Dryocrassin ABBA Inhibits the Function of LLO and SortaseA to Alleviate the Virulence of Listeria monocytogenes
Jiahui Lu, Junlu Liu, Hanbing Zhou, Yifan Duan, Zehua Wang, Guizhen Wang

TL;DR
A natural compound called Dryocrassin ABBA reduces the harmful effects of Listeria monocytogenes by inhibiting key proteins involved in infection.
Contribution
Dryocrassin ABBA is shown to inhibit LLO and SrtA, reducing Listeria's virulence without directly killing the bacteria.
Findings
ABBA inhibits LLO oligomer formation and hemolytic activity.
ABBA reduces SrtA activity, decreasing biofilm formation and cell invasion.
ABBA shows protective effects in L. monocytogenes-infected Galleria mellonella without cytotoxicity.
Abstract
Listeria monocytogenes is a deadly foodborne pathogen, its virulence factors Listeriolysin O (LLO) and Sortase A (SrtA) play a critical role in its infection and have been identified as key targets for developing its inhibitor. This study reveals that the natural compound Dryocrassin ABBA (ABBA) alleviates the toxicity of L. monocytogenes by effectively inhibiting the functions of LLO and SrtA. ABBA bound to the binding pocket of LLO and inhibited the formation of its oligomers, which in turn reduced the ability of LLO to lyse mammalian erythrocytes. When co-cultured with L. monocytogenes, ABBA reduced the hemolytic activity of the cultural supernatant. In addition, ABBA bound to SrtA and reduced its transpeptide activity, which in turn resulted in a reduction in the formation of bacterial biofilm, the adhesion and invasion of L. monocytogenes to cells. Thr415 and Lys505 of LLO, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
