RNA-binding protein Rbm38 as a multifaceted post-transcriptional regulator in zebrafish pancreatic development
Xiangmin Zhang, Xianpeng Li, Rui Liu, Lu Wang, Yunchao Wang, Ailong Zhang, Shuaiqi Yang, Hongyan Li

TL;DR
This study explores how the RNA-binding protein Rbm38 regulates pancreatic development in zebrafish and affects gene expression.
Contribution
The study reveals Rbm38's multifaceted role in post-transcriptional regulation during zebrafish pancreatic development.
Findings
Rbm38 is expressed in the zebrafish pancreas during larval stages and is regulated by specific transcription factors.
Loss of Rbm38 causes abnormal pancreatic enlargement and disrupts post-transcriptional regulation of key genes.
Rbm38 destabilizes pdx1 transcripts and regulates alternative splicing of isl1a, smad2, and nkx2.2a.
Abstract
RNA-binding motif protein 38 (Rbm38), also known as RNPC1, is a major regulator of post-transcriptional gene expression. It represents a potential candidate gene linked to the susceptibility of type 2 diabetes, and decreased RBM38 expression can enhance the proliferation of pancreatic cancer cells in humans. However, its role in pancreatic development remains elusive. In this study, we explored the function of Rbm38 using zebrafish as a model. Pancreatic expression of Rbm38 is present at larval stages and is controlled by several transcription factors acting on specific rbm38 promoter regions. The loss of Rbm38 leads to abnormal pancreatic enlargement. Mechanistically, Rbm38 is involved in several aspects