 ♀ (SCM); • Iran: Province Hamadan, Nahavand 34°02.756'N, 48°22.614'E, 1851 m, 26.06.2005, L. Ábrahám, 1 ♀ (USMB); • Iran: Province Fars, S of Nüräbäd, 30°12'38"N, 51°32'02"E, 900 m, 29.04.2016, netting, R. Królik, 1 ♀ (USMB); • Israel: Tabgha 10 km N. v. Tiberias, 25.V.1967, C. A. W. Jeekel, 1 ♂ 2 ♀ (RMNH); • Jordan: Oberes Jordantal, 06.1999, G. Müller, 1 ♂ (SCM); • Syria: Damascus, Mts. Hermon, 1 km E Burqus, 33°28.261'N, 36°01.636'E, 1140 m, 12.06.2006, N. Rahmé, A. Kotán, A. Márkus, D. Szalóki and K. Székely, 1 ♀ (SCM); • Turkey: Anatolia orient., vil. Elâzıg˘, valico Gözeli, m 900, 24.VI.1968, V. Sbordoni, 1 ♂ (MNSG); • Turkey: Bingöl, 38°55'11.3"N, 40°20'8.3"E, T. and W. Garrevoet and N. Vandorpe, 1 ♀ (RMNH); • Turkey: Bitlis, Kuzkun Kiran Gecidi, 2100 m, 28.VII.1992, J. A. W. Lucas, 1 ♀ (RMNH); • Turkey: Hakkâri, Süvarihalil Geçidi, 1800–2400 m, 37°29'49.7"N, 43°22'19.8"E, 11.VII.2004, Dils J. and Faes J., 4 ♀ (RMNH); • Turkey: İçel (present Mersin), Güzeloluk, 950 m, 38°41'54"N, 34°09'52"E, 15.VI.1999, Dils J. and Faes J., 1 ♂ 3 ♀ (RMNH); • Turkey: Izmir, Foça, 50 m, 38°39'33"N, 26°49'16"E, 9.VI.1999, Dils J. and Faes J., 1 ♂ 1 ♀ (RMNH); • Turkey: Gevaş 1870 m, 38°16'31"N, 43°03'52"E, 16.VII.2006, Dils J. and Faes J., 2 ♀ (RMNH); • Turkey: Mut, VI–VII.1966, H.H.F. Hamann, 1 ♂ 1 ♀ (HUAC); • Turkey: Mut, 5–8.VI.1968, Josef Schmidt, 1 ♀ (HUAC); • Turkey: Mut, 8.VI.1968, Jos. Schmidt, 1 ♂ (HUAC); • Turkey: Mut, 12.VI.1965, Jos. Schmidt, 4 ♀ (HUAC); • Turkey: Urfa, 30–31.V.1978, J. Schmidt, 1 ♂ 1 ♀ (HUAC); • Turkey: Urfa, 10.VI.1988, J. Schmidt 1 ♂ 1 ♀ (HUAC); • Turkey: Province Agri, 7 km W of Aydintepe, 39°49'N, 42°30'E, 1800 m, 05–06.08.1988, Gyulai, Hreblay, Ronkay and Ronkay, 2 ♂♂ (SCM); • Turkey: Kusgunkiran Gecidi, 12.07.1989, ?, 1 ♀ (SCM); • Turkey: mer., village Icel Orta Toroslar, Hocali, 500 m, 03–04.06.2004, Z. Rahmé, L. Nádai and K. Székely, 2 ♀♀ (SCM); • Turkey: Province Mersin, 10 km E of Aydincik, 37°08'N, 36°34'E, in meadows, 01.06.2001, R. Dobosz, 3 ♂♂ 4 ♀♀ (USMB); • Turkey: Province Mersin, 10 km E of Aydincik, 37°08'N, 36°34'E, in meadows, 01.06.2001, J. Kurzawa, 1 ♂ 2 ♀♀ (USMB); • Turkey: Province Mersin, 3 km N of Erdemli, 36°37'N, 34°19'E, oak woods, 350 m, A. Lasoń, 1 ♀ (USMB); • Turkey: Province Mersin, 3 km N of Erdemli, Taşeli Platosu, 36°37'N, 34°15'E, 05.06.2011, A. Lasoń, 1 ♂ (USMB); • Turkey: Province Şanliurfa, Yeni Halfeti, 37°14'N, 37°53'E, 08.06.2011, A. Lasoń, 1 ♂ (USMB); • Turkey: Province Şanliurfa, Yeni Halfeti, 37°14'N, 37°53'E, 08.06.2011, R. Królik, 1 ♂ 1 ♀ (USMB); • Turkmenistan: Turkmenia SW, КАЛА-Kala, V.1993, [S.] Naglis, 1 ♂ (ex coll. Gallo, 2000), det. Badano, 2012 (MSNG).
