Preventive Effects of Probiotic and Postbiotic Lacticaseibacillus paracasei HY2782 on DSS-induced Colitis in Mice: Comparable Efficacy of Live and Heat-Killed Forms
Daehyeop Lee, Hyeonjun Gwon, Ji-Woong Jeong, Joo-Yun Kim, Jae-Jung Shim, Jae-Hwan Lee

TL;DR
This study shows that both live and heat-killed forms of Lacticaseibacillus paracasei HY2782 can prevent colitis in mice, suggesting they are equally effective for treating intestinal inflammation.
Contribution
The study demonstrates that both probiotic and postbiotic forms of HY2782 are comparably effective in preventing DSS-induced colitis in mice.
Findings
Both live and heat-killed HY2782 reduced disease activity and preserved colon length in DSS-induced colitis.
HY2782 treatment suppressed pro-inflammatory cytokines and restored tight junction-related gene expression.
Gut microbiota changes were limited and similar between live and heat-killed HY2782 forms.
Abstract
Ulcerative colitis is chronic inflammatory bowel disease characterized by intestinal inflammation and barrier dysfunction. Probiotics and postbiotics have been proposed as dietary interventions for intestinal health; however, their comparative preventive effects remain unclear. In this study, we evaluated the preventive effects of probiotic and postbiotic forms of Lacticaseibacillus paracasei HY2782 in a dextran sulfate sodium (DSS)-induced colitis mouse model. Male C57BL/6 mice were orally administered live or heat-killed HY2782 prior to DSS exposure. Disease activity index, colon length, histopathological damage, inflammatory cytokine expression, and intestinal barrier-related gene expression were assessed, and gut microbiota composition was analyzed using 16S rRNA gene sequencing. Both probiotic and postbiotic forms of HY2782 significantly alleviated DSS-induced colitis, as evidenced…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
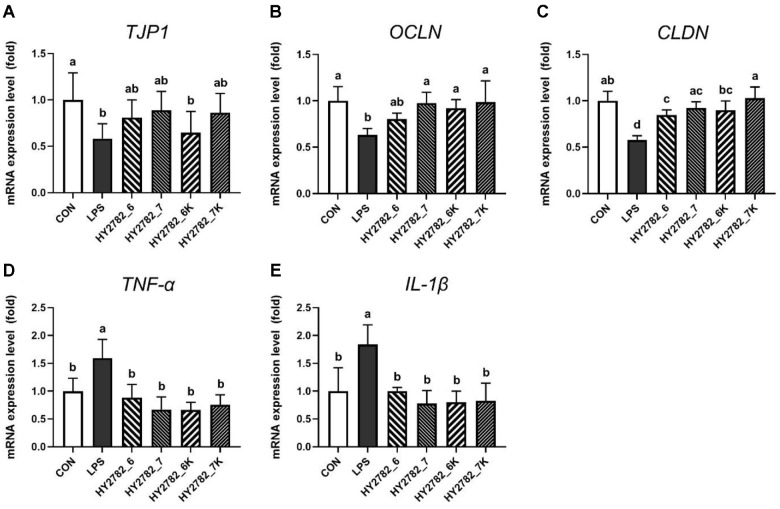 Figure 1
Figure 1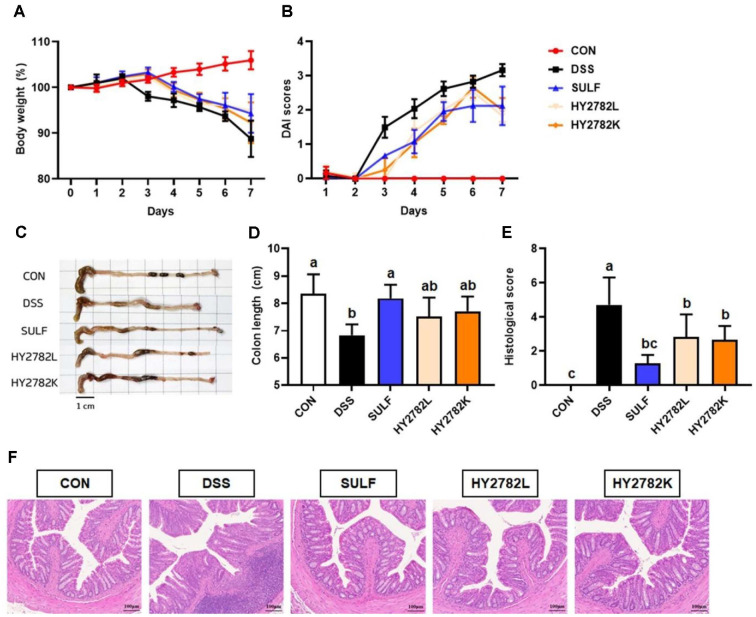 Figure 2
Figure 2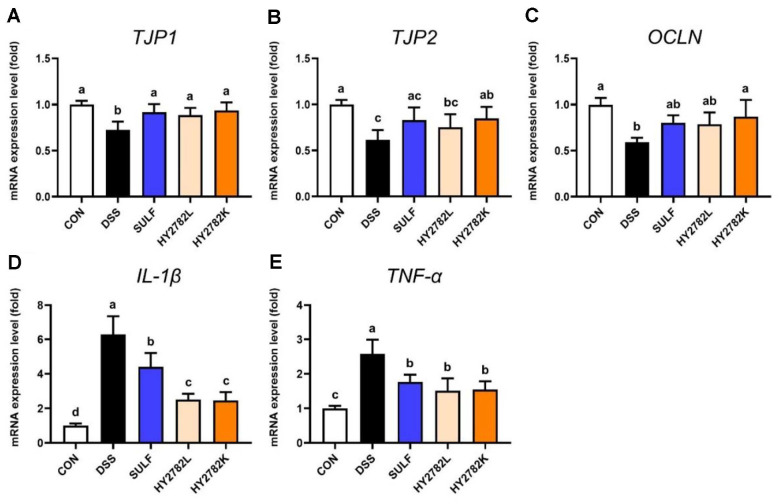 Figure 3
Figure 3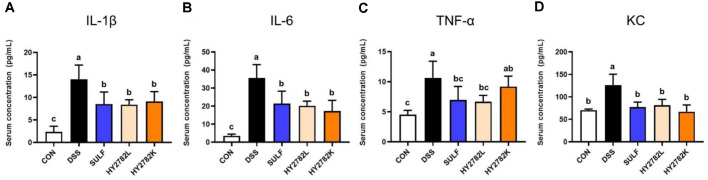 Figure 4
Figure 4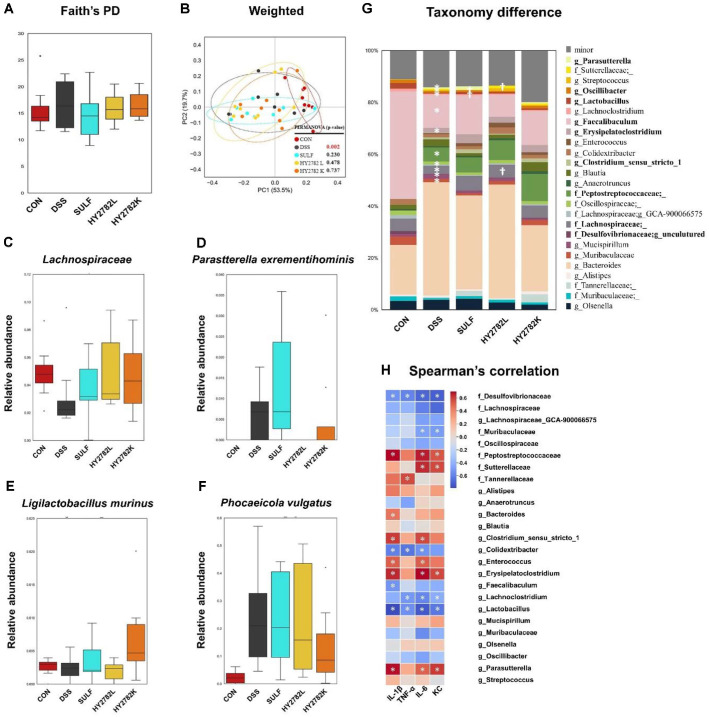 Figure 5
Figure 5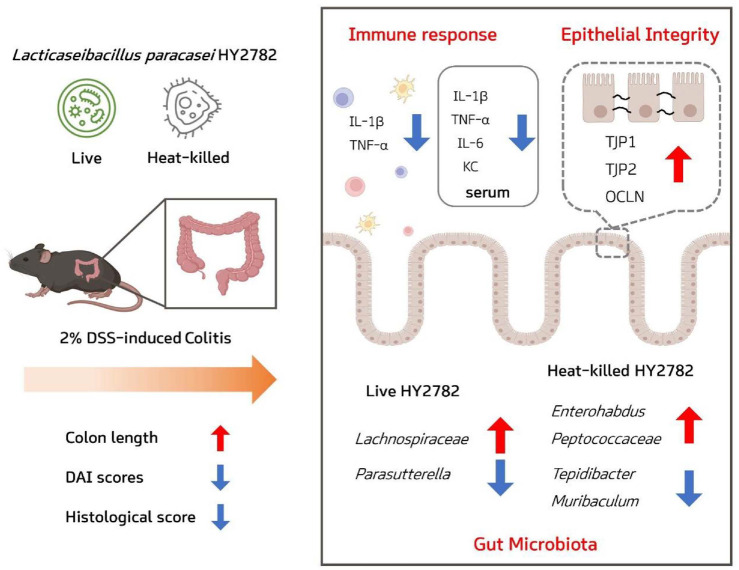 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Probiotics and Fermented Foods · Inflammatory Bowel Disease
Introduction
Inflammatory bowel disease (IBD), which includes Crohn’s disease (CD) and ulcerative colitis (UC), is a group of chronic gastrointestinal disorders characterized by inflammation, chronic diarrhea, abdominal pain, and rectal bleeding [1-3]. The incidence and prevalence of UC are rising globally; its precise pathogenesis remains incompletely defined [4]. Increasing evidence suggests that UC develops through a multifactorial interplay involving genetic susceptibility, environmental exposures, microbial infections, immune dysregulation, and gut microbiota alterations [5]. Dysbiosis, or disruption of the intestinal microbial community, is particularly implicated