In Vivo Evidence of Myofascial Force Transmission Along the Posterior Spiral Chain: Functional Connectivity Linking the Contralateral Latissimus Dorsi, Thoracolumbar Fascia, and Gluteal Region
Saverio Colonna, Greta Maietti, Federico Cuoghi

TL;DR
This study shows that manipulating the posterior spiral chain affects trunk rotation, supporting the role of myofascial force transmission in body mechanics.
Contribution
Provides in vivo evidence of functional connectivity in the posterior spiral chain through myofascial force transmission.
Findings
Isometric activation of the PSC caused a redistribution of trunk rotation with increased left rotation and reduced right rotation.
Stretching the PSC partially rebalanced the asymmetry induced by activation, with a direction-specific effect on rotational balance.
Total rotational range of motion remained largely unchanged despite altered rotational patterns.
Abstract
Myofascial force transmission has been proposed as a mechanism by which mechanical stimuli applied to one body region can influence the behavior of distant segments through fascial continuity. The posterior spiral chain (PSC), linking the latissimus dorsi, thoracolumbar fascia, and contralateral gluteal region, has been described anatomically, but its functional relevance in vivo remains incompletely understood. A single-group pre-post experimental study was conducted on 73 healthy participants. Trunk rotation in the transverse plane was assessed using inertial digital goniometry under four conditions: two baseline assessments, isometric activation of the PSC via right hip abduction-external rotation, and post-stretching of the caudal segment of the PSC (right or left side, alternating allocation procedure). Left and right trunk rotation, total rotational range of motion (ROM), and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
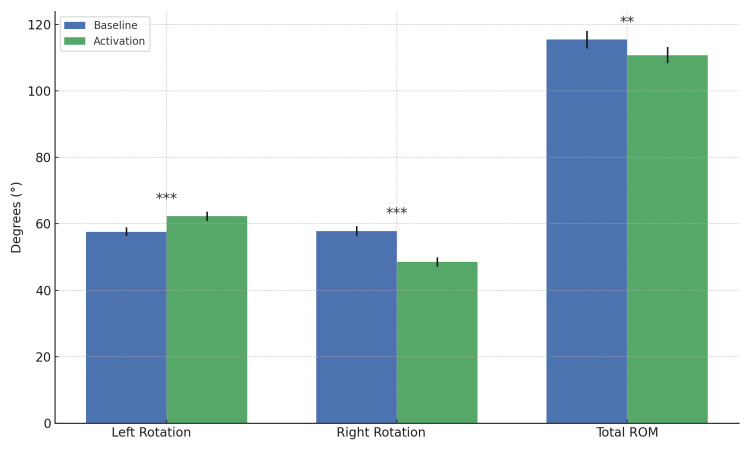 Figure 1
Figure 1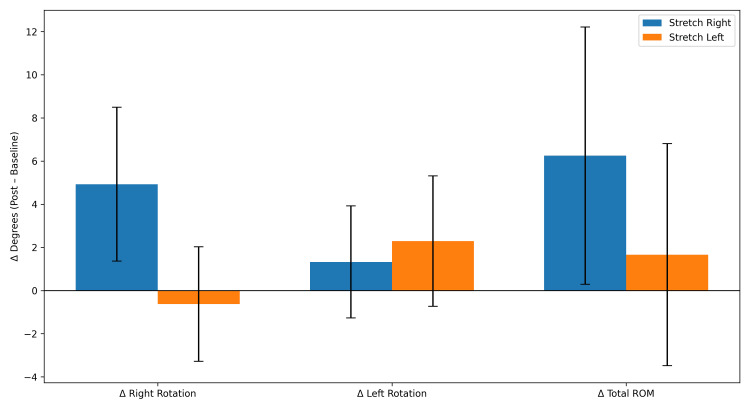 Figure 2
Figure 2| Parameter | Left Rotation (°) | Right Rotation (°) | Mean Difference (Left / Right) | t-value | p-value | Hedges’ g | 95% CI (Difference) |
| Baseline | 57.60 ± 11.02 | 57.80 ± 12.05 | –0.20° | –0.32 | 0.749 | –0.04 | –2.27 to +1.64 |
| Rotation | Female (n = 26) | Male (n = 47) | t-value | p-value | Hedges’ g | 95% CI of Difference (Male – Female) |
| Left Rotation (°) | 57.03 ± 11.31 | 57.91 ± 10.97 | 0.321 | 0.750 | 0.08 | –4.62 to 6.37 |
| Right Rotation (°) | 56.57 ± 12.56 | 58.48 ± 11.85 | 0.635 | 0.529 | 0.16 | –4.14 to 7.96 |
| Rotation Parameter | Right-Dominant (n = 67) | Left-Dominant (n = 6) | t-value | p-value | Hedges’ g | 95% CI (Right – Left) |
| Left Rotation (°) | 56.88 ± 10.41 | 65.54 ± 15.41 | –1.35 | 0.231 | –0.79 | –24.78 to +7.47 |
| Right Rotation (°) | 56.90 ± 11.09 | 67.84 ± 18.37 | –1.44 | 0.207 | –0.92 | –30.17 to +8.29 |
| Rotation Parameter | Left Rotation (°) | Right Rotation (°) | t-value | p-value | Hedges’ g | 95% CI (Left – Right) |