of post-transcriptional regulation of pancreatic gene expression. It destabilizes pdx1 transcripts by binding to the 3′-untranslated region and regulates alternative splicing of key…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
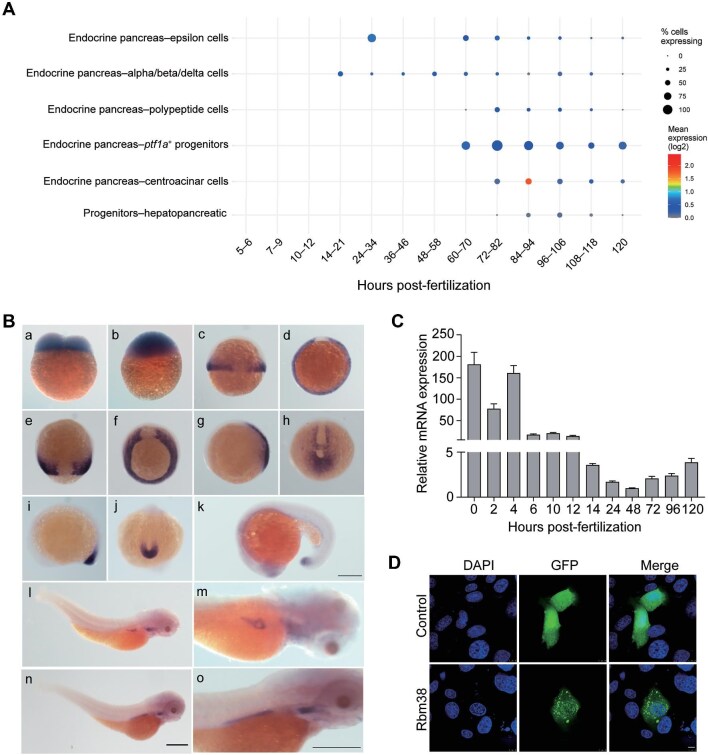 Figure 1
Figure 1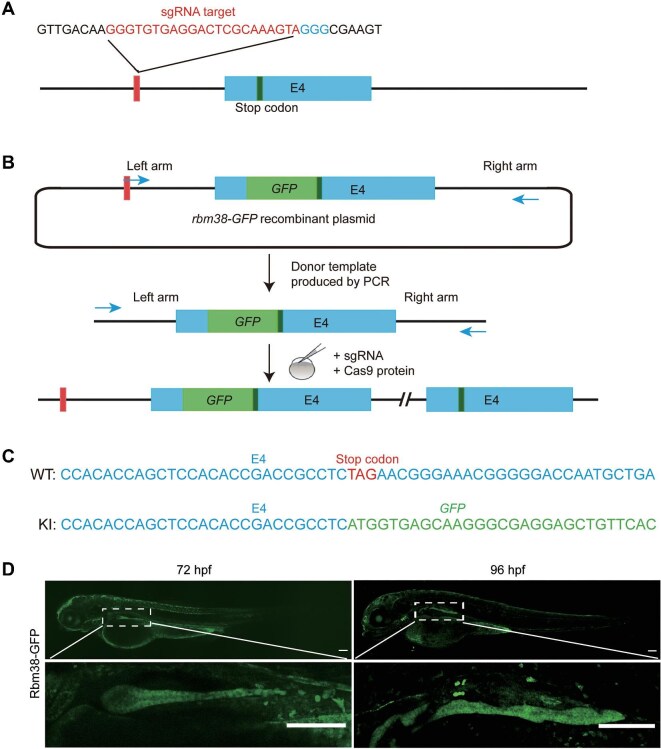 Figure 2
Figure 2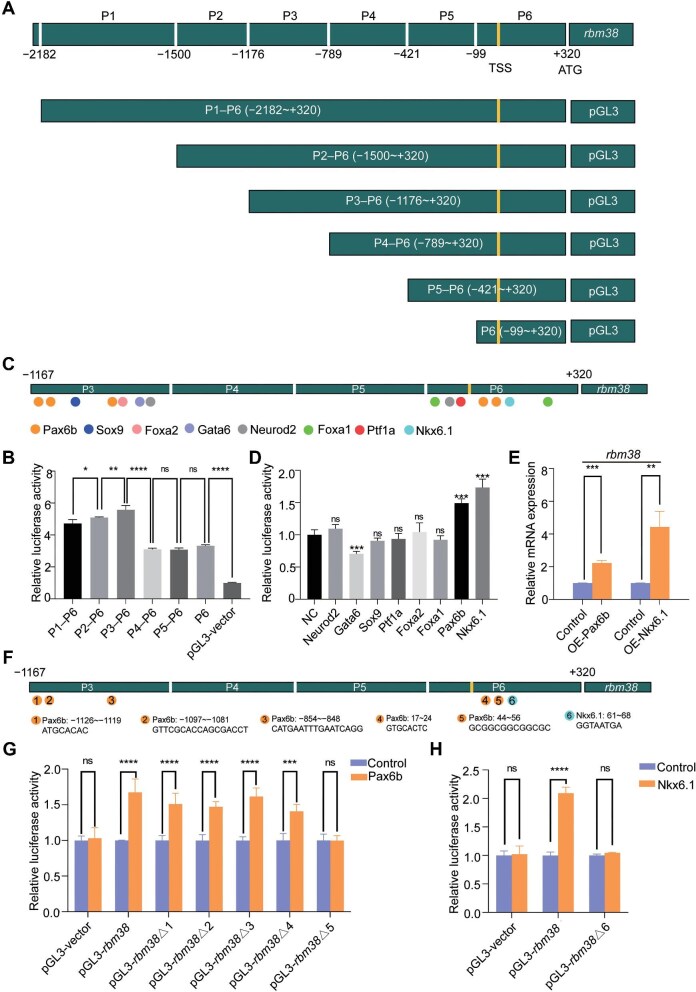 Figure 3
Figure 3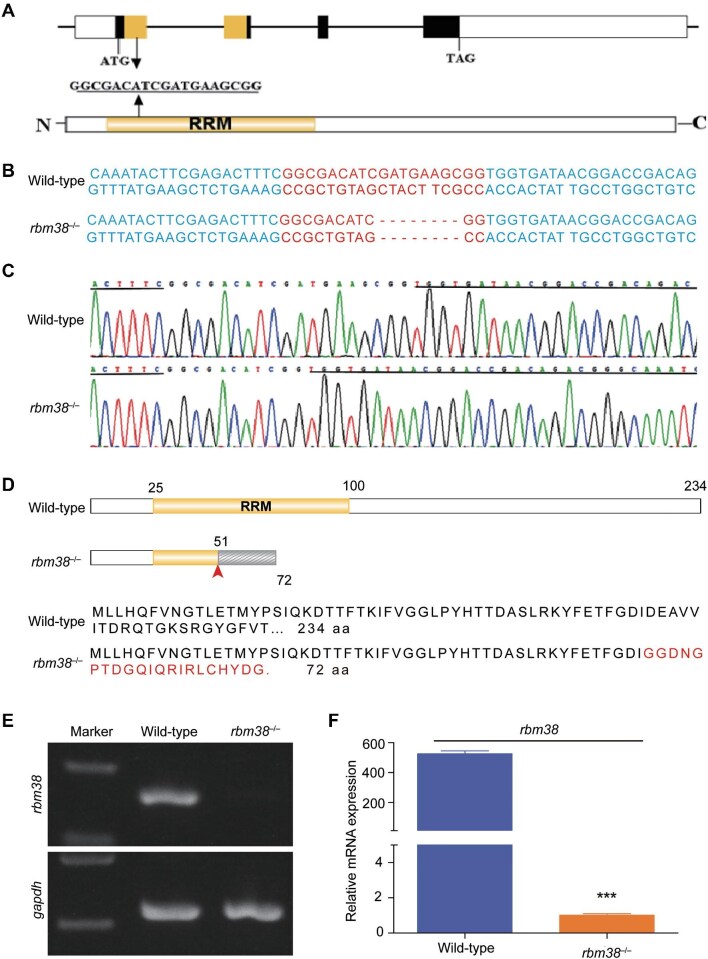 Figure 4
Figure 4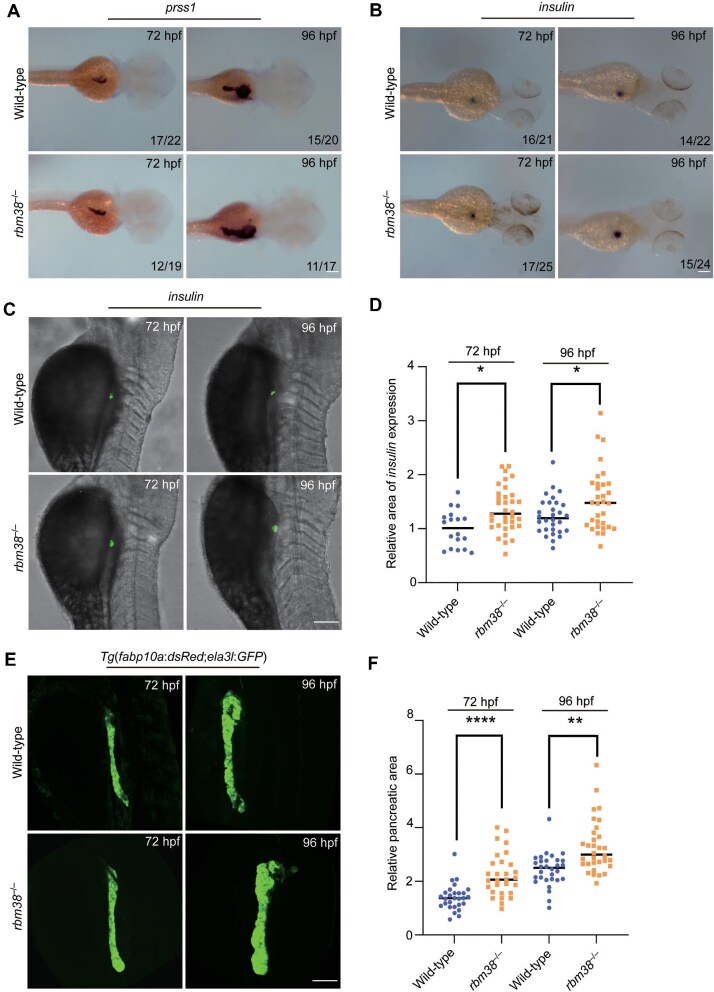 Figure 5
Figure 5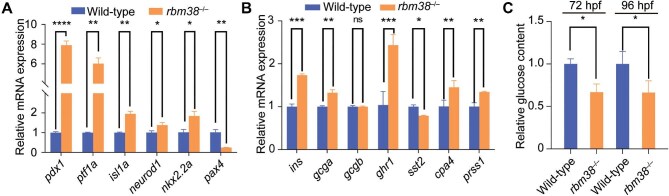 Figure 6
Figure 6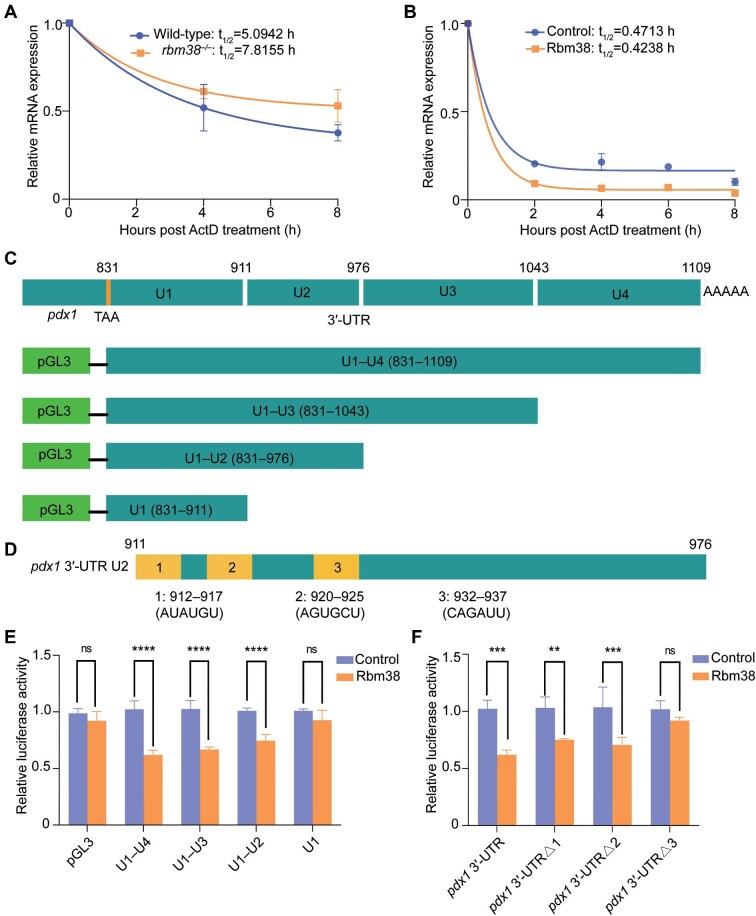 Figure 7
Figure 7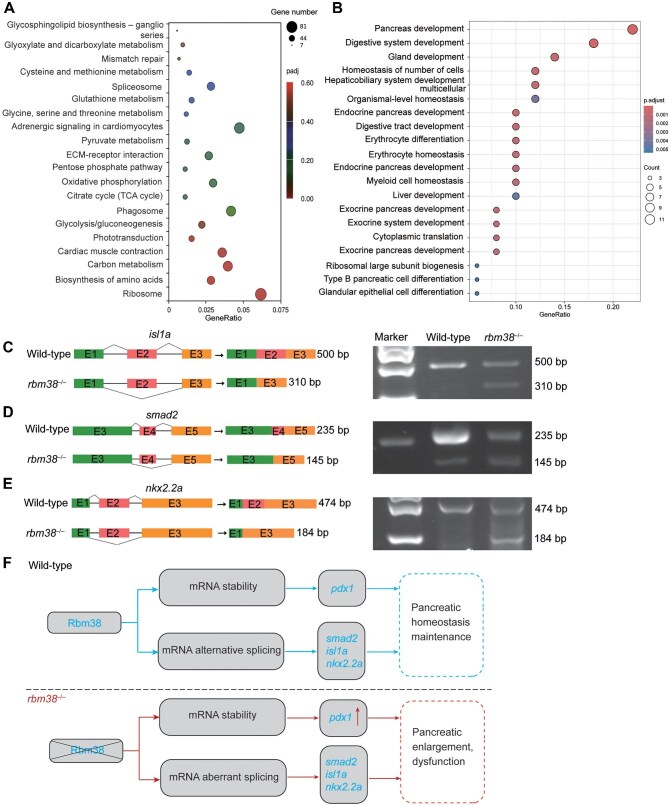 Figure 8
Figure 8- —Science and Technology Innovation Project of Longhua Hospital10.13039/501100013090
- —National Natural Science Foundation of China10.13039/501100001809
- —Qingdao Postdoctoral Science Foundation10.13039/100018936
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · RNA Research and Splicing · MicroRNA in disease regulation
Introduction
The development and maintenance of pancreatic homeostasis are governed by a complex interplay of regulatory mechanisms, encompassing both transcriptional and post-transcriptional controls (Apelqvist et al., 1999; Hebrok, 2003; Fujitani et al., 2006; Glauser and Schlegel, 2009; Leung, 2010; Jennings et al., 2015; Mi et al., 2024). Although previous works mostly centered on transcriptional regulation, there is a growing body of evidence indicating that post-transcriptional regulation is equally