Diagnostic characters.
Forewing unmarked; hindwing posterior margin curved, convex; hindwing with faint marking below pterostigma, usually just a shade around crossveins; male ectoproct with branch stout, in apical third, well after mid-length, after branch ectoproct narrowing and progressively broadening caudally; setae on branch for its entire length; setae on branch and tip of ectoproct long and thin.
Redescription.
Size (mm, based on 5 specimens). Head + thorax length 6.01–7.04; antenna length 10.83–12.78; fore wing length 16.30–17.87; fore wing width 4.14–5.38; hindwing length 10.61–13.02; hindwing width 3.84–4.78.
Head. Vertex yellowish brown; setae dense, long, pale yellow (Fig. 22C). Para-ocular band yellow. Frons yellowish brown; setae dense, long, pale yellow. Clypeus and labrum yellowish brown; setae long, pale. Mandible yellowish brown, darker apically. Labium and palp yellowish brown, of the same color of mandible (Fig. 22B). Antenna with pedicel with dense pale and dark setae, flagellomeres brown, nodes slightly darker; club brown, paler anteriorly (Fig. 22A).
Deleproctophylla variegata (Klug, 1834), Cyprus. A. Habitus, female; B. Head, frontal view, male; C. Head and thorax, dorsal view, male; D. Head and thorax, lateral view, male.
Thorax. Cervical sclerite yellowish brown. Pronotum brown; setae long, pale yellow. Mesonotum brown with yellowish brown markings: an anterior stripe-like pair, a median triangular pair, and a posterior rounded pair; mesoscutellum dark brown, with yellowish brown posterior margins. Metanotum yellowish brown, with dark brown median marking; setae pale yellow (Fig. 22B). Pleurae yellowish; setae pale yellow (Fig. 22D).
Legs. Pro- and mesothoracic legs entirely yellowish brown. Metathoracic leg yellowish brown, usually unmarked, occasionally with a faint brown marking. Setae of coxae pale yellow setae of femora and tibiae thin and pale yellow dorsally, thicker and blackish ventrally; tibial spurs narrow; setae of tarsi blackish (Fig. 22D).
Wings. Forewing long, apex rounded. Pterostigma with ~4 unforked brown veinlets; membrane brown. Presectoral area with ~6 crossveins. RP with four branches. MA gradually curving toward hind margin in distal portion. MP-CuA area with ~3 irregular rows of cells. Anal area hind margin straight; axillary lobe slightly prominent. Forewing venation brown, paler toward wing base and at tip; Sc and R yellowish. Forewing membrane hyaline, unmarked. Hindwing broad, wider at RP fork; hind margin markedly convex, with a pronounced curve. Presectoral area with three crossveins. RP with three branches. MA largely straight, slightly curved toward hind margin distally. MP-CuA area with ~2 irregular rows of cells. Hindwing venation brown, paler near wing base; Sc yellowish (Fig. 3E, F, 22A). Hindwing membrane hyaline, distal section with an inconspicuous brown marking, as a shade bordering crossveins from pterostigma to first branch of RP (Fig. 3E, F, 22A).
Abdomen. Tergites pale brown, each with a caudal pair of yellowish markings. Sternites pale brown, paler than tergites. Pleural membrane pale brown.
Male genitalia. Ectoproct with branch bifurcating at two-third of length; ectoproct abruptly narrowing after branching and gradually expanding caudally. Setae of ectoproct long, dark brown; setae of branch arranged on entire length, sparse and gradually denser toward apex, black, long, and slightly curved; setae on tip of ectoproct, black, long, and slightly curved, denser apically (Fig. 23A–C). Tergite 9 sub-rectangular in lateral view, with large ventral thumb-like projection; setae long and black, longer and denser on projection. Sternite 9 with three lobes, lateral pair slightly prominent, median pair large; setae thin, thicker on median lobe (Fig. 23A–C). Gx11 slightly curved downward; gp11 as wide as gx9, forming a furrow, with short setae; gx9 set apart, relatively broad, almost subrectangular, strongly sclerotized, folded downward (Fig. 23F–J).