| Activation | 62.26 ± 12.06 | 48.50 ± 12.00 | 9.95 | 3.7 × 10⁻¹⁵ | 1.15 | +11.00 to +16.52 |
| Rotation Direction | Baseline (°), Mean ± SD | Activation (°), Mean ± SD | Mean Change (°) | t-value | p-value | Hedges’ g | 95% CI of Change |
| Left Rotation | 57.60 ± | 62.26 ± | +4.67° | –4.77 | 9.4 × 10⁻⁶ | 0.55 | +2.71 to +6.62 |
| Right Rotation | 57.80 ± | 48.50 ± | –9.30° | +7.92 | 2.2 × 10⁻¹¹ | –0.92 | –11.64 to –6.96 |
| Parameter | Baseline ROM (°) Mean ± SD | Activation ROM (°) Mean ± SD | Mean Change (°) | t-value | p-value | Hedges’ g | 95% CI of Change |
| Total ROM (Left + Right) | 115.40 ± 22.34 | 110.76 ± 20.96 | –4.63° | 2.79 | 0.0068 | –0.32 | –7.95 to –1.32 |
| Parameter (Δ Post–Baseline) | Stretch Right (n = 39) Mean ± SD | Stretch Left (n = 34) Mean ± SD | t-value | p-value | Hedges’ g | 95% CI |
| Δ Right rotation | +4.93 ± 10.84 | +2.29 ± 9.67 | 0.56 | 0.575 | 0.13 | −6.70 to +11.99 |
| Δ Left rotation | +1.33 ± 9.95 | +4.82 ± 10.21 | −1.54 | 0.128 | −0.36 | −9.43 to +1.35 |
| Δ Total ROM (Left / Right) | +6.26 ± 15.41 | +7.11 ± 14.88 | −0.24 | 0.809 | −0.06 | −8.34 to +6.64 |
| Δ Rotational Shift Left / Right) | −3.60 ± 9.12 | +2.92 ± 8.47 | −2.14 | 0.036 | −0.72 | −12.59 to −0.44 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMyofascial pain diagnosis and treatment · Body Contouring and Surgery · Musculoskeletal pain and rehabilitation
Introduction
The fascial system is currently regarded as a continuous anatomical network capable of connecting even distant body regions and contributing to the transmission and modulation of muscular forces. Experimental studies have shown that a substantial proportion of the force generated by muscle contraction is not transmitted exclusively through the myotendinous junction but also via intra-, inter-, and extramuscular myofascial pathways, potentially affecting structures that are not directly adjacent to the activated muscle [1-4].
Fascial tissue exhibits sufficient stiffness to transmit approximately 15% to 37% of muscular force to adjacent structures rather than conveying it entirely to the tendinous insertion [5]. The magnitude of this force transmission depends on the length and relative spatial arrangement of the involved tissues [6,7]. Moreover, these forces may be distributed over a distance, thereby influencing the range of motion of segments remote from the initial site of muscular contraction [8]. This long-distance phenomenon has been defined by Wilke et al. [9] as the “remote effect,” referring to biomechanical or functional changes observed in body regions other than the one directly subjected to treatment. Accordingly, a related hypothesis is that myofascial connectivity contributes to the effects of “remote stretching” through force transmission along myofascial chains. Given the rich innervation of these tissues, the term "neuro-myofascial systems" may be more appropriate.
