Evolution of sensory systems underlies the emergence of predatory feeding behaviors in nematodes
Marianne Roca, Güniz Göze Eren, Leonard Böger, Olena Didenko, Wen-Sui Lo, Monika Scholz, James W. Lightfoot

TL;DR
This paper shows how the nematode Pristionchus pacificus evolved to use both touch and chemical senses to hunt prey, revealing how new behaviors can emerge through changes in sensory systems and neural circuits.
Contribution
The study identifies a mechanosensory module linked to predatory behavior in P. pacificus, revealing a novel circuit-level mechanism for behavioral evolution.
Findings
P. pacificus uses mechanosensory and chemosensory systems together to detect prey.
Ppa-mec-6 is a key mechanosensory gene expressed in IL2 neurons crucial for predation.
Disrupting mechanosensation and chemosensation together has a synergistic effect on prey detection.
Abstract
Understanding how new behaviors evolve is a major challenge in biology. In this study, we show the nematode Pristionchus pacificus has evolved to use both mechanical and chemical sensory modalities to detect and attack prey. By screening sensory genes and analyzing behavior with machine learning, we identified a mechanosensory module associated with the evolution of predation. Furthermore, these pathways are integrated within a specific set of neurons required for prey detection, revealing a circuit-level mechanism for predatory behavior that is distinct from the function of homologous neurons in Caenorhabditis elegans. These findings reveal how evolution can modify sensory modalities and reconfigure neural pathways to generate new behaviors, shedding light on the mechanisms that drive behavioral innovation. Understanding how animal behavior evolves remains a major challenge, with few…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