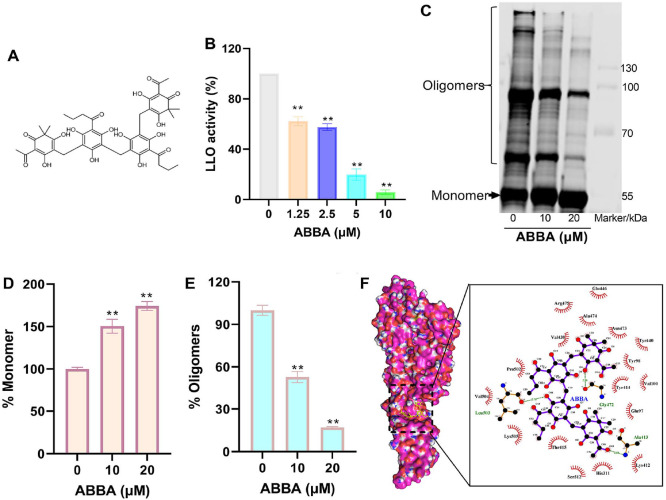 Figure 1
Figure 1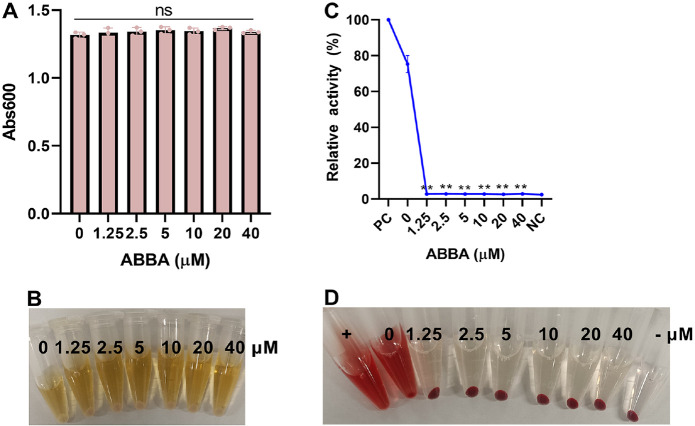 Figure 2
Figure 2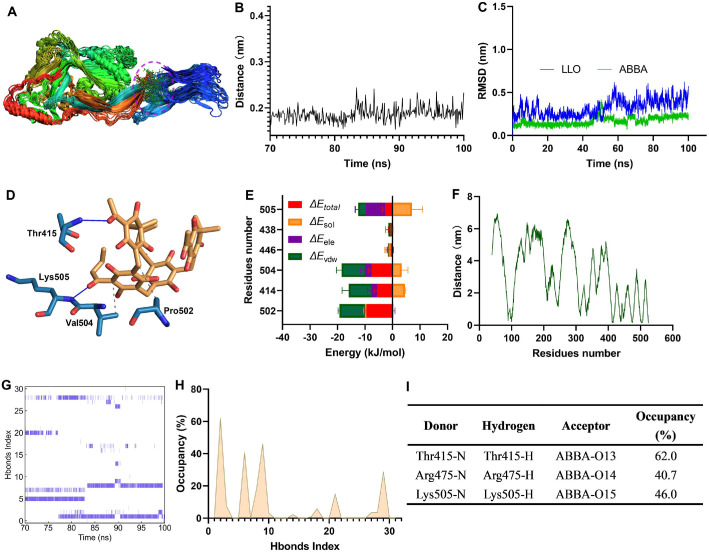 Figure 3
Figure 3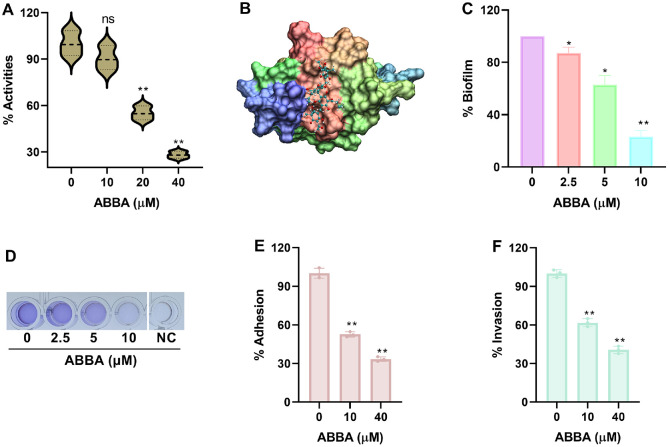 Figure 4
Figure 4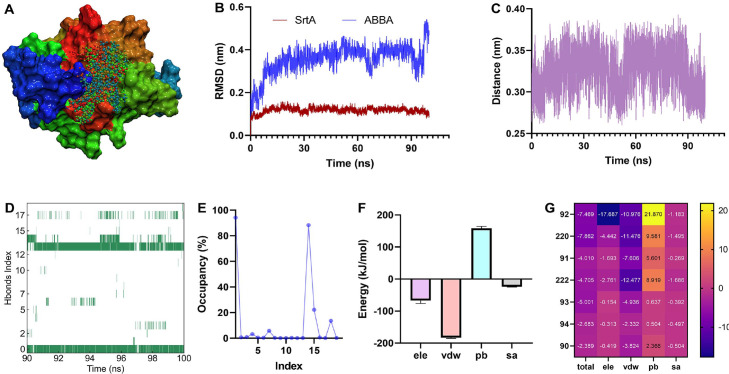 Figure 5
Figure 5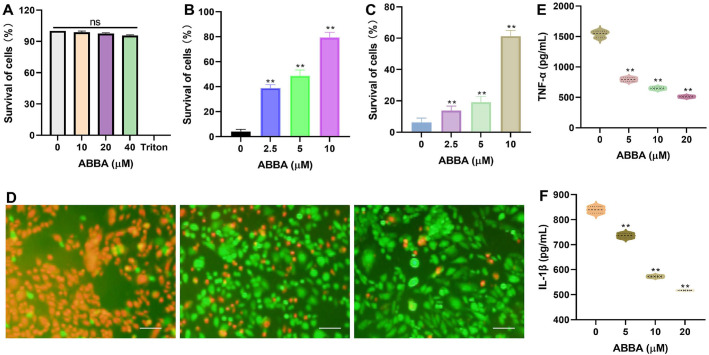 Figure 6
Figure 6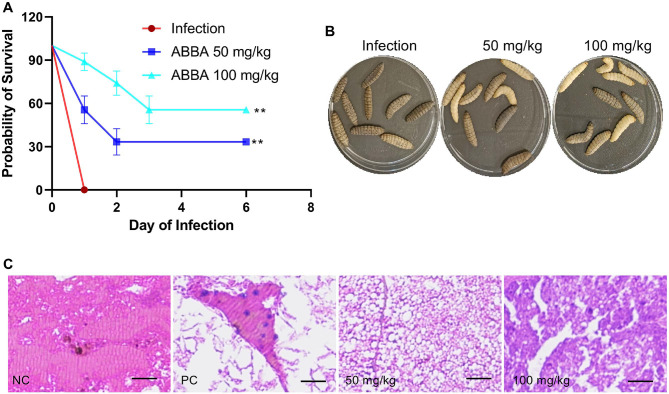 Figure 7
Figure 7- —Scientifc Research Project of Jilin Province
- —Natural Science Foundation that from Jilin Provincial Department of Science and Technology
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiochemical and Structural Characterization · Transgenic Plants and Applications · Antimicrobial Peptides and Activities
Introduction
Listeria monocytogenes is one of the deadliest foodborne pathogens, it can cross the blood-brain barrier and placental barrier after infecting people, causing gastroenteritis, bacteremia, meningitis and many other diseases, the mortality of L. monocytogenes infections can reach as high as 30%, which bring expensive medical costs and seriously threaten public health [1-3]. Besides, L. monocytogenes also infect chickens, cattle, sheep, pigs and many other animals in the poultry industry to affect their healthy growth, which would reduce the production and quality of poultry industry and cause economic losses [4-6]. L. monocytogenes infection is widespread all over the world, the abuse of antibiotic led to L. monocytogenes severe resistance, the continuing development of L. monocytogenes resistance exerted challenge to veterinary clinical, food safety and public health [7-9]. Therefore, developing alternative strategy and inhibitors to combat this pathogen is urgent.