A systematic review by Wilke et al. [9] provided strong evidence, primarily based on anatomical dissection studies, for the existence of at least three myofascial chains proposed by Myers [10]: the superficial back line, the back functional line, and the front functional line. In contrast, the evidence supporting the spiral line and the lateral line was rated as moderate to strong but less consistent.
Studies investigating remote effects have predominantly focused on myofascial systems with a mainly sagittal orientation [9,11-17], particularly the superficial back line. When examining the myofascial connectivity models proposed in the literature, all authors [10,18-20] agree on the existence of a posterior sagittal antigravitational system connecting the cranial and podalic regions bilaterally. However, not all models describe a system linking the right cranial region with the contralateral (left) podalic region. Among the systems involved in rotational body mechanics, Busquet [18] refers to these connections as crossed, Myers [10] defines them as functional, while Stecco [19] and Colonna [21] describe them as spiral, emphasizing their three-dimensional spatial organization.
In contrast to the interpretations proposed by Stecco [19] and Colonna [21], the spiral line described by Myers [10] appears to be more difficult to interpret biomechanically, as it is conceptualized as a concatenation of an anterior spiral component that continues into a posterior sagittal component.
Within the framework of posterior spiral chains, the thoracolumbar fascia (TLF) represents a crucial biomechanical hub. Its anatomical connections with the latissimus dorsi (LD) and the gluteus maximus (GMax) create a force-transmission pathway that crosses the lumbosacral region and functionally links the shoulder girdle to the contralateral lower limb [22]. Cadaveric anatomical studies have shown that tension applied to either the LD or the GMax produces measurable displacements within the TLF, including on the contralateral side, supporting the existence of a true mechanical continuity along this crossed posterior pathway [23,24].
Within this context, the concept of the posterior spiral chain (PSC) provides a three-dimensional functional interpretation of this system: a myofascial pathway connecting the upper limb through the latissimus dorsi, the thoracolumbar fascia, and the contralateral lower limb via the gluteal region [25]. This model integrates features of the crossed chains described by Busquet [18] and the “functional lines” reported in the literature [10].
The PSC is particularly involved in trunk rotation tasks and in movements requiring crossed pelvic stabilization, in which one myofascial spiral acts as the functional agonist of rotation while the contralateral spiral must effectively lengthen to serve as a functional antagonist. For example, during left trunk rotation, active engagement of the right anterior spiral and the left posterior spiral must be accompanied by adequate relaxation of the opposing chains, namely, the left anterior spiral and the right posterior spiral, to avoid mechanical restriction of movement.
In vivo evidence has begun to support this model. Carvalhais et al. [23] demonstrated that increasing tension in the latissimus dorsi can modify the passive mechanical properties of the contralateral hip, providing early experimental support for myofascial force transmission along what has been described as the “back functional line” [1,10,26], the “posterior oblique sling” [22], or the posterior spiral chain [19,21,27]. More recent studies in runners have shown that latissimus dorsi contraction is associated with changes in lumbar stiffness and passive characteristics of the contralateral hip, further reinforcing the hypothesis of a functional LD-TLF-GMax coupling [23,25].
Despite the growing body of anatomical and experimental evidence on myofascial force transmission, the in vivo functional relevance of this crossed posterior chain remains insufficiently clarified. In particular, it is still unclear whether, and to what extent, modulation of tension at the caudal end of the PSC, for instance through isometric contraction or stretching of the hip abductors and external rotators, can alter trunk rotational range of motion in the transverse plane without changing the overall movement amplitude, but rather by redistributing its directional expression [1].
The aim of the present study was to test this hypothesis by investigating whether an increase in active tension (via isometric contraction) and/or a reduction in passive tension (via muscle energy stretching techniques) at the caudal segment of the PSC induces measurable changes in trunk rotational range of motion in the transverse plane. Demonstrating such changes in the absence of modifications in total rotational amplitude would provide further in vivo evidence of myofascial force transmission along the PSC and support its functional role in the regulation of trunk rotation and postural spirals [23,25].