Deleproctophylla variegata (Klug in Ehrenberg, 1834), Cyprus. Male terminalia: A. Dorsal view; B. Ventral view; C. Lateral view. Female terminalia: D. Ventral view, E. Lateral view. Male genitalia: F. Ventral view; G. Anteroventral view; H. Caudal view; I. Lateral view; J. Dorsal view.
Types of Deleproctophylla spp. A, B. Theleproctophylla dusmeti Navás, 1914, paralectotype in MNHN; photo by YZ; A. Habitus; B. Labels; C, D. Ascalaphus variegatus Klug, 1834, holotype in MfN, photo by Hongyu Li; C. Habitus; D. Labels.
Female genitalia. Ectoproct elongated, with ventral projection, setae long, black. Tergite 9 sub-rectangular. Gx9 wider than long, wider than tergite; setae long. Tergite 8 subrectangular. Gx8 wider than long, with long setae. Gp8 ventrally prominent; setae thin (Fig. 23D, E).
Comparative notes.
Deleproctophylla variegata differs from most species of Deleproctophylla due to the highly reduced hindwing pigmentation, which is often limited to a slight shading around the crossveins in the radial area (Fig. 3E, F). Deleproctophylla variegata is easily distinguishable by the male ectoproct with branch arising at two-third of length—the most apical position among Deleproctophylla species (Fig. 23A–C). Additionally, the caudal narrowing of ectoprocts after the branch is unique to this species.
Distribution.
Cyprus, Greece (Aegean and Dodecanese islands), Iran, Iraq, Israel, Jordan, Kazakhstan?, Kyrgyzstan?, Lebanon, Russia? Turkey, Turkmenistan (NEW RECORD), Uzbekistan (H. Aspöck et al. 2001; Monnerat and Ábrahám 2020; Oswald 2025) (Fig. 25).
Distribution map of Deleproctophylla spp.
Remarks.
Deleproctophylla variegata is a species with a wide distribution (Oswald 2025), but it is likely that some old records belong to other species (see D. tengri). The specimen reported as D. variegata from Azerbaijan by Kerimova et al. (2022) is D. australis, as shown by molecular data (Fig. 1) and confirmed by examination of the voucher photograph by RD. Records of D. variegata from Kazakhstan, Kyrgyzstan, and Russia are also likely misidentifications and require verification.
Discussion
The two diurnal owlfly genera, Deleproctophylla and Libelloides, are characteristic elements of the warmer regions of the Palaearctic, with their southern distribution limits largely coinciding with the biogeographic boundaries of the region itself. Compared with Libelloides, Deleproctophylla is less diverse, more southerly distributed, and more strictly associated with arid habitats. Traditionally, these genera have been considered closely related based on their shared diurnal activity, the presence of large wing markings, the sub-triangular shape of the hindwings, and the presence of prominent projections on the male ectoprocts (van der Weele 1908). Based on these characters, van der Weele (1908) established the tribe Ascalaphini to include a few genera with these features, namely Libelloides (then Ascalaphus), Deleproctophylla, Puer Lefèbvre, 1842, and Ascalaphodes McLachlan, 1871 (the only one with unmarked wings), a classification later followed by Navás (1912a). Later, Pantaleoni and Loru (2018) renamed this tribe Libelloidini Pantaleoni & Loru, 2018. The enigmatic genus Puer, while resembling these genera in its heavily marked wings, differs significantly in having a less prominent male ectoproct, a distinct male genital conformation, and a remarkably different larval morphology, suggesting distant relationship to the other Palaearctic owlflies (U. Aspöck and H. Aspöck 1987; Badano and Pantaleoni 2012). Conversely, the poorly known genus Parascalaphus Alexandrova-Martynova, 1926, is likely related to Libelloides. Most phylogenetic studies on owlflies have focused on resolving their affinity within Neuroptera and Myrmeleontidae, with relatively few studies investigating their internal relationships. The only comprehensive phylogenetic analysis of owlflies, conducted by Jones (2019), did not recover a close relationship between Libelloides and Deleproctophylla, instead placing the latter as sister to Ascalohybris Sziraki, 1998. However, larval morphology supports the traditional view because Libelloides and Deleproctophylla share a highly similar larval form, differing only in minor traits, whereas Ascalohybris exhibits substantial differences in body shape, chaetotaxy, and the shape and arrangement of setiferous processus (Badano and Pantaleoni 