in UC onset, as it impairs the epithelial barrier, promotes mucosal immune activation, and drives pro-inflammatory cytokine production. This compromise in epithelial integrity increases intestinal permeability, accelerating the progression of the disease. In this context, tight junction proteins (TJPs), which are essential for maintaining intestinal epithelial barrier function, play a key role in UC pathophysiology [6]. Collectively, these interrelated factors support the view that UC is an immune-mediated inflammatory disorder driven by disturbances within the intestinal ecosystem [7].
Current therapeutic agents, including corticosteroids, 5-aminosalicylic acid (5-ACA) derivatives, immunomodulators, and biologics, aim to reduce inflammation for managing IBD. However, these treatments are frequently associated with notable adverse effects, ranging from nephrotoxicity and drug hypersensitivity to gastrointestinal symptoms such as abdominal pain, nausea, vomiting, and exacerbation of diarrhea [8, 9]. Moreover, several therapeutic challenges remain, including primary unresponsiveness or secondary loss of response to treatment (e.g., patients ceasing to respond after an initial benefit) and inability to address the underlying microbiota imbalance [10]. These limitations have redirected attention toward interventions, particularly those capable of modulating mucosal barrier integrity and restoring gut microbiota homeostasis. Within this therapeutic framework, probiotics and postbiotics have emerged as potential modalities to reinforce epithelial barrier function, regulate mucosal immune activity, and restore microbial balance.
Probiotics, defined as live microorganisms that confer health benefits to the host, modulate the immune system and enhance the epithelial barrier function through microbial signaling, metabolite production, and host-microbe interaction [11, 12]. Postbiotics, by contrast, are non-viable microbial cells or components derived from probiotics, exerting similar beneficial effects, including immunomodulation and mucosal barrier reinforcement [13, 14]. Given the pathological hallmarks of UC, including mucosal inflammation, impaired barrier integrity, and dysbiosis, both probiotics and postbiotics represent distinctive therapeutic approaches. Probiotics achieve their effects through viability-associated functions like colonization, metabolic activity, and direct immune modulation, while postbiotics provide increased stability and safety by acting independently of microbial viability [15-17]. The therapeutic efficacy of these two forms may vary based on the disease context, with probiotics offering sustained effects through microbial ecosystem modulation, and postbiotics providing reproducible, safer outcomes by reducing inflammation and restoring epithelial integrity [18, 19]. Understanding the contribution of probiotics and postbiotics is essential for optimizing UC treatment strategies and exploring optimal personalized therapeutic approaches [20].