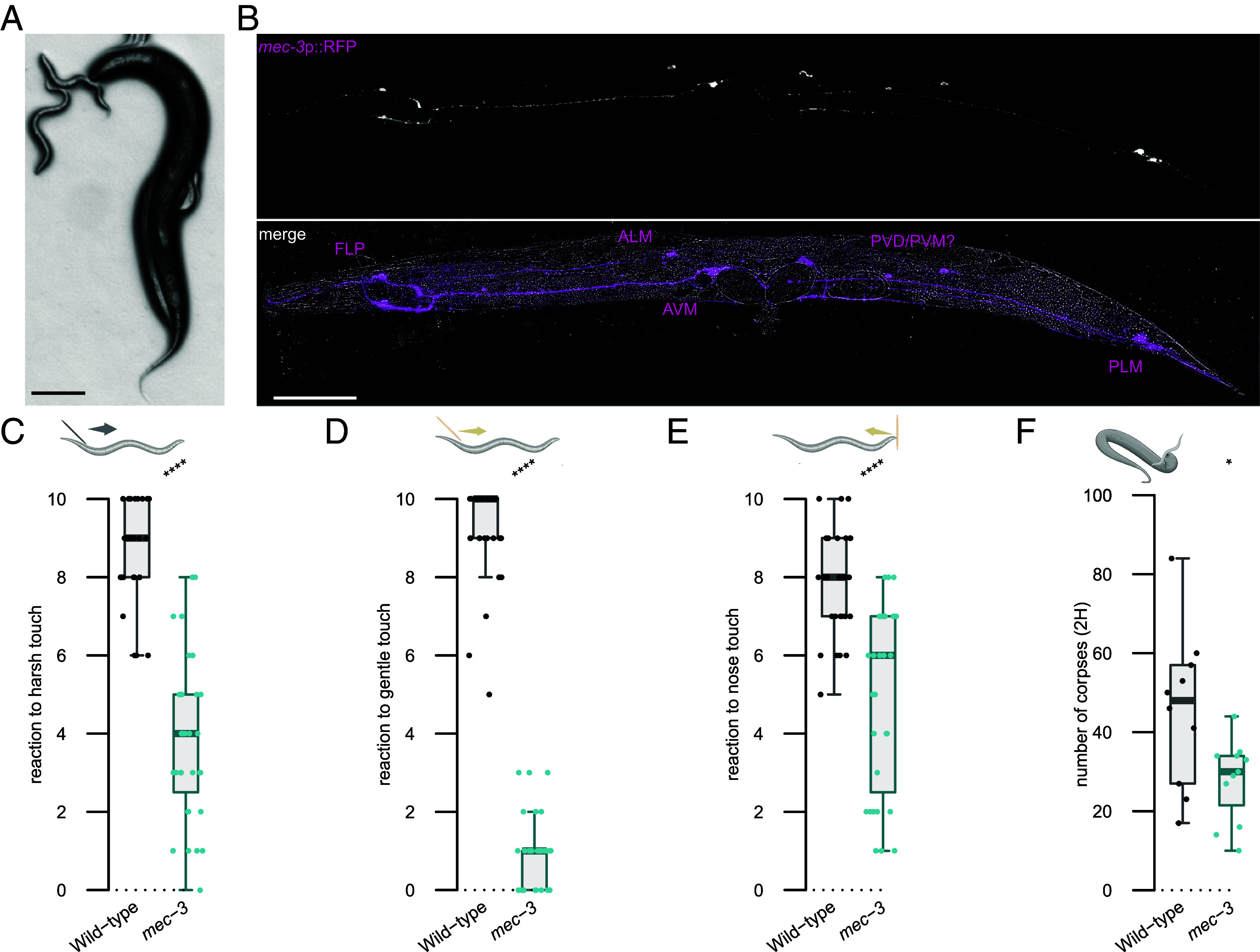 Fig. 1
Fig. 1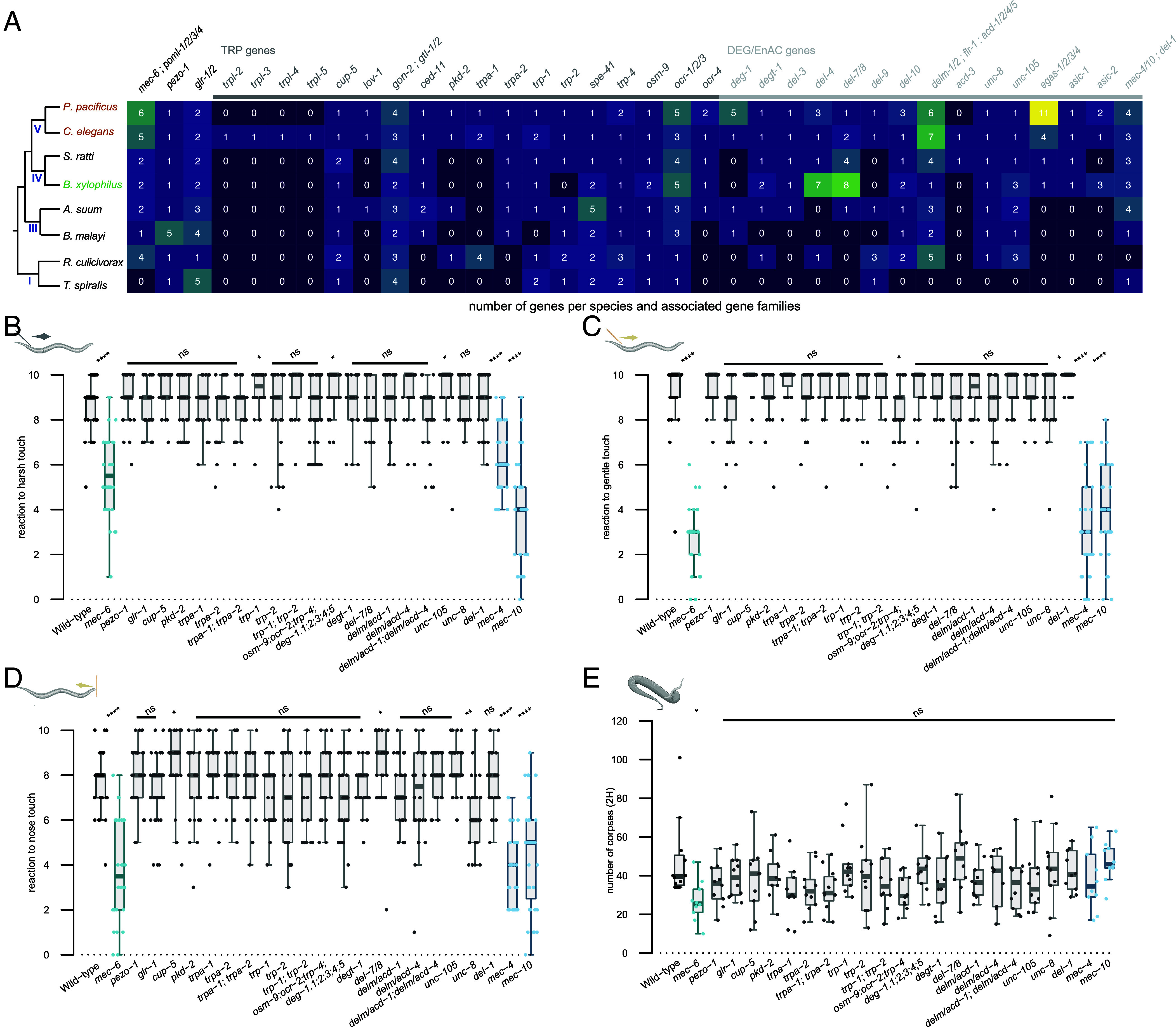 Fig. 2
Fig. 2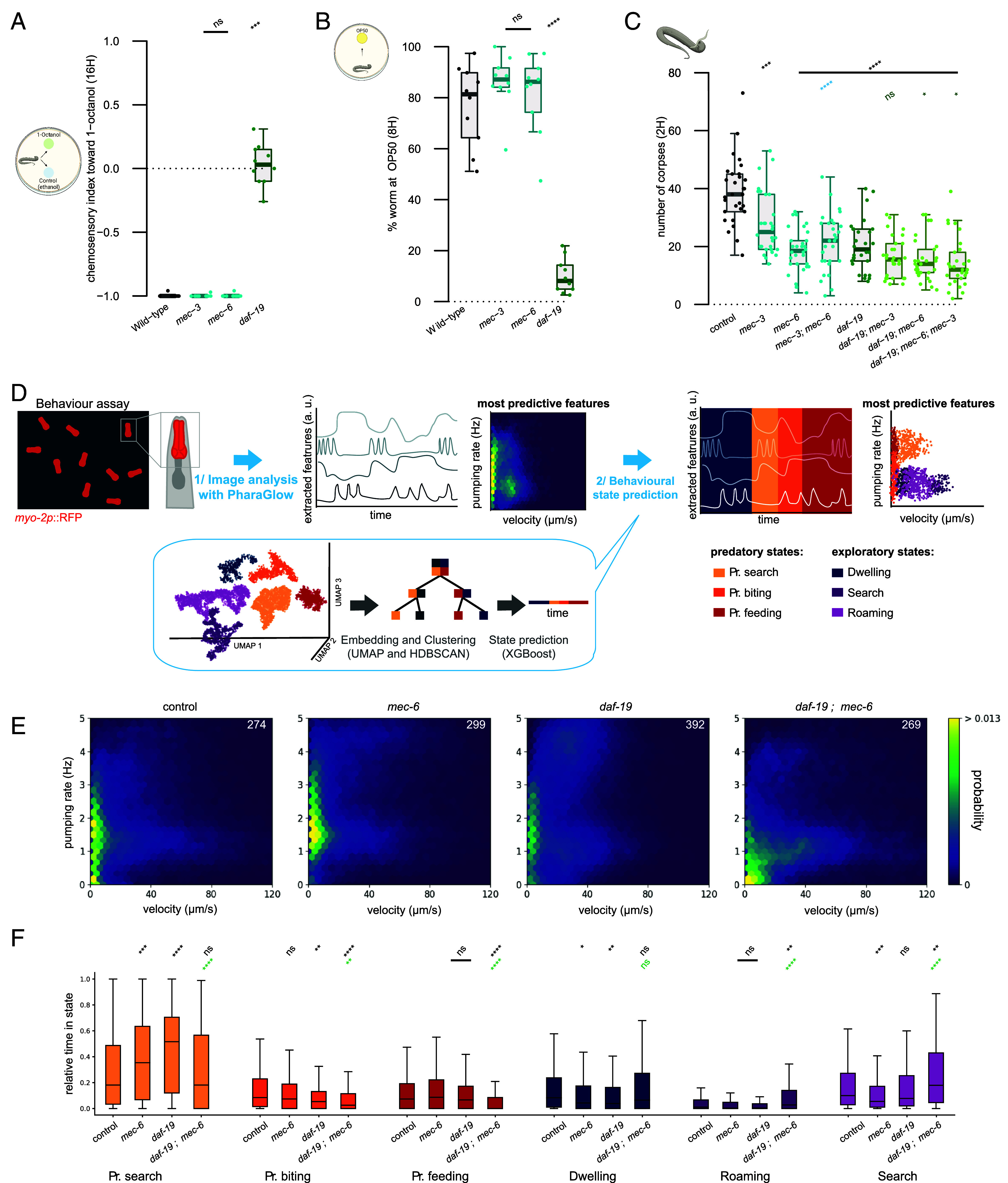 Fig. 3
Fig. 3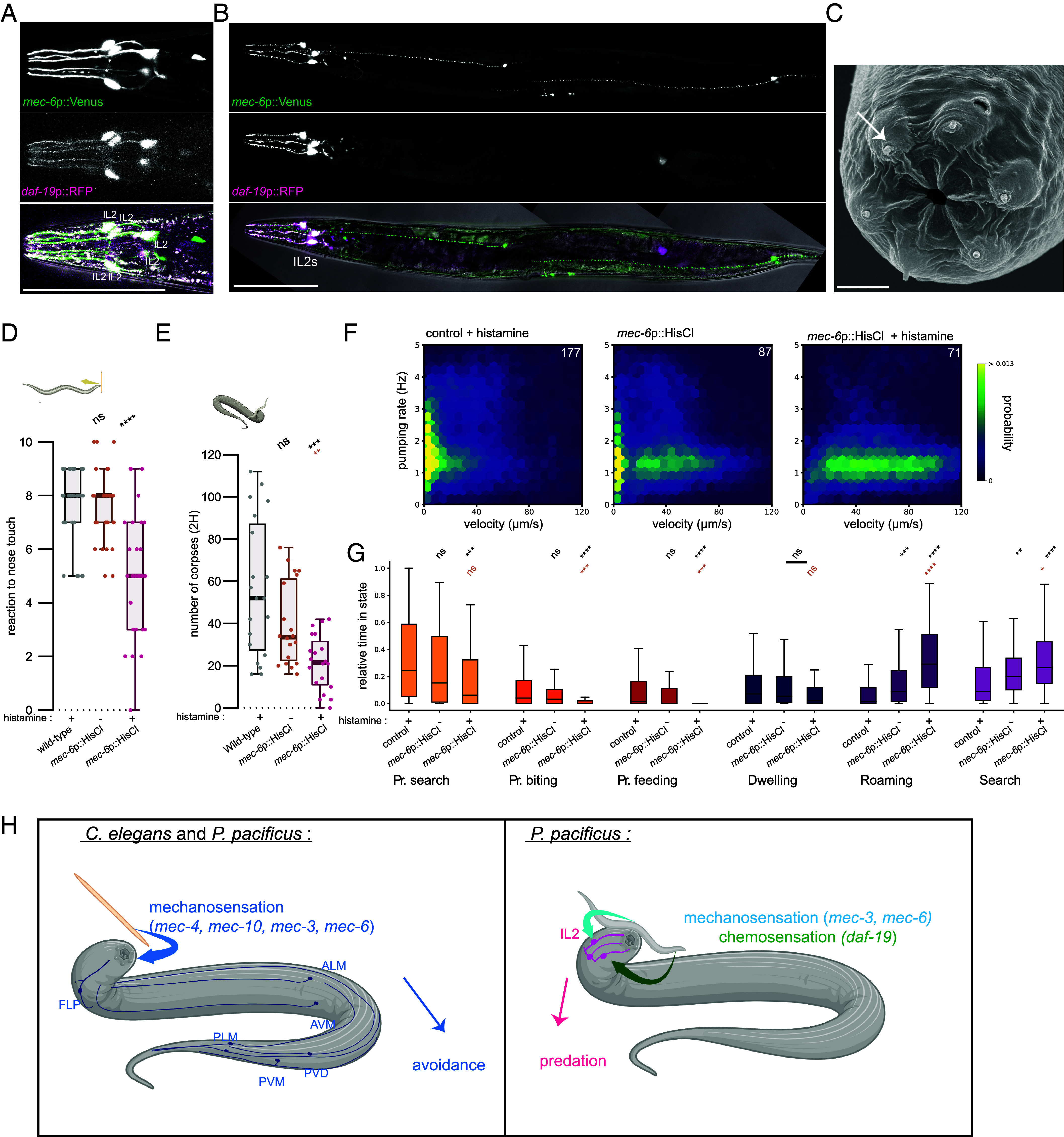 Fig. 4
Fig. 4- —Max Planck Society
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Nematode management and characterization studies · Parasite Biology and Host Interactions
Sensory systems represent the primary mechanism through which an organism acquires environmental information. These signals can be used for finding food, avoiding predators, detecting mates, and navigating through complex surroundings. Detecting these environmental cues is dependent on specialized sensory systems which are often finely tuned to specific stimuli. Crucially, these systems are not static and their modification can profoundly affect the evolutionary trajectory of a species (1). Mechanisms of sensory system evolution include modifications to receptors, such as changes to receptor selectivity or sensitivity, as well as associated gene losses or gains (1). Examples of such events have been observed in several invertebrate species with changes in receptor function influencing feeding ecology in fruit flies (2), mosquitoes (3), and cockroaches (4). Similarly, in vertebrates, changes to sweet receptors have modified the feeding behaviors of songbirds and bats (5, 6) while in deep sea fish, a greatly expanded number of opsins has evolved to maximize their visual sensitivity in minimal light environments (7). Thus, sensory system evolution shapes the behavioral repertoire of an organism and facilitates its adaptation to its ecological niche.
