Delimiting species boundaries in Hosta section Capitatae (Asparagaceae) using MIG-seq and morphological analyses: taxonomic revision with new taxa from Korea and Japan
Ami Oh, Ji Young Yang, Won Seok Lee, Takashi Shiga, Seiko Fujii, Shota Sakaguchi, Shukherdorj Baasanmunkh, Seung-Chul Kim, Hyeok Jae Choi

TL;DR
This study uses genetic and morphological data to revise the classification of Hosta species in Japan and Korea, identifying new taxa.
Contribution
The paper introduces two new Hosta taxa and provides a comprehensive taxonomic revision of the section Capitatae using MIG-seq and morphological analyses.
Findings
Three species and two varieties were identified in the section Capitatae, including two new taxa.
MIG-seq and PCA revealed clear genetic and morphological differentiation between the Kochi lineage, H. capitata, and H. nakaiana.
The Wando population was found to be embedded within the H. nakaiana cluster.
Abstract
Hosta capitata, which has recently been placed in the monotypic section Capitatae, and H. nakaiana, were originally described in Iya Valley, Japan and Mt. Baegun, Korea, respectively, and have been considered the same from a morphological perspective. However, considering the significant genetic distance between these groups, the identity of H. nakaiana deserves further investigations. Recently, the populations of H. capitata from Kochi, Japan, and Wando Island, Korea have been distinguished from the other groups by their distinctive morphological traits. On the basis of these observations, the present study aimed to provide a complete taxonomic revision of the section Capitatae in Korea and Japan based on extensive morphological observations and multiplexed inter-simple sequence repeats genotyping by sequencing (MIG-seq) analysis. Samples of the section Capitatae were collected from…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
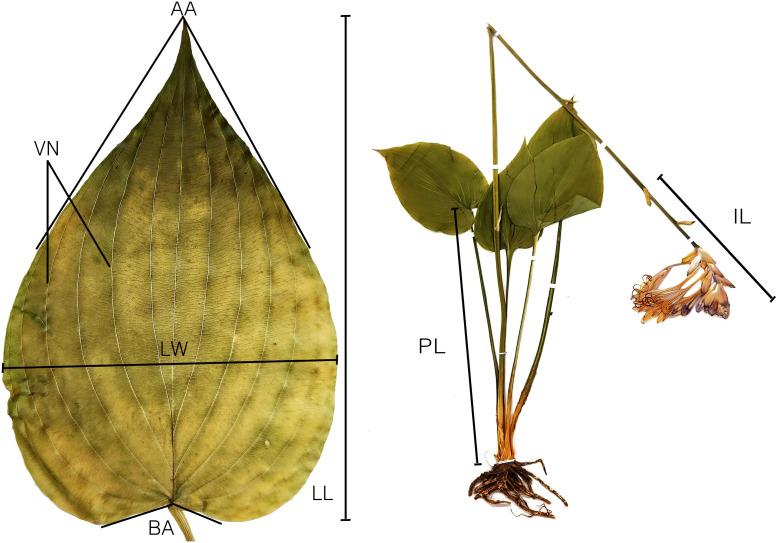 Figure 1
Figure 1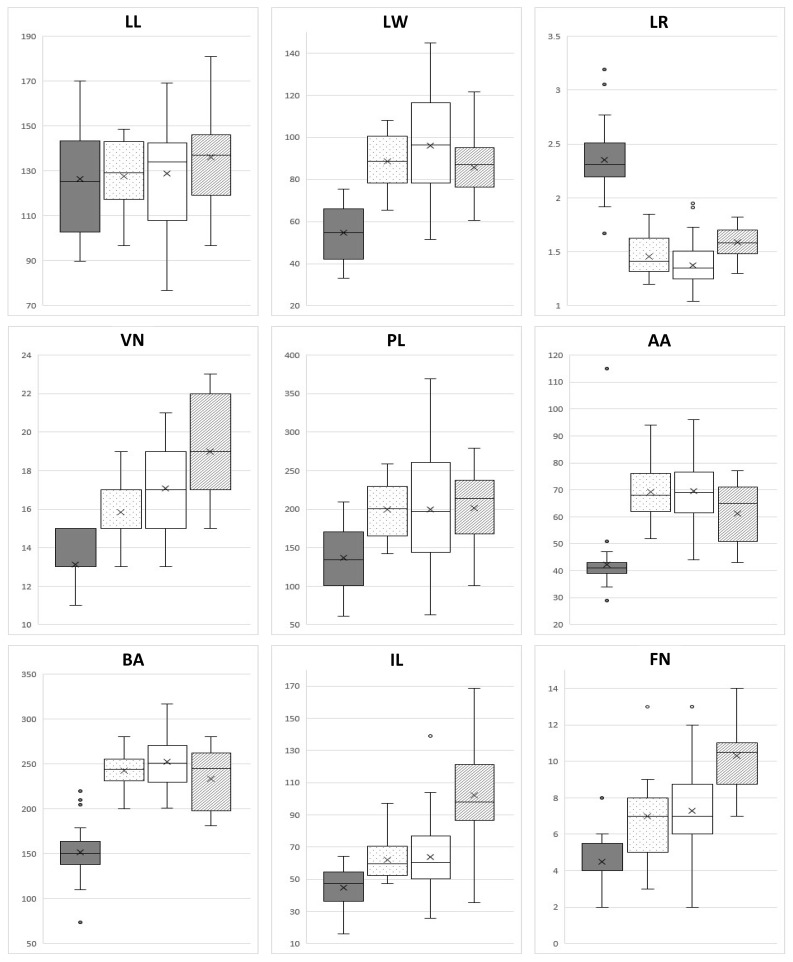 Figure 2
Figure 2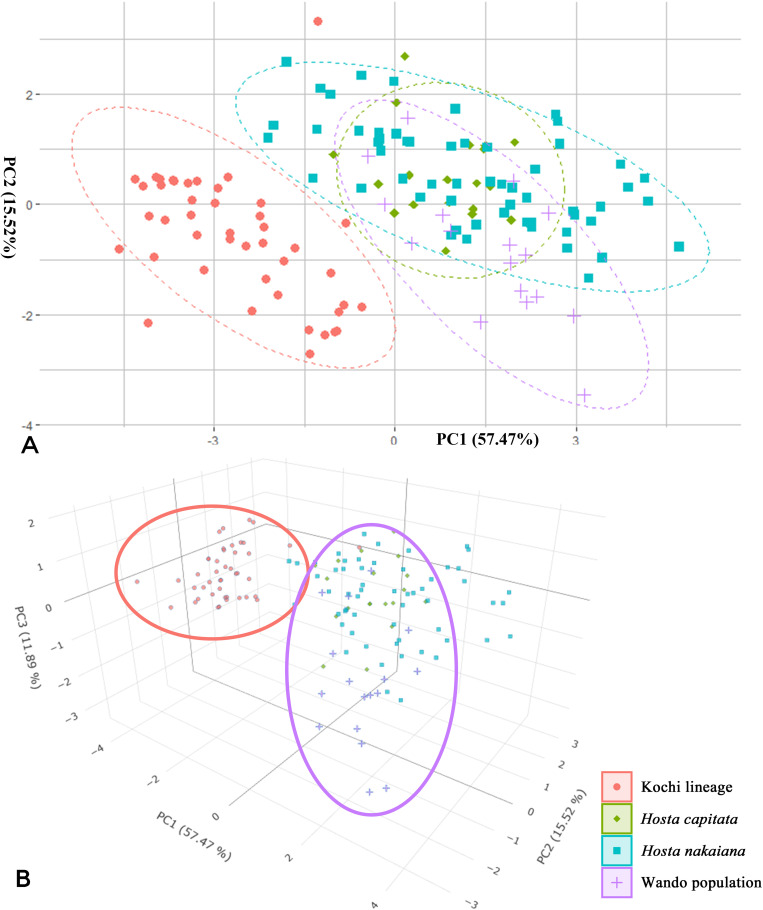 Figure 3
Figure 3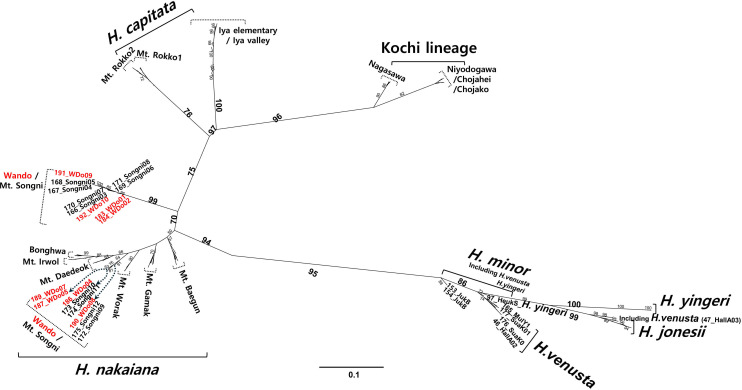 Figure 4
Figure 4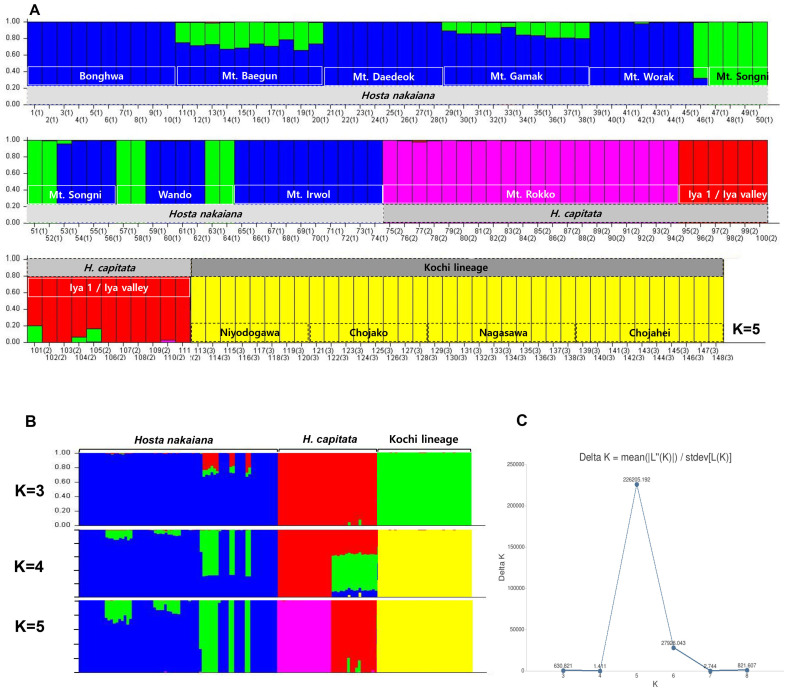 Figure 5
Figure 5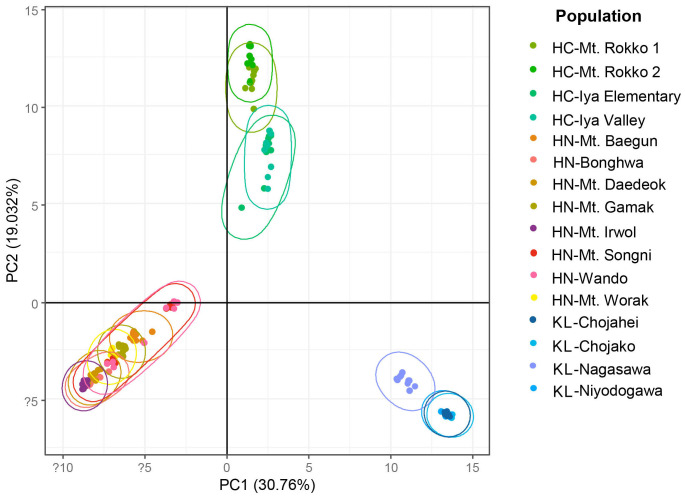 Figure 6
Figure 6 Figure 7
Figure 7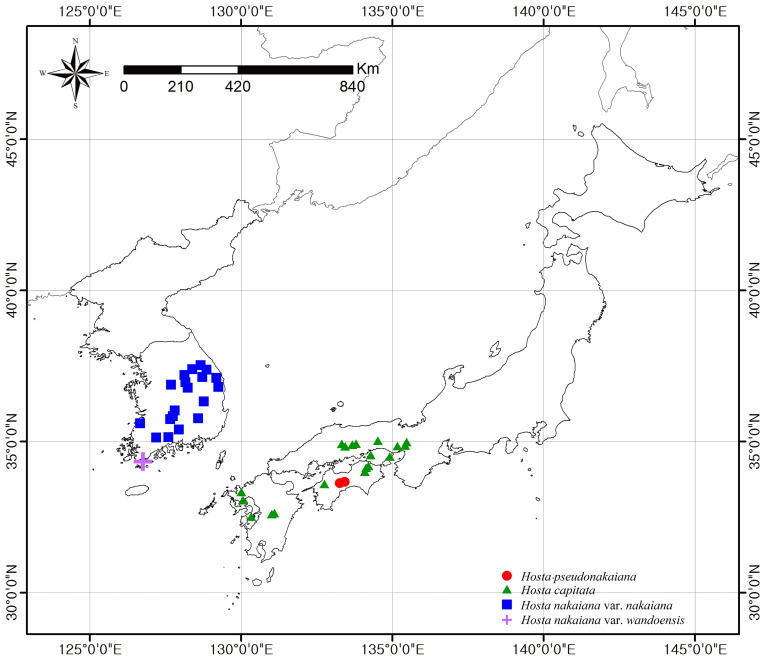 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12| Scientific name | Voucher ID | Locality | Latitude and longitude |
|---|---|---|---|
|
|
| Mt. Rokko, | Available on request |
|
|
| Mt. Rokko, | Available on request |
|
|
| Iya elementary school, | Available on request |
|
|
| Iya valley, | Available on request |
|
|
| Jeopdo Isl., | N34.37624 E126.29005 |
|
|
| Namhae natural forest, Gyeongsangnam-do, Korea | N34.75220 E128.01727 |
|
|
| Mt. Geumsan, Gyeongsangnam-do, Korea | N34.75228 E127.98214 |
|
|
| Mt. Bulmosan, Gyeongsangnam-do, Korea | N35.18017 |
|
|
| Mt. Cheongnyangsan, Gyeongsangbuk-do, Korea | N36.78724 E128.89820 |
|
|
| Mt. Haewolbong, Gyeongsangbuk-do, Korea | – |
|
|
| Mt. Jukbyeonsan, Gangwon-do, Korea | N38.30397 E128.46625 |
|
|
| Mt. Hanusan, Gyeongsangnam-do, Korea | – |
|
|
| Mt. Sinbulsan, Gyeongsangnam-do, Korea | – |
|
|
| Gasa forest, | N34.37150 E126.92563 |
|
|
| Bonghwa, Gyeongsangbuk-do, Korea | – |
|
|
| Mt. Baegun, | N35.10241 E127.65130 |
|
|
| Mt. Daedeok, | N37.23967 E128.91963 |
|
| HN-GA01 | Mt. Gamak, Gyeongsangnam-do, Korea | – |
|
| HN-WR01 | Mt. Worak, Chungcheongbuk-do, Korea | – |
|
| HN-IW01 | Mt. Irwol, Gyeongsangbuk-do, Korea | – |
|
|
| Mt. Songni, Chungcheongbuk-do, Korea | – |
| Wando population |
| Wando, | N34.34675 E126.69531 |
| Kochi lineage |
| Chojako, | Available on request |
| Kochi lineage |
| Chojahei, | Available on request |
| Kochi lineage |
| Niyodogawa, | Available on request |
| Kochi lineage |
| Nagasawa, | Available on request |
|
|
| Mt. Hallasan, | N33.35938 E126.46326 |
|
|
| Mt. Muryeongarioreum, | N33.36845 E126.69331 |
|
|
| Suakgyo Brdg., | N33.33699 E126.61111 |
|
|
| Mt. Chillaksan, Jeollanam-do, Korea | N34.68240 E125.42640 |
|
|
| Hongdo Isl., Jeollanam-do, Korea | – |
| Character | Measurement minimum (mean ± SD) and maximum | |||
|---|---|---|---|---|
| Kochi lineage |
|
| ||
| Other populations | Wando population | |||
| Leaf | ||||
| Blade length (LL, mm) | 89.7–170 (126.49±24.14) | 96.79–148.5 (127.69±15.86) | 76.76–169.18 (128.72±21.82) | 96.7–194.66 (136.16±24.21) |
| Blade width (LW, mm) | 33.16–75.27 (54.66±12.70) | 65.23–108.02 (88.69±13.11) | 51.54–144.89 (96.04±23.77) | 60.66–121.58 (85.83±14.07) |
| Apex angle (AA, °) | 29–115 (42.41±11.92) | 52–94 (69.26±10.26) | 44–96 (69.42±11.15) | 43–77 (61.24±10.66) |
| Base angle (BA, °) | 74–220 (152.07±27.10) | 200–280 (242.26±20.74) | 201–317 (252.40±27.91) | 181–280 (233.06±32.33) |
| Lateral vein number (VN, no.) | 11–15 (13.14±1.37) | 13–19 (15.84±1.76) | 13–21 (17.07±2.31) | 15–23 (19±2.83) |