In this study, we investigated the preventive potential of Lacticaseibacillus paracasei HY2782, in both its probiotic (live) and postbiotic (heat-killed) forms, against DSS-induced colitis. By focusing on the reinforcement of the intestinal epithelial barrier and the modulation of gut microbiota, we sought to elucidate the distinct and shared mechanisms by which these two bacterial formulations mitigate colonic inflammation.
Materials and Methods
Preparation of Live and Heat-Killed Probiotics
L. paracasei HY2782 (HY2782) was cultured in Man-Rogosa-Sharp broth (Difco Laboratories, USA) at 37°C for 24 h. After incubation, bacterial cells were harvested by centrifugation at 3,000 ×g for 20 min. The supernatants were discarded, and the bacterial pellet was washed twice with sterile phosphate-buffered saline (PBS) and resuspended in PBS to a concentration of 1 × 10^9^ CFU/ml. For heat-killed probiotics preparation, the suspension was subjected to boiling for 20 min in a water bath with gentle mixing. The complete loss of viability was confirmed by observing no visible colonies on MRS agar plates. For heat-killed preparations, bacterial concentrations were expressed as CFU equivalents based on viable counts measured prior to heat treatment. The same bacterial suspensions were subsequently heat-inactivated and used at equivalent cell densities.
Cell Culture Conditions and Samples Treatment
RAW 264.7 cells and Caco-2 cells were obtained from the American Type Culture Collection (ATCC; USA) and the Korean Cell Line Bank (KCLB; Republic of Korea), respectively. RAW 264.7 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin, and the growth medium was changed every 2-3 days. Caco-2 cells were maintained in DMEM containing 10% FBS and 1% antibiotic-antimycotic. Caco-2 cells were differentiated for 21 days, with the culture medium replaced every 2-3 days. All cells were incubated at 37°C in a humidified atmosphere containing 5% CO_2_.
After reaching sufficient confluency, cells were treated with 10^6^ and 10^7^ CFU/ml of live HY2782 (HY2782_6 and HY2782_7, respectively) or with heat-killed HY2782 at equivalent concentrations (HY2782_6K and HY2782_7K, respectively) for 24 h. Subsequently, lipopolysaccharide (LPS, 1 μg/ml) was added, and cells were incubated for an additional 24 h.
Gene Expression Analysis by qRT-PCR in RAW 264.7 and Caco-2 Cells
Total RNA was extracted using the Easy-spin Total RNA Extraction Kit (iNtRON Biotechnology, Republic of Korea). Complementary DNA (cDNA) was synthesized from 2 μg of total RNA using the Omniscript Reverse Transcription Kit (Qiagen, Germany). qRT-PCR was conducted using TaqMan Gene Expression Assays (Applied Biosystems, USA).
For RAW 264.7 cells, the following targets were analyzed: tumor necrosis factor (TNF-α, Mm00443258_m1), interleukin 1 beta (IL-1β, Mm00434228_m1), interleukin 6 (IL-6, Mm00446190_m1), nuclear factor of kappa B (NF-κB, Mm00476361_m1), and toll-like receptor 4 (TLR4, Mm00445273_m1). For Caco-2 cells, the target genes included tight junction protein 1 (TJP1, Hs01551871_m1), occludin (OCLN, Hs05465837_g1), claudin 1 (CLDN, Hs00221623_m1), TNF-α (Hs00174128_m1), and IL-1β (Hs01555410_m1).
Relative mRNA expression levels were calculated using the 2^-ΔΔCT^ method. Gene expression in RAW 264.7 cells was normalized to GAPDH (Mm99999915_g1), and expression in Caco-2 cells was normalized to GAPDH (Hs02786624_g1) expression.
Animals Study Design
Male C57BL/6 mice (6 weeks old) were purchased from Dooyeol Biotech (Republic of Korea) and acclimated for one week prior to experiments. The mice were housed under conditions at a temperature of 20-22°C with 40-60% humidity, a 12 h light/dark cycle, and free access to standard rodent diet and water. All animal studies were approved by the Institutional Animal Care and Use Committee (IACUC) of hy Co. Ltd. (approval number, AEC-2025-0002-Y).
After acclimation, the mice were randomly assigned into five groups (n = 8 per group): CON, normal group; DSS, 2% DSS-induced colitis group; SULF, 2% DSS + sulfasalazine (100 mg/kg/day, positive control); HY2782L, 2% DSS + live HY2782 probiotic (1 × 10^9^ CFU/ml); HY2782K, 2% DSS + heat-killed HY2782 postbiotic (1 × 10^9^ CFU/ml). Mice were orally administered sulfasalazine or each sample at a fixed volume of 200 μl daily. The CON and DSS groups administered an equivalent volume of PBS for the same duration.
Mice were orally administered HY2782 for a total of 12 days. Mice were first administered samples for 7 days. After this pre-treatment period, drinking water in all groups except the CON group was replaced with 2% DSS (MP Bio-medicals, USA; molecular weight 36,000-50,000 Da) for 5 days to induce colitis. Throughout both the pre-treatment and DSS induction periods, mice continued to be orally administered sulfasalazine or both forms of HY2782, followed by 2 days for recovery.
During the experimental period, body weight, food intake, and water consumption were recorded daily. On the final day of the experiment, mice were euthanized using CO_2_ gas, and blood, colon, and cecum tissues were collected for further analyses.
Determination of Disease Activity Index (DAI) Scores
The DAI was evaluated based on the mean of three parameters: body weight loss, stool consistency, and hematochezia. Body weight loss was scored on a scale of 0-4 as follows: 0, no change; 1, 0-5% loss; 2, 5-10% loss; 3, 10-15% loss; and 4, >15% loss. Stool consistency was scored from 0 to 3, where 0 indicated normal stool, 2 indicated loose or soft stool, and 3 indicated watery diarrhea. The degree of hematochezia was scored from 0 to 4, with 0 representing no detectable blood, 2 indicating the presence of occult or visible traces of blood in the stool, and 4 indicating gross rectal bleeding visually observed.