In nematodes, a remarkable range of behaviors has evolved in accordance with the diverse ecologies found across the phyla. In particular, with their exceptional array of genetic and molecular tools, (8?–10) alongside their distinct feeding ecologies, Caenorhabditis elegans and Pristionchus pacificus represent a powerful interspecies system for understanding the evolutionary adaptations influencing sensory perception. More specifically, while C. elegans is a microbial feeder, P. pacificus is an omnivorous species and not only feeds on bacteria but is also a predator of other nematodes (11). This expanded feeding behavior is linked to a mouth-form plasticity with P. pacificus nematodes capable of developing one of two morphs. These are either the stenotomatous (st) or eurystomatous (eu) form. The st mouth is narrow with a single tooth and only permits bacterial feeding or scavenging, whereas the eu mouth is wider and possesses an additional tooth which facilitates both bacterial feeding and predation (11). Furthermore, predatory behavior is directed toward not only other nematode species but also con-specifics. However, closely related strains are spared due to the presence of a robust kin-recognition system (12). Crucially, how sensory systems evolved to accommodate the predatory behaviors is an open question.
In C. elegans, bacterial food is detected through well described foraging behavior using their chemosensory and texture sensing systems (13, 14). In P. pacificus, little is known of its food and prey sensing mechanisms, however, they show specific sensory adaptations relevant to their ecology. In particular, P. pacificus are attracted to many bacterial species found concurrently in their environment and are also attracted to specific insect pheromones as Pristionchus are frequently found associated with scarab beetles (15). During food sensing and foraging, P. pacificus transitions between the docile bacterial feeding and aggressive predatory feeding states through the internal balance of octopamine and tyramine. These neuromodulators act on a group of head sensory neurons expressing the corresponding receptors (16). Contact between the nose of the predator and the surface of the prey during aggressive predatory bouts can initiate the activity of their movable tooth to puncture through the cuticle of the prey (11, 17, 18). However, the precise stimuli responsible for detecting prey contacts are currently unknown.
Here, to understand how the P. pacificus sensory systems have contributed to the emergence of predatory feeding we have investigated the role of mechanosensory adaptations for these behaviors. Using a candidate-based approach, we identified a crucial role for mechanosensation in predation. We find Ppa-mec-6 is necessary for this process and may contribute to these behaviors through a previously undescribed predatory-associated mechanosensory channel. Furthermore, we demonstrate that mechanosensation is required alongside chemosensory inputs and that these acts synergistically to execute efficient predation behaviors. We also reveal that chemosensory and mechanosensory components are expressed in the same externally projecting and environmentally exposed neurons, indicating a sensory hub for prey detection. Finally, we show that the functional inhibition of these neurons disrupts predation.
Results
Mechanosensation Is Involved in Prey Detection.
P. pacificus is a voracious predator of other nematode larvae including C. elegans and this feeding behavior requires contact between predator and prey (Fig. 1A). Accordingly, we investigated the importance of P. pacificus mechanosensation for detecting prey and a potential role in the evolution of its predation behaviors. Cel-mec-3 is a critical transcription factor involved in the development and function of mechanosensory neurons in C. elegans, and animals with mutations in this gene fail to respond to tactile stimuli (19). In P. pacificus, using a Ppa-mec-3 transcriptional reporter, we observed expression in several neurons with similar cell body placements to the C. elegans mechanosensory neurons FLP, AVM, ALM, PVM, PVD, and PLM (Fig. 1B), (19?–21). To assess its role in mechanosensation in P. pacificus, we generated a mutation in the Ppa-mec-3 orthologue using CRISPR/Cas9. Similar to C. elegans, we found Ppa-mec-3 mutants fail to react to several classic mechanosensory assays including harsh touch, gentle touch, and nose touch assays (Fig. 1 C–E). Taken together, this indicates a conserved role for Ppa-mec-3 between species. Next, to assess a potential role for Ppa-mec-3 in predatory feeding, we performed previously established corpse assays (11). In corpse assays, starved P. pacificus adults are placed on a plate saturated with C. elegans larval prey and predation success is determined by the number of corpses after a designated time interval (SI Appendix, Fig. 1A). Here, Ppa-mec-3 predators show a significant reduction in killing ability, with fewer larval corpses generated by predators than observed in control assays with wildtype predators (Fig. 1F). Thus, mechanosensory systems have acquired additional functions in P. pacificus and are necessary for efficient prey detection.
*Ppa-mec-3 regulates touch avoidance and prey detection. (A) A Pristionchus pacificus adult predator (Right) attacks a Caenorhabditis elegans larvae (Left). (Scale bar, 100 μm.) (B) Representative image of a P. pacificus adult expressing Ppa-mec-3p::RFP (Top, magenta). Putative P. pacificus neuronal identity is based on C. elegans soma placement and known mechanosensory function. (Scale bar, 100 μm.) (C) Mechanosensory assays to harsh touch, (D) gentle touch, and (E) nose touch. Each assessment is the result of ten consecutive trials of each worm. At least 30 worms were tested per strain. (F) Number of C. elegans corpses counted after 2 h of contact with the indicated P. pacificus strains as predator. At least 10 assays were performed. Statistical tests: two-tailed Wilcoxon–Mann–Whitney, nonsignificant (ns), *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001. Schematics were made with biorender.
Mechanosensory Genes Have Diversified Across Evolution.
Having established the importance of mechanosensation for predation, we next explored the conservation of mechanosensory gene networks across nematode evolution. Frequent gene losses and gains are observed between nematode species and this can be associated with novel evolutionary processes (22). Therefore, we initially conducted a bioinformatic study of the mechanosensory systems across 8 nematode species to investigate the evolution of this sensory modality across the nematode phylum (Fig. 2A). Our analysis consisted of the two free-living species C. elegans and P. pacificus belonging to clade 5 as well as several obligate parasite species of both plants and animals (22). We included orthologues of the C. elegans predicted mechanotransduction channels belonging to the degenerin–epithelial Na+ channels (DEG/ENaC) and transient receptor potential (TRP) gene families (21). We also included other known mechanosensory genes such as mec-6, a paraoxonase-like protein which acts as a chaperone to ensure channel function (23), and pezo-1, due to its reported role in food sensation (24). Compared to free-living nematodes, parasitic nematodes frequently had a lower number of genes encoding mechanotransduction channels, perhaps indicative of adaptations to their specific lifestyle. In contrast, comparisons between the free-living nematodes C. elegans and P. pacificus generally showed a similar representation of gene families with a few exceptions (Fig. 2A). These include the four trpl genes which are specific to C. elegans but absent in other species including P. pacificus (21). Additionally, there are two large gene expansions in P. pacificus among the DEG/ENaC gene family with five copies of deg-1 in P. pacificus compared to one in C. elegans and 11 egas genes in P. pacificus compared to four in C. elegans. Little is known regarding the function of egas in C. elegans and many of the P. pacificus egas paralogues lack RNA-seq data to validate their expression (10). However, deg-1 has been shown in C. elegans to be required for the induction of mechanoreceptor current in the sensory ASH neuron (25). Therefore, mechanosensory gene networks appear relatively stable between C. elegans and P. pacificus but are more stochastic in parasitic nematodes with their more complex life-styles and host interactions.