Materials and methods
Study design
A single-group pre-post experimental study was conducted to investigate the effects of myofascial tension modulation at the caudal segment of the Posterior Spiral Chain (PSC) on trunk rotational range of motion in the transverse plane. Each participant completed four testing conditions in the following order: Baseline 1; Baseline 2; Isometric activation of the right PSC; Post-stretching of the PSC (right or left side, randomly assigned). All measurements were obtained using inertial digital goniometry.
Participants
Seventy-five healthy participants (48 males, 27 females) were initially recruited-all students from the OSCE school (Osteopathic Spine Center Education); two participants were excluded a posteriori due to acquisition errors related to device calibration. Therefore, data from 73 participants were included in the final analysis.
Participants were aged between 18 and 40 years and had no history of spinal disorders that could limit trunk rotation. Exclusion criteria included the presence of radicular pain or neurological signs, significant spondylolisthesis, spinal stenosis, structural scoliosis, or a history of spinal surgery.
Instrumentation
Trunk rotation was assessed using the Free4Act system (F4A LetSense Srl, Bologna, Italy), a wireless digital goniometer based on inertial sensors. This system has been validated in the literature for the analysis of trunk and pelvic motion [28,29].
Participant positioning and movement standardization
Across all experimental conditions, participants were seated on the edge of an examination table with hips and knees flexed at 90°, feet fully supported on the floor, and a bar positioned across the shoulders and held with the forearms. Pelvic motion was manually controlled by the examiner using a standardized hand placement to minimize compensatory movements. All assessments were performed by the same trained examiner to ensure procedural consistency across participants. Examiner positioning and pelvic stabilization are illustrated in Supplementary Appendix 1.
During the activation condition, isometric activation was consistently performed involving the right lower limb in order to standardize the direction of myofascial tension along the posterior spiral chain across participants.
Stretching was performed using a modified muscle energy technique (MET) targeting the caudal segment of the PSC, as described above. The side of stretching was assigned using a systematic alternating allocation procedure: the first participant underwent stretching on the left side, and subsequent participants were alternately assigned to right and left stretching in sequence. This approach was adopted to ensure a balanced distribution of stretching laterality across the sample.
A detailed description of participant positioning, trunk rotation execution, activation protocol, and stretching procedures is provided in Supplementary Appendix 1.
Statistical analysis
Normality of the data was assessed using the Shapiro-Wilk test. Since all primary outcomes were normally distributed, parametric statistics were applied. Paired t-tests were used to compare within-subject conditions (baseline vs. activation; left vs. right rotation), while independent-samples t-tests (with Welch’s correction when appropriate) were used for between-group comparisons (sex, limb dominance, stretching side).
No formal correction for multiple comparisons was applied. Given the exploratory design of the study and the presence of predefined, hypothesis-driven comparisons, results were interpreted using p-values in conjunction with effect sizes (Hedges’ g) and 95% confidence intervals.
Statistical significance was set at p < 0.05 (two-tailed), and effect sizes with 95% confidence intervals were used to support interpretation.
Standardized effect sizes were estimated using Hedges’ g rather than Cohen’s d, as Hedges’ g provides a bias correction for small sample sizes and unequal group distributions [30,31].
In the absence of robust preliminary data allowing reliable a priori sample size estimation, the study was designed using an adaptive approach [32]. An exploratory analysis of the first 30 participants revealed medium-to-large effects (Hedges’ g ≈ 0.7-0.8) for the primary outcomes related to trunk rotation changes during muscle activation. Based on these estimates, a paired t-test with α = 0.05 and 90% power would have required approximately 30 participants. To improve the robustness of subgroup analyses, recruitment was therefore continued up to a total sample of 75 participants.
Results
Data from 73 participants (48 males and 27 females) were analyzed. The mean age was 24.03 ± 3.95 years, with a mean body weight of 69.06 ± 12.91 kg and a mean height of 171.87 ± 7.31 cm.
Shapiro-Wilk testing confirmed normal distribution of baseline trunk rotation values, supporting the use of parametric statistical analyses.
Baseline trunk rotation showed no lateral asymmetry (Table 1), confirming the absence of a baseline rotational bias.