Histological Analysis Using Hematoxylin and Eosin (H&E) Staining
Colon tissues were fixed in 10% formalin (Sigma Aldrich, USA) at room temperature for 24 h. After that, the tissues were embedded in paraffin and sectioned. The paraffin sections were stained with hematoxylin and eosin (H&E). The images were obtained using a Zeiss Axiovert 100 M microscope (Carl Zeiss AG, USA). Histological damage was evaluated using a composite scoring system. Inflammatory cell infiltration, crypt damage, and mucosal erosion were each scored individually on a scale of 0 to 3 according to severity, and the individual scores were summed to obtain a total histological score for each sample (DooYeol Biotech).
Gene Expression Analysis by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) in Colon Tissues
Total RNA from distal colon tissues was extracted, and cDNA was synthesized as mentioned above. The target genes analyzed in this study were as follows: TJP1 (Mm01320638_m1), TJP2 (Mm00495620_m1), occludin (Mm00500910_m1), IL-1β (Mm00434228_m1), and TNF-α (Mm00443258_m1). The relative mRNA expression levels were calculated using the 2^-ΔΔCT^ method, and all data were normalized to GAPDH (Mm99999915_g1) expression.
Measurement of Pro-Inflammatory Cytokines in Serum
Blood samples were centrifuged at 3,000 ×g for 20 min at 4°C to obtain serum. The concentration of pro-inflammatory cytokines, including TNF-α, IL-1β, IL-6, and KC, was measured using the Mouse High Sensitivity T Cell Magnetic Bead Panel (Merck Millipore, USA). The cytokine measurements were performed using the Luminex Multiplex Assay system (Thermo Fisher Scientific, USA) at LABISKOMA (Republic of Korea).
DNA Extraction and 16S rRNA Gene Sequencing
Total DNA was extracted from the cecum, and the composition of the microbiota was analyzed through 16S rDNA sequencing using a next-generation sequencing (NGS) platform (Illumina, USA). The universal primer sequences employed for sequencing were as follows: V3-F (5’-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACG GGNGGCWGCAG-3') and V4-R (5'-CTCGTGGGCTCGGAGATGTGT ATAAGAGACAGG ACTACHVGGTATCTAATCC-3'). DNA extraction and 16S rDNA sequencing were conducted at Macrogen (Republic of Korea) using an Illumina MiSeq i100 Plus instrument (Illumina).
Analysis of 16S rRNA Gene Sequencing Data
Amplicon sequencing data were processed with QIIME2 (version 2023.9). Raw sequences were first demultiplexed using the q2-demux plugin, followed by filtering and denoising of low-quality sequences with the DAD2 plugin to generate amplicon sequence variants (ASVs). The ASVs were aligned using MAFFT, and a rooted phylogenetic tree was constructed with FastTree 2 for subsequent phylogenetic analysis. Taxonomic assignment was carried out with the q2-feature-classifier plugin, utilizing the pre-trained SILVA 138 reference database (99% identity threshold). Alpha diversity metrics were computed to assess within-sample diversity using Faith’s Phylogenetic Diversity (Faith’s PD). The Kruskal-Wallis test was employed to evaluate group differences in alpha diversity. Beta diversity was measured weighted UniFrac distance matrices, with results visualized via Principal Coordinates Analysis (PCoA). Group Differences in microbial community composition were analyzed using Permutational Multivariate Analysis of Variance (PERMANOVA). Taxonomic composition was assessed by comparing the relative abundances of microbial taxa across groups. Taxa with a relative abundance of ≥ 1% in at least one group were categorized as major taxa, while others were grouped as minor taxa. Differentially abundant taxa were identified using LEfSe (Linear Discriminant Analysis Effect Size) with a significance threshold of LDA score > 2.0. Spearman’s rank correlation coefficient was used to examine correlations between gut microbiota relative abundance and biochemical markers, with analyses performed in R (Version 3.6.6). All sequencing data are available in the NCBI Sequence Read Archive under the accession number PRJNA1308642.
Statistical Analysis
All data were expressed as mean ± standard deviation (SD). Differences among multiple groups were analyzed using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. Statistical analyses were performed using GraphPad Prism version 6 (GraphPad Software, USA), and p < 0.05 was considered statistically significant.
Results
Before evaluating the in vivo efficacy, the anti-inflammatory potential of HY2782 was confirmed in RAW 264.7 macrophages (Fig. S1). Subsequently, we assessed whether these protective effects extended to the intestinal epithelial barrier using Caco-2 cells.
Effects of Live and Heat-Killed HY2782 on Gene Expression in Caco-2 Cells
The effects of live and heat-killed HY2782 on tight junction and inflammation-related genes were evaluated in the LPS-induced Caco-2 cell model. Treatment with LPS significantly decreased the expression of tight junction genes, TJP1, OCLN, and CLDN, by 0.58-, 0.63-, and 0.58-fold, respectively, compared to the control. The expression of these three tight junction genes was dose-dependently restored following treatment with both live and heat-killed HY2782. Specifically, the expression of the TJP1 gene was significantly restored in the HY2782_6 group (0.81-fold) and the HY2782_7 group (0.89-fold). Treatment with two different doses of heat-killed HY2782 also upregulated TJP1 expression, however, there was no significant difference in the HY2782_6K group. The LPS-induced downregulation of the OCLN gene was significantly restored following treatment with both live and heat-killed HY2782. Low-dose and high-dose treatments with live HY2782 resulted in a 0.80- and 0.97-fold increase in expression, respectively, while treatment with heat-killed HY2782 led to a 0.92- and 0.99-fold mRNA expression. Similarly, the expression of CLDN was also upregulated, with significant increases observed in the HY2782_6 group (0.85-fold), HY2782_7 group (0.92-fold), HY2782_6K group (0.90-fold), and HY2782_7K group (1.03-fold) (Fig. 1A-1C).