*Mechanosensation is required for efficient predation. (A) Table analyzing the number of mechanosensory gene paralogues compared to C. elegans. Species of free-living nematodes (brown), plant parasite nematodes (green), and animal parasite nematodes (black) are compared. Their evolutionary tree is presented on the Left with the nematode clade marked. (B) Mechanosensory assays to harsh touch, (C) gentle touch, and (D) nose touch for P. pacificus mutants in putative mechanosensory genes. Each assessment is the result of ten consecutive trials of each worm. At least 30 worms were tested per strain. (E) Number of C. elegans corpses counted after 2 h of contact with the indicated P. pacificus putative mechanosensory mutants as predators. At least 10 assays were performed per mutant. Statistical tests: two-tailed Wilcoxon–Mann–Whitney with Benjamini–Hochberg correction, nonsignificant (ns), *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001. Schematics were made with biorender.
With the identification of genes potentially acting as mechanosensory channels in P. pacificus, we conducted a candidate-based approach to identify components necessary for efficient prey detection through P. pacificus nose contact. Candidates were selected based on known mechanosensory phenotypes in C. elegans together with further validation through available P. pacificus expression data (10, 26). Accordingly, we selected genes to target by CRISPR/Cas9 which included both TRP and DEG/ENaC families as well as other associated mechanosensory genes. We selected the DEG/ENaC channels Ppa-mec-4 and Ppa-mec-10 as well as Ppa-mec-6. In C. elegans, Cel-mec-4 and Cel-mec-10 are both involved in the gentle touch response (21, 27) and Cel-mec-6 has been shown to directly interact with Cel-mec-4 and Cel-mec-10 to aid the function and stability of this mechanosensitive channel (23, 28). We targeted Ppa-osm-9, Ppa-ocr-2, and Ppa-trp-4 together as in C. elegans these genes have an additive effect reducing its nose touch responsiveness (29). Only P. pacificus triple mutants for these genes were analyzed*. C. elegans* nose touch response also requires Cel-delm-1 and Cel-delm-2 which are expressed in glial cells (30). However, we could not identify one-to-one orthologues for these two genes in P. pacificus as both paralogous sequences are closer to each other than to either Cel-delm or Cel-acd genes. Therefore, these were named delm/acd and we mutated both candidates in P. pacificus. In C. elegans, Cel-del-7 and Cel-del-8 are DEG/ENaC ion channel family members, with the former involved in mechanotransduction (31). However, there is only one paralogue in P. pacificus, which we mutated and named Ppa-del-7/8. We also generated a quintuple mutant of all paralogues of DEG/ENaC Ppa-deg-1, and targeted the DEG/ENaC potential proprioception genes Ppa-del-1, Ppa-unc-8, and Ppa-unc-105 (21, 32). To further represent the TRP gene family, we included Ppa-trp-1, Ppa-trp-2, Ppa-pkd-2, Ppa-cup-5, and Ppa-trpa-1, which, in C. elegans, are expressed in many neurons and most have roles in mechanotransduction (29, 30, 33, 34). Finally, we targeted the ionotropic glutamate receptor Ppa-glr-1 due to its role in mediating excitatory neurotransmission in touch receptor neurons (35), and Ppa-pezo-1 due to potential roles in food sensing (24). This resulted in 26 genes of interest, and we investigated two alleles for each of these genes, which were predicted to cause loss-of-function mutations (SI Appendix, Table S1 and Dataset S1).
Ppa-mec-6 Is Required for Efficient P. pacificus Predation.
Initially, we tested all of our mutants for mechanosensory defects using the three touch assays adapted from previous C. elegans studies. These assays measure the animal response to harsh touch, gentle touch, and nose touch (21). The majority of the P. pacificus mutants did not show mechanosensory deficient phenotypes. However, Ppa-mec-4, Ppa-mec-10, and Ppa-mec-6 showed the strongest mechanosensory defects similar to those described in C. elegans, and also similar to those observed in Ppa-mec-3, resulting in a response to touch only half the time or less (Fig. 2 B–D and SI Appendix, Fig. 1 B–D). Furthermore, all four mutants were also more lethargic than wildtype worms, phenocopying C. elegans mechanosensory mutants (27). Thus, as in C. elegans, Ppa-mec-4, Ppa-mec-10, Ppa-mec-3, and Ppa-mec-6 are required for touch sensation in P. pacificus. While previous reports in C. elegans have not identified a role for harsh touch sensation in Cel-mec-4 (29), this discrepancy between the two species may stem from subtle species-specific sensitivity differences. Next, we tested all of our mutants for defects in predation using the previously described corpse assays. While the majority of mutants displayed wildtype levels of predation, reduced killing was observed in Ppa-mec-6 (Fig. 2E and SI Appendix, Fig. 1E). This is similar to the defect observed in the Ppa-mec-3 mutants (Fig. 1F). Importantly, no predatory defect was observed in Ppa-mec-4 or Ppa-mec-10, indicating that the predatory abnormality is not the consequence of a general mechanosensory defect, but rather is specific to the function of Ppa-mec-3 and Ppa-mec-6. In C. elegans, Cel-mec-6 is involved in the assembly and function of mechanosensory ion channels and interacts with Cel-mec-4 and Cel-mec-10 to form ion channel complexes that are essential for mechanotransduction (23, 28). Our data indicate that while the Ppa-mec-4 and Ppa-mec-10 channels are not required for prey detection in P. pacificus, Ppa-mec-6 may function as part of an additional, as yet unknown, mechanosensory ion channel involved in detecting prey contact. Thus, specific mechanosensory inputs are necessary for prey detection in P. pacificus. However, as predation is not fully abolished, additional sensory inputs also contribute to prey detection ability.
Mechanosensation and Other Predatory-Associated Traits.
Predation in P. pacificus is dependent on the formation of the eu mouth morph which is determined by a multitude of environmental and genetic factors (36, 37). To assess if mechanosensation also influences the mouth morph fate, we screened our mutant library for any effect on mouth form ratio. All mutants showed wildtype levels of eu to st morphs (SI Appendix, Fig. 2 A and B). In addition, P. pacificus has a robust kin-recognition mechanism which prevents attacks on its own progeny and close relatives, while facilitating the cannibalism of other con-specific competitors (12). Due to the prey detection defect observed in Ppa-mec-3 and Ppa-mec-6 we also tested these mutants for kin-recognition defects. For both Ppa-mec-3 and Ppa-mec-6 mutants, kin-recognition was robustly maintained indicating no deficiency in this process (SI Appendix, Fig. 2C). Therefore, defects in mechanosensation are specific to prey detection and do not influence other predatory traits.
Chemosensation and Mechanosensation Synergistically Influence Prey Detection.