Listeriolysin O (LLO) is an exocrine toxin of L. monocytogenes, it is encoded by the hly gene and it belongs to the cholesterol-dependent cytolysin family [10]. LLO can specifically bind to cholesterol on the cell membrane of mammalian to form oligomers that contain 35-40 monomers, the oligomers can pierce the cell membrane, trigger cytotoxicity, and affect the normal physiological functions of cells [11]. LLO also helps L. monocytogenes escape from phagocytic vesicles by punching holes on the cell membrane to promote bacteria survival and proliferation [12-14]. LLO have also been reported to trigger inflammatory responses by intervening in host inflammation-related signalling pathways [15, 16]. The hly knockout strain was unable to escape phagosomes and could hardly colonise in the spleen and liver of mice [17]. Sortase A (SrtA) is a transpeptidase expressed by L. monocytogenes, encoded by the srtA gene, which can specifically recognise virulence proteins containing LPXTG motifs, and cleave the peptide bonds between T and G, covalently anchoring virulence proteins to the surface of bacterial cell walls [18]. More than 40 genes encoding LPXTG proteins have been identified in L. monocytogenes [19], many of them are internalin family protein, such as InlA protein that helps L. monocytogenes adhesion and invasion, the InlH protein that affects host immune response, and the InlL that affects biofilm formation [19, 20]. The srtA knockout strain was unable to adhere to and invade host cells, and its pathogenicity was significantly reduced in mouse models [21-23]. Therefore, LLO and SrtA are key targets for the development of novel anti-L. monocytogenes agents.
Dryocrassin ABBA (ABBA) is mainly exists in medicinal plant that named Dryopteris crassirhizoma Nakai [24]. ABBA has been confirmed to possess anti-inflammatory, anti-cancer, and antibacterial properties [25, 26]. Researches indicated that ABBA has the potential to inhibit the virulence of Streptococcus pneumoniae and Staphylococcus aureus [24-26]. However, ABBA inhibits L. monocytogenes pathogenicity by targeting LLO and SrtA has not been disclosed. Here, we found that ABBA inhibited the hemolytic activity of LLO protein and L. monocytogenes culture supernatant by directly binding with LLO and reduced its secretion to the cultural medium. In addition, ABBA inhibited the formation of L. monocytogenes biofilm and its adhesion and invasion to host cells by targeting SrtA. Furtherly, ABBA reduced the cytotoxicity and inflammation mediated by L. monocytogenes and demonstrated a comprehensive protective effect against L. monocytogenes-infected Galleria mellonella model. The elucidation of the molecular mechanism of ABBA interacted with LLO and SrtA established the foundation for its application.
Materials and Methods
Reagents, Proteins, Cell Lines, Strains, and Growth Conditions
ABBA (purity ≥98%) was purchased from Chengdu Herpurify Co., Ltd. (China). Trypticase Soy Broth (TSB) and agar were purchased from Haibo Bio-Technology Co., Ltd., (China). The G. mellonella was purchased from Tianjin Huiyude Biotechnology Co., Ltd., (China). The substrate peptide of SrtA (Dabcyl-QALPETGEE-Edans) was purchased from GL Biochem (China), Ltd. The pET-28a vector that carried the hly or SrtA gene was constructed in our previous research, the LLO (wild type or its mutants) and SrtA proteins were purified and stored at -80°C based on previously reported methods [27, 28]. L. monocytogenes ATCC19115, mouse macrophage J774A.1 and Hela cells were purchased from American Type Culture Collection (ATCC). L. monocytogenes was cultured in TSB medium at 37°C with shaking or static.
LLO and L. monocytogenes Culture Supernatant Hemolytic Activity Inhibition
The hemolytic activity assay was carried out as described previously [29]. In brief, purified LLO protein (1 μg) was incubated with different concentrations of ABBA (0, 1.25, 2.5, 5, and 10 μM) at 37°C for 20 min, and then sterile defibrinated sheep blood was added (final concentration 2.5%). After incubation for 10 min, the samples were centrifuged at 12,000 rpm for 2 min, and equal volumes of the supernatant were collected to measure the absorbance at 543 nm (Abs543) to analyze the inhibitory effect of ABBA on LLO activity. For L. monocytogenes cultural supernatant hemolytic activity assay, equal volume supernatant was added to sterile phosphate buffered saline (PBS) buffer that contained sterile defibrinated sheep blood (2.5%) and incubated at 37°C for 10 min, then samples were detected Abs543 after centrifuged. Sheep blood cells treated with sterile water or PBS were used as positive control (PC) or negative control (NC).