| Ratio of blade length to width (LR, L:W) | 1.67–3.19 (2.35±0.27) | 1.20–1.85 (1.45±0.18) | 1.04–1.95 (1.38±0.19) | 1.30–1.82 (1.59±0.14) |
| Petiole length (PL, mm) | 60.82–208.97 (137.25±41.31) | 141.87–258.6 (199.90±33.68) | 62.32–368.7 (199.91±72.17) | 100.69–279.33 (201.44±47.68) |
| Flower | ||||
| Inflorescence length (IL, mm) | 16.2–64.11 (44.95±13.21) | 47.34–97.2 (62.00±13.18) | 25.7–139.04 (63.67±20.41) | 35.71–168.59 (102.10±29.99) |
| Flower number (FN, no.) | 2–8 (4.48±1.38) | 3–13 (7±2.25) | 2–13 (7.29±2.34) | 7–14 (10.29±1.83) |
| Character | Kochi lineage |
|
| |
|---|---|---|---|---|
| Other populations | Wando population | |||
| Leaf | ||||
| Shape | Narrowly ovate | Ovate to broadly ovate | Ovate to broadly ovate | Broadly ovate |
| Base | Rounded to subcordate | Subcordate to cordate | Subcordate to cordate | Subcordate to cordate |
| Apex | Acuminate to acute | Acute to obtuse | Acute to obtuse | Acute to obtuse |
| Inflorescence | ||||
| Shape | Compact spike-like raceme | Compact spike-like raceme | Compact spike-like raceme | Loose spike-like raceme |
| Flowering season | June to July | End of June to July | End of June to July | Early to end of June |
| Perianth | ||||
| Shape | Bell shape | Bell shape | Bell shape | Bell shape |
| Lobe | Lanceolate | Broadly ovate | Broadly ovate | Broadly ovate |
| Character | Component | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| LL | 0.2196 | -0.6251* | 0.3663 |
| PL | 0.3645* | -0.2397 | 0.3145 |
| LW | 0.4151* | -0.0365 | 0.2281 |
| VN | 0.3763* | -0.0963 | -0.0510 |
| AA | 0.3394 | 0.4379* | 0.0162 |
| BA | 0.3616* | 0.2869 | 0.0142 |
| IL | 0.2042 | -0.2699 | -0.6597* |
| FN | 0.2663 | -0.2375 | -0.5256* |
| LR | -0.3827* | -0.3712 | 0.0169 |
| Eigenvalue (%) | 57.47 | 15.52 | 11.89 |
| Cumulative of eigenvalue (%) | 57.47 | 72.99 | 84.88 |
| Source of variation | DF | Pillai | Approx. F | num DF | den DF | Pr(>F) |
|---|---|---|---|---|---|---|
| Lineage | 2 | 1.4683 | 12.426 | 28 | 126 | < 2.2e-16*** |
| Residuals | 75 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Mediterranean and Iberian flora and fauna
Introduction
1
The genus Hosta Tratt. is widely used in gardening because of its ground-covering, glossy foliage, leaf patterns, and showy purple-to-white flowers (Jones, 1989; Chung, 1990). It is characterized by a leaf base that is abruptly tapered into the petiole, six stamens, united tepals, and purple-to-white flowers (Chen and Boufford, 2000). With approximately 22–45 species, Hosta is naturally distributed in Korea, Japan, China, and the Russian Far East, mainly on rocky substrates, forest edges, or grasslands in alpine regions (Jones, 1989; Chen and Boufford, 2000; Chung, 2007; Yahara et al., 2023). Hosta was traditionally classified under Liliaceae (Maekawa, 1940; Fujita, 1976; Chung, 1990; Chung et al., 1991; Schmid, 1991) or Agavaceae (Moran, 1949; Takhtajan, 1980), but it has recently been placed in the subfamily Agavoideae in Asparagaceae (APG IV, 2016; Jo and Kim, 2017; Lee et al., 2018). The genera Hemerocallis, Agave, Hesperocallis, Leucocrimum, Yucca, Manfreda, and Camassia are the most closely related to Hosta (Chung and Jones, 1989; Liu et al., 2011).
The taxonomy within the genus Hosta is highly complex and confusing due to its natural and artificial hybridization, abundant cultivars, and extensively variable morphology (Lee et al., 2019; Yang et al., 2021; Yoo et al., 2021). Specifically, hybridization and introgression which are common in this genus, have complicated species delimitation of Hosta (Yang et al., 2021; Yoo et al., 2021). Many previous studies have revised the infrageneric classification of Hosta, reporting diverse taxonomic systems (Maekawa, 1940; Fujita, 1976; Schmid, 1991; Zonneveld and Van Iren, 2001). In general, three subgenera, Hosta, Bryocles, and Giboshi, have been recognized in Hosta (Schmid, 1991; Zonneveld and Van Iren, 2001; Chung, 2007; Yang et al., 2021) while different numbers of sections were identified within the genus (Schmid, 1991; Zonneveld and Van Iren, 2001). One of the most recent and frequently cited studies by Schmid (1991) reported ten sections in Hosta. Among these, Hosta sect. Lamellatae (F.Maek.) F.Maek. is morphologically characterized by an erect and ridged scape (Maekawa, 1938). The three species in this section, Hosta capitata (Koidz.) Nakai, H. minor (Baker) Nakai, and H. venusta F. Maek. are found in Korea and Japan (Maekawa, 1938; Chung, 2007; Tamura and Fujita, 2016). However, H. capitata has recently been placed in its monotypic sect. Capitatae (F.Maek.) J.C.Yang, due to its unique morphology, which is a compact spike-like raceme (Lee et al., 2018). Notably, in the genus Hosta, morphological characteristics such as the presence of rhizomes, stamen connection, shape of leaf, scape, inflorescence, and flower, have historically been employed in taxonomic studies and have supported a stable classification (Maekawa, 1938; Tamura and Fujita, 2016).