No sex-related differences were observed in baseline trunk rotation (Table 2). Accordingly, the sample was considered homogeneous with respect to sex-related baseline trunk rotational mobility.
Baseline trunk rotation did not differ significantly between right- and left-dominant individuals (Table 3). Left-dominant participants tended to show higher rotation values in both directions; however, these differences did not reach statistical significance. The small number of left-dominant participants (n = 6) limited the statistical power of this analysis.
Muscle activation induced a marked redistribution of trunk rotational mobility (Tables 4-6). Compared with baseline, activation was associated with a significant increase in left rotation and a concomitant reduction in right rotation, resulting in a pronounced leftward shift in rotational capacity (Figure 1). Total trunk rotational ROM showed only a small change, indicating that activation primarily altered the directional distribution of rotation rather than producing a substantial net gain or loss of overall mobility.
*Baseline vs. activation trunk rotation (left, right, total ROM).Bar chart showing left rotation, right rotation, and total trunk rotation range of motion (ROM) at baseline and during activation. Bars represent mean values with standard error of the mean (SEM). Asterisks indicate statistical significance of paired comparisons: ***p < 0.001, **p < 0.01, p < 0.05, and ns = not significant.
Following stretching, trunk rotational mobility showed a partial rebalancing of the activation-induced asymmetry (Table 7, Figure 2). When changes from baseline were compared between participants stretched on the right versus the left lower limb, no significant between-group differences were observed for left rotation, right rotation, or total ROM. However, a significant difference emerged for the rotational shift (left/right), indicating a direction-specific effect of stretching on rotational balance.
Side-specific effects of stretching on trunk rotation (Δ post–baseline).Bar chart showing changes from baseline (Δ post–baseline) in right rotation, left rotation, and total ROM after stretching. Participants are grouped by stretched side (right vs. left). Bars represent mean ± SEM. No significant differences between-group were observed.
Discussion
The findings of the present study contribute to the growing body of research on myofascial force transmission, which has demonstrated that forces generated within a given body region can influence the mechanical behavior of non-contiguous segments through fascial and epimuscular continuities [1,33]. Both cadaveric and in vivo studies have documented load transmission between the latissimus dorsi, the thoracolumbar fascia, and the lower limb [15,22-24]. The present work extends these observations by providing in vivo evidence obtained during a functional task, namely, trunk rotation.
Specifically, this study provides in vivo evidence supporting the existence of a functional, beyond purely anatomical, relationship between adjacent segments belonging to the posterior spiral chain (PSC). The results demonstrate that modulation of myofascial tension at the caudal level is capable of systematically altering the pattern of trunk rotation in the transverse plane.
Effect of activation: directional shift without an increase in total ROM
The most robust finding of the present study concerns the isometric activation phase of the PSC. Under this condition, a significant reduction in rightward rotation was observed, accompanied by a concomitant increase in leftward rotation, without an expansion of the overall rotational cone. This pattern suggests that muscle activation does not generically increase joint mobility but rather induces a directional redistribution of the available rotational range of motion.
From a biomechanical perspective, this behavior is consistent with the spiral myofascial chain model described by Colonna [21], according to which, during rotational movements, one chain acts as the functional agonist while the contralateral chain must effectively lengthen to permit motion. Increased active tension at one extremity of the PSC, therefore, appears to facilitate contralateral rotation while constraining ipsilateral rotation, resulting in a shift of the rotational cone rather than its expansion. Consistent with this interpretation, total trunk rotational ROM was found to be slightly, but significantly, reduced during the activation phase.
The response observed during the stretching phase further supports this conceptual framework and aligns with the work of Stecco [19], who emphasized the role of fascia as a structure capable of modulating intermuscular coordination and force distribution, rather than acting as a purely passive element. The present findings are also consistent with experimental evidence reported by Carvalhais et al. [23], who demonstrated force transmission between the latissimus dorsi and the contralateral gluteus maximus, as well as with the reviews by Wilke et al. [1], which highlighted the capacity of myofascial chains to influence global kinematics even in the absence of direct muscular continuity.
By extending these observations to the axial skeleton, the present study shows that such myofascial effects are detectable at the level of the spine during a functional task such as trunk rotation.