In addition, the effects of live and heat-killed HY2782 on the expression of pro-inflammatory cytokines TNF-α and IL-1β were evaluated in the LPS-induced Caco-2 cell model. Treatment with LPS significantly increased the expression of both TNF-α and IL-1β to 1.59- and 1.84-fold of the control, respectively. However, treatment with both live and heat-killed HY2782 led to restoration of expression across all groups. In the HY2782_6 group, TNF-α expression was restored to 0.88-fold, and IL-1β expression was restored to 1.00-fold, nearly returning to the control levels. The HY2782_7 group showed a more modest recovery, with TNF-α expression restored to 0.67-fold and IL-1β expression restored to 0.78-fold. Likewise, treatment with heat-killed HY2782 resulted in restoration of gene expression. In the HY2782_6K group, TNF-α and IL-1β expression were restored to 0.66-fold and 0.80-fold, respectively. And the expression of TNF-α and IL-1β was significantly downregulated to 0.75- and 0.83-fold, respectively, in the HY2782_7K group (Fig. 1D and 1E).
Effects of Live and Heat-Killed HY2782 on Physiological Parameters
The effects of the live or heat-killed HY2782 on body weight and DAI score are evaluated. When the initial body weight was set to 100%, the CON group exhibited a normal increase throughout the experimental period, reaching 105.92 ± 2.01%. In contrast, the DSS group showed a pronounced reduction in body weight, decreasing to 88.78 ± 3.97%. Treatment with sulfasalazine (SULF group) mitigated this weight loss, resulting in a final weight of 94.29 ± 4.22%. The HY2782L and HY2782K groups similarly alleviated DSS-induced weight loss, maintaining body weights of 94.89 ± 2.78% and 92.26 ± 4.45%, respectively (Fig. 2A).
DAI scores were determined based on body weight change, stool consistency, and fecal blood. The CON group showed no alterations in these parameters and maintained a DAI score of 0 throughout the study. In the DSS group, the DAI score began to increase on day 3 (1.50 ± 0.31) and continued to rise, reaching 3.17 ± 0.18 on day 7. In contrast, the SULF group showed a milder progression, with the score increasing to 0.67 on day 3 and 2.13 ± 0.56 on day 7. The HY2782L and HY2782K groups also demonstrated improvement relative to the DSS group, with DAI scores beginning to rise on days 2 and 3, respectively, and reaching 1.83 ± 0.18 and 2.00 ± 0.36 by day 7 (Fig. 2B).
Effects of Live and Heat-Killed HY2782 on Histological Parameters
In the DSS-induced colitis mouse model, administration of HY2782 probiotics and postbiotics improved physiological indicators, including colon length and histological scores. DSS treatment markedly shortened colon length. The CON group exhibited a normal colon length of 8.36 ± 0.71 cm, whereas DSS exposure significantly reduced the length to 6.83 ± 0.41 cm. Treatment with sulfasalazine alleviated this reduction, restoring colon length to 8.19 ± 0.50 cm. Similarly, the HY2782L and HY2782K groups showed significant recovery, with colon length of 7.51 ± 0.70 cm and 7.71 ± 0.54 cm, respectively (Fig. 2C and 2D).
Histological evaluation of colon tissues was performed using H&E staining to assess morphological changes. The total histological score was determined by summing the individual scores for inflammatory cell infiltration, crypt damage, and mucosal erosion. The DSS group exhibited marked inflammatory cell infiltration, crypt damage, and signs of edema. The histological score in the DSS group was significantly elevated to 4.71 ± 1.60 compared with the CON group. In contrast, the SULF group demonstrated substantial improvement, with a reduced score of 1.29 ± 0.49. Both HY2782L and HY2782K groups exhibited the restored histological levels, resulting in scores of 2.83 ± 1.33 and 2.67 ± 0.82, respectively, indicating improved colon structure and reduced inflammation and crypt injury (Fig. 2E and 2F).
Effects of Live and Heat-Killed HY2782 on mRNA Expressions of Genes Related to Tight Junctions and Inflammation
The effects of HY2782 probiotics and postbiotics on the expression of tight junction and inflammation-related genes in colon tissue were evaluated. DSS treatment markedly reduced the mRNA expression levels of the tight junction genes, TJP1, TJP2, and OCLN to 0.73 ± 0.09, 0.63 ± 0.11, and 0.61 ± 0.05 relative to the CON group, respectively. In the SULF group, the gene expression levels were restored to 0.92 ± 0.09, 0.85 ± 0.14, and 0.82 ± 0.09. Likewise, both the HY2782L and HY2782K groups significantly recovered the expression levels of all three tight junction genes (Fig. 3A-3C).
Then, the expression of the pro-inflammatory cytokines IL-1β and TNF-α was examined. DSS treatment led to a significant increase in their expression, reaching 6.29 ± 1.07 and 2.58 ± 0.41 relative to the CON group. In contrast, administration of sulfasalazine reduced their expression to 4.41 ± 0.08 and 1.77 ± 0.21. Administration of HY2782 probiotics further decreased IL-1β and TNF-α expression to 2.50 ± 0.35 and 1.51 ± 0.36, respectively, while the HY2782 postbiotics showed a similar reduction, 2.46 ± 0.49 and 1.54 ± 0.24, respectively. These results indicate that both live and heat-killed HY2782 effectively attenuate DSS-induced inflammatory responses and preserve tight junction integrity in colon tissue (Fig. 3D and 3E).
Effect of Live and Heat-Killed HY2782 on Inflammatory Parameters in Serum
The serum concentration of the pro-inflammatory cytokines IL-1β, IL-6, and TNF-α, as well as chemokine KC, was analyzed. In the CON group, IL-1β, IL-6, and TNF-α levels were 2.35 ± 1.26 pg/ml, 3.49 ± 0.97 pg/ml, and 4.51 ± 0.73 pg/ml, respectively. After DSS administration, their levels significantly elevated to 14.06 ± 3.13 pg/ml, 35.54 ± 7.44 pg/ml, and 10.65 ± 2.76 pg/ml, respectively. Supplementation with sulfasalazine markedly reduced these cytokines 8.53 ± 2.67 pg/ml, 21.40 ± 6.88 pg/ml, and 6.95 ± 2.26 pg/ml. Similarly, the HY2782L group showed significant reductions, with levels of 8.40 ± 1.07 pg/ml (IL-1β), 20.14 ± 2.59 pg/ml (IL-6), and 6.47 ± 1.05 pg/ml (TNF-α). The HY2782K group also attenuated cytokine production, with levels of 9.10 ± 2.19 pg/ml, 17.25 ± 5.93 pg/ml, and 9.20 ± 1.73 pg/ml, respectively (Fig. 4A-4C).