Hosta capitata and H. nakaiana F.Maek. were originally described in the Iya Valley, Shikoku, Japan (Nakai, 1930a) and Mt. Baegun, Jeollanam-do, Korea (Maekawa, 1935), respectively. However, from a morphological perspective, both species are considered the same, and thus, H. nakaiana is regarded as a synonym of H. capitata (Fujita, 1976; Tamura and Fujita, 2016; Jo and Kim, 2017). Therefore, H. capitata (=H. nakaiana) shows a disjunct distribution, occurring in the main mountain range (central and southern parts) of Korea and the western parts of Japan (Fujita, 1976; Chung et al., 1991).
In a chloroplast whole genome-based phylogenetic study by Yang et al. (2021), species belonging to the sect. Lamellatae s.l., including H. minor, H. venusta, and H. capitata (=H. nakaiana), did not form a single clade, and the monotypic sect. Capitatae was clearly established as a distinct evolutionary lineage. In terms of the pairwise sequence distance (K2P), the average distance between the Korean and Japanese populations of H. capitata (=H. nakaiana) was 0.00020 ± 0.00003. For comparison, the K2P between sister species in the genus Hosta, H. jonesii M.G.Chung and H. tsushimensis N.Fujita, and between H. ventricosa (Salisb.) Stearn and H. sieboldiana Engl. were 0.00026 and 0.00008, respectively. Additionally, the distance between H. minor and H. venusta, two species endemic to Korea, was 0.00003 ± 0.00002 (Yang et al., 2021). Consequently, the taxonomic identity of H. nakaiana remains controversial (Schmid, 1991; Sauve et al., 2005).
Recently, notable morphological variability has been recognized within H. capitata, where the population from Kochi, Japan, exhibits a clear morphological distinction from the other groups (personal observation). Specifically, the Kochi population is distinguished from other groups by narrower leaf and perianth lobes, and fewer flowers per inflorescence. This observation warrants further research to determine whether this population represents a distinct lineage. Meanwhile, during a field trip to Wando Island, Korea, we observed a population of H. capitata with a distinctively looser inflorescence, which deserves an in-depth investigation.
In this study, we combined morphological data with multiplexed inter-simple sequence repeats genotyping by sequencing (MIG-seq) (Suyama and Matsuki, 2015), a method well-suited for reconstructing phylogenetic relationships among closely related species, and population genetic analyses (Binh et al., 2018; Strijk et al., 2020; Zhang et al., 2020; Yahara et al., 2021; Kusuma et al., 2022; Tamaki et al., 2023; Yahara et al., 2023). This integrative approach was applied to revise the taxonomy of Hosta sect. Capitatae in Korea and Japan.
Materials and methods
2
Taxon Sampling
2.1
Samples of H. capitata (=H. nakaiana), including two putative new taxa, were collected from Japan and Korea during extensive field surveys between 2013 and 2022 (Table 1). Samples from Japan were collected from Mt. Rokko in Hyogo, Iya in Tokushima, and Kochi. Samples from Korea were collected from eight populations: Bonghwa, Mt. Baegun, Mt. Daedeok, Mt. Gamak, Mt. Worak, Mt. Songni, Mt. Irwol, and Wando. For the phylogenetic analysis, H. minor, H. yingeri, H. jonesii, and H. venusta were collected from their habitats in Korea (Table 1). Voucher specimens were deposited in the herbarium of Changwon National University, Republic of Korea (CWNU).
Morphological analysis
2.2
Morphological analyses were conducted using living plants from the habitats and herbarium specimens from the following herbaria: CWNU, KH, KYO, MBK, OSA, and TI (abbreviations are according to Thiers (2023)). The voucher specimens used for the measurement of morphological characteristics are marked with an asterisk (*) in the list of additional specimens examined. Photographs of the type specimens were also used for the comparative observation of key characteristics within the section. Some plants collected from these habitats were transplanted into an experimental field at Changwon National University to observe their growth patterns.
Nine quantitative characteristics, including leaf blade length (LL), leaf blade width (LW), leaf apex angle (AA), leaf base angle (BA), leaf lateral vein number (VN), ratio of leaf length to width (LR), petiole length (PL), inflorescence length (IL), and flower number (FN), were measured and statistically analyzed (Table 2, Figures 1, 2). Length and width were measured in 0.01 mm units using electronic Vernier calipers (Mitutoyo Corp. CD-15APX), and angles in 1° units using a protractor. The measured traits were organized into tables in CSV files and used for principal component analysis (PCA). The qualitative characteristics, such as the shape of the leaf, inflorescence, and perianth, were also investigated (Table 3).