Oligomer Formation Inhibition Assay
This assay referred the method reported previously [17]. LLO protein (4 μg) was incubated with with different concentrations of ABBA (0, 10, and 20 μM) at 37°C for 20 min, following sterile defibrillated sheep blood was added with the final concentration of 1%. After incubated on ice for 2 min, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer without β-mercaptoethanol was added and all samples were treated at 55°C for 10 min. The samples were separated by using a 6% SDS-PAGE gel, and the proteins were transferred to PVDF membranes. After blocked with 5% skimmed milk, samples were incubated with His-tag antibody for 2 h at room temperature (1:1000, Sangon Biotech, D191001), after washed, the PVDF membranes was incubated with a sheep anti-mouse secondary antibody that conjugated with HRP. Image was collected to analyze the effect of ABBA on the formation of LLO oligomers. The LLO monomer detection was based on coomassie brilliant blue dyeing.
Computational Biology Assay
This assay was performed by referencing methods described previously [30]. Simply, the crystal structures of LLO (PDB ID 4CDB) and SrtA protein (PDB ID 5HU4) were used as receptors, ABBA (CAS) was used as ligand. A docking box with the spacing of 1 angstrom (Å) was generated, AutoDock Vina was used to perform the docking calculation. After obtaining the complexes, the molecular dynamics simulations assays were performed by using GROMACS 2020.6 to confirm the reliability of the binding. Amber 99SB-ILDH force field and TIP3P water model were applied. Root-mean-square deviation (RMSD) was analyzed to evaluate the stability of the conformations. Binding free energy was analyzed to determine the critical residues involving in the binding. The mutants proteins of LLO that used for calculating the binding free energies were generated by using swiss-PdbViewer 4.10 [31].
Bacterial Growth Assay
L. monocytogenes and various concentrations of ABBA (0, 1.25, 2.5, 5, 10, 20, and 40 μM) were incubated at 37°C for 8 h, the Abs600 values of each sample were determined to evaluate the effect of ABBA on bacterial growth. The minimum inhibitory concentration assay was performed as follows: serious concentrations (0, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120 μM) of ABBA in TSB medium was prepared, then bacteria were added to reach a final concentration of 5 × 10^5^ colony forming units per milliliter (CFUs/ml).
SrtA Activity Experiment
L. monocytogenes SrtA protein with different concentrations of ABBA (0, 10, 20, and 40 μM) was incubated at 37°C for 30 min, then the substrate peptides with fluorophores at both ends were added and samples were incubated at 37°C in the dark. After 60 min, the fluorescence intensity of the samples were measured with the excitation wavelength of 350 nm and emission wavelength of 520 nm. The inhibitory effect of ABBA on the activity of L. monocytogenes SrtA was analyzed [32].
L. Monocytogenes Biofilm Formation Inhibition Experiment
L. monocytogenes was co-cultured with different concentrations of ABBA (0, 2.5, 5, 10, 20, 40, and 80 μM) at 37°C for 24 h. Then the samples were washed with sterile PBS after the culture medium was discarded, and stained with 0.1% crystal violet for 20 min. Subsequently, samples were washed after the excess crystal violet was discarded and treated with 33% glacial acetic acid. Abs570 values were detected to analyze the effect of ABBA on the formation of L. monocytogenes biofilms [33].
Adhesion and Invasion Experiments
Human colorectal cancer cells (CaCO_2_) were cultured in Dulbecco's Modified Eagle's Medium (DMEM) with 10% fetal bovine serum (FBS) at 37°C with 5% CO_2_. Cells were seeded into 24 well plate with a density of 1 × 10^5^ cell/ml and cultured overnight. The next day, the culture medium was replaced with DMEM containing L. monocytogenes and various concentrations of ABBA (0, 10, 40 μM). After 60 min of infection, the culture medium was removed, cells were washed with sterile PBS harvested. After diluting, samples were coated on TSB agar plates and incubated overnight at 37°C. The effect of ABBA on L. monocytogenes adhesion was evaluate by analyzing the number of the colonies. For the invasion assay, cells with L. monocytogenes and ABBA co-cultured for 1.5 h, then the medium was removed and gentamicin (100 μg/ml) was used to treat cells for 1 h. Following, the medium was removed, cells were lysed after washing, and samples were coated onto TSB medium and cultured overnight [33]. The inhibitory effect of ABBA on L. monocytogenes invasion was evaluated by counting the colonies number.