Similarly, chemokine KC levels in serum were 70.26 ± 2.60 pg/ml in the CON group but were significantly increased to 126.13 ± 24.33 pg/ml following DSS exposure. In the SULF group, the KC concentration was reduced to 77.20 ± 11.07 pg/ml. The HY2782L and HY2782K group also significantly reduced KC levels, reaching 81.04 ± 13.46 pg/ml and 66.92 ± 15.04 pg/ml, respectively (Fig. 4D).
Effects of Live and Heat-Killed HY2782 on the Composition of the Gut Microbiota
Alpha diversity was assessed using Faith’s PD. Faith’s PD showed a tendency to increase following DSS treatment. In contrast, supplementation with sulfasalazine, live or heat-killed HY2782, did not result in significant changes to alpha diversity. Beta diversity analysis revealed significant shifts in microbial community composition after DSS induction, whereas minimal changes were observed in the HY2782L and HY2782K groups (Fig. 5A and 5B).
At the genus level, the DSS group exhibited an increased abundance of Peptostreptococcaceae, Erysipelatoclostridium, Parasutterella, and Enterococcus, while showing a reduction in Bifidobacterium, Lachnoclostridium, Lachnospiraceae, Lactobacillus, and Faecalibaculum. In contrast, the SULF group showed an increase in Lactobacillus and a decrease in Sutterellaceae, Acetatifactor, and Akkermansia. The HY2782L group demonstrated an increase in Lachnospiraceae and a decrease in Parasutterella. The HY2782K group exhibited increases in Enterohabdus, Peptococcaceae, and Tepidibacter, with a reduction in Muribaculum (Fig. 5C-5G).
Correlation between Inflammatory Markers and Gut Microbial Profiles after Live and Heat-Killed HY2782 Treatment
Correlation analysis revealed strong positive associations of Peptostreptococcaceae, Erysipelatoclostridium, and Parasutterella with IL-1β, TNF-α, IL-6, and KC. In contrast, Desulfovibrionaceae, Bifidobacterium, Colidextribacter, Lactobacillus, Faecalibaculum, and Lachnospiraceae exhibited negative correlations with these inflammatory markers. Notably, Lactobacillus showed strong negative correlations with all measured inflammatory indicators (Fig. 5H).
Discussion
Inflammatory bowel disease (IBD) is a common intestinal disorder affecting people worldwide, yet its underlying mechanisms remain poorly understood [21, 22]. Current treatments often have side effects, emphasizing the need for safer and more effective therapies that can modulate inflammation and restore dysbiosis with fewer complications [23]. Probiotics are beneficial in IBD, aligning with its pathogenesis, as they enhance intestinal health and help manage or prevent intestinal disorders by blocking inflammatory reactions and altering the bacterial flora [24]. However, since probiotics involve the consumption of live bacteria, there are potential limitations associated with their use. The use of heat-killed probiotics, which retain their functional characteristics, including immunomodulatory and barrier-enhancing effects, could minimize such risks or potential side effects [25, 26], while providing comparable antioxidant and anti-inflammatory benefits to their live counterparts [13, 14]. Consequently, we aim to investigate the preventive potential of both live and heat-killed L. paracasei HY2782 in alleviating colitis. The primary focus of this study was to understand their distinct mechanisms in reinforcing the intestinal barrier and modulating microbial homeostasis to prevent the onset and progression of colonic inflammation.
In preliminary in vitro screening assays, both live and heat-killed HY2782 exhibited anti-inflammatory potential in macrophage-based models, supporting their suitability for subsequent in vivo evaluation. Given the limited translational relevance of isolated macrophage responses, the present study focused primarily on epithelial barrier protection and inflammatory outcomes in a DSS-induced colitis mouse model. In this context, inflammatory responses in colitis are known to involve multiple pattern-recognition receptor–associated signaling pathways, including the TLR4–NF-κB axis, which provides a general biological framework for interpreting inflammation-related gene expression changes observed under DSS-induced conditions [27-29].
We also assessed whether these protective effects extended to intestinal epithelial cells, which are crucial for maintaining mucosal barrier integrity in UC. In Caco-2 cells, we examined tight junction-related genes (TJP1, OCLN, and CLDN) and pro-inflammatory cytokines (TNF-α and IL-1β) to evaluate the effects of live and heat-killed HY2782 on epithelial barrier integrity under inflammatory conditions. Impaired epithelial barrier triggers inflammatory cytokine release, disrupts tight junctions, and increases intestinal permeability, contributing to infection and chronic inflammatory diseases [30]. LPS exposure reduced tight junction gene expression and increased cytokine levels, reflecting epithelial dysfunction. However, treatment with live or heat-killed HY2782 restored tight junction gene expression and suppressed cytokine production, suggesting that both probiotics and postbiotics forms of HY2782 may contribute to the preservation of epithelial barrier integrity under inflammatory conditions.
Based on these in vitro findings, which demonstrate that both the probiotic and postbiotic forms of HY2782 can enhance epithelial integrity and modulate inflammatory responses, we next conducted an in vivo experiment using a DSS-induced colitis mouse model. The DSS model is widely used for its simplicity, reproducibility, and ability to replicate key features of human UC, including acute epithelial injury, mucosal inflammation, and colon-specific pathology [5, 31]. After DSS induction, DAI scores, calculated based on body weight loss, stool consistency, and fecal bleeding, were significantly increased. Additionally, colon length was markedly reduced, and histological scores, including inflammatory cell infiltration, crypt damage, and ulceration, were elevated compared to the CON group. However, the HY2782L and HY2782K groups demonstrated improvements in these physiological and histological alterations, indicating a reduction in disease severity. While the in vitro results indicated that live and heat-killed HY2782 treatments can modulate inflammatory responses and epithelial integrity, in vivo findings demonstrated their protective effects against DSS-induced epithelial injury and mucosal inflammation. These results suggest that live and heat-killed HY2782 may modulate colonic inflammation and barrier dysfunction through multiple complementary actions.