Diagram showing the morphological characters of Hosta sect. Capitatae measured in this study. LL, blade length (mm); LW, blade width (mm); AA, apex angle (°); BA, base angle (°); VN, lateral vein number (no.); LR, ratio of blade length to width (L:W); PL, petiole length (mm); IL, inflorescence length (mm); FN, flower number (no.).
Quantitative data of morphological characteristics of four Hosta taxa. Boxplots show the median, 25th and 75th percentiles (box), 10th and 90th percentiles (whiskers), and outliers (closed circle). From left to right: Kochi lineage, H. capitata, H. nakaiana, Wando population. LL, blade length (mm); LW, blade width (mm); AA, apex angle (°); BA, base angle (°); VN, lateral vein number (no.); LR, ratio of blade length to width (L:W); PL, petiole length (mm); IL, inflorescence length (mm); FN, flower number (no.). Units of the y-axes are mm for LL, LW, PL, IL, degree (°) for AA, BA, and count (no.) for VN, FN.
PCA was performed for nine quantitative characters using FactoMineR v.1.04, stats v.4.1.3, and vegan packages v.2.0–10 in R v.4.1.3 (Figure 3, Table 4; Lê et al., 2008; Oksanen et al., 2013; R Core Team, 2022). The FactoMineR package was used to standardize numerical variables. The stats package was used to cluster the data of the nine quantitative factors. To assess the statistical reliability of the clusters, we performed a multivariate analysis of variance (MANOVA) using the vegan package (Table 5).
Cluster analysis based on quantitative morphological factors of four Hosta taxa. (A) Scatter plot of the first two principal components PC1-PC2. (B) Three-dimensional scatter plot. The red oval indicates the cluster of the Kochi lineage and the purple oval indicates the cluster of the Wando population.
DNA isolation, MIG-seq library construction, and genotyping
2.3
Total DNA was extracted from the silica gel-dried leaves using a DNeasy Plant Mini Kit (Qiagen, Carlsbad, CA, USA). The MIG-seq library for 192 samples of H. capitata, H. nakaiana, the Kochi lineage, H. venusta, H. minor, H. yingeri and H. jonesii was constructed using two-step amplification (Suyama and Matsuki, 2015). To amplify the target inter-simple sequence repeats (ISSR), we conducted the first PCR using ISSR primers developed by Suyama and Matsuki (2015). A second PCR was performed to add individual indices to each sample using barcoding primers designed by Suyama and Matsuki (2015). The amplicons were pooled and purified in selected sizes of 350–800 bp, and sequenced on an Illumina MiSeq platform (Illumina, San Diego, CA, USA) using the Illumina MiSeq Reagent Kit V3 (150 cycles, Illumina). The ‘DarkCycle’ option was used to skip the sequencing of SSR primer regions and anchors. Low-quality reads and extremely short reads containing index tags were removed from the next generation sequencing (NGS) data using FASTX Toolkit 1 (RRID: SCR_005534) and TagDust 1.12 (Lassmann et al., 2009). To maintain the quality of the dataset, 13 samples with extensive missing data were removed after screening using vcftools (Danecek et al., 2011). To identify SNPs in the processed reads, we used STACKS v.1.48 (Catchen et al., 2013) and generated data matrices in multiple formats (STURCTURE, Phylip, and variant call format (VCF)) for subsequent phylogenetic and population genetic analyses. Specifically, in STACKS, we applied sequential analyses that are ‘ustacks’, ‘cstacks’, and ‘sstacks’ with the following parameters: the minimum depth of coverage required to create a stack (m) = 3, maximum distance between stacks (M) = 2, and maximum mismatches between loci when building the catalog (n) = 2. In addition, we conducted ‘population’ analysis with the following parameters: at least 75% (–r 0.75) of the minimum percentage of individuals across populations, minimum two populations present to process a locus (–p 2), minimum minor allele frequency at a locus of 0.05 (–min-max 0.05), and maximum observed heterozygosity at a locus of 0.6 (–max-obs-het 0.6).
Phylogenetic and population structure analyses
2.4
A phylogenetic tree based on genome-wide 3,165 SNPs in the MIG-seq dataset for 176 Hosta samples was inferred using the maximum likelihood (ML) method implemented in IQ-TREE v.1.6.12 (Nguyen et al., 2015) with the best-fit model of “TVM+F+ASC+R2”. The bootstrap support value (BS) was calculated from 1,000 bootstrap replicates, using Ultrafast bootstrapping implemented in IQ-TREE. To determine the population structure of the section Capitatae, STRUCTURE v.2.3.4 (Pritchard et al., 2000) was used. The analysis was run for 147 samples of H. capitata, H. nakaiana, and the Kochi lineage, and the log-likelihood for each model was estimated using different numbers of populations (K = 2–9). The optimal K value was determined using the Delta K method described by Evanno et al. (2005) in Clumpak (Kopelman et al., 2015). Finally, to examine the genetic similarities and relationships among the individuals, Principal Coordinate Analysis (PCoA) was conducted by calculating the eigenvalues and eigenvectors using GenoDive v.3.06 (Meirmans, 2020). We used R v.4.1.3 (R Core Team, 2022) to construct the PCoA plot.
Results
3
Morphological characters
3.1
In our study, among the nine quantitative characteristics assessed, leaf blade width, ratio of leaf blade length to width, and angle of leaf apex and base exhibited clear separation between the Kochi lineage and the other groups of the section Capitatae (Table 2; Figure 2). Regarding qualitative characteristics, the shape of the inflorescence and flowering time showed distinct division between the Wando population and other groups (Table 3). PCA was performed using the nine quantitative trait data (Figure 3), with the first three principal components accounting for 84.88% of the total variance. PC1 accounted for 57.47% of the total variance, PC2 for 15.52%, and PC3 for 11.90%. When examining the contribution of leaf data to the principal components, PC1 showed substantial involvement, encompassing attributes such as leaf blade width, length-to-width ratio, base angle, petiole length, and vein number (Table 4). PC2 also exhibited contributions from the leaf data, specifically leaf blade length and apex angle (Table 4). In contrast, flower and inflorescence data, including inflorescence length and flower number per inflorescence, solely influenced PC3 (Table 4). The PCA results identified two groups: the Kochi lineage and others (Figure 3). Within the cluster other than the Kochi lineage, H. capitata showed high morphological similarity to H. nakaiana in Korea. Notably, the Wando population of H. nakaiana was slightly separated from the remaining taxa within the cluster. This group showed looser spike-like racemes and more flowers (Table 3). The morphological PCA results were supported by multivariate analysis of variance (MANOVA) (Table 5).
Phylogenetic tree reconstruction using MIG-seq
3.2
In the process of MIG-seq data generation, where the 1^st^ PCR and the 2^nd^ PCR were conducted, PCR products were successfully obtained for all the 96 primer combinations in the 2^nd^ PCR. The number of loci recovered ranged from 1,617 to 15,759 (Supplementary Table 1). In total, 63,613,860 raw reads (144,483 ± 52,080 reads per sample) were obtained using MIG-seq. After quality control, 4,165,030 reads (21,806 ± 8,550 reads per sample) were used for further analyses. To infer the phylogenetic relationships among the 176 accessions of Hosta species, including H. jonesii, H. minor, H. venusta, H. yingeri, and the section Capitatae (H. capitata, H. nakaiana, and the Kochi lineage), ML analysis was performed (Figure 4). The unrooted ML tree strongly supported the monophyly of the section Capitatae (94% bootstrap value; BS). The section Capitatae consisted of three distinct clusters: the Kochi lineage, H. capitata, and H. nakaiana, which were genetically distant from the remaining species. The cluster of the Kochi lineage was recovered as monophyletic (96% BS) and was composed of two geographically distinct groups from the Nagasawa and Shikoku/Niyodo regions. Hosta capitata, which was most closely related to the Kochi lineage in the tree, was paraphyletic and consisted of two geographically differentiated groups from Mt. Rokko and the Iya region. Within H. nakaiana, which was recovered as paraphyletic and included eight populations from Korea, most populations showed monophyly, whereas other populations, such as those from Wando and Mt. Songni, did not. Notably, two clades composed of samples from both Wando and Mt. Songni were embedded in H. nakaiana, with bootstrap values of 91% and 99%. Among the other Hosta species, H. minor, H. yingeri, H. venusta, and H. jonesii, H. jonesii alone was identified as monophyletic (99% BS). Hosta venusta, a Korean endemic species, was embedded in both H. yingeri and H. minor clusters, whereas an individual of H. yingeri was nested within the H. minor cluster. The H. yingeri cluster was sister to the H. jonesii cluster, and the cluster that included both H. yingeri and H. jonesii appeared to be sister to the H. minor/H. venusta cluster.
Unrooted Maximum likelihood (ML) tree generated using IQ-TREE based on the MIGseq-derived 3165 SNPs from 147 accessions of sect Capitatae (H. capitata, H. nakaiana, and Kochi lineage). Numbers on branches represent bootstrap support (BS) values of > 50%, based on 1000 replicates. Red letters indicate accessions collected from Wando, Korea.