Detection of Lactate Dehydrogenase Activity
J774A.1 were cultured in DMEM with 10% FBS at 37°C with 5% CO_2_. The cells were seeded into 96 well plate (1.5 × 10^4^ cells/well) and cultured overnight. The next day, L. monocytogenes with various concentrations of ABBA (0, 2.5, 5, 10 μM) was used to treat cells. Five hours later, the cultural medium were used to detect the lactate dehydrogenase (LDH) level by referring the protocol of a kit (Beyotime, China). The cells that received 10 μM ABBA treatment were stained with live/dead reagents (Beyotime), images were obtained to observe the survival of the cells [29]. The cytotoxicity of ABBA or it alleviated the cytotoxicity from LLO was determined by detecting the LDH levels.
Inflammatory Factor Detection Experiment
J774A.1 cells were seeded in a 6-well plate (1 × 10^6^ cells/well) and cultured overnight. The next day, the culture medium was replaced with fresh DMEM containing L. monocytogenes and ABBA (0, 5, 10, 20 μM), and samples were treated for 5 hours at the same cultural condition. Then the culture medium was transferred to clean tubes and centrifugated (4°C, 12000 rpm, 5 min)[29]. The levels of tumor necrosis factor α (TNF-α) and interleukin 1β (IL-1β) that released to the medium were detected by using an enzyme-linked immunosorbent assay (ELISA) kit (Sangon Biotech, China).
L. monocytogenes Infect G. mellonella Model
G. mellonella used in this assay were purchased from Tianjin Huiyude Biotechnology Co., Ltd. Healthy and without melanin samples were used to carry out the assay. L. monocytogenes were injected into each G. mellonella (2 × 10^5^ CFUs) by using a microinjection pump to construct the infection model, the infected G. mellonella were divided into three groups randomly, and nine individual was arranged to each group. 30 min after infection, ABBA (50 mg/kg, 100 mg/kg) was injected into the infected samples, the infected G. mellonella that received equal volume solvent treatment was defined as infection group, which was used as positive control (PC), healthy and without melanin samples that got solvent treatment was defined as negative control (NC) group[34]. The survival of the G. mellonella were monitored everyday to evaluate the protect effect of ABBA against L. monocytogenes infection in vivo. Samples from each group was stained with hematoxylin-eosin (H&E) after fixing with 4% formaldehyde, the pathological tissue damage of each sample was observed to determine the alleviation of ABBA against L. monocytogenes infection symptoms.
Data Statistics and Analysis
The research data that obtained from three independent experiments were shown as means with standard deviations (SDs), the analysis of significant differences between data was performed by using GraphPad Prism software (version 9.5.0) which built-in t test analytical method, p ≤ 0.05 was defined as significant difference.
Results
ABBA Inhibits LLO Activity by Binding with and Suppressing Its Oligomer Formation
The molecular structure of ABBA is shown in Fig. 1A. The LLO activity in the group that did not receive ABBA was defined as 100%, when various concentrations of ABBA was added to the reaction system, the LLO activity reduced to 62.32%, 57.65%, 19.81% and 5.84% respectively (Fig. 1B). As a member of cholesterol-dependent cytolysin (CDC), forming oligomer is critical for LLO activity, we speculate that ABBA inhibits the formation of LLO oligomers, which was confirmed by the oligomer formation assay. As shown, the LLO monomer was approximately 55 kDa (Fig. S1), the amount of LLO monomer increased along with the increasing ABBA concentrations (100% to 150.52%, 174.29%), but the oligomer formation decreased gradually with the values from 100% to 52.82%, 17.02% (Fig. 1C-1E), suggesting a direct binding between ABBA and LLO was generated, this was confirmed by a docking calculation assay. After docking, nine binding mode was obtained, eight of them were bound at the interface between domains 2 and 4 of LLO, and only one was bound at the interface between domains 1 and 3 (binding mode 6) (Fig. S2), indicating that ABBA is more likely to bind at the junction between domains 2 and 4. The binding mode 1 that with the affinity of -9.97 kcal/mol was shown in Fig. 1F, the binding sites analysis showed that Val504, Pro502, Leu503, Tyr414, Arg475 generated interactions with ABBA (Fig. 1F). These findings suggest that ABBA inhibits oligomer formation by directly binding to LLO proteins, thereby reducing its hemolytic activity.
ABBA Inhibits the Activity of L. monocytogenes Culture Supernatant
The bacteria reach the same growth endpoint when L. monocytogenes was co-cultured with different concentrations of ABBA for 8 h (Fig. 2A and 2B), indicating that ABBA does not affect the normal growth of L. monocytogenes within the test concentration, the CFUs analysis also confirm this result (Fig. S3). The minimum inhibitory concentration of ABBA against L. monocytogenes was 100 μM. The hemolytic activity of the L. monocytogenes and ABBA cultural supernatant was analyzed, as shown, the amount of heme that released to the medium in the PC group was defined as 100%, the relative activity of L. monocytogenes cultural supernatant was 75.27%, while, the hemolytic activity of the samples that received ABBA treatment decreased sharply to 2.78%, which is comparable to the NC group (Fig. 2C-2D).
