Characterization of two reductases MaLAR and MaANR revealed their roles in proanthocyanidin biosynthesis in mulberry
Zhiheng Feng, Peijun Li, Guang Yang, Yining Wang, Mengqi Li, Jiangting Wu, Nan Chao, Li Liu

TL;DR
This study identifies two enzymes, MaLAR and MaANR, that are crucial for proanthocyanidin biosynthesis in mulberry, affecting fruit quality and stress defense.
Contribution
The study functionally characterizes MaLAR and MaANR in mulberry, revealing their distinct roles in proanthocyanidin biosynthesis.
Findings
MaLAR and MaANR were cloned and confirmed to catalyze the formation of catechin and epicatechin, respectively.
Knock-down of either gene reduced proanthocyanidin content in mulberry leaves, while overexpression increased it in Arabidopsis.
MaLAR and MaANR show distinct expression patterns during fruit ripening, influencing proanthocyanidin composition.
Abstract
Proanthocyanidins (PAs), polymers of flavan-3-ols, are crucial for the sensory quality and stress defense of mulberry (Morus spp.). The biosynthesis of their monomers, catechin and epicatechin, are catalyzed by leucoanthocyanidin reductase (LAR) and anthocyanidin reductase (ANR), respectively, representing key rate-limiting steps that determine PA composition and abundance. In this study, systematic functional analyses including phylogenetic analysis, quantified spatio-temporal expression profiles during fruit development (S1-S4 stages), in vitro enzymatic assay, knock-down using Virus-Induced Gene Silencing (VIGS) in mulberry leaves and heterologous overexpression in Arabidopsis thaliana were conducted to reveal their roles in proanthocyanidin biosynthesis in mulberry. Results showed that MaLAR (969 bp) and MaANR (1014 bp) were successfully cloned and phylogenetically conserved.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
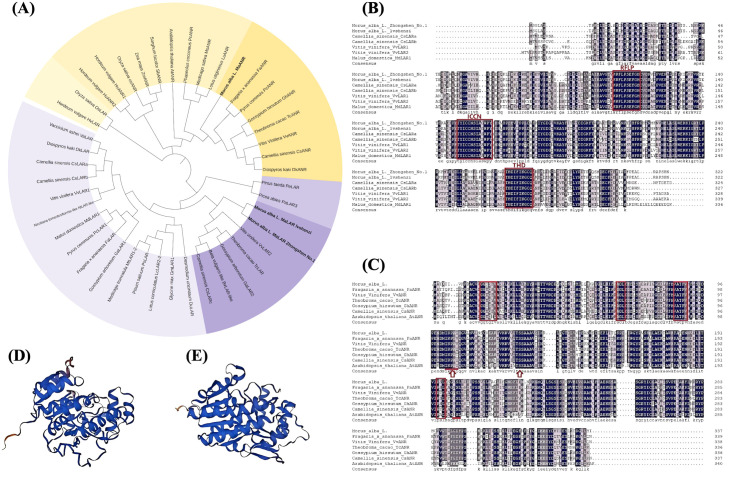 Figure 1
Figure 1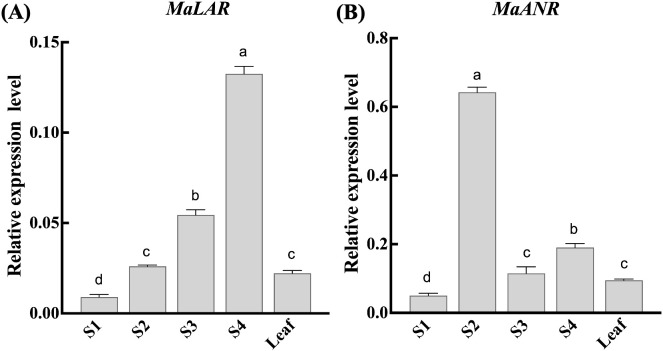 Figure 2
Figure 2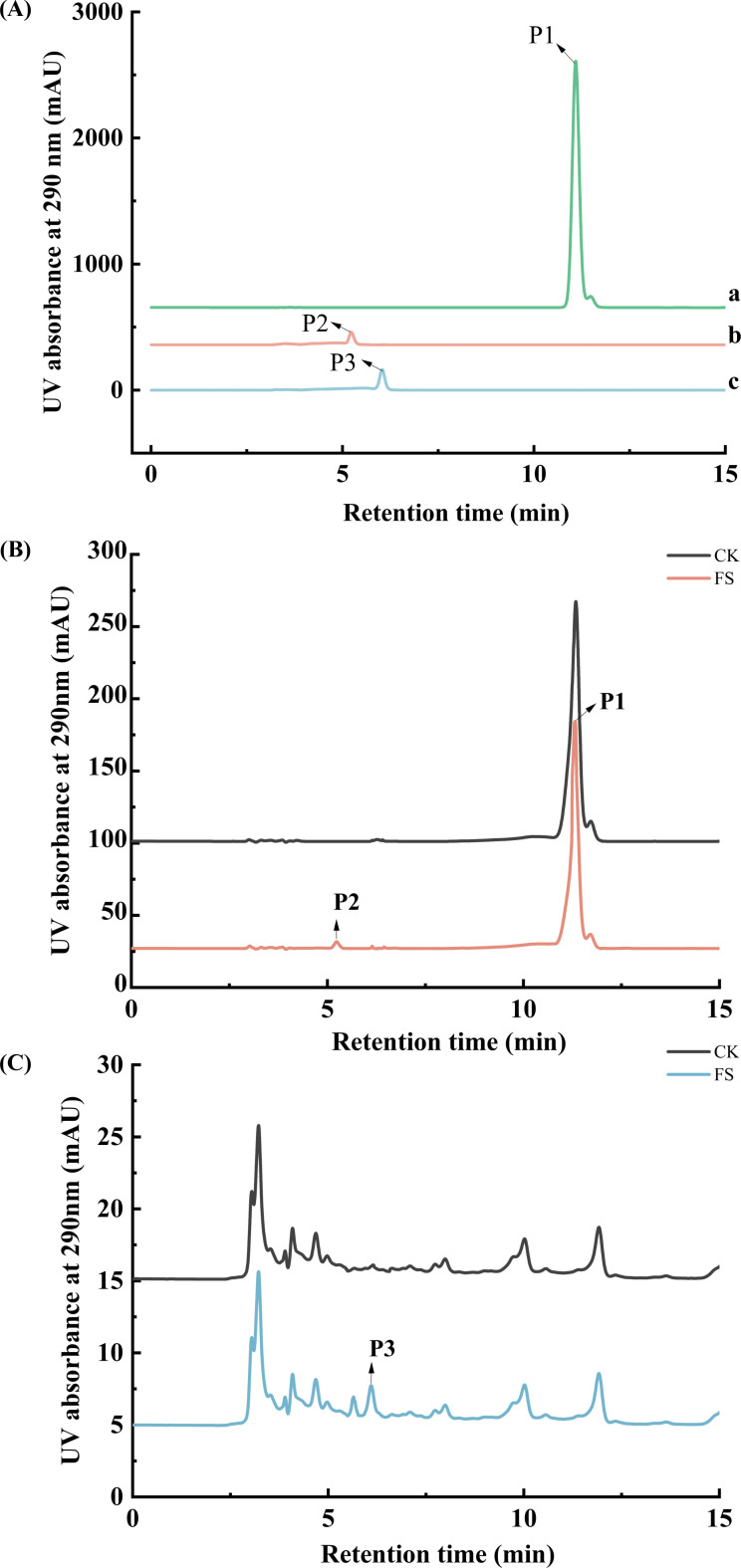 Figure 3
Figure 3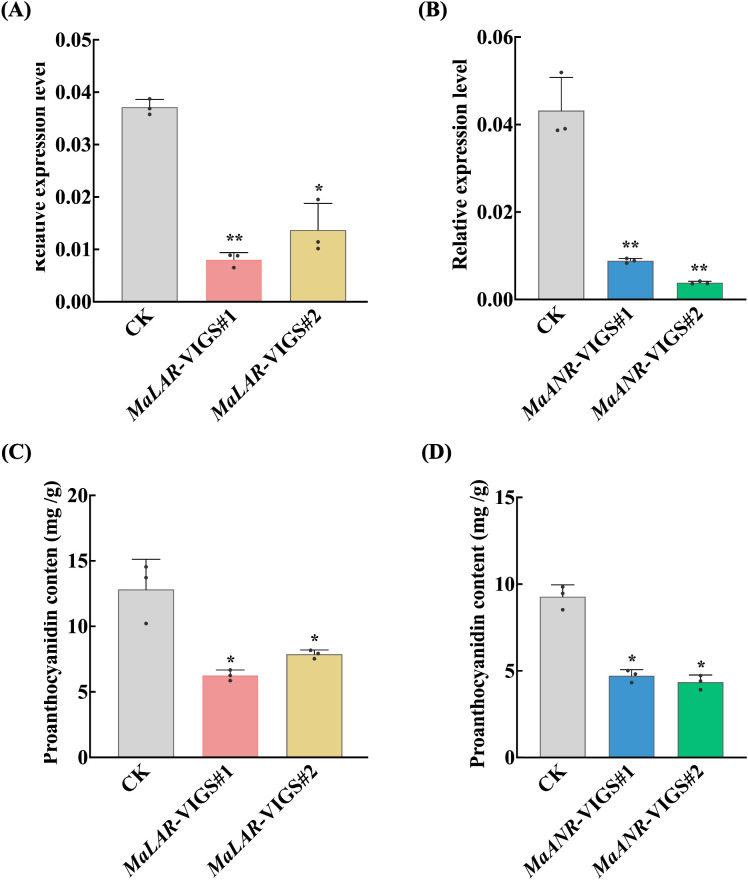 Figure 4
Figure 4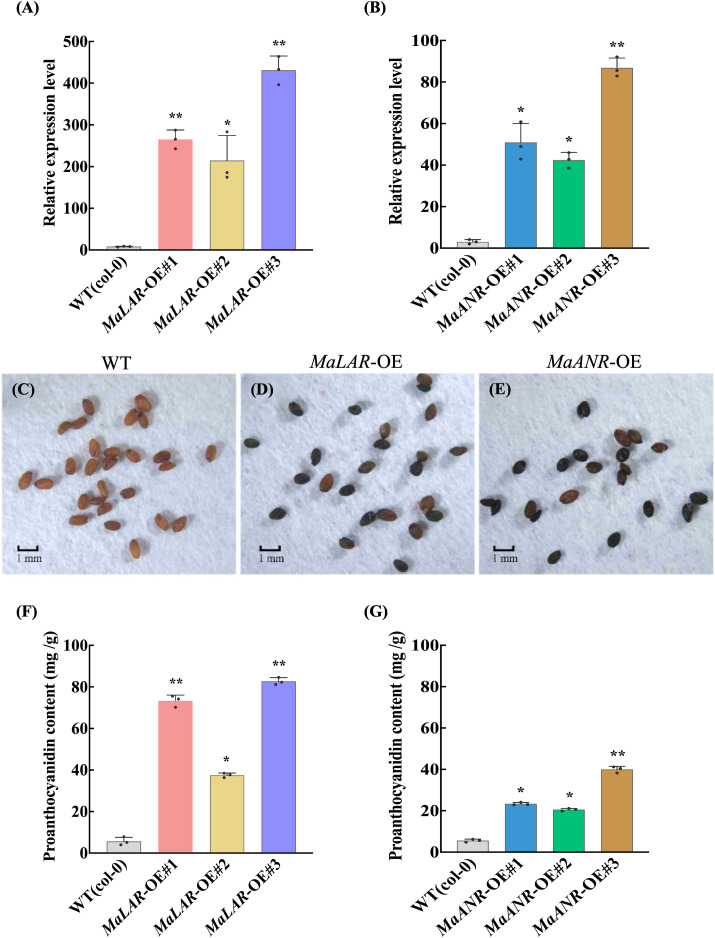 Figure 5
Figure 5| NO. | Gene ID | E-value | Score | Gene name |
|---|---|---|---|---|
| 1 | L484_022305.p01 | 3e-80 | 658 |
|
| 2 | L484_014834.p01 | 6e-90 | 679 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Phytochemicals and Antioxidant Activities · Postharvest Quality and Shelf Life Management
Introduction