Population structure analyses using MIG-seq
3.3
The genetic structures of 147 accessions from the three lineages of the section Capitatae, the Kochi lineage, H. capitata, and H. nakaiana, were determined using genetic assignment analysis (Figures 5A, B). The optimal K, based on the ΔK method, was 5 (Figures 5B, C). When K = 5 (Figure 5A), the individuals of H. nakaiana were assigned to two clusters: cluster 1 is represented in blue and cluster 2 in green, although most of the individuals were assigned to cluster 1. Individuals from Bonghwa, Mt. Daedeok, and Mt. Irwol were assigned to cluster 1, whereas all individuals from Mt. Baegun and Mt. Gamak showed slight admixture signals. The Mt. Songni and Wando populations contained three and four individuals that were completely assigned to cluster 1, whereas six and four individuals from each population were classified as pure cluster 2. In the case of H. capitata, individuals from Mt. Rokko and the Iya region were separately assigned to two clusters, cluster 3 represented in pink and cluster 4 in red. All the individuals of the Kochi lineage were completely assigned to cluster 5 in yellow.
(A) Bar plot of STRUCTURE analysis for 147 accessions of Hosta sect. Capitatae (H. capitata, H. nakaiana, and Kochi lineage) at ΔK = 5. (B) Comparative bar plots showing population structure at K = 3, 4, and 5. (C) Delta K distribution graph used to identify the best-fit K value using the method of Evanno et al. (2005) in Clumpak.
The genetic structures of the three lineages of the section Capitatae assessed using PCoA (Figure 6) showed genetic differentiation between H. capitata, H. nakaiana, and the Kochi lineage. The PC1 axis distinguished all three groups, whereas H. nakaiana and the Kochi lineage were separated from H. capitata along the PC2 axis. In the plot, H. nakaiana comprised continuously distributed genetic clusters, with some individuals from Wando and Mt. Songni located separately. In H. capitata, two genetically and geographically differentiated clusters were observed. Furthermore, in the Kochi lineage, two distinct genetic clusters were identified: the Nagasawa population and others. All the populations of the Kochi lineage, except for the Nagasawa population, were completely clustered.
Principal coordinate analysis (PCoA) score plot for 147 accessions of sect Capitatae (H. capitata (HC), H. nakaiana (HN) and Kochi lineage (KL)).
Discussion
4
Taxonomic revision of the section Capitatae
4.1
Hosta, popular in landscaping, medicine, and horticulture, has been long investigated taxonomically; however, species delimitation in this genus has been challenging (Li et al., 2012; Yang et al., 2021; Yoo et al., 2021). Notably, the monotypic section Capitatae has been only recently introduced into the genus (Lee et al., 2018), and its taxonomy, based on morphological or phylogenetic analyses, is largely lacking. In this study, the taxonomy of the section Capitatae was reassessed based on comprehensive morphological observations, phylogenetic inferences, and population structure analyses. We identified three species and two varieties in the section Capitatae. The Kochi lineage from Shikoku, Japan, was proposed as a new species and the Wando population from Korea as a new variety, contributing to the establishment of an advanced classification system for the section Capitatae.
Our findings indicate that the morphological characteristics analyzed in our study are of taxonomic utility in the section Capitatae. Specifically, the quantitative characteristics measured exhibited separation between the Kochi lineage in Shikoku, Japan, and the other groups inhabiting the southern part of Korea and Japan (Table 2, Figure 2). In addition, PCA results divided the section into two major groups: the Kochi lineage and remaining group (Figure 3). Phylogenetic and population structure analyses based on MIG-seq clarified and supported the morphology-based taxonomy of the section Capitatae (Figures 4, 5A). In the phylogenetic tree, the Kochi lineage displayed monophyly, supporting its distinct entity as a new taxon (Figure 4). Notably, in the tree, H. capitata and the Kochi lineage were closely related; however, the degree of genetic differentiation was higher between these two groups than between H. jonesii and H. yingeri, two well-established Hosta species (Figure 4, Yang et al., 2021). This observation supports that the Kochi lineage should be considered a new species in the section Capitatae. In addition, the results of the STRUCTURE analysis, in which the Kochi lineage was assigned to a single distinct cluster, suggested that this lineage is genetically independent and should be treated as a novel species (Figure 5A). Finally, PCoA successfully supported the clear genetic separation of the Kochi lineage from the remaining taxa (Figure 6). Notably, the detection of a distinct Nagasawa population in the Kochi lineage confirmed the results of the phylogenetic analysis (Figures 4, 6).
Although H. nakaiana was previously treated as a synonym of H. capitata (Tamura and Fujita, 2016; Jo and Kim, 2017) and our morphological analysis strongly supported the high similarity between them (Tables 2, 3; Figures 2, 3), our phylogenetic tree showed that H. capitata and H. nakaiana were genetically separate (Figure 4). STRUCTURE analysis revealed that these two groups had distinct genetic structures (Figure 5A). Furthermore, PCoA exhibited significant genetic separation between these taxa (Figure 6). A recent study by Yang et al. (2021) recognized that H. capitata populations in Japan and Korea are reciprocally monophyletic and genetically differentiated. Considering that the pairwise sequence distance between H. capitata in Japan and H. nakaiana in Korea is comparable to or larger than those between well-established Hosta species (Yang et al., 2021), it is highly likely that H. nakaiana is a different species from H. capitata and not a synonym. Notably, however, the sequence distances mentioned in our study, which are based on plastomes, should be interpreted with caution given that sequence divergence in plastomes of Hosta is quite low. Meanwhile, the identical morphology of these two groups indicates that H. nakaiana may be a cryptic species of H. capitata with divergent genetic and evolutionary traits.
The Wando population of H. nakaiana exhibits distinct morphological and phenological traits compared to the other groups, supporting its recognition as a novel taxon. In a previous phylogenetic analysis based on whole plastomes, the Wando population was genetically separated from the remaining H. capitata populations in Korea and Japan (Supplementary Figure 1). However, based on the results of the present study where the Wando population is deeply embedded in H. nakaiana, this group should be described as a new variety rather than a new species (Figure 4). Here, the Wando population is considered an incompletely distinct lineage of H. nakaiana. Meanwhile, phylogenetic analysis identified two clades containing samples from both Wando and Mt. Songni, indicating that these populations are genetically closely related (Figure 4). In addition, the STRUCTURE analysis revealed that the genetic compositions of the Mt. Songni and Wando populations share similarities (Figure 5A). Overall, the close relationship between the Wando and Mt. Songni populations in the phylogenetic tree, STRUCTURE plot, and PCoA plot suggests that the genetics, morphology, and evolution of these two populations should be investigated in parallel to better characterize this new variety from Wando.
In the previous studies on Hosta, the phylogenetic relationships among the species native to Korea, such as H. minor, H. venusta, H. yingeri, and H. jonesii, have been investigated in depth, and have shown both consistent and contrasting results (Lee et al., 2019; Yang et al., 2021; Yoo et al., 2021). At the same time, these species generally exhibited monophyly in these studies (Yang et al., 2021; Yoo et al., 2021), although Yoo et al. (2021) observed that the individuals of H. venusta were embedded within H. minor clade. In our phylogenetic analysis, regarding species genetically distant from the section Capitatae in the tree, H. yingeri cluster was sister to H. jonesii cluster, whereas the cluster comprising H. yingeri and H. jonesii was genetically close to H. minor/H. venusta group (Figure 4). Interestingly, the rough pattern of the genetic relationships aforementioned is consistent with the result of a previous phylogenomic analysis based on whole plastome sequences as well as that of another phylogenetic study that used the Hyb-Seq method, except that our analysis generally did not exhibit monophyly of the species (Yang et al., 2021; Yoo et al., 2021). However, in another phylogenetic analysis based on 246 nuclear genes, H. yingeri, the second most basal clade to the other Hosta species included, did not display sisterhood with H. jonesii (Yoo et al., 2021). Further phylogenetic analyses using more samples and various markers are needed to resolve this inconsistency and clarify the relationships between these Hosta species native to Korea.
Our study provides the most comprehensive framework for the classification of the section Capitatae, ultimately strengthening the taxonomy of the genus Hosta. This study also effectively complements Korean and Japanese flora by identifying the distributional patterns of newly reported taxa within the section Capitatae. Notably, in our study, the report of the new species and variety involves both morphological evaluation and molecular analysis, which greatly improved the quality and reliability of the report. Moreover, for the first time, our study used MIG-seq to successfully revise and update the taxonomy of the section Capitatae. Future studies employing genome-wide molecular data, extensive sampling, and state-of-the-art analysis tools may further elucidate the taxonomy of the Hosta sect. Capitatae based on the findings from our study.
Taxonomic treatment
4.2
A key to the species of Hosta sect. Capitatae
-
Leaves narrowly ovate, with leaf blade length:width ratio 1.67–3.19, perianth lobe lanceolate ————— H. pseudonakaiana
-
Leaves ovate to broadly ovate, with leaf blade length:width ratio 1.04–1.95, perianth lobe broadly ovate ————————— 2
-
Leaf blade length:width ratio 1.45 ± 0.18, 9.7–14.9 cm long, 6.5–10.8 cm wide, base truncate to subcordate ———— H. capitata
-
Leaf blade length:width ratio 1.38 ± 0.19, 7.7–16.9 cm long, 5.6–14.5 cm wide, base subcordate to cordate ——— H. nakaiana
Hosta pseudonakaiana Shiga & H.J.Choi, sp. nov. (Figures 7A, 8, 9). — TYPE: JAPAN. Kochi Pref.: Agawa-gun, Niyodogawa-cho, July 6, 2019 [fl], M.J.Choi & T.Shiga 190706-001* (holotype, KB; isotypes, four sheets, KH, KIOM, MBK, NGU)!.