Elucidation of the Interactive Mechanism between ABBA and LLO
To explore the interactive mechanism between LLO and ABBA, we performed molecular dynamics simulation assay. The trajectory files were obtained based on a 100 ns simulation, the structural superposition of ABBA and LLO at different time-points shows that ABBA consistently occupy the initial binding position of LLO during the simulation process (Fig. 3A). The distance between ABBA and LLO fluctuated around 0.2 nm (Fig. 3B), further confirming the stability of the binding. The RMSD of LLO and ABBA fluctuated around 0.32 nm and 0.17 nm, respectively during the simulation process (Fig. 3C), suggesting that they maintained excellent configuration throughout the dynamics process. Thr415, Lys505, Pro502, and Val504 of LLO generated interaction with ABBA (Fig. 3D). These interactions were confirmed by the residue energy decomposition, besides, Tyr414 also contribute more energy (Fig. 3E), the closer distance between these residues and ABBA (Fig. 3F) prove the energy contributions. Hydrogen bonds (Hbonds) analysis reveal more than 30 pairs of Hbonds between ABBA and LLO were generated during 70-100 ns, though many of them have a low existence (Fig. 3G-3H), there are three pairs Hbonds have a higher occupancy (Fig. 3G-3H). The -NH of Thr415, Arg475, and Lys505 interacted with the -O13, -O14, and -O15 in ABBA, with occupancy of 62.0%, 40.7%, and 46.0%, respectively (Fig. 3I). Besides, the binding free energies between ABBA and the LLO mutants and the inhibitory effects of ABBA against the LLO mutants proteins activity decreased significantly (Tables S1 and S2). As shown in Fig. S4, The LLO protein consists of four domains, namely domain 1, 2, 3, and 4. Domain 4 specifically binds to cholesterol of host cells, triggering a series of structural changes in the LLO protein. Subsequently, multiple monomers assemble into a barrel-shaped oligomer, which inserts into the cell membrane, causing leakage of cellular contents and leading to disease. Although it is the fourth domain that interacts with the host receptor, each domain of LLO is crucial for oligomer formation. In this study, ABBA binds at the junction between Domain 2 and Domain 4, where residues Lys505, Pro502, Val504, and Tyr414 form weak interactions that affect the formation of LLO oligomers, thereby inhibiting LLO's hemolytic activity.
ABBA Inhibits Biofilm Formation, Adhesion and Invasion of L. monocytogenes by Affecting SrtA Function
The activity of SrtA from sample that did not receive ABBA treatment was defined as 100%, while, when ABBA was added, the activity of SrtA reduced to 90.50%, 55.12%, and 28.12% respectively (Fig. 4A), suggesting ABBA generated direct interaction with SrtA, this was confirmed by molecular docking assay, the result showed that ABBA bound to the active pocket of SrtA with an affinity of -8.67. Kca/mol (Fig. 4B). Surface proteins that anchored by SrtA involving in L. monocytogenes biofilm formation, adhesion and invasion of the bacteria to host cells, therefore, we evaluated these indicators. For biofilm formation, when various concentrations of ABBA was used to treat L. monocytogenes, the biofilm formation reduced from 100% to 87.19%, 62.81% and 23.11% (Fig. 4C), the visual image demonstrate the quantitative results (Fig. 4D). For adhesion and invasion assay, we found that when different concentrations of ABBA was used to treat samples, the adhesion of L. monocytogenes to Caco2 cells reduced from 100% to 52.71% and 33.45% (Fig. 4E), and the invasion reduced to 61.78% and 40.67% (Fig. 4F). These results indicate that ABBA binds to SrtA and inhibits its biological function, thereby reducing biofilm formation and the adhesion and invasion of bacterial to host cell.
ABBA Generates Interactions with Asn92, Pro220, Val91 and Lys222
To elucidate the interactive molecular mechanism between ABBA and SrtA, a 100 ns molecular dynamics simulation experiment was performed, and we found that ABBA was always located in the initial binding pocket of SrtA throughout the simulation process by analyzing the relative positions of ABBA and LLO at different moment (Fig. 5A). During the simulation, the RMSD of ABBA and SrtA fluctuated around 0.36 nm and 0.12 nm, respectively (Fig. 5B), and the distance between ABBA and SrtA fluctuated around 0.33 nm (Fig. 5C), indicating that ABBA and SrtA maintained stable structures and binding modes during the simulation. There are nearly 20 pairs of hydrogen bonds at 90-100 ns (Fig. 5D), but most of them were ephemeral, two pairs were stable existence with the occupancy of 94.20% and 88.30%, respectively (Fig. 5E), the -NH and -O groups of Asn92 interacted with the -O12 and -O16 of ABBA, respectively (Table 1). We found that van der Waals force (vdw) and electrostatic action (ele) are the main interaction forces with the values of -182.68 kJ/mol and -67.28 kJ/mol (Fig. 5F), more exactly, Asn92, Pro220, Val91, Lys222 contributed more ele and vdw to promote the binding (Fig. 5G).