1
Proanthocyanidins (PAs), also known as condensed tannins, are a class of secondary metabolites widely present in plants (Marles et al., 2003). They are polymers formed through the C-C bond condensation of flavan-3-ol monomers, primarily catechin and epicatechin (Rauf et al., 2019). In plants, PAs play vital roles in defense mechanisms and stress adaptation, offering protection against UV radiation, pathogens, and herbivores (He et al., 2008). Furthermore, PAs are critical determinants of the sensory quality of fruits, vegetables, and beverages, significantly influencing color stability, astringency, and mouthfeel (Yu et al., 2020). For human health, PAs are valued for their potent antioxidant properties and potential benefits in preventing chronic diseases (Xie et al., 2003).
Mulberry (Morus alba L.), a woody plant cultivated globally for its economic, edible, and medicinal value, accumulates a significant number of PAs in its fruits and leaves (Xin et al., 2020). The profile and concentration of these compounds are directly linked to the fruit’s sensory quality, processing suitability, and potential health benefits. Moreover, PA accumulation is hypothesized to contribute to phenotypic variations and stress resilience among different mulberry varieties (Jin et al., 2017).
The biosynthesis of PAs is a distinct branch of the flavonoid metabolic pathway, sharing precursors with anthocyanin synthesis. Two key rate-limiting enzymes, leucoanthocyanidin reductase (LAR, EC 1.17.1.3) and anthocyanidin reductase (ANR, EC 1.3.1.112), are crucial in determining the monomeric composition and abundance of PAs (Lu, 2024). LAR catalyzes the reduction of leucoanthocyanidins (e.g., leucocyanidin) to form 2,3-trans-flavan-3-ols (catechin), while ANR converts anthocyanidins (e.g., cyanidin) into 2,3-cis-flavan-3-ols (epicatechin) (He et al., 2008). The coordinated action of LAR and ANR thus governs the catechin/epicatechin ratio, which in turn dictates the degree of polymerization, structural isomerism, and the terminal/extension unit composition of the final PA polymer. These physicochemical characteristics ultimately define the fruit’s astringency, color stability, and processing attributes (Rauf et al., 2019; Zhao et al., 2023; Kennedy et al., 2001).