Type specimens of Hosta sect. Capitatae. (A)H. pseudonakaiana (holotype KB); (B)H. capitata (holotype TI); (C)H. nakaiana var. nakaiana (holotype TI); (D)H. nakaiana var. wandoensis (holotype HIBR).
General distribution of Hosta sect. Capitatae species.
Herbs perennial, rhizomatous. Leaves basal, erect-patent, not shiny; petiole 6.1–20.9 cm long, base purple-dotted; leaf narrowly ovate, green, 9.0–17.0 × 3.3–7.5 cm, veins in 11–15 lateral pairs, base rounded to subcordate, apex acuminate to acute, papillose on abaxial surfaces. Scapes lamellar, ridged longitudinally. Inflorescences compact spike-like raceme, 2–8 flowered; bracts navicular, ovate, acuminate to acute at apex, purplish white, sometimes greenish near apex, deciduous at fruiting. Flowers bell-shaped; perianth pale purple to purple with dark purple nerves, lobes lanceolate; stamens 6, longer than perianth; anthers yellow with purple dots; ovary 3-loculed. Capsules cylindric. Seeds winged, black.
Note: This new species is morphologically very similar to H. capitata in having a ridged scape and compact spike-like racemes; however, several morphological characteristics of the leaves, inflorescences, and perianths differentiate them. In particular, H. pseudonakaiana differs in having narrowly ovate leaves vs. ovate to cordate leaves in H. capitata, 2–8 flowers per inflorescence vs. 3–13 flowers in H. capitata, and lanceolate perianth lobes vs. broadly ovate lobes in H. capitata (Tables 2, 3; Figures 9, 10). Schmid (1991) and Sauve et al. (2005) recognized this new species as being taxonomically distinct from H. capitata. Morphologically similar specimens were collected in Kagawa and Kumamoto Prefectures, and further analysis using SNP data, similar to the present study, will be necessary in the future to clearly identify these specimens.
Hosta pseudonakaiana. (A) habit; (B, C) leaf (B, adaxial; C, abaxial); (D) cross-section of scape; (E) flower; (F, G) perianth (F, adaxial; G, abaxial); (H) capsule; (I) seed. Photos from M.J.Choi & T.Shiga 190706-001.
Etymology: The specific epithet “pseudonakaiana” refers to the fact that this species had been misidentified as H. nakaiana in various herbaria.
Local name: Tosano-kanzashi-giboshi
Phenology: Flowering in June to July and fruiting in August to October.
Distribution and habitat: H. pseudonakaiana is endemic to Shikoku, Japan (Figure 8). This species grows under moist and half-shadowed forest conditions.
Additional specimens examined (Paratypes): JAPAN. Kochi Pref.: Agawa-gun, Ino-cho, Oomorigawa-keikoku (gorge), 7 Jul. 2013 [fl], Y.Kokami FOS-004908, 004909 (MBK)!; Nagasawa, 25 Jul. 1983 [fr], Y.Koukami M83-264 (MBK)!; 04 Jul. 2004 [fl], T.Nakamura et al., FOK-067205 (MBK)!; Choja-ko, 24 Aug. 2016 [fl], S.Fujii (NGU)!; 9 Oct. 2016 [fr], T.Shiga 9261-9264 (NGU)!; 06 Jul. 2019 [fl], M.J.Choi et al., 190706-Chojako(Kochi)-001* (CWNU)!; Choja-hei, 6 Jul. 2019 [fl], T.Shiga 11296 (NGU)!; Takaoka-gun, Niyodo-mura, 9 Aug. 1970 [fr], N.Fujita & K.Okamura 1 (KYO)!; Gyobuyabu, 9 Sep. 1970 [fr], N.Fujita & K.Okamura 2 (KYO)!; Shimanto-cho, Eshi, 4 Jul. 1995 [fl], Y.Koukami 95-130 (MBK)!; Tsuno-cho, Shiraishi-ko, 25 Aug 2016 [fr], S.Fujii, 160825-Takaoka(Kochi)-001* (CWNU)!; Tosa-gun, Motoyama-mura, 26 Jun. 1967 [fl], T.Yamanaka (KYO)!.
Hosta capitata (Koidz.) Nakai, Bot. Mag. (Tokyo) 44: 514 (Nakai, 1930b). (Figures 7B, 8, 10). — Basyonym: H. caerulea var. capitata Koidz., Bot. Mag. (Tokyo) 30: 326 (1916). — TYPE: JAPAN. Tokushima Pref.: Awa-shi, Mima-gun, Higashiiya-mura, June 29, 1915 [fl] (holotype, TI)!.
Hosta capitata. (A) habit; (B) inflorescence; (C) root; (D, E) leaf (D, adaxial; E, abaxial); (F) flower; (G, H) perianth (G, adaxial; H, abaxial); (I) capsule; (J) seed. Photos from H.J.Choi 190707-Rokko(Gobe)-001(A, G, H) and H.J.Choi 161008-Iya-001(B-F, I, J).
Herbs perennial, rhizomatous. Leaves basal, erect-patent, not shiny; petiole 14.2–25.9 cm long, base purple-dotted; leaf broadly ovate, green, 9.7–14.9 × 6.5–10.8 cm, veins in 13–19 lateral, base subcordate to cordate, apex acuminate to acute, papillose on abaxial surfaces. Scapes lamellar, ridged longitudinally. Inflorescences compact spike-like raceme, 3–13 flowered; bracts navicular, ovate, acute to obtuse at apex, purplish white, sometimes greenish near apex, deciduous at fruiting. Flowers bell-shaped; perianth pale purple to purple with dark purple nerves, lobes broadly ovate; stamens 6, longer than perianth; anthers yellow with purple dots; ovary 3-loculed. Capsules cylindric. Seeds winged, black.
Note: H. capitata was treated as the same species as H. nakaiana and H. pseudonakaiana in Hosta sect. Capitatae (Fujita, 1976; Tamura and Fujita, 2016; Jo and Kim, 2017). In previous studies, differences in a few traits, such as the shape of the leaf base and the ratio of leaf blade length to width, were treated as variations within a species. However, according to the MIG-seq results, the genetic distinction was clear within the taxa; therefore, they were treated as separate species (Figures 4-6). In Japan, the vernacular name “Kanzashi-giboshi” has traditionally been applied to H. capitata (e.g., Tamura and Fujita, 2016). This name was originally introduced in association with a description of H. nakaiana (Maekawa, 1935). Accordingly, the name “Iya-giboshi” (Koidzumi, 1915), which was historically linked to H. capitata, is more appropriate for this species.
Local name: Iya-giboshi (Koidzumi, 1915)
Phenology: Flowering from the end of June to July and fruiting from August to October.
Distribution and habitat: H. capitata is endemic to southern Honshu, Shikoku, and Kyushu, Japan (Figure 8). This species is typically found in rocky valleys with limestone, granite, serpentine outcrops, and forest margins with open canopies (Fujita, 1976; Tamura and Fujita, 2016).