ABBA Alleviates Cytotoxicity and Inflammation Mediated by Listeria monocytogenes
When J774A.1 cells were treated with various concentrations of ABBA, the survival of the cells was comparable to that of the negative control group (only DMEM treatment group), indicating that ABBA had no cytotoxic effect within the test concentration range (Fig. 6A). For LLO treated cells, the survival was 38.81%, 48.73% and 79.45% when various concentrations of ABBA was applied (Fig. 6B), and the survival was 13.75%, 19.13%, and 61.33% when cells treated with L. monocytogenes and ABBA (Fig. 6C). For live/dead cells stain, when different concentrations of ABBA were applied, the red fluorescence significantly decreased, whereas the green fluorescence significantly increased, indicating that dead cells decreased and live cells increased (Fig. 6D). The above results demonstrate that ABBA can significantly alleviate the cytotoxic effects mediated by LLO or L. monocytogenes from both qualitative and quantitative perspectives. For ELISA assay, when J774A.1 cells were treated with L. monocytogenes, a large amount of TNF-α (1540.07 pg/ml) and IL-1β (839.26 pg/ml) were detected in the culture medium, and the levels of TNF-α decreased to 795.66 pg/ml, 647.95 pg/ml, and 511.32 pg/ml (Fig. 6E), respectively. The level of IL-1β decreased to 736.16 pg/ml, 572.03 pg/ml, and 516.86 pg/ml (Fig. 6F), indicating that L. monocytogenes can trigger an inflammatory response in J774A.1 cells, while ABBA can significantly alleviate L. monocytogenes-mediated cellular inflammatory responses.
ABBA Protects G. mellonella from L. monocytogenes Infection
All the G. mellonella in the infected group died at 24 h, the 50 mg/kg and 100 mg/kg ABBA treatment groups also found died G. mellonella at 24 h, but the survival of these groups were 33.33% and 55.56%, respectively (Fig. 7A). For the infectious symptoms, individual in the infection group died completely, they were swelling and the whole body was filled with melanin, but these characters were alleviated in the samples from the ABBA treatment group (Fig. 7B). Sample from the NC group exhibited complete, undamaged tissue structure, and there was no inflammatory infiltration and melanin; samples from the PC group showed complete tissue destruction and infiltration of inflammatory factors, while, these characters were alleviated in the ABBA treatment group samples (Fig. 7C). These results indicate that L. monocytogenes infection could cause serious tissue damage, trigger inflammation, and result in death, whereas ABBA treatment could significantly alleviate pathological tissue damage, reduce the inflammatory response, and improve the survival of the infected G. mellonella.
Discussion
LLO serves as the “Swiss army knife” of L. monocytogenes [35, 36], considering its crucial role in L. monocytogenes infection, research on its inhibitors is compelling. Some bioactive properties from medicinal plants have been identified as inhibitors of LLO, such as amentoflavone, atractylodin, fisetin, and so on [37-39]. These inhibitors suppress L. monocytogenes-mediated inflammatory responses and their pathogenicity by intervening in LLO function [40]. But reports on plant activity ingredients targeting both LLO and SrtA are rare, phloretin and isoflavone glucoside genistin showed inhibiting effect to these toxins [28, 41]. This study found that ABBA simultaneously targets LLO and SrtA to suppress L. monocytogenes virulence. By targeting LLO and SrtA, ABBA reduced the haemolytic activity of L. monocytogenes culture supernatant and biofilm formation, and it inhibited the adhesion and invasion of L. monocytogenes to the host cells. We speculate that the inhibition of SrtA function results in a reduction of surface protein that anchored to the bacterial cell wall, such as inlA, which lead to the reduction of bacterial adhesion and invasion to cells, which may be explored in our future work.
The crystal structures of LLO and SrtA have been elucidated [42, 43]. The LLO crystal structure is divided into four domains; the residues in domain four interact with the host cell receptor and form oligomer to pierce cells. However, previous studies have demonstrated that the other three domains are crucial for the formation of LLO oligomers [42, 44]. The binding sites of previously reported inhibitors with LLO differ slightly; but they can inhibit the formation of LLO oligomers and affect its haemolytic activity [17, 29]. This confirms the importance of different domains in the formation of LLO oligomers and their haemolytic activity. In this study, based on molecular dynamics simulation analysis, Thr415, Lys505, Pro502, and Val504 in LLO protein generated interactions with ABBA and promote their binding. For SrtA, His127, Arg197 and Cys188 have been identified as its active sites that interact with the substrates, as SrtA lost almost all transpeptide activity after these residues were mutated to Ala [45, 46]. In this study, we found that ABBA bound to the back of the active pocket of L. monocytogenes SrtA, Asn92, Pro220, Val91 and Lys222 are important residues on promoting their binding. Unlike previous reports, ABBA did not directly bind to L. monocytogenes SrtA activity center in this study, and we speculate that ABBA may affect SrtA function through non-competitive inhibition, which worth further exploring in the future.