UC is characterized by impaired epithelial integrity and disrupted tight junction structures, which compromise the intestinal barrier’s selective permeability, increasing susceptibility to microbial invasion and toxin diffusion [32]. Tight junction proteins such as TJP1, TJP2, and OCLN are essential for maintaining barrier function, so strategies that enhance epithelial integrity could provide an important therapeutic approach for UC [33, 34]. In this study, both HY2782L and HY2782K groups restored the DSS-induced downregulation of TJP1, TJP2, and OCLN, indicating their ability to preserve tight junction integrity under inflammatory conditions. Additionally, the expression of pro-inflammatory cytokines, including IL-1β and TNF-α, was significantly reduced in the HY2782L and HY2782K groups compared to the DSS group. Balanced cytokine expression is essential for maintaining intestinal homeostasis, whereas IBD is characterized by disrupted cytokine regulation due to excessive pro-inflammatory cytokine production [35]. The suppression of IL-1β and TNF-α in Caco-2 cells suggests that both HY2782 probiotic and postbiotic can regulate the inflammatory response in DSS-induced colitis, potentially reducing mucosal damage and improving overall disease severity.
Despite these promising findings, some limitations should be acknowledged. While we evaluated serum cytokines at the protein level, our molecular analyses in colonic tissue were limited to mRNA expression, which did not capture post-transcriptional regulation or protein-level alterations in local inflammatory signaling. In addition, cytokine regulation involves complex interactions among various immune cell populations, which were not comprehensively assessed in this study. Although both live and heat-killed HY2782 exhibited protective effects, the exact mechanisms warrant further elucidation. Based on our findings, the comparable efficacy between the two forms likely results from a functional convergence of host-centered responses. In the acute DSS model, which is primarily driven by epithelial injury and innate immune activation, host-mediated pathways such as pattern-recognition receptor (PRR) signaling may play a dominant role in maintaining barrier integrity. Consistent with this, our in vitro data showed that both live and heat-killed HY2782 modulated TLR4 and NF-κB expression in macrophages (Fig. S1). This suggests that non-viable components retain the capacity to activate immune pathways that attenuate inflammation and reinforce the intestinal barrier. In this acute injury context, these shared host-mediated mechanisms may override viability-dependent effects, leading to similar phenotypic outcomes. Importantly, this similarity reflects the constraints of an acute injury model rather than identical biological actions. Future studies using chronic or therapeutic models are needed to further distinguish the unique mechanistic pathways of probiotics versus postbiotics.
Furthermore, the serum concentrations of IL-1β, IL-6, TNF-α, and KC were significantly elevated in the DSS group compared to the CON group. These cytokines are key mediators of inflammatory signaling and immune cell activation, playing a pivotal role in exacerbating intestinal inflammation and contributing to tissue damage. Specifically, IL-6 and TNF-α modulate intestinal tight junctions and increase permeability in IBD patients [36, 37]. Excessive IL-1β further increases permeability and activates dendritic cells and macrophages, while KC recruits neutrophils, leading to additional epithelial destruction [38, 39]. There was a significant reduction in the HY2782L and HY2782K groups of these cytokines, suggesting that both forms not only suppressed localized inflammation in the colon but also attenuated the systemic spread of inflammatory responses.
The reduction in serum inflammatory markers and the increased expression of tight junction genes in the HY2782L and HY2782K groups suggest that both probiotics and postbiotics may help reduce inflammation in colitis and improve gut microbiota imbalance [40, 41]. To investigate the relationship between inflammatory markers and microbial alterations, alpha- and beta-diversity analyses were performed. Consistent with previous studies, DSS induction caused a typical dysbiotic shift, characterized by an increase in Faith’s PD [42, 43]. Given that Faith’s PD reflects phylogenetic diversity rather than species richness, its elevation following DSS treatment likely reflects the expansion of phylogenetically diverse opportunistic taxa driven by epithelial barrier disruption and increased intestinal permeability. Although live and heat-killed HY2782 supplementation showed modest trends toward alpha diversity restoration, the severely compromised intestinal environment under acute DSS exposure appeared to limit substantial community-level recovery. Accordingly, beta-diversity analysis revealed marked DSS-driven microbial composition with minimal separation between the supplementation and DSS groups, suggesting that acute inflammatory pressure masked detectable improvements in overall microbial community composition.
After evaluating alpha and beta diversity, taxonomic profiling was performed to assess the microbial groups’ abundance across the CON, DSS, SULF, HY2782L, and HY2782K groups. DSS treatment reduced bacterial families associated with short-chain fatty acids (SCFAs) production, such as Lachnospiraceae, Faecalibaculum, and Lactobacillus, while increasing genera linked to inflammation and epithelial dysfunction, including Erysipelatoclostridium, Peptostreptococcaceae, and Parasutterella [44, 45]. Despite limited recovery in community structure, selective microbial shifts were observed in the HY2782L and HY2782K groups. Live HY2782 treatment increased Lachnospiraceae and completely suppressed Parasutterella excrementihominis, a bacterium associated with colonic inflammation [46]. In the HY2782K group, inflammation-associated taxa, including Bacteroides, were reduced, and Lachnospiraceae increased to levels similar to those observed in the HY2782L and SULF groups, suggesting a compositional shift toward microbial taxa commonly associated with intestinal homeostasis. Notably, Ligilactobacillus murinus, which supports epithelial integrity, was substantially increased in the HY2782K group, surpassing levels in the CON group, an effect not observed in the HY2782L group. One possible explanation is that differences in bacterial viability may influence host–microbe interactions and microbial competitiveness under DSS-induced inflammatory stress. Viable and non-viable bacterial preparations are known to differ in their interactions with the intestinal environment, which may affect the persistence or expansion of specific taxa during acute inflammation. In the present study, however, the precise mechanisms underlying the differential abundance of specific taxa, including L. murinus, were not directly examined and therefore cannot be conclusively determined, although previous studies have suggested that host pattern-recognition receptors such as TLR2 and NOD2 play important roles in mediating epithelial–microbial interactions [47]. In contrast, there was an increase in Lactobacillus in the SULF group, suggesting restoration of the colonic barrier, creating a more favorable environment for Lactobacillus growth [48]. The SULF group did not show a reduction in Parasutterella excrementihominis, which distinguishes its microbial response from that of the HY2782L and HY2782K group. This pattern is consistent with the pharmacological profile of sulfasalazine, which is metabolized in the colon into 5-ACA and primarily exerts anti-inflammatory effects through local immune pathways rather than through modulation of the gut microbiota [49]. Given the limited recovery of overall microbial diversity, the observed taxonomic changes are more appropriately interpreted as being associated with, rather than driving, the anti-inflammatory and barrier-protective effects observed in vivo. As a result, sulfasalazine typically induces only limited shifts in microbial composition, explaining why meaningful microbiota restructuring was not observed in this group despite improvements in inflammatory readouts.