While the functions of LAR and ANR have been extensively studied in model plants like Arabidopsis (Marinova et al., 2007) and commercially important crops such as grape (Vitis vinifera) (Bogs et al., 2007), Persimmon (Diospyros kaki), and Litchi (Litchi chinensis Sonn.) (Zhong et al., 2024), systematic identification and functional validation of these enzymes in mulberry remain notably insufficient. Validated functional evidence still needs to be provided for identifying and characterizing LAR and ANR to reveal their roles in proanthocyanidin biosynthesis.
In the present study, we identified and cloned the candidate genes MaLAR and MaANR from M. atropurpurea var. ‘Zhongshen No.1’ (Mazs), and then conducted detailed sequence conservation and phylogenetic analyses. Spatiotemporal expression patterns in various tissues and during fruit ripening were characterized. enzymatic assay indicated MaLAR and MaANR were responsible for catechin and epicatechin formation respectively in mulberry. Further transgenic analyses revealed that they are positive regulators for PA biosynthesis.
Materials and methods
2
Plant materials and sample collection
2.1
The mulberry variety used in this study was M. atropurpurea var. ‘Zhongshen No.1’ (Mazs). The plants were cultivated and maintained at the National Mulberry Germplasm Resource Nursery (Zhenjiang, China; 32°12′ N, 119°27′ E), affiliated with the Sericultural Research Institute of the Chinese Academy of Agricultural Sciences (CAAS) and Jiangsu University of Science and Technology (JUST).
Tender leaves collected from the shoot apices were immediately frozen in liquid nitrogen and stored at -80 °C for subsequent total RNA extraction and gene cloning.
Mulberry fruit samples (cv. Mazs) were collected at four distinct developmental stages based on ripening time and phenotype: the green-ripe stage (S1, 7 days post-anthesis, DPA), the color-turning stage (S2, 14 DPA), the mature stage (S3, 21 DPA), and the fully-ripe stage (S4, 28 DPA) (Fu et al., 2025). All harvested fruit samples were immediately snap-frozen in liquid nitrogen and stored at -80°C until use.
Methods
2.2
Gene cloning and bioinformatics analysis
2.2.1
Amino acid sequences of LAR and ANR from Arabidopsis thaliana, Vitis vinifera L., and Camellia sinensis L. were used as queries for a BLASTP search against the M. notabilis genome (https://Morus.biodb.org/Morusdb) (He et al., 2013) (E-value ≤ 1 × 10^−20^, top 10% score). This search identified the candidate genes L484_022305.p01 (designated MnLAR) and L484_014834.p01 (designated MnANR).