The partial defect in prey detection observed in the Ppa-mec-3 and Ppa-mec-6 mutants is similar to that previously described in cilia deficient mutants (17). Cilia are complex organelles, and in nematodes they are essential for the detection of many environmental cues. In both C. elegans and P. pacificus, some of the strongest cilia deficiencies are observed in mutants defective for the transcription factor, DAF-19, which acts as the master regulator of ciliogenesis (17, 38, 39). Curiously, while mutations in Cel-daf-19 cause aberrant dauer entry, no abnormal dauer phenotypes result from Ppa-daf-19 mutations in P. pacificus, reinforcing the distinct evolutionary trajectories regulating this developmental process between these species (40). Furthermore, accompanying the numerous chemosensory defects in Ppa-daf-19, these mutants are also defective in prey detection (17). Therefore, to disentangle the roles of chemosensation and mechanosensation, we assessed the Ppa-mec-3 and Ppa-mec-6 mutants alongside Ppa-daf-19. We found that while Ppa-daf-19 mutants showed chemosensory defects in their attraction to a bacterial food source and in their aversion to octanol, both Ppa-mec-3 and Ppa-mec-6 responded as wildtype animals to these cues (Fig. 3 A and B). This indicates both Ppa-mec-3 and Ppa-mec-6 are not defective for chemosensation. In contrast, while both Ppa-mec-3 and Ppa-mec-6 are mechanosensory defective, the touch response of Ppa-daf-19 mutants was similar to wild type animals (SI Appendix, Fig. 3 B–D). A similar observation has also been reported in C. elegans, where only mild mechanosensory defects are observed in Cel-daf-19 mutants (41). Thus, mechanosensory defects appear specific to Ppa-mec-3 and Ppa-mec-6, while *Ppa-daf-*19 mutants are mostly associated with chemosensory defects. As both groups of mutants are defective for prey detection, but neither fully abrogates the P. pacificus predatory abilities, we next assessed if these sensory inputs act synergistically. The Ppa-mec-6; Ppa-mec-3 double mutants phenocopied the Ppa-mec-6 single mutants revealing no additive effect in mechanosensory defects. However, we found that predation defects were further exacerbated in double mutants of Ppa-mec-6; Ppa-daf-19 or in Ppa-mec-3; Ppa-mec-6; Ppa-daf-19 triple mutants although, crucially, some killing was still maintained (Fig. 3C). Therefore, chemosensation and mechanosensation act together for prey detection. Remaining killing abilities may act through some residual function in these sensory pathways or through another as yet unidentified stimuli.
*Chemosensation and mechanosensation synergically influence prey detection. (A) Chemotaxis index analyzing the aversive response of Ppa-mec-3, Ppa-mec-6, and Ppa-daf-19 to 1-octanol. Each genotype was assessed 10 times. (B) Percentage of Ppa-mec-3, Ppa-mec-6, and Ppa-daf-19 mutants finding a bacterial OP50 food source after 8 h. Each genotype was assessed 10 times. (C) Number of C. elegans corpses generated after two hours of exposure to the indicated P. pacificus mutants as predator. 30 assays were performed. (D) Schematic of behavioral tracking and machine learning workflow to track feeding behavior and determine behavioral states based on a Ppa-myo-2::RFP pharyngeal fluorescence marker (16). This pipeline uses the most predictive behavioral features (velocity and pumping rate) to visualize behavioral states on probability density maps. Predatory states occupy a low velocity space. It also determines the mean fraction of time spent in each behavioral state per animal. (E) Joint probability density map of velocity (μm/s) and pumping rate (Hz) for animals corresponding to the genotypes Ppa-mec-6, Ppa-daf-19, and Ppa-mec-6; Ppa-daf-19. The number of worms is indicated in the top right corner. (F) Time spent in each behavioral state normalized to the total track duration. Statistical tests: Significance from comparison to wild-type (black), to mec-3 single mutant (blue) and to daf-19 single mutant (green) was assessed using a two-tailed Wilcoxon–Mann–Whitney with Benjamini–Hochberg correction (A–C), Mann–Whitney U test with a Bonferroni correction (F). nonsignificant (ns), *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001. Schematics were made with biorender. Predatory (Pr.)
Behavioral Tracking and State Predictions.
Quantifying the number of C. elegans corpses generated by P. pacificus predators provides a robust assessment of overall killing ability (11). However, understanding the sensory-motor transformations underpinning prey contact requires a more detailed behavioral analysis. We recently developed an automated behavioral tracking and machine learning model to identify and quantify aggressive behavioral states associated with predation and further analyze these complex behaviors (16, 42). Using this method, the P. pacificus behavioral repertoire can be divided into six states. These consist of three predatory associated states including “predatory search,” “predatory biting” and “predatory feeding” (Fig. 3D). Additionally, three nonpredatory behaviors including a ‘dwelling’ state and two ‘roaming’ states exist which are similar to behavioral states observed in C. elegans (14, 16). The two most predictive features of predation states are velocity and pumping rate. By observing the joint distribution of these two features, it is possible to visualize the prevalence of predatory and nonpredatory states (Fig. 3D). We used a machine learning model that was previously trained on behavioral data using both unsupervised and supervised methods (16) to investigate behavioral state occupancy in our predation mutants compared to wildtype animals (Fig. 3 D–F and SI Appendix, Fig. 3 E–G). In the mechanosensory defective Ppa-mec-6 and Ppa-mec-3, as well as the chemosensory defective Ppa-daf-19 mutants, we found a significant increase in the duration and total time spent in the “predatory search” state. This may be indicative of a compensatory mechanism whereby the loss of one sense enhances utilization of the remaining senses. This is further supported by the absence of the increased “predatory search” state in mutants lacking both sensory modalities (Fig. 3F). Instead, these Ppa-mec-6; Ppa-daf-19 double mutants show a significant increase in the nonpredatory “search” and “roaming” states, indicating a switch away from predatory states and into exploratory behaviors. This is similar to observations in C. elegans where the loss of one or multiple sensory modalities results in modifications to the performance of the remaining senses and induces behavioral adjustments (27). Furthermore, “predatory biting” state occupancy was similar to wildtype animals in both Ppa-mec-6 and Ppa-mec-3 mutants while, in Ppa-daf-19 this state was significantly reduced. Furthermore, in Ppa-daf-19 there are additional roaming defects (SI Appendix, Fig. 3G) which are similar to observations in Cel-daf-19 (38). Therefore, predatory behavioral states are modulated through signals received through the P. pacificus sensory systems with both chemosensory and mechanosensory modalities influencing distinct but overlapping aspects of prey detection.
IL2 Neurons Represent a Sensory Hub for Prey Detection.
As distinct mechanosensory and chemosensory inputs contribute to the P. pacificus predatory abilities, we investigated if they act through discrete independent neurons, or overlapping sensory pathways. To resolve this, we generated a reporter strain expressing Ppa-mec-6p::Venus and Ppa-daf-19p::RFP and identified the specific prey detection neurons involved. Ppa-mec-6p::Venus and Ppa-daf-19p::RFP colocalized and were robustly expressed in the P. pacificus IL2 neurons (Fig. 4 A and B). These were identified based on soma position and their unique morphological features (43). Importantly, the anterior processes from this group of six neurons project into the external environment through exposed sensory endings making them strong candidates for prey detection (Fig. 4C). Additionally, we observed that Ppa-mec-6 is coexpressed with Ppa-mec-3 in the putative P. pacificus ALM and PLM body neurons and the FLP head neurons (SI Appendix, Fig. 4 A and B). Notably, while Ppa-mec-6 is observed in the IL2 neurons, we did not detect Ppa-mec-3 in these cells. This divergence may account for the different prey detection phenotypes observed between Ppa-mec-3 and Ppa-mec-6 mutants. As in C. elegans, many sensory neurons in P. pacificus are ciliated and this is not restricted to the IL2 sensory neurons (39). Therefore, it is somewhat surprising that the most robust Ppa-daf-19 expression is observed in the IL2 neuron cluster, although there is also low expression in a few other head neurons (Fig. 4B). However, as there are several large introns in the Ppa-daf-19 gene structure which contain enhancers and other epigenetic signatures (44), we predict these may contribute additional regulatory elements for further expression in other ciliated neurons. These may currently be missing from our Ppa-daf-19p::RFP reporter. Thus, both mechanosensory and chemosensory prey detection systems are integrated within the same sensory hub with projections exposed to the external environment. These likely act as the first point of contact with potential prey.