2014). Further research is therefore needed to clarify the internal relationships within Ascalaphinae and their position within Myrmeleontidae. The striking appearance of most owlflies has historically led to poor taxonomic practices relying on superficial wing pattern characters, resulting in taxonomic inflation and misidentifications (e.g., van der Weele 1908; Navás 1912a). Deleproctophylla exemplifies this issue, as many species were historically distinguished based on wing markings alone, leading to widespread confusion (Monserrat et al. 2014; Monnerat and Ábrahám 2020; Japaridze et al. 2024). However, the present morphological comparisons combined with barcode-based species delimitation confirm that these characters, albeit visually prominent, are highly variable and unreliable for species identification. Both species delimitation methods employed in this study, ASAP and mPTP, corroborated the morphological taxonomy, identifying five distinct species: D. australis, D. bleusei, D. dusmeti, D. variegata, and the newly described D. tengri (Fig. 1). Deleproctophylla australis and D. dusmeti exhibit considerable overlap in wing pattern, identical larval morphology, and geographic proximity, but morphological traits and COI data support their distinction (Fig. 1). The most reliable diagnostic feature between D. australis and D. dusmeti is the shape of the male ectoproct. The remaining analyzed species were only available as single individuals. In all cases, pairwise genetic distances between the specimens exceeded the 3% distance threshold, which appears effective for delimit the species of Myrmeleontidae (Ascenzi et al. 2025). One unexpected result was the identification of a potential mislabeled GenBank sequence, a finding previously noted by Japaridze et al. (2024). Recently collected specimens suitable for DNA extraction were unavailable D. dandizenor sp. nov., the latter of which was collected in a region currently inaccessible due to geopolitical turmoil. Nevertheless, this species is morphologically highly distinct from all other members of Deleproctophylla, making misidentification unlikely.
Supplementary Material
XML Treatment for Deleproctophylla
XML Treatment for Deleproctophylla australis
XML Treatment for Deleproctophylla bleusei
XML Treatment for Deleproctophylla dandizenor
XML Treatment for Deleproctophylla dusmeti
XML Treatment for Deleproctophylla gelini
XML Treatment for Deleproctophylla tengri
XML Treatment for Deleproctophylla variegata
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ábrahám L (2017) New data to the Moroccan Myrmeleontiformia (Nemopteridae, Myrmeleontidae, Ascalaphidae) fauna. Natura Somogyiensis 30(30): 75–138. 10.24394/Nat Som.2017.30.75 · doi ↗
- 2Ábrahám L, Mészáros Z (2004) Ascalaphid studies III. Remarks on the morphology of Deleproctophylla gelini (Navás, 1919) (Neuroptera: Ascalaphidae). Somogyi Múzeumok Közleményei 16: 333–336.
- 3Aistleitner E (2019) Schmetterlingshafte der Westpalaearktis – faunistische und phaenologische Aufzeichnungen und Anmerkungen (Neuropterida, Planipennia, Ascalaphidae). Mitteilungen der Münchener Entomologischen Gesellschaft 109: 117–137.
- 4Aistleitner E, Lencina GF (2020) On the knowledge of the lacewing fauna in the Spanish province of Albacete (I) (Neuroptera, Nemopteridae, Ascalaphidae). Mitteilungen des Internationalen Entomologische Verein 43: 97–116.
- 5Ascenzi A, Nania D, Cristiano A, Badano D, Pacifici M, Cerretti P (2025) Neglected predatory insects trigger potential Key Biodiversity Areas in threatened coastal habitats. Biodiversity and Conservation 34(5): 1671–1688. 10.1007/s 10531-025-03035-8 · doi ↗
- 6Aspöck U, Aspöck H (1987) Wiederentdeckung von Puer maculatus (Olivier) in Europa (Neuropteroidea: Planipennia: Ascalaphidae). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen 39: 5–11.
- 7Aspöck U, Aspöck H (2008) Phylogenetic relevance of the genital sclerites of Neuropterida (Insecta: Holometabola). Systematic Entomology 33(1): 97–127. 10.1111/j.1365-3113.2007.00396.x · doi ↗
- 8Aspöck H, Hölzel H (1996) The Neuropteroidea of North Africa, Mediterranean Asia and of Europe: a comparative review (Insecta). In: Canard M, Aspöck H, Mansell MW (Еds) Pure and Applied Research in Neuropterology, 31–86. Proceedings of the Fifth International Symposium on Neuropterology. Meeting: 2–6 May 1994, Cairo, Egypt. Privately printed, Toulouse, France, 341 pp.