Specific primers were designed based on the coding sequences (CDS) of the target genes, flanked with sequences compatible with seamless cloning using the pNC-Blunt vector system. Total RNA was extracted from tender leaves of the ‘Mazs’ cultivar using the EASYspin Plus Complex Plant RNA Kit (RN5301, Aidlab, Beijing, China). First-strand cDNA was synthesized with the HiScript II Q RT SuperMix for qPCR (+gDNA wiper) kit (R223-01, Vazyme, Nanjing, China), following the manufacturer’s instructions. The full-length CDS of MaLAR (969 bp) and MaANR (1014 bp) were amplified using a high-fidelity DNA polymerase. The PCR products (MaLAR and MaANR CDS; see Supplementary Figure S1) were confirmed by 1% agarose gel electrophoresis, purified using the SanPrep Column DNA Gel Extraction Kit (B518131, Sangon, Shanghai, China), and subsequently ligated into the pNC-Blunt vector using the Ready-to-Use Seamless Cloning Kit (GS2288, Bai’ao Laibo, Beijing, China). The resulting constructs were transformed into Escherichia coli TOP10 competent cells. Positive clones were selected and verified by Sanger sequencing, yielding the final constructs MaLAR and MaANR derived from the ‘Mazs’ variety.
Spatiotemporal expression analysis by qRT-PCR
2.2.2
Total RNA was extracted from tender leaves and fruit at four developmental stages (S1, S2, S3, S4) of ‘Mazs’, with three biological replicates per tissue type. RNA quality and concentration were assessed using a NanoDrop spectrophotometer (A260/A280 and A260/A230 ratios), and integrity was verified by 1% agarose gel electrophoresis. Genomic DNA was removed using gDNA Eraser, and first-strand cDNA was synthesized using the HiScript III 1st Strand cDNA Synthesis Kit (+gDNA wiper) (R312-01, Vazyme, Nanjing, China).
qPCR primers were designed using the Primer-BLAST platform (amplification efficiency 95–105%, the primer sequences for MaLAR and MaANR are provided in Supplementary Table S1). Mulberry Actin (Shukla et al., 2019) GenBank: HQ123583) was used as the internal reference gene. qRT-PCR reactions were performed using SYBR^®^ Premix Ex Taq™ II (Q341-02, Vazyme, Nanjing, China) in a 20 μL system (10 μL Premix, 0.4 μM of each primer, 2 μL cDNA) on an ABI 7500 Real-Time PCR System. The thermal cycling conditions were: 95°C for 30 s; 40 cycles of 95°C for 5 s and 60°C for 34 s (fluorescence acquisition). A melt curve analysis (95°C for 15 s, 60°C for 1 min, 95°C for 15 s) was conducted to ensure amplification specificity. Three technical replicates were performed for each sample. Relative expression levels were calculated using the 2^−ΔΔCt^ method and are presented as the mean ± SD.
Recombinant protein expression, purification, and HPLC-based enzyme assay
2.2.3
The sequence-verified CDS of MaLAR and MaANR were cloned into the pET-28a (+) vector (N-terminal His_6_ tag) and transformed into E. coli BL21 (DE3) cells. Cultures were grown at 37°C in LB medium (50 μg/mL kanamycin) to an OD_600_ of 0.6–0.8. Protein expression was induced with 0.5 mM IPTG and incubated overnight at 16°C.
Cells were harvested, resuspended in lysis buffer, and disrupted by sonication. The soluble fraction was purified by Ni^2+^-NTA affinity chromatography, followed by dialysis to remove free imidazole. Protein purity and molecular weight were confirmed by SDS–PAGE (Supplementary Figure S2).
The in vitro enzymatic assays for MaLAR and MaANR were performed as previously described, with minor modifications (Gagné et al., 2009). For the MaLAR assay, taxifolin (dihydroquercetin; V91397, MedMol, Shanghai, China) was used as the substrate. The reaction mixture (300 µL total volume) contained 5 µL taxifolin (2000 ng/µL), 152 µL 0.1 M Tris-HCl buffer (pH 7.5; T301503, Aladdin, Shanghai, China), 2 µL NADPH (N276326, Aladdin, Shanghai, China), 70 µL purified MaDFR1, 70 µL purified MaLAR, and 1 µL dithiothreitol (DTT; D755747, Aladdin, Shanghai, China). For the MaANR assay, cyanidin chloride (B20784, MedMol, Shanghai, China) was used as the substrate. The reaction mixture (500 µL total volume) contained 100 µL cyanidin chloride (2000 ng/µL), 195 µL 0.1 M Tris-HCl buffer (pH 6.0; T301503, Aladdin, Shanghai, China), 5 µL NADPH (N276326, Aladdin, Shanghai, China), and 200 µL purified MaANR. Control reactions were performed using heat-inactivated enzymes. Reactions were incubated in a constant-temperature water bath at 35 °C for 45 min. Reaction products were extracted with ethyl acetate (E116141, Aladdin, Shanghai, China) by vortexing (the extraction step for ANR assays was repeated once). The combined organic phases were dried under nitrogen, redissolved in HPLC-grade methanol (M433267, Aladdin, Shanghai, China), centrifuged, and filtered prior to HPLC analysis.
Enzymatic products were detected at 290 nm. Substrate consumption and product formation were quantified by comparing retention times with authentic standards using the external standard method.