Additional specimens examined: JAPAN. Hyogo Pref.: Kobe-si, Kita-ku, Kamakura-kyo, 27 Jul. 1977 [fr], S.Hosomi 17670 (KYO)!; Yamada-cho, Aina, 25 Jun. 1978 [fl], T.Kobayashi 9381 (KYO)!; 9 Jul. 1978 [fl], N.Fukuoka 9757 (KYO)!; 20 Jul. 2019 [fl], T.Kobayashi 57482 (KYO)!; Dojyo-cho, Shimotanaka, 20 Jun. 1993 [fl], T.Fujii 3280 (OSA)!; Nada-ku, Higashirokko, Mt. Rokko, 16 Aug. 1992 [fr], T.Kobayashi 21247 (KYO)!; 2 Jul. 1994 [fl], T.Kobayashi 25704 (KYO)!; Rokkosan-cho, Mt. Minami-rokko, 04 Sep 2016 [fr], T.Kurazono 160904-Minamirokko-001* (CWNU)!; 07 Jul 2019 [fl], H.J.Choi et al., 190707-Rokko(Gobe)-001** (CWNU)!; Mt. Nishitaniyama, 04 Sep 2016 [fr], T.Kurazono 160904-Nishitaniyama-001 (CWNU)!; Takarazuka-si, Takedao, 16 Sep. 1991 [fr], T.Kobayashi 19596 (KYO)!; Ako-gun, Kamigori-cho, 2 Jul. 1989 [fl], S.Fujii 2743 (OSA)!; 15 Aug. 1989 [fr], S.Fujii & M.Kuribayashi 2782 (KYO)!; 11 Oct. 1990 [fr], H.Nagamasu 4401 (KYO)!; Sep. 1989 [fr], M.Kuribayashi 230 (KYO)!; 27 Jul. 1991 [fr], T.Kobayashi 19061 (KYO)!; Oct. 1993 [fr], T.Takahashi & M.Sawada 3035 (OOM)!; 24 Jun. 1999 [fl], T.Takahashi 2698 (KYO)!; 29 Jun. 1992 [fl], N.Fukuoka 13946 (OSA)!; 2 Jul. 1989 [fl], M.Kuribayashi 204 (OSA)!; 17 Jun. 1995 [fl], S.Fujii 4280 (OSA)!; border of Kamigori-cho and Mikaduki-cho, Todoma valley, 4 Jun. 2002 [fl], S.Fujii 9190 (MBK)!; Tsuna-gun, Hokudan-cho, Mt. Joryuji, 30 Jun. 1991 [fl], T.Kobayashi 18778 (KYO)!; Sanda-shi, Yayoiga-oka, Fukada-park, 29 May 1996 [fl], T.Fujii 5701 (OSA)!. Okayama Pref.: Mitsu-gun, Kamogawa-cho, Enjyo, 16 Oct. 1971 [fr], N.Fujita 218 (KYO)!; Kamifusa-gun, 26 Jun. 1989 [fl], Y.Kohata & S.Kano (KYO)!; Takahashi-shi, Nariwa-cho, 8 Jul. 1989 [fl], I.Okubo 327 (KYO)!. Hiroshima Pref.: Hiba-gun, Tojo-cho, 26 Jul. 1987 [fr], N.Kurosaki 16168 (KYO)!. Tokushima Pref.: Miyoshi-city, Ikeda-cho, Ori, 08 Oct 2016 [fr], H.J.Choi et al.*, 161008-Iya-001** (CWNU)!; Mima-gun, Higashi-Iya-yama-son, 29 Jun. 1915 [fl], G.Koidzumi (isotype, KYO)!; Waki-machi, Shimizu, 15 Oct. 1971 [fr], N.Fujita 258 (KYO)!. Kagawa Pref.: Takamatsu-si, shionoe-cho, Mt. Otaki, 13 Sep. 1961 [fl], S.Sakaguchi (KYO)!. Ehime Pref.: Kita-gun, Uchiko-cho, 19 Jul. 1973 [fl], T.Yamanaka 64945 (KYO)!. Saga Pref.: Imari-si, Ohkawauchi-cho, 6 Sep. 2021 [fr], Y.Inoue SS210906 (KYO)!; Fujitsu-gun, Tara-cho, Mt. Tara, 1 Nov. 1919 [fr], I.Kawauchi (KYO)!; 13 Oct. 1992 [fr], Z.Tashiro (KYO)!; 22 Jul. 2011 [fr], Y.Inoue (KYO)!. Kumamoto Pref.: Yatsushiro-si, Izumi-mura, 10 Aug. 1977 [fl], H.Tomita 1295 (KYO)!. Miyazaki Pref.: Higashiusuki-gun, Shiiba-mura, Kiritachi-goe, Mt. Shiroiwa, 15 Aug. 1961 [fr], M.Hotta 6475 (KYO)!; 24 Sep. 1988 [fr], M.Furuse 54560 (KYO)!.
Hosta nakaiana F.Maek., J. Jap. Bot. 11(10): 687. (1935) (Figure 7C). — TYPE: KOREA. Jeollanam-do: Mt. Baegunsan, July 6, 1935 [fl] ((holotype, TI)!).
A key to varieties of Hosta nakaiana
-
Inflorescence compactly spike-like, scape lamellar ridged, 2.6 (6.4 ± 2.0) 13.9 cm long, 2 (7.3 ± 2.3) 13 flowers per inflorescence, flowering at the end of June to July ——————— var. nakaiana
-
Inflorescence loosely spike-like, scape smooth, 3.6 (10.2 ± 3.0)16.9 cm long, 7 (10.3 ± 1.8) 14 flowers per inflorescence, flowering at early to end of June ———————————— var. wandoensis
Hosta nakaiana var. nakaiana (Figures 8, 11).
Hosta nakaiana var. nakaiana. (A) habit; (B) inflorescence; (C, D) leaf (C, adaxial; D, abaxial); (E) flower; (F, G) perianth (F, adaxial; G, abaxial); (H) capsule; (I) seed. Photos from H.J.Choi 150806-Gwangyangsi(Baegunsan)-046.
Herbs perennial, rhizomatous. Leaves basal, erect-patent, not shiny; petiole 6.2–36.9 cm long, base purple-dotted; leaf broadly ovate, green, 7.7–16.9 × 5.2–14.5 cm, veins in 13–21 lateral, base subcordate to cordate, apex acuminate to acute, papillose on abaxial surfaces. Scapes lamellar, ridged longitudinally. Inflorescences compact spike-like raceme, 2–13 flowered; bracts navicular, ovate, acute to obtuse at apex, purplish white, sometimes greenish near apex, deciduous at fruiting. Flowers bell-shaped; perianth pale purple to purple with dark purple nerves, lobes broadly ovate; stamens 6, longer than perianth; anthers yellow with purple dots; ovary 3-loculed. Capsules cylindric. Seeds winged, black.
Note: Despite their morphological similarities (Tables 2, 3, Figure 2), H. nakaiana and H. capitata were genetically distinct groups based on the results of the MIG-seq analyses (Figures 4-6). In addition, H. nakaiana and H. capitata are distributed in Korea and Japan, respectively.
Local name: Ilwol-bibichu
Phenology: Flowering from the end of June to July and fruiting from August to October.
Distribution and habitat: H. nakaiana var. nakaiana is endemic to Korea (Figure 8). This species grows in rocky valleys, mountain summits, ridges, and slopes (Jo and Kim, 2017).
Additional specimens examined: KOREA. Gangwon-do: Yeongwol-gun, Sangdong-eup, Taebaeksan-ro, 3432-74, 37°04’25.3”N, 128°49’53.3”E, 1,174 m a.s.l., 27 Jul. 2008 [fl], J.O.Hyun, 1001019* (KH)!; 37°04’02.4”N, 128°50’00.5”E, 1,263 m a.s.l., 27 Jul. 2008 [fl], J.O.Hyun, 1001023* (KH)!; Yeongwol-eup, Munsan-ri, San161, 37°16’34.9”N, 128°29’58.5”E, 769 m a.s.l., 17 Jul. 2008 [fl], J.O.Hyun & H.J.Gwon, NAPI-0414* (KH)!; Jeongseon-gun, Yeoryang-myeon, Bongjeong-ri, San1, 37°26’34.7”N, 128°45’30.4”E, 1,053 m a.s.l., 30 Jul. 2010 [fl], K.S.Kim & H.J.Kim, K0801020* (KH)!; Jeongseon-eup, 26 Jun. 2013 [fl], B.U.Oh, KH00719* (KH)!; Taebaek-si, Changjuk-dong, san76, 37.23967°N, 128.91963°E, 1,161 m a.s.l., 29 Jul. 2016 [fl], H.J.Choi et al., 160729-Taebaeksi(Daedeoksan)-001* (CWNU)!. Chungcheongbuk-do: Jecheon-si, Cheongpung-myeon, Hakhyeon-ri, 289-1, 37°00’40”N, 128°12’56”E, 279 m a.s.l., 14 Aug. 2006 [fr], G.H.Nam & J.E.Koh, CHJ60438* (KH)!; San6-1, 37°01’36.4”N, 128°13’22.8”E, 780 m a.s.l., 18 Oct. 2006 [fr], S.H.Park et al., ParkSH63602* (KH)!; Goesan-gun, Chilseong-myeon, Saeun-ri, San5-1, 36°44’06.8”N, 127°53’11.7”E, 553 m a.s.l., 21 Jun. 2006, W.K.Paik et al., Sangjusi (Songnisan)-060621-112* (KH)!; Goesan-gun, Mt. Songnisan, 26 Jun. 2013 [fl], B.U.Oh et al., Goesangun-130627-133* (KH)!; 5 Aug. 2013 [fr], B.U.Oh et al., Goesangun-130805-008* (KH)!; Boeun-gun, Mt. Songnisan, 11 Oct. 2002 [fr], Y.M.Kim et al., L-60077* (KH)!; 23 Jun. 2013 [fl], B.U.Oh et al., Boeungun-130623-012* (KH)!. Jeollabuk-do: Muju-Gun, Mupung-myeon, Samgeo-ri, San1-11, 35°52’52.3”N, 127°50’26.9”E, 938 m a.s.l., 23 Jun. 2008 [fl], J.O.Hyun et al., NAPI-0293* (KH)!; Jangsu-gun, Gyebuk-myeon, Yangak-ri, San35-2, 35°46.563’N, 127°41.061’E, 1,252 m a.s.l., 07 Aug. 2013 [fr], H.J.Choi et al., 130807_hjchoi065* (KH)!; Wanju-gun, Mt. Moaksan, Bukbong-Moakjeong, 30 Jun. 2013 [fl], B.U.Oh, 130630-Moaksan-027* (KH)!; 30 Jun. 2013 [fl], B.U.Oh, 130630-Moaksan-023* (KH)!; Gochang-gun, Asan-myeon, Samin-ri, San35, 35.51770°N, 126.57699°E, 423 m a.s.l., 28 Sep. 2013 [fr], H.J.Choi et al., 130928-Gochanggun(Seonunsan)-020* (CWNU)!. Jeollanam-do: Suncheon-si, Juam-myeon, Ullyong-ri, San142. 