*Predation is disrupted by inhibition of Ppa-mec-6 expressing cells. (A) Head of a worm expressing mec-6p::Venus (Top, green) and daf-19p::RFP (Middle, magenta). Coexpression of both Ppa-mec-6 and Ppa-daf-19 visible in the merge image. IL2 neurons are indicated. (Scale bar, 100 μm.) (B) Representative image of a worm expressing Ppa-mec-6p::Venus (Top, green) and Ppa-daf-19p::RFP (Middle, magenta) and the merge (Bottom). (Scale bar, 100 μm.) (C) Scanning electron microscopy image of the anterior of a P. pacificus adult. The P. pacificus mouth opening is surrounded by protrusions of the 6 inner labial IL2 sensilla neurons (arrow). (Scale bar, 3 μm). (D) Mechanosensory assays of nose touch for P. pacificus worms for each condition: wild-type worms treated with histamine and worms expressing HisCl channel under the control of Ppa-mec-6 promotor in absence (–) or presence (+) of histamine. Each datapoint is the result of ten consecutive trials on an individual worm. At least 30 worms were tested per condition. (E) Number of C. elegans corpses generated after 2 h of exposure to the indicated P. pacificus worms as predator in absence (–) or presence (+) of histamine. 20 assays were performed. (F) Joint probability density map of velocity (μm/s) and pumping rate (Hz) for animals corresponding to condition described in (D). The number of worms is indicated in the Top Right corner. (G) Time spent in each behavioral state normalized to the total track duration. (H) Putative model of the evolutionary diverse role for mechanosensation between C. elegans and P. pacificus. In both species, mechanosensation triggers an avoidance response which rely on mec-3, mec-6, mec-4, and mec-10. In addition, in P. pacificus, contact can trigger predatory behavior when the IL2 perceive mechanical or chemical stimuli. The perception of the predatory specific mechanical stimuli, acts through the mechanosensory pathway components Ppa-mec-3 and Ppa-mec-6. Statistical tests: Significance from comparison to wild-type (black) and to Ppa-mec-6p::HisCl untreated worms (brown) was assessed using two-tailed Wilcoxon–Mann–Whitney with Benjamini–Hochberg correction (D–E), Mann–Whitney U test with a Bonferroni correction (G). nonsignificant (ns), *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ***P ≤ 0.0001. Predatory (Pr.) Schematic was made with biorender.
Functional Inhibition of Ppa-mec-6 Expressing Neurons Disrupts Mechanosensation and Predation.
Finally, given the integration of sensory stimuli in the IL2 neurons and the importance of Ppa-mec-6 for prey detection, we investigated the requirements of Ppa-mec-6 expressing cells for predation. We expressed a histamine-gated chloride channel (HisCl) under the control of the Ppa-mec-6 promoter. This method, recently established in P. pacificus, enables inducible inhibition of target neurons upon the exogenous addition of histamine as neurons become hyperpolarized (45). We first verified that the construct and treatment did not affect worm health. Developmental time was the same across wildtype and transgenic Ppa-mec-6p::HisCl animals, both alone and in the presence of histamine, indicating no adverse effects (SI Appendix, Fig. 4C). Furthermore, mouth form also remained at wildtype ratios indicating Ppa-mec-6 expressing cells do not influence this trait (SI Appendix, Fig. 4D). Next, we assessed the requirement of the Ppa-mec-6 expressing neurons for mechanosensation and chemosensation. Both body touch and nose touch response were significantly reduced upon silencing of the Ppa-mec-6 positive neurons although the effect on nose touch was much greater than the body touch response consistent with its robust expression in head neurons (Fig. 4D and SI Appendix, Fig. 4 E and F). We also observed no defect in the response to our chemosensory stimuli upon neuronal silencing (SI Appendix, Fig. 4 G and H). Finally, we tested the consequences of this inhibition on predation. Both the number of corpses and prevalence of predatory states were reduced when Ppa-mec-6p::HisCl expressing worms were treated with histamine (Fig. 4 E–G and SI Appendix, Fig. 4I). Automated identification of the different behavioral states indicated that time dedicated to all three predatory behaviors—search, biting, and feeding were also reduced. These reductions were accompanied by an increase in two of the exploratory behaviors—search and roaming. Notably, silencing all Ppa-mec-6 expressing cells has a stronger effect on predation than has been previously reported by the silencing of the IL2 neurons alone (16). Thus, neurons expressing Ppa-mec-6 are required for efficient predation.
Discussion
A central aspect associated with the evolution of predatory behaviors hinges on an organism’s ability to detect potential prey. Fundamental to this are adaptations to sensory abilities that enable an organism to perceive, process, and respond to potential prey cues and direct predatory behaviors appropriately. In the nematode P. pacificus, we have found that the evolution of predation is associated with the diversification and specialization of both mechanosensory and chemosensory modalities to also encompass prey detection (Fig. 4H). This is beyond their previously described functions in the model species C. elegans. The co-option and neofunctionalization of sensory systems are a frequently observed phenomenon. For example, the lateral line system, used to detect water movements and vibrations in many fish species, has evolved electroreception capabilities in sharks, rays, and skates (46). Similarly, certain snake species including pit vipers, pythons, and some boas have evolved infrared detection through adaptations to the temperature-sensitive skin thermotransduction receptor TRPA1. This enables these species to hunt warm blooded prey in near total darkness, while in other snakes it is a mechanism to regulate body temperature (47). Examples of sensory co-option events are not restricted to animals and can also be readily found in plants including carnivorous Venus flytrap and sundew species, whereby ancestral mechanosensory channels are instead used for sensing and prey capture (48). Therefore, our findings support a well-established link between sensory system evolution and behavioral diversity across a wide range of taxa.
In P. pacificus, the co-option and integration of multiple sensory modalities, suggests that predation is not solely dependent on enhancements and adaptations to a single sensory pathway but rather on the evolution of multiple interacting systems which fine-tune prey capture. In this nematode species, it is both chemosensation and mechanosensation that are required for efficient predation. Similar multimodal mechanisms of predation can be observed in other invertebrates, including other species which also lack visual inputs or those which hunt in low light environments. These include species of octopus, which utilize chemotactile receptors on their tentacles to identify potential prey through a mechanism of contact-dependent aquatic chemosensation (49). Additionally, cone snails are carnivorous mollusks that hunt fish, worms, or other snails and utilize chemosensation to first detect the presence of chemicals secreted by their prey, and subsequently use vibration sensing and mechanoreception to determine prey position. They then shoot a venomous harpoon-like tooth to immediately immobilize and feed on their target (50). In P. pacificus, our data show that both chemosensation and mechanosensation are important for predation, and defects in either system significantly reduce killing ability. Additionally, from our behavioral state studies of these mutants, we find defects in Ppa-daf-19 chemosensory mutants are more severe and are associated with aberrations in the “predatory biting” state, alongside increased “predatory search” occupancy. A similar “predatory search” occupancy increase is also detected in Ppa-mec-3 and Ppa-mec-6 mutants. We hypothesize that the increased “predatory search” state occupancy in both sensory modalities is a compensatory mechanism whereby P. pacificus switches dependency to the unimpaired sense in an attempt to maintain its predatory behaviors. This is further validated by the loss of this increase in mutants lacking both the predatory-associated mechanosensory and chemosensory components. Indeed, sensory loss in C. elegans has also been shown to alter the performance of remaining sensory modalities resulting in distinct behaviors (27). Furthermore, while the mechanosensory Ppa-mec-6 and Ppa-mec-3 as well as chemosensory Ppa-daf-19 mutants show reduced killing abilities, Ppa-mec-6 and Ppa-mec-3 mutants were found to occupy “predatory biting” states similar to wildtype, while this state was reduced in Ppa-daf-19. Therefore, both sensory inputs likely have specific functions during predation, such that chemosensation is potentially required throughout these behaviors while mechanosensory systems may be associated with the initial steps of the predator–prey contact.
Both daf-19 and *mec-*3 are transcription factors and influence genes associated mostly with chemosensation and mechanosensation respectively (19, 39). This makes it difficult to disentangle the precise molecular mechanisms associated with the emergence of the predatory behavior in P. pacificus using these mutants alone. However, by focusing on the role of mechanosensation, we have been able to identify Ppa-mec-6 as a specific molecular component that has acquired additional functions in P. pacificus associated with its predatory feeding. In C. elegans, Cel-mec-6 is a known to interact with several DEG/EnAC type subunits with the best characterized being Cel-mec-4 and Cel-mec-10 which form a mechanosensory ion channel responsible for the gentle touch response (23, 28, 51). Within this complex Cel-mec-6 is a small auxiliary protein that is not a pore-forming subunit itself, but is necessary for proper channel function. In P. pacificus, we find mutations in any of Ppa-mec-4, Ppa-mec-10, or Ppa-mec-6 result in mechanosensory defects, but it is only Ppa-mec-6 that has additional predation-specific abnormalities. Thus, while Ppa-mec-6 may interact with Ppa-mec-4 and Ppa-mec-10 to regulate mechanosensation as in C. elegans, it is also likely involved with other as yet unknown components and may form a distinct mechanosensory channel involved in prey detection. These components require further identification but mechanosensory genes with expanded families in P. pacificus are prime candidates. These include the egas gene family which is greatly increased in P. pacificus as well as asic-2 that is expressed in potential neurons of interest (52).