Effect of stretching: direction-specific response and individual variability
The stretching phase produced more complex and less uniform results compared with the activation phase. Stretching of the left lower limb was associated with a significant increase in leftward trunk rotation, whereas stretching of the right lower limb did not elicit comparable statistically significant changes in contralateral rotation. However, when the rotational balance shift (left/right) was analyzed, a relevant finding emerged: stretching induced a displacement of the rotational cone toward the treated side, independently of changes in total rotational ROM.
These findings suggest that the primary effect of stretching is not an increase in absolute joint excursion but rather a modification of the tensile balance between the two hemichains, consistent with a mechanism of selective reduction in passive tension along the PSC. The substantial inter-individual variability observed is in agreement with previous literature on fascial tissue responses to mechanical stimuli, which are known to depend on factors such as tissue stiffness, collagen organization, and individual motor strategies, as described by Schleip et al. [34].
An additional relevant aspect is that the right lower limb was consistently involved during the activation phase preceding stretching. It is therefore plausible that a persistence of activation-related effects attenuated the response to stretching on the same side, suggesting a potential interaction between active and passive tensioning mechanisms that warrants further experimental investigation.
Lateral dominance and subgroup analyses
Subgroup analyses did not reveal significant differences in baseline rotational values between males and females, indicating a largely sex-homogeneous sample. With respect to limb dominance, left-dominant participants exhibited higher mean rotational values; however, the extremely small number of left-dominant individuals substantially limited the statistical power of this comparison. In this context, the use of Hedges’ g allowed estimation of effect magnitude while reducing bias associated with small and unbalanced sample sizes, in accordance with methodological recommendations for exploratory studies.
Clinical implications
The structural complexity and three-dimensional continuity of the fascial system render this tissue particularly vulnerable to repetitive mechanical overload and maladaptive responses, which may contribute to the development of chronic musculoskeletal disorders and persistent pain syndromes. In recent years, fascia has no longer been regarded as a purely passive support structure but rather as a mechanosensitive and reactive tissue, characterized by rich innervation and the presence of contractile cellular elements, particularly myofibroblasts, capable of modifying tissue mechanical properties in response to mechanical stimuli [35].
These characteristics help explain the involvement of the myofascial system in a wide range of clinical disorders, including chronic neck pain, low back pain, adhesive capsulitis of the shoulder, and nerve entrapment syndromes. In such disorders, pain is often present not only in association with stiffness or hyperlaxity but also at end ranges of motion, suggesting an active role of fascia in modulating nociceptive perception and movement control [36-38].
The results of the present study fit within this framework, demonstrating that remote modulation of myofascial tension can produce measurable changes in trunk kinematics without the need for direct intervention at the spinal segment. In particular, the observation that both activation and stretching primarily induced a directional shift of the rotational cone, rather than an increase in total ROM, suggests that clinical interventions targeting the myofascial system may be more effective in rebalancing altered movement patterns than in pursuing a nonspecific increase in joint mobility.
From an applied perspective, these findings support a global approach to assessment and treatment, in which targeted activation or stretching strategies along specific myofascial chains may be used to influence the functional behavior of the spine and hip. This approach appears particularly relevant for injury prevention and rehabilitation, as well as within sports medicine, where the myofascial system is increasingly recognized as a key contributor to performance and mechanical load management [34].
In light of recent advances, both clinical and experimental research are increasingly focused on the dynamic adaptation and the structural and biochemical changes of fascia in response to mechanical loading. Progress in this field will require a closer integration of biomechanics, therapeutic exercise, and functional assessment tools, with the aim of developing more targeted, effective, and individualized intervention strategies.
Clinically, the present findings suggest that interventions applied to caudal segments of the PSC, through either activation or stretching, can induce measurable changes in trunk kinematics even in the absence of direct treatment of the spine. This supports a global approach to the evaluation and management of trunk rotational dysfunctions, consistent with biomechanical models attributing a central role to the thoracolumbar fascia in load transfer between the lower limbs and the spine, as described in the work of Vleeming et al. [22] and Barker [24].