Virus-Induced Gene Silencing and knock-down analysis
2.2.4
Gene-specific 300 bp fragments of MaLAR and MaANR were predicted using the online SGN VIGS Tool (https://vigs.solgenomics.net/), and the primers used for gene silencing are listed in Supplementary Table S1. Fragments were amplified with primers containing NC-linker arms, verified by electrophoresis, and ligated into the linearized pNC-TRV2-GFP vector via seamless cloning to construct pNC-TRV2-GFP-MaLAR and pNC-TRV2-GFP-MaANR.
The recombinant TRV2 plasmids and the TRV1 vector were co-transformed (1:1 molar ratio) into Agrobacterium tumefaciens GV3101. Cultures were grown to OD_600_ ≈ 1.0, collected, and resuspended in MMA infiltration buffer (10 mM MgCl_2_, 10 mM MES, pH 5.6, 150 μM acetosyringone), and incubated at room temperature for 3 h (Mo et al., 2024).
Sterile seedlings of M alba var. fengchi (4–6 true leaves) were infiltrated on the abaxial leaf side via syringe. Plants were grown at 24°C (16 h light/8 h dark). GFP fluorescence was monitored 2–3-week post-infiltration under UV light to select positive plants. Total RNA was extracted from GFP-positive leaves, and transcript levels of MaLAR and MaANR were quantified by qRT-PCR (Actin internal control) using the 2^−ΔΔCt^ method to evaluate gene silencing efficiency. (Supplementary Figure S3).
Generation of overexpression Arabidopsis and phenotypic analysis
2.2.5
The full-length CDSs of MaLAR (969bp) and MaANR (1,014bp) were cloned into the pNC-Cam13FN vector under the control of the CaMV 35S promoter. Recombinant plasmids were introduced into A. tumefaciens GV3101 via triparental mating. A. thaliana (Col-0) plants were transformed using the floral dip method (Clough and Bent, 1998).
Harvested T_0_ seeds were screened on 1/2 MS medium containing 50 μg/mL kanamycin. Positive transformants were grown and self-pollinated to obtain T_2_ generation seeds.
Transgene integration in T_2_ plants was confirmed by genomic DNA PCR using gene-specific primers identical to those employed for qRT-PCR analysis. Transgene expression levels were quantified by qRT-PCR using cDNA as the template, with AtACT7 as the internal control. Relative expression was calculated using the 2^−ΔΔCt^ method. All primer sequences used for genomic DNA PCR and qRT-PCR analyses are listed in Supplementary Table S1. PA distribution in T_2_ seed coats was visualized by staining seeds with 0.1% DMACA solution for 10 min and observing them under an optical microscope. Total PA content in seedlings was quantified colorimetrically using the DMACA method (640 nm absorbance) (Li et al., 1996), with epicatechin as the standard.
Results
3
Identification and characterization of mulberry LAR and ANR
3.1
Using the protein sequences of LAR and ANR from A. thaliana, V. vinifera, and C. sinensis as queries, BLASTP searches were performed against the M. notabilis genome. Two candidate genes were identified: L484_022305.p01 and L484_014834.p01, which were subsequently designated as MnLAR and MnANR, respectively (Table 1). MaLAR and MaANR were then successfully cloned from M. atropurpurea var. Zhongshen No.1 (Mazs). The CDS of MaLAR was 969 bp in length, encoding a protein of 322 amino acids, whereas the CDS of MaANR was 1,014 bp, encoding a 337-amino-acid protein. The sequences of MaANR and MaLAR analyzed in this study have been deposited in the GenBase database (NGDC; https://ngdc.cncb.ac.cn/genbase/) under the accession numbers C_AA132743.1 and C_AA132744.1, respectively.
Phylogenetic analysis showed that MaANR and MaLAR clustered with previously characterized ANR and LAR proteins from other plant species, respectively (Figure 1A). Multiple sequence alignment and conserved domain analysis revealed that MaANR contains the conserved NADPH-binding site, and its predicted substrate-binding residues are highly consistent with those of reported ANRs. MaLAR harbors the three conserved motifs RFLP, ICCN, and THD, which are characteristic of this reductase family (Wang et al., 2018; Liao et al., 2015; Ferraro et al., 2014) (Figures 1B, C). Homology-based 3D structural modeling indicated clear structural differences between MaANR and MaLAR, consistent with their distinct catalytic functions (Figure 1D).
Cloning and identification of MaLAR and MaANR genes in mulberry. (A) Phylogenetic tree of MaLAR and MaANR proteins from mulberry aligned with homologous amino acid sequences from other species. The MaLAR and MaANR are indicated in bold. (B) Multiple sequence alignment of LAR from mulberry and other species. The three conserved domains are indicated by square boxes. (C) Multiple sequence alignment of MaANR from mulberry and other species. The arrow, square, and horizontal line indicate the predicted active site, the NADPH coenzyme-binding site, and the substrate-binding site, respectively. (D) Predicted tertiary structure of the MaLAR protein from mulberry. (E) Predicted tertiary structure of the MaANR protein from mulberry.
Spatiotemporal expression patterns of MaLAR and MaANR
3.2
The spatiotemporal expression patterns of MaLAR and MaANR were assessed by quantitative real-time PCR (qRT-PCR) using cDNA synthesized from Ma leaves and fruits collected at four developmental stages (S1, S2, S3, S4) (Figure 2). Both genes displayed pronounced tissue specificity and dynamic transcriptional changes during fruit maturation.
Analysis of MaLAR and MaANR gene expression levels at different developmental stages. (A)MaLAR organ expression profile. (B)MaANR organ expression profile. S1 Green-ripe stage; S2 Color-turning stage; S3 Mature stage; S4 Fully-ripe stage; Leaf. Values represent the mean ± SD of three biological replicates (n = 3). Different lowercase letters above the bars indicate statistically significant differences among samples, as determined by one-way ANOVA followed by Tukey’s multiple comparison test (P < 0.05).