Phocaeicola vulgatus, also known as Bacteroides vulgatus, is commonly found in the colonic microbiota of UC patients, with some exhibiting elevated serum antibodies against its OmpA protein, which promotes bacterial adherence. Protease-producing P. vulgatus strains are linked to increased intestinal permeability and active colitis through the secretion of dipeptidyl peptidases [50]. In this study, DSS treatment increased P. vulgatus abundance, with a relatively minor reduction in the SULF and HY2782L groups, while a significant decrease in the HY2782K group. Live bacteria likely exert their effects through direct competition with P. vulgatus by metabolizing organic acids and creating an unfavorable environment for their growth. In contrast, heat-killed bacteria activate intestinal immune responses, enhancing tight junction protein expression through immune receptors like TLR2 and NOD2, thereby strengthening the epithelial barrier [47]. Additionally, heat-killed bacteria do not induce competitive inhibition as strongly as live bacteria, which aligns with the observed increase in TJP gene expression in the colon in the HY2782K group. The results indicate that live and heat-killed HY2782 suppress P. vulgatus through different mechanisms, contributing to improved intestinal barrier function and reduced inflammation in the DSS-induced colitis model. Taken together, the microbiome data indicate that administration of live or heat-killed HY2782 did not induce large-scale restructuring of the gut microbial community under acute DSS-induced inflammatory conditions. Instead, the observed selective taxonomic shifts are more appropriately interpreted as being associated with, rather than driving, the anti-inflammatory and barrier-protective effects observed in vivo.
In summary, the present findings indicate that L. paracasei HY2782, administered as either live or heat-killed formulations, has preventive potential against colitis-associated inflammation and epithelial barrier disruption in a DSS-induced model. In line with this objective, the present study was intentionally designed as a preventive proof-of-concept model, and therapeutic administration following disease onset was not evaluated. Taken together, these findings suggest that both live and heat-killed forms of HY2782 can ameliorate inflammatory responses and epithelial barrier dysfunction primarily through host-centered effects rather than extensive microbiota remodeling (Fig. 6). However, these results should be interpreted with caution, as mechanistic differences between live and inactivated preparations were not fully resolved, and the relatively short duration of administration limits conclusions regarding long-term efficacy and microbiome remodeling. Consequently, additional studies incorporating extended preventive dosing strategies, detailed mechanistic investigations, and diverse colitis models will be required to more definitively establish the preventive potential and translational applicability of HY2782.
Supplemental Materials
Supplementary data for this paper are available on-line only at http://jmb.or.kr.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rodriguez-Nogales A Algieri F Garrido-Mesa J Vezza T Utrilla MP Chueca N 2018 Intestinal anti-inflammatory effect of the probiotic Saccharomyces boulardii in DSS-induced colitis in mice: Impact on micro RN As expression and gut microbiota composition J. Nutr. Biochem.61129139 https://doi.org/10.1016/j.jnutbio.2018.08.005 10.1016/j.jnutbio.2018.08.00530236870 · doi ↗ · pubmed ↗
- 2Celiberto LS Bedani R Rossi EA Cavallini DCU 2017 Probiotics: The scientific evidence in the context of inflammatory bowel disease Crit. Rev. Food. Sci. Nutr.5717591768 https://doi.org/10.1080/10408398.2014.941457 10.1080/10408398.2014.94145725996176 · doi ↗ · pubmed ↗
- 3Zhang T Zhang W Feng C Kwok L-Y He Q Sun Z 2022 Stronger gut microbiome modulatory effects by postbiotics than probiotics in a mouse colitis model NPJ Sci. Food 653 https://doi.org/10.1038/s 41538-022-00169-9 10.1038/s 41538-022-00169-936379940 PMC 9666507 · doi ↗ · pubmed ↗
- 4Ma W Lian L Guo L Wu Y Huang L 2025 A synbiotic combination of mixed probiotics and oligofructose restores intestinal microbiota disturbance in DSS-induced colitis in mice Front. Microbiol.161582155 https://doi.org/10.3389/fmicb.2025.1582155 10.3389/fmicb.2025.158215540778208 PMC 12330387 · doi ↗ · pubmed ↗
- 5Li Z Peng C Sun Y Zhang T Feng C Zhang W 2024 Both viable Bifidobacterium longum subsp. infantis B 8762 and heat-killed cells alleviate the intestinal inflammation of DSS-induced IBD rats Microbiol. Spectr.12e 0350903523 https://doi.org/10.1128/spectrum.03509-23 10.1128/spectrum.03509-2338647334 PMC 11237488 · doi ↗ · pubmed ↗
- 6Hu Q Yu L Zhai Q Zhao J Tian F 2023 Anti-inflammatory, barrier maintenance, and gut microbiome modulation effects of Saccharomyces cerevisiae QHNLD 8L 1 on DSS-induced ulcerative colitis in mice Int. J. Mol. Sci.246721 https://doi.org/10.3390/ijms 24076721 10.3390/ijms 2407672137047694 PMC 10094816 · doi ↗ · pubmed ↗
- 7Huang C Hao W Wang X Zhou R Lin Q 2023 Probiotics for the treatment of ulcerative colitis: a review of experimental research from 2018 to 2022 Front. Microbiol.141211271 https://doi.org/10.3389/fmicb.2023.1211271 10.3389/fmicb.2023.121127137485519 PMC 10358780 · doi ↗ · pubmed ↗
- 8Liu YW Su YW Ong WK Cheng TH Tsai YC 2011 Oral administration of Lactobacillus plantarum K 68 ameliorates DSS-induced ulcerative colitis in BALB/c mice via the anti-inflammatory and immunomodulatory activities Int. Immunopharmacol.1121592166 https://doi.org/10.1016/j.intimp.2011.09.013 10.1016/j.intimp.2011.09.01321996541 · doi ↗ · pubmed ↗