35.116342°N, 127.204698°E, 193 m a.s.l., 24 Jun. 2009 [fl], Y.H.Cho et al., WR-090624-159* (KH)!; Gwangyang-si, Jinsang-myeon, Beakhak-ro, 1243, 35°06’22”N, 127°37’19”E, 1,205 m a.s.l., 12 Aug. 2010 [fr], D.O.Lim et al., LIM1462* (KH)!; Eochi-ri, San308, 35.10805°N, 127.62758°E, 1,084 m a.s.l., 06 Aug. 2015 [fr], H.J.Choi et al., 150806-Gwanyangsi(Baegunsan)-046* (CWNU)!. Gyeongsangbuk-do: Yecheon-gun, Yongmun-myeon, Nosa-ri, 290. 36°40’55.15”N, 128°21’41.11”E, 236 m a.s.l., 06 Jul. 2008 [fl], C.G.Jang et al., VP-KNU-368061-0011* (KH)!; Uiseong-gun, Gaeum-myeon, Hyeolli-ri, 11-6, 36°12’12.6”N, 128°47’45.5”E, 879 m a.s.l., 14 Jul. 2006 [fl], C.G.Jang, Jang310* (KH)!; Yeongyang-gun, Subi-myeon, Suha-ri, San26-1, 36°49’03.4”N, 129°16’08.9”E, 396 m a.s.l., 31 Aug. 2013 [fr], K.H.Bae et al., KH-130148** (KH)!; Bonghwa-gun, Chunyang-myeon, Seobyeok-ri, San103, 37°01’16.8”N, 128°47’12.5”E, 992 m a.s.l., 19 Jul. 2010 [fl], S.H.Oh et al., C100836 (KH)!; Seokpo-myeon, Banya-gil, 176-37, 37°02’15.1”N, 129°09’77.9”E, 883 m a.s.l., 20 May 2010 [fl], K.H.Bae et al., KH-1000181* (KH)!; Mungyeong-si, Dongno-myeon, Saengdal-ri, 493, 36°48’58.4”N, 128°15’40.6”E, 761 m a.s.l., 14 Jul. 2006 [fl], W.K.Paik et al., Mungyeongsi (Hwangjangsan)-060714-808* (KH)!; Seongju-gun, Geumsu-myeon, Muhak-ri, Mt. Baebawisan, 13 Oct. 2007 [fr], B.U.Oh, 071013-Baebawisan-009* (KH)!; Gimcheon-si, Daedeok-myeon, Deoksan-ri, San42-4, 35.92462°N, 127.90825°E, 495 m a.s.l., 29 Jun 2013, H.J.Choi et al., 130629-Gimcheonsi(Daedeoksan)-019* (CWNU)!; Daehang-myeon, Jurye-ri, San1, 36.10550°N, 127.96708°E, 923 m a.s.l., 02 Jul 2012 [fl], H.J.Choi et al., 120702-Gimcheonsi(Hwangaksan)-543* (CWNU)!; Daegu-si, Dalseong-gun, Gachang-myeon, Samsan-ri, San171-1, 35.72109°N, 128.68133°E, 452 m a.s.l., 28 Jun 2013 [fl], H.J.Choi et al., 130628-Daegusi(Sangwonsan)-079* (CWNU)!. Gyeongsangnam-do: Sancheong-gun, Samjang-myeon, Honggye-ri, San336, 35°20’44.97”N, 127°51’24.31”E, 396 m a.s.l., 20 Jun. 2007 [fl], J.C.Yang, YangJC 70056* (KH)!; Danseong-myeon, Un-ri, San147, 35°20’51.51”N, 127°52’55.14”E, 687 m a.s.l., 27 Jun. 2007 [fl], J.C.Yang, YangJC 70199* (KH)!; San1, 35°20’41.03”N, 127°53’34.81”E, 360 m a.s.l., 27 Jun. 2007 [fl], J.C.Yang, YangJC 70206* (KH)!; Sangcheong-eup, Nae-ri, San204, 35°22’48”N, 127°52’37”E, 200 m a.s.l., 20 Jun. 2007 [fl], J.C.Yang, YangJC 70011* (KH)!; San158-12, 35°23’00.83”N, 127°52’16.13”E, 350 m a.s.l., 28 Jun. 2007 [fl], J.C.Yang, YangJC 70274* (KH)!; Geochang-gun, Goje-myeon, Bongsan-ri, San252, 35°52’29.0”N, 127°51’20.9”E, 963m a.s.l., 24 Jun. 2008 [fl], J.O.Hyun et al., NAPI-0304* (KH)!.
Hosta nakaiana var. wandoensis H.J.Choi, var. nov. (Figures 7D, 8, 12). — TYPE: KOREA. Jeollanam-do: Wando-gun, Wando-eup, Daesin-ri, San8-1, 34.34675°N, 126.69531°E, 516 m a.s.l., June 23, 2021 [fl], Wando-210623-001* (holotype, HIBR; isotypes, three sheets, KB, KH, KIOM)!.
Hosta nakaiana var. wandoensis. (A) habit; (B) inflorescence; (C) root; (D, E) leaf (D, adaxial; E, abaxial); (F) flower; (G, H) perianth (G, adaxial; H, abaxial); (I) capsule; (J) seed. Photos from H.J.Choi Wando-210623-001.
Herbs perennial, rhizomatous. Leaves basal, erect-patent, not shiny; petiole 10.1–27.9 cm long, base purple-dotted; leaf broadly ovate, green, 9.7–19.5 × 6.1–12.2 cm, veins in 15–23 lateral, base subcordate to cordate, apex acuminate to acute, papillose on abaxial surfaces. Scapes lamellar ridged. Inflorescences loose, spike-like racemes, 7–14 flowered; bracts navicular, ovate, acute to obtuse at apex, purplish-white, sometimes greenish near apex, deciduous at fruiting. Flowers bell-shaped; perianth pale purple to purple with dark purple nerves, lobes broadly ovate; stamens 6, longer than perianth; anthers yellow with purple dots; ovary 3-loculed. Capsules cylindric. Seeds winged, black.
Note: This new variety is morphologically very similar to H. nakaiana var. nakaiana in having a ridged scape and bell-shaped flowers, but several morphological characteristics of the shapes of leaves and inflorescences easily differentiate them (Figures 7, 11, 12): var. wandoensis differs by having broadly ovate leaves and cordate base vs. ovate to cordate leaves and subcordate to cordate base in var. nakaiana. In addition, flowering from early to the end of June vs. flowering from the end of June to July in var. nakaiana and 7–14 flowers per inflorescence with a loose spike-like raceme vs. 2–13 flowers per inflorescence with a compact spike-like raceme in var. nakaiana (Tables 2, 3).
Etymology: The specific epithet was derived from the name of the type locality (Wando Island).
Local name: Wando-bibichu
Phenology: Flowering in June and fruiting in July to August.
Distribution and habitat: H. nakaiana var. wandoensis is endemic to Korea (Wando, Jeollanam-do; Figure 8). This species grows in shaded mountain ranges and open habitats on mountain summits.
Additional specimens examined (Paratypes): KOREA. Jeollanam-do: Wando-gun, Wando-eup, Daesin-ri, San8-1, 34.34675°N, 126.69531°E, 516 m a.s.l., 19 Jun 2019 [fl], H.J.Choi et al., 190619-Wandogun(Sangwangbong)-001* (CWNU)!*; 20 Jun 2025 [fl], H.J.Choi et al., 250620-Wandogun(Sangwangbong)-001 (CWNU)!.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1APG IV (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants. Bot. J. Linn. Soc 181, 1–20. doi: 10.1111/boj.12385 · doi ↗
- 2Binh H. T. Ngoc N. V. Tagane S. Toyama H. Mase K. Mitsuyuki C. . (2018). A taxonomic study of Quercus langbianensis complex based on morphology and DNA barcodes of classic and next generation sequences. Phyto Keys 7, 37–70. doi: 10.3897/phytokeys.95.21126, PMID: 29674926 PMC 5904329 · doi ↗ · pubmed ↗
- 3Catchen J. Hohenlohe P. A. Bassham S. Amores A. Cresko W. A. (2013). Stacks: an analysis tool set for population genomics. Mol. Ecol. 22, 3124–3140. doi: 10.1111/mec.12354, PMID: 23701397 PMC 3936987 · doi ↗ · pubmed ↗
- 4Chen X. Boufford D. E. (2000). Hosta. Flora of China. Available online at: https://flora.huh.harvard.edu/China/PDF/PDF 24/hosta.pdf [Accessed March 15, 2024].
- 5Chung M. G. (1990) in A biosystematic study of the genus Hosta Tratt. (Liliaceae/Funkiaceae) in Korea. [Ph. D. Thesis]. [Athens (GA)]: University of Georgia.
- 6Chung Y. C. (2007). “ Hosta,” in The Genera of Vascular Plants of Korea. Flora of Korea Editorial Committee. Ed. Park C.-W. ( Academy Publishing Co., Seoul), 1304–1306.
- 7Chung M. G. Jones S. B.Jr (1989). Pollen morphology of Hosta Tratt.(Funkiaceae) and related genera. Bull. Torrey Botanical Club 116, 31–44. doi: 10.2307/2997107 · doi ↗
- 8Chung M. G. Jones S. B. Hamrick J. L. (1991). Morphometric and isozyme analysis of the genus Hosta (Liliaceae) in Korea. Plant Species Biol. 6, 55–69. doi: 10.1111/j.1442-1984.1991.tb 00210.x · doi ↗