ABBA did not exhibited anti-L. monocytogenes and cytotoxicity effects at the effective inhibiting concentrations. Moreover, ABBA treatment significantly reduced L. monocytogenes-mediated cytotoxicity and inflammatory responses, and it protected G. mellonella from L. monocytogenes infection, which results from the inhibitory effects of ABBA against LLO and SrtA simultaneously, although there may be other underlying mechanisms need to be explore in future. The bioavailability of ABBA can reach as high as 50.1%, and its metabolic kinetics parameters have been well-defined [47]. These results provide a significant basis for future applications of ABBA.
Conclusion
ABBA, a natural active ingredients, inhibited the hemolytic activity of LLO and L. monocytogenes culture supernatants by suppressing oligomer formation. In addition, ABBA directly bound with SrtA to inhibit its transpeptidase activity, which resulted in a reduction of bacterial cell wall surface proteins that anchored by SrtA, thus the formation of bacterial biofilm was defective, and the adhesion and invasion of bacteria to host cell were alleviated. ABBA and LLO or SrtA maintained stable and reliable binding model, weak interaction such as Hbonds, vdw and ele were generated during the interaction. ABBA did not exhibit anti-L. monocytogenes or cytotoxicity effects at the effective inhibition concentrations, but it alleviated the cytotoxicity and inflammation mediated by LLO or bacteria, and protected G. mellonella from L. monocytogenes infection. These research results provide evident for the application of ABBA on combating L. monocytogenes infection.
Supplemental Materials
Supplementary data for this paper are available on-line only at http://jmb.or.kr.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Quereda JJ Morón-García A Palacios-Gorba C Dessaux C García-Del Portillo F Pucciarelli MG 2021 Pathogenicity and virulence of Listeria monocytogenes: a trip from environmental to medical microbiology Virulence 1225092545 http://doi.org/10.1080/21505594.2021.197552 10.1080/21505594.2021.19755234612177 PMC 8496543 · doi ↗ · pubmed ↗
- 2Valenti M Ranganathan N Moore LS Hughes S 2021 Listeria monocytogenes infections: presentation, diagnosis and treatment Br. J. Hosp. Med. (Lond)8216 https://doi.org/10.12968/hmed.2021.0107 10.12968/hmed.2021.010734726931 · doi ↗ · pubmed ↗
- 3Vázquez-Boland JA Kuhn M Berche P Chakraborty T Domínguez-Bernal G Goebel W 2001 Listeria pathogenesis and molecular virulence determinants Clin.Microbiol. Rev.14584640 https://doi.org/10.1128/CMR.14.3.584-640.2001 10.1128/CMR.14.3.584-640.200111432815 PMC 88991 · doi ↗ · pubmed ↗
- 4Demaître N Van Damme I De Zutter L Geeraerd AH Rasschaert G De Reu K 2020 Occurrence, distribution and diversity of Listeria monocytogenes contamination on beef and pig carcasses after slaughter Meat Sci.169108177 https://doi.org/10.1016/j.meatsci.2020.108177 10.1016/j.meatsci.2020.10817732544760 · doi ↗ · pubmed ↗
- 5Matle I Mbatha KR Madoroba E 2020 A review of Listeria monocytogenes from meat and meat products: epidemiology, virulence factors, antimicrobial resistance and diagnosis Onderstepoort J. Vet. Res.87e 1e 20 https://doi.org/10.4102/ojvr.v 87i 1.1869 10.4102/ojvr.v 87i 1.1869 PMC 756515033054262 · doi ↗ · pubmed ↗
- 6Jeong J Song H Kim WH Chae M Lee JY Kwon YK 2023 Tracking the contamination sources of microbial population and characterizing Listeria monocytogenes in a chicken slaughterhouse by using culture-dependent and -independent methods Front. Microbiol.141282961 https://doi.org/10.3389/fmicb.2023.1282961 10.3389/fmicb.2023.128296138098672 PMC 10720907 · doi ↗ · pubmed ↗
- 7Kayode AJ Okoh AI 2022 Antibiotic resistance profile of Listeria monocytogenes recovered from ready-to-eat foods surveyed in South Africa J. Food Prot.8518071814 https://doi.org/10.4315/JFP-22-090 10.4315/JFP-22-09036075088 · doi ↗ · pubmed ↗
- 8Lachtara B Wieczorek K Osek J 2023 Antimicrobial resistance of Listeria monocytogenes serogroups I Ia and I Vb from food and food-production environments in Poland J. Vet. Res.67373379 https://doi.org/10.2478/jvetres-2023-0050 10.2478/jvetres-2023-005037786847 PMC 10541657 · doi ↗ · pubmed ↗