The expression of MaLAR increased progressively throughout fruit development, reaching its maximum at the fully ripe stage (S4) (Figure 2A). In contrast, MaANR exhibited transient induction pattern, with transcript levels peaking at the color-turning stage (S2) before declining at later stages (Figure 2B).
These distinct expression trajectories indicate that MaLAR and MaANR may fulfill complementary yet non-redundant functions in regulating proanthocyanidin biosynthesis during mulberry fruit development.
In vitro enzymatic assay of MaLAR and MaANR
3.3
To further validate the roles of MaLAR and MaANR in the PA biosynthetic pathway, recombinant proteins were expressed and purified, and in vitro enzymatic assay systems were established. The purity and molecular weights of the purified proteins were confirmed by SDS–PAGE (Supplementary Figure S2), and these proteins were then used for enzymatic activity assays. The standard products, catechin and epicatechin serving as references for product identification were detected using HPLC-VWD at 290 nm with retention time of 5.226 min and 6.035 min respectively, (Figure 3A). Enzymatic assay showed that MaLAR can catalyze leucoanthocyanidin produced by DFR using dihydroquercetin as substrates (Figure 3B). Furthermore, the MaANR activity assay revealed a characteristic epicatechin peak at 6.072 min. This product was also not detected in the heat-inactivated control (Figure 3C), confirming its ability to reduce anthocyanidin to epicatechin.
Chromatogram of MaLAR and MaANR enzymatic assay. (A) Chromatograms of dihydroquercetin, catechin and epicatechin standards; A-a, dihydroquercetin; A-b, Catechins; A-c, epicatechin. (B) Chromatogram of MaLAR enzyme catalytic activity assay. FS: Experimental group, Reaction system + MaLAR; CK: Control group, Reaction system + MaLAR (heat-inactivated). (C) Chromatogram of MaANR enzyme catalytic activity assay. FS: Reaction system + MaANR; CK: Reaction system + MaANR (heat-inactivated).
Collectively, the in vitro enzymatic evidence unequivocally confirms that MaLAR and MaANR are responsible for the formation of catechin and epicatechin, respectively, further establishing their critical and rate-limiting roles in the mulberry PA biosynthetic pathway (Sunkar et al., 2003; Tucker, 2003; Pang et al., 2013).
Knock-down of either MaLAR or MaANR using VIGS resulted in reduction of PAs
3.4
To elucidate the in vivo functional roles of MaLAR and MaANR in the mulberry PA biosynthetic pathway, virus-induced gene silencing (VIGS) constructs (pNC-TRV2-GFP-MaLAR and pNC-TRV2-GFP-MaANR) were generated and introduced into M. alba var. fengchi.
qRT-PCR analysis verified effective suppression of the target genes in the silenced lines. MaLAR transcript levels were reduced by 78.28% and 63.06% in lines #1 and #2, respectively (Figure 4A). Similarly, MaANR expression decreased by 79.48% and 91.11% in the corresponding #1 and #2 lines (Figure 4B).
Changes in gene expression levels and proanthocyanidin content after VIGS treatment. (A) Expression levels of MaLAR in mulberry leaves after VIGS treatment. (B) Expression levels of MaANR in mulberry leaves after VIGS treatment. (C) Proanthocyanidin content in leaves of MaLAR-knockdown lines. (D) Proanthocyanidin content in leaves of MaANR- knockdown lines; Statistical significance was assessed using Student’s t-test. Significance levels were indicated as follows: 0.01 < P ≤ 0.05 (), and 0.001 < P ≤ 0.01 (**).*
Consistent with the extent of gene silencing, total PA levels also showed pronounced declines. Relative to the control plants, silencing MaLAR reduced total PAs content by 51.16% and 38.57% in lines #1 and #2, respectively (Figure 4C). Silencing MaANR resulted in decreases of 49.15% and 53.11% in lines #1 and #2, respectively (Figure 4D). Given that catechin and epicatechin serve as essential monomeric precursors for PA polymerization, impaired synthesis of these intermediates directly contributed to the diminished PA accumulation observed in the silenced plants.
Collectively, these VIGS assays provide strong evidence for MaLAR and MaANR as key in vivo contributors to PA biosynthesis in mulberry, and downregulation of either gene leads to substantial reduction in total PA content.
Heterologous overexpression of MaLAR and MaANR in A. thaliana resulted in accumulation of PAs
3.5
To further substantiate the functional roles of MaLAR and MaANR in PA biosynthesis, each genes was heterologously overexpressed in A. thaliana independently. The resulting transgenic lines were systematically evaluated for phenotypic characteristics, transcriptional changes, and metabolite accumulation.
qRT-PCR analysis confirmed substantial upregulation of the target genes, with MaLAR transcript levels elevated by 25.54- to 52.37-fold and MaANR levels increased by 12.93- to 27.53-fold relative to the wild type (WT) (Figures 5A, B). DMACA staining of T_2_ seeds revealed markedly intensified blue coloration in the transgenic lines compared with WT seeds (Figures 5C–E), indicating enhanced PA deposition in the seed coat.
OE-Arabidopsis seed staining and PAs quantification. (A) Expression level of MaLAR in transgenic Arabidopsis(B) Expression level of MaANR in transgenic Arabidopsis(C) Wild-type Arabidopsis thaliana seeds (D) Overexpression of MaLAR A. thaliana seeds (E) Overexpression of MaANR A. thaliana seeds (F) Proanthocyanidin content determination in MaLAR-overexpressing Arabidopsis(G) Proanthocyanidin content in MaANR-overexpressing Arabidopsis. Statistical significance was assessed using Student’s t-test. Significance levels were indicated as follows: 0.01 < P ≤ 0.05 (), and 0.001 < P ≤ 0.01 (**).*
Consistent with the staining results, quantitative measurements showed dramatic increases in total PA content. In the MaLAR overexpression lines, PA levels were 5.61- to 13.58-fold higher than in WT, whereas MaANR overexpression lines exhibited 2.69- to 6.18-fold increases (Figures 5F, G). This substantial accumulation is attributable to the heightened synthesis of catechin and epicatechin—key monomeric precursors required for PA polymerization—thereby driving elevated PA deposition and intensified staining signals.
In conclusion, the heterologous overexpression results in Arabidopsis demonstrate that increased transcription of MaLAR and MaANR markedly enhances the biosynthetic flux toward catechin and epicatechin, ultimately promoting PA accumulation in the seed coat. These findings, in concordance with the VIGS silencing assays and the in vitro enzymatic characterization, collectively confirm the essential positive regulatory roles of MaLAR and MaANR in the mulberry PA biosynthetic pathway.
Discussion
4
Core functions of LAR and ANR in proanthocyanidin biosynthesis
4.1
LAR and ANR are key terminal enzymes in the PA biosynthetic pathway, catalyzing the formation of catechin and epicatechin, respectively (Jun et al., 2021). By controlling both the availability and stereochemical configuration of flavan-3-ol monomers, these enzymes play central roles in shaping PA composition and structural diversity. Although the core PA biosynthetic pathway is broadly conserved among higher plants, increasing evidence has revealed substantial enzymatic complexity and functional diversification of LAR and ANR across species (Yu et al., 2023; Lin et al., 2024).
Notably, in several plant species, ANR exhibits expanded catalytic activities, generating not only epicatechin starter units but also putative intermediates such as 2,3-trans-flavan-3,4-diols or their corresponding thiol derivatives, which can serve as extension units during PA polymerization (Jun et al., 2021). Such substrate versatility highlights the species-specific catalytic preferences of ANR and underscores its influence on PA polymer length and composition. Similarly, in species such as grapevine and Medicago truncatula, LAR has been reported to catalyze not only the conversion of leucoanthocyanidins to catechin but also the reverse transformation of 4β-(S-cysteinyl)-epicatechin to epicatechin. This capacity modulates the balance between starter and extension units, ultimately shaping PA chain length (Xie et al., 2004; Liang et al., 2023).
These functional characteristics contrast sharply with the metabolic landscape of Arabidopsis thaliana, which lacks an endogenous LAR gene and accumulates proanthocyanidins composed almost exclusively of epicatechin units derived from the ANR pathway (Gagné et al., 2009; Chen et al., 2022; Bogs et al., 2005). In the present study, however, heterologous expression of MaLAR or MaANR in Arabidopsis resulted in a significant increase in total PA content, as quantified by the DMACA colorimetric assay. Although this method does not discriminate between catechin- and epicatechin-derived units, it reliably reflects overall PA accumulation. The observed increase in PA levels suggests that the introduction of functional MaLAR activity—which is otherwise absent in Arabidopsis—may enhance the overall flux of flavan-3-ols toward PA biosynthesis. Crucially, such enhancement does not necessarily require detectable accumulation of free catechin monomers, as newly formed catechin units may be rapidly incorporated into PA polymers or indirectly influence PA production by redirecting metabolic intermediates away from competing flavonoid branches.
Previous research has reported that co-expression of multiple flavonoid biosynthetic genes can enable ectopic catechin biosynthesis, as demonstrated by the simultaneous expression of CsANS, CsLAR, and CsANR from the tea plant in tobacco (Yang et al., 2025). In addition, the ban and ldox mutants of Arabidopsis thaliana are well-characterized lines impaired in proanthocyanidin biosynthesis due to defects in key enzymatic steps of the flavonoid pathway, making them suitable genetic backgrounds for complementation-based functional validation (Abrahams et al., 2002; Rafique et al., 2016). Accordingly, genetic complementation using Arabidopsis ban or ldox mutants, together with co-expression of MaLAR and MaANR, could be explored in future work to further substantiate their roles in PA biosynthesis. Consistent with this interpretation, previous studies in species possessing functional LAR, including cacao and grapevine, have demonstrated that LAR activity contributes to PA accumulation primarily through modulation of flavonoid metabolic flux rather than through substantial accumulation of free catechin monomers. Collectively, these findings support the notion that, although the catalytic functions of LAR and ANR are central to PA biosynthesis across diverse plant lineages, species-specific metabolic contexts ultimately determine PA composition and structural complexity (Chen et al., 2022; Bogs et al., 2005; He et al., 2008; Lin et al., 2024; Liu et al., 2013; Yu et al., 2023).
Evolutionary patterns and regulatory networks of LAR and ANR genes
4.2
At the genomic level, LAR and ANR genes are broadly distributed across angiosperms; however, their copy number and structural organization frequently exhibit species-specific expansion and diversification (Jiao et al., 2023). In several economically important crops, such as grape and tea, multiple functional homologs of LAR and ANR have been identified, and their expression levels show strong correlations with the accumulation of catechin and epicatechin (Lin et al., 2024).
The spatiotemporal transcriptional regulation of LAR and ANR is primarily orchestrated by the MYB–bHLH–WD40 (MBW) regulatory complex (Shan et al., 2025). In the Arabidopsis seed coat, the TT2–TT8–TTG1 module constitutes the core regulatory unit that activates ANR expression, and homologous regulatory components have been identified and shown to be functionally conserved in species such as grape, strawberry, and tobacco (Jiang et al., 2023).
Beyond the canonical MBW complex, hormonal cues (e.g., auxin and ethylene) and environmental stimuli (e.g., UV-C irradiation) can further modulate LAR and ANR expression (Bulanov et al., 2025). For example, in apple, ethylene-responsive transcription factors indirectly enhance MdLAR and MdANR expression through upstream regulation of MYB activators (Wang et al., 2022b).
Collectively, these findings indicate that LAR and ANR gene regulation operates through multilayered and species-specific networks, reflecting divergent evolutionary strategies that enable plants to meet distinct biochemical and physiological demands for PA biosynthesis under developmental and environmental constraints (Jiang et al., 2023).
Application potential and future directions of PA metabolic engineering
4.3
With the growing recognition of the roles of PAs in enhancing plant stress resistance and nutritional quality (Yu et al., 2023), metabolic engineering targeting LAR and ANR has emerged as an effective strategy for crop trait improvement (Jiang et al., 2023). The heterologous expression of MYB, bHLH, and LAR/ANR genes has enabled the establishment of PA-accumulating systems in plants that typically do not synthesize PAs (Escaray et al., 2023). For instance, robust PA production has been achieved in tobacco leaves and white clover, resulting in enhanced nitrogen use efficiency and reduced methane emissions in ruminant livestock (Lu et al., 2023). In addition, upregulation of LAR/ANR in crops such as cotton and mulberry has been shown to improve disease resistance and promote fiber development (Wang et al., 2022a).
Despite these advances, excessive PA accumulation can negatively affect plant growth. Therefore, recent studies have increasingly focused on tissue-specific or inducible expression systems to mitigate potential developmental penalties associated with constitutive overaccumulation (Khan et al., 2023). Looking forward, comprehensive elucidation of the LAR/ANR regulatory network, together with emerging technologies such as gene editing and precision transcriptional regulation, will further facilitate the development of optimized PA metabolic engineering strategies aimed at improving crop quality, enhancing stress resilience, and increasing the nutritional value of forage crops (Das et al., 2024; Wang et al., 2025).
Conclusion
5
This study leveraged M. notabilis genomic resources to comprehensively elucidate the molecular characteristics and functional roles of the key reductases MaLAR and MaANR in the mulberry proanthocyanidin (PA) biosynthetic pathway.
Our results demonstrate that both genes are highly conserved in mulberry and exhibit distinct spatiotemporal expression dynamics during fruit development: MaLAR expression progressively increased throughout fruit maturation, whereas MaANR expression reached its maximum at the color-turning stage. In vitro enzymatic assays unequivocally confirmed that MaLAR and MaANR catalyze the formation of catechin and epicatechin, respectively. In vivo functional validation further substantiated their roles, as VIGS-mediated silencing of either gene significantly reduced total PA content in mulberry leaves, while heterologous overexpression in Arabidopsis markedly enhanced PA accumulation in the seed coat.
In conclusion, the converging evidence from in vitro enzymatic characterization, in vivo gene silencing, and heterologous overexpression firmly establishes MaLAR and MaANR as essential positive regulators of PA biosynthesis in mulberry. These findings not only deepen our understanding of the molecular mechanisms governing secondary metabolism in mulberry but also provide valuable molecular targets and theoretical guidance for modulating PA content and improving fruit quality through molecular breeding and metabolic engineering.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abrahams S. Tanner G. J. Larkin P. J. Ashton A. R. (2002). Identification and biochemical characterization of mutants in the proanthocyanidin pathway in Arabidopsis. Plant Physiol. 130, 561–576. doi: 10.1104/pp.006189, PMID: 12376625 PMC 166587 · doi ↗ · pubmed ↗
- 2Bogs J. Downey M. O. Harvey J. S. Ashton A. R. Tanner G. J. Robinson S. P. (2005). Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 139, 652–663. doi: 10.1104/pp.105.064238, PMID: 16169968 PMC 1255985 · doi ↗ · pubmed ↗
- 3Bogs J. JafféF. W. Takos A. M. Walker A. R. Robinson S. P. (2007). The grapevine transcription factor Vv MYBPA 1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 143, 1347–1361. doi: 10.1104/pp.106.093203, PMID: 17208963 PMC 1820911 · doi ↗ · pubmed ↗
- 4Bulanov A. N. Andreeva E. A. Tsvetkova N. V. Zykin P. A. (2025). Regulation of flavonoid biosynthesis by the MYB-b HLH-WDR (MBW) complex in plants and its specific features in cereals. Int. J. Mol. Sci. 26, 734. doi: 10.3390/ijms 26020734, PMID: 39859449 PMC 11765516 · doi ↗ · pubmed ↗
- 5Chen Q. Liang X. Wu C. Liu Y. Liu X. Zhao H. . (2022). Overexpression of leucoanthocyanidin reductase or anthocyanidin reductase elevates tannins content and confers cassava resistance to two-spotted spider mite. Front. Plant Sci. 13, 994866. doi: 10.3389/fpls.2022.994866, PMID: 36061805 PMC 9433999 · doi ↗ · pubmed ↗
- 6Clough S. J. Bent A. F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana: Floral dip transformation of Arabidopsis. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x, PMID: 10069079 · doi ↗ · pubmed ↗
- 7Das S. Kwon M. Kim J.-Y. (2024). Enhancement of specialized metabolites using CRISPR/Cas gene editing technology in medicinal plants. Front. Plant Sci. 15, 1279738. doi: 10.3389/fpls.2024.1279738, PMID: 38450402 PMC 10915232 · doi ↗ · pubmed ↗
- 8Escaray F. J. Valeri M. C. Damiani F. Ruiz O. A. Carrasco P. Paolocci F. (2023). Multiple b HLH/MYB-based protein complexes regulate proanthocyanidin biosynthesis in the herbage of Lotus spp. Planta 259, 10., PMID: 38041705 10.1007/s 00425-023-04281-2PMC 10693531 · doi ↗ · pubmed ↗