The fact that both activation and stretching predominantly resulted in a redistribution of the rotational cone, rather than an increase in total ROM, further suggests that myofascial treatments may be particularly useful for restoring directional balance in movement patterns, rather than simply increasing joint mobility. This perspective may have important implications for the management of low back pain, hip dysfunctions, and disorders characterized by persistent rotational asymmetries.
Strengths and limitations
Maintaining a final sample size of 73 participants enhanced the overall robustness of the analyses and allowed more reliable exploration of several subgroup comparisons, including sex-related differences (males vs. females), lower limb dominance (right vs. left), and, in a subsequent phase, the side of stretching (right vs. left). A larger sample also helped reduce the risk of type II error, improving the stability of variability estimates and the precision of confidence intervals. Overall, the final sample size strengthens the methodological validity of the findings.
The main limitations of this study include the absence of a control group, inter-individual variability in the intensity of isometric activation, the operator-dependent nature of the stretching protocol, and the unbalanced distribution of left-dominant participants. Furthermore, it was not possible to definitively identify the specific anatomical pathway of force transmission, although the thoracolumbar fascia represents the most plausible biomechanical candidate, as suggested by anatomical and histological evidence [22].
Future studies should combine controlled experimental designs with direct measurements of fascial stiffness and neuromuscular activation to further clarify the direction-specific role of the posterior spiral chain and its clinical relevance.
Conclusions
This study provides in vivo evidence supporting the functional relevance of the posterior spiral chain as a myofascial pathway capable of modulating trunk rotational behavior. Modulation of myofascial tension at the caudal level, through either isometric activation or targeted stretching, did not primarily alter the overall amplitude of trunk rotation but rather induced a direction-specific redistribution of the available range of motion. These findings suggest that myofascial interventions act mainly by shifting the rotational cone, influencing movement balance rather than increasing total mobility.
The observed effects during activation and stretching are consistent with current biomechanical models of spiral myofascial connectivity and support the concept that remote segments can influence axial kinematics through fascial continuity. From a clinical perspective, this highlights the potential value of addressing myofascial chains when assessing and managing trunk rotational dysfunctions, particularly in disorders characterized by asymmetrical movement patterns. Future research employing controlled designs and direct measurements of fascial mechanical properties is warranted to further clarify the mechanisms underlying direction-specific myofascial force transmission.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1What is evidence-based about myofascial chains: a systematic review Arch Phys Med Rehabil Wilke J Krause F Vogt L Banzer W 4544619720162628195310.1016/j.apmr.2015.07.023 · doi ↗ · pubmed ↗
- 2Myofascial continuity: Review of anatomical and functional evidence J Bodyw Mov Ther Kalichman L 5695754520254131662210.1016/j.jbmt.2025.09.020 · doi ↗ · pubmed ↗
- 3Intermuscular force transmission between human plantarflexor muscles in vivo J Appl Physiol (1985) Bojsen-Møller J Schwartz S Kalliokoski KK 1608161810920102088483810.1152/japplphysiol.01381.2009 · doi ↗ · pubmed ↗
- 4Extramuscular myofascial force transmission also occurs between synergistic muscles and antagonistic muscles J Electromyogr Kinesiol Huijing PA van de Langenberg RW Meesters JJ Baan GC 6806891720071738389810.1016/j.jelekin.2007.02.005 · doi ↗ · pubmed ↗
- 5Significance of epimuscular myofascial force transmission under passive muscle conditions J Appl Physiol (1985) Maas H 1465147312620193060539810.1152/japplphysiol.00631.2018 · doi ↗ · pubmed ↗
- 6Spastic muscle properties are affected by length changes of adjacent structures Muscle Nerve Smeulders MJ Kreulen M Hage JJ 2082153220051593787510.1002/mus.20360 · doi ↗ · pubmed ↗
- 7Myofascial force transmission and tendon transfer for patients suffering from spastic paresis: a review and some new observations J Electromyogr Kinesiol Smeulders MJ Kreulen M 6446561720071736905210.1016/j.jelekin.2007.02.002 · doi ↗ · pubmed ↗
- 8A review of the theoretical fascial models: biotensegrity, fascintegrity, and myofascial chains Cureus Bordoni B Myers T 012202010.7759/cureus.7092 PMC 709601632226693 · doi ↗ · pubmed ↗