By analyzing the localization of Ppa-mec-6, we observed robust expression in several head neurons. These include the P. pacificus FLP neurons as well as the six head sensory IL2 neurons where it is also expressed in C. elegans (20). All these neurons are easily identified in P. pacificus by their morphology and soma location, although the IL2 neurons show a distinct placement compared to C. elegans (43, 53). In C. elegans, IL2 neurons regulate dauer formation and nictation behaviors, and are thought to be both chemosensory and mechanosensory (54, 55). Furthermore, they release extracellular vesicles potentially involved in communication (52, 56). The FLP neurons in C. elegans are also polymodal detecting both thermosensory and mechanosensory cues and they are involved in its response to nose touch (57). Our results show that in P. pacificus, inhibition of the Ppa-mec-6 expressing cells including the IL2 and FLP neurons additionally prevents the acquisition of sensory information necessary for prey detection. Notably, the silencing of Ppa-mec-6 expressing cells induces a stronger defect than has been reported by silencing the IL2 cells alone (16). In P. pacificus, the FLP neurons show altered connectivity compared to C. elegans, and this has been proposed to reflect a shift in function from aversion to attraction (43). This shift has been suggested as a potential predatory adaptation, and our data further support a role for the FLP neurons in predatory behavior. Crucially, recent studies in P. pacificus also identified the IL2 neurons as regulators of predatory feeding states which depend on the balancing actions of octopamine and tyramine (16). Octopamine induces predatory states while tyramine initiates more docile bouts and IL2 neurons express octopamine receptors necessary for the predatory states in P. pacificus. Accordingly, our findings that these neurons express Ppa-mec-6 as well as Ppa-daf-19 reinforces their importance as prime candidates for detecting and determining prey contact events. Moreover, the integration of both mechanosensory and chemosensory inputs, as well as behavioral state modulatory mechanisms in the P. pacificus IL2 neurons demonstrates that even relatively simple circuits can be remodeled to accommodate complex and dynamic behaviors including predation. Therefore, taken together, our findings provide important insights into how sensory systems are co-opted and refined to generate behavioral diversity, and demonstrate the importance of sensory perception and evolutionary adaptations for establishing novel behavioral traits.
Materials and Methods
Worm Maintenance.
P. pacificus (PS312 and derived strains) or C. elegans (N2), were maintained on nematode growth media (NGM) 6 cm plates seeded with 300 μL Escherichia coli (OP50) at 20 °C. All P. pacificus strains used are listed in SI Appendix, Table S1. The C. elegans N2 strain was provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440).
Orthologue Identification.
We generated a mechanotransduction channel list from wormbook (21) and obtained published genome assemblies for eight nematode species belonging to different clades identified by Blaxter et al. (22). These species include: P. pacificus (10), C. elegans (release WS271, 2019, WormBase website (58)), Strongyloides ratti (59), Bursaphelenchus xylophilus (60), Brugia malayi (61, 62), Ascaris suum (63), Romanomermis culicivorax (64), and Trichinella spiralis (65). When multiple annotated isoforms were available, only one was considered for analysis. Orthologous were determined using OrthoFinder (66).
Generation of New Strains.
Mutations were induced by CRISPR/Cas9 (IDT) by coinjection of the sgRNA for the gene of interest while using a sgRNA targeting Ppa-prl-1 as a marker as previously described (67). sgRNAs were made by mixing crRNA (IDT) and tracrRNA (IDT) in equal volume and quantity and incubated for 5 min at 95 °C. After letting them cool down for 5 min at room temperature, up to three sgRNA were mixed with the Cas9 and 5 min later Tris-EDTA (TE, Promega) was supplemented to reach the concentration of 18.1 μM for sgRNA and 12.5 μM for Cas9. This mix was centrifuged for 10 min at maximum speed at 4 °C. The resulting mix was used to inject P. pacificus adults in the gonads area (Axiovert 200, Zeiss and Eppendorf set up). P0 injected worms were singled out and laid eggs overnight. Once the progeny had developed enough, plates were screened for a roller phenotype induced by the dominant mutation in Ppa-prl-1 pointing to successful injection. Around 50 worms were genotyped from identified plates for the gene of interest using PCR (Qiagen) followed by sanger sequencing (Eurofins). The sgRNA and primer sequences used as well as the wild-type sequences and generated mutation are provided in Dataset S1. Two guide RNA were used for Ppa-mec-3 to target both isoforms. All mutations generated are predicted to lead to an early stop codon or frameshift with the exception of one allele of Ppa-mec-3 (bnn61) and one allele of Ppa-trp-2 (bnn51) presented in supplementary figures. Mutations were generated in the P. pacificus PS312 strain.
Behavioral tracking and state predictions required an integrated Ppa-myo-2p::RFP (16). In addition, mouth morph ratio is influenced by Ppa-daf-19 which results in a strong st bias unsuitable for predation assays (17). Therefore, predatory morphs eu were induced alongside Ppa-daf-19 through generation of additional mutations in Ppa-nag-1; Ppa-nag-2 (17) in the Ppa-myo-2p::RFP background. Accordingly, this strain JWL118 has 100% Eu mouth morphs (SI Appendix, Fig. 3A). For experiments involving Ppa-daf-19, all comparative experimental strains also carry Ppa-nag-1; Ppa-nag-2 mutations based on the control strain JWL147 (SI Appendix, Table S1).
Generation of Reporter Lines.
TurboRFP and GFP sequences optimized for P. pacificus were retrieved from plasmid pZH009 and pZH008 respectively (9). A Venus reporter was optimized for P. pacificus as previously described (9). Promotor sequences consisted of the indicated number of nucleotides preceding the ATG according to P. pacificus genome (10): Ppa-mec-6 (949 bp), Ppa-mec-3 (932 bp), Ppa-daf-19 (719 bp) promotor sequences. Ppa-daf-19p::RFP (pMR027, Genscript) and Ppa-mec-6p::Venus (pMR022, eurofins) were made by gene synthesis and Ppa-mec-3p::RFP (pMR005) by cloning. For generating stable transgenic lines, animals were injected with a mix containing 60 ng/μL purified gDNA (NEB Monarch purification kit) along with 5 to 10 ng/μL plasmid of interest all digested with HindIII. Prior to each injection, the mix was centrifuged at maximum speed at 4 °C for 10 min. Injections were made in P. pacificus adult gonads (Axiovert 200, Zeiss, and Eppendorf set up). Progeny worms carrying the construct of interest were identified using the epi-fluorescence microscope (Axio Zoom V16; Zeiss). Confocal images were acquired using a stellaris microscope (Leica) with a 63× objective at water immersion.
Chemosensory Assays.
Attraction toward OP50 was tested using previously developed assays with minor modifications (39). 20 μL of an OP50 overnight culture was pipetted onto a 6 cm unseeded plate 1.5 cm from the edge and left at room temperature overnight. Plates were then kept in the fridge until use. 50 adult worms from plates almost depleted of food were placed at the opposite side of the bacterial lawn at around 1.5 cm of the border making sure to not transfer any bacteria when doing so. After 8 h the number of worms at the bacterial lawn and the total number of worms still alive in the plates were determined. Plates were scored if they had at least 30 worms left at the end of the assay.
To assess worm behavior toward 1-octanol, we adapted existing protocols (40). Adult worms were washed off with M9 and left on an unseeded plate for 3 h to remove bacteria. Prior to starting the assay, two drops of 1.2 μL of sodium azide 1 M was deposited 3 cm apart on opposite side of a 6 cm unseeded plate. 100 worms were placed in the middle line between the two drops and immediately 1μL of 100% ethanol was put as control on top of one of the sodium azide deposits and 1 μL of 100% 1-octanol (sigma-aldrich) over the other. The assay plates were left for 16 h at 20 °C before assessing worm position. Only worms within a 2 cm radius of the drop up to the midline were taking into account leading to a total of at least 40 worms. Chemosensation index was calculated as the ratio between the number of worm closer to the 1-octanol minus those closer to the ethanol relative to the total number of worms. For both assays 10 replica were made over at least three different days.
Touch Assays.
Touch assays were performed as described for C. elegans with minor modifications (21). Worms were transferred to unseeded plates and left to recover for several minutes prior to being utilized in one of the three assays. Harsh touch assays consisted of touching the worm between its vulva and its tails with the platinum pick used in every day worm maintenance. For the gentle touch assays, the worm was caressed with an eyelash (taped to a wood skewer pick for handling) between its vulva and its tails. For both assays, each worm was tested ten times and the number of times that trigger a reaction was assessed. Due to the fact that some of our strains appear lethargic we did not only consider forward movement as a reaction but any change in worm state occurring upon contact. To assess sensation of the nose, the eyelash was presented to the nose of the worm ten times and again all signs of detection were considered (backward movement or head bending). At least 30 worms over at least 3 different days were assessed for each test and strain.
Corpse Assay.
Predatory behavior was tested as previously described with modifications (11, 16). C. elegans larvae were collected from plates depleted in food just after hatching using M9 and passed through 20 µm filter to remove the other developmental stages. After centrifugation (2,000 rpm, 1 min), ~1 μL of larvae was put off center of a 6 cm unseeded plate, see SI Appendix, Fig. 1A. We visually assessed each plate to maintain a consistent and saturated larval environment for each assay plate. In parallel, predators consisting of young adult P. pacificus of the strain of interest were picked onto another unseeded plate. Both predators and prey were left for 2 h to allow the larval prey to spread and predator to starve. 5 predators were added on the plates and a copper arena 2.25 cm^2^ was put around them. Assays were left for 2 h then two pictures of the arena were taken at ~20× (each field covered a quarter of the arena) at least 10 s apart (Axio Zoom V16; Zeiss). The two images were processed with Fiji using the “max” set up and corpses were manually counted on the resulting image. To assess kin recognition PS312 P. pacificus larvae were used as prey with 20 predators and the assay took place without the arena for 24 h. Corpses were counted under the bench microscope (stemi 508, Zeiss). For both assay 10 replicates were conducted at least three different days. To increase the power of our assay and detect smaller variation between strain, 30 replicas were made for Fig. 3.
Mouth Form Assessment.
Worms were grown in OP50 plates until a high number of adults was reached. Worms were then washed in M9 and placed on a slide containing sodium azide to immobilize them. The number of eu and st worms were counted under an Axiovert 200 (Zeiss) at 10× or 20×. Mouth form was controlled from 5 different plates over at least three different days with a minimum of 30 animals scored per assay.
Feeding assay with PharaGlow.
PharaGlow experiments were performed as described previously (16). Briefly, adult worms with a fluorescent pharynx marker (Ppa-myo-2p::RFP) were picked onto an unseeded plates and left for a 2 h starvation period. 30 worms were picked onto unseeded assay plates containing C. elegans larvae made as for the killing assays (see above) but with twice the density to saturate the plate in larvae. To maintain the worms inside the field of view, a copper arena 2.25 cm^2^ was put around the assay animals. Worms were acclimated to their new environment for 15 min before recording. Recording was made with an epi-fluorescence microscope (Axio Zoom V16, Zeiss) at 1× effective magnification (60 N-C ⅔’’ 0.5 x, Zeiss) and Basler camera (acA3088-57um, BASLER). Light intensity was set at 100% and acquisition time was adjusted to use the full range of intensity without saturating the signal. Images were acquired at 30 Hz for 10 min. Images were then compressed and processed with the python package PharaGlow (https://github.com/scholz-lab/PharaGlow), (42) adapted for P. pacificus (16) with the following parameters (subtract:1, smooth:3, dilate:3, tfactor:0.9, thresholdWindow:50, bgWindow:500, length:70, watershed:100, minSize:250, maxSize:1,000, searchRange:10, memory:30, minimalDuration:1,800, widthStraight:10, pad:5, nPts:50, linewidth:2). The quality of extracted movies was manually assessed and any worms with no sign of activity throughout the recording length were considered dead or damaged and removed from the analysis. Postprocessing to calculate the velocity and pumping rate was done using a previously developed python script and finally automatic behavioral prediction were assigned using the git-hub package PpaPred as described previously (https://github.com/scholz-lab/PpaPred), (16). Worms were recorded over at least three different days.
Inhibition of Neurons by Histamine Chloride Channels.
The Ppa-mec-6 promotor from our reporter construct Ppa-mec-6p::Venus (pMR022) was cloned in front of the P. pacificus codon optimized histamine chloride channel from plasmid pLJ149 (16) forming plasmid pLB010. Transgenic line JWL237 carrying both pMR022 and pLB010 was generated as described above in the wildtype PS312 background. JWL237 was then crossed in the Ppa-myo-2p::RFP background (JWL27) for tracking experiments. Only worms displaying green fluorescence were used for experiments.
Histamine plates were made by the addition of histamine chloride to the NGM before pouring to reach a final concentration of 10 mM. Plates were kept and used in the dark to preserve the histamine. Touch, chemosensation, and feeding assays were made as described above with worms transferred to the histamine plates at least 10 min before performing the assay or from the starvation period when applicable. For corpses assays, copper arenas were not used as they reacted with the histamine chloride and due to the wide field of exploration, images could not be taken and instead corpses were counted manually after 2 h exposure to predators. For developmental assay and mouth form assessment, histamine plates were seeded as usual with 300 μL of OP50 and around 100 J2 larvae were added following filtration (20 µm filter, Millipore). After 4 d at 20 °C, the number of eu or st animals were assessed.
Data Representation and Statistical Analysis.
Box plot illustrates data distribution with quartiles Q1, Q2, and Q3 representing the 25th, 50th, and 75th percentiles, respectively. The box, bounded by Q1 and Q3, includes a thicker line at the median (Q2). Whiskers show the range where most values fall. Analysis of chemosensory assay, touch assay, corpses assays, and mouth form ratio was made with R software and its package prettyR with significance determined using two-direction Wilcoxon–Mann–Whitney with Benjamini–Hochberg correction (68). Analysis of the state prediction results (relative time in state, mean bout duration, transition rates) were done with python as described previously (16) relying on Wilcoxon–Mann–Whitney with Bonferroni correction. In figures, statistics results are displayed as follow: nonsignificant (ns), *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001.
Supplementary Material
Appendix 01 (PDF)
Dataset S01 (XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1P. Oteiza, M. W. Baldwin, Evolution of sensory systems. Curr. Opin. Neurobiol. 71, 52–59 (2021).34600187 10.1016/j.conb.2021.08.005 · doi ↗ · pubmed ↗
- 2T. O. Auer , Olfactory receptor and circuit evolution promote host specialization. Nature 579, 402–408 (2020).32132713 10.1038/s 41586-020-2073-7PMC 7100913 · doi ↗ · pubmed ↗
- 3C. S. Mc Bride , Evolution of mosquito preference for humans linked to an odorant receptor. Nature 515, 222–227 (2014).25391959 10.1038/nature 13964 PMC 4286346 · doi ↗ · pubmed ↗
- 4A. Wada-Katsumata, J. Silverman, C. Schal, Changes in taste neurons support the emergence of an adaptive behavior in cockroaches. Science 340, 972–975 (2013).23704571 10.1126/science.1234854 · doi ↗ · pubmed ↗
- 5H. Jiao , Loss of sweet taste despite the conservation of sweet receptor genes in insectivorous bats. Proc. Natl. Acad. Sci. U.S.A. 118, e 2021516118 (2021).33479172 10.1073/pnas.2021516118 PMC 7848599 · doi ↗ · pubmed ↗
- 6Y. Toda , Early origin of sweet perception in the songbird radiation. Science 373, 226–231 (2021).34244416 10.1126/science.abf 6505 · doi ↗ · pubmed ↗
- 7Z. Musilova , Vision using multiple distinct rod opsins in deep-sea fishes. Science 364, 588–592 (2019).31073066 10.1126/science.aav 4632 PMC 6628886 · doi ↗ · pubmed ↗
- 8G. G. Eren, M. Roca, Z. Han, J. W. Lightfoot, Genomic integration of transgenes using UV irradiation in Pristionchus pacificus. Micro Publ. Biol. 2022, e 000576 (2022).10.17912/micropub.biology.000576 PMC 918722335693893 · doi ↗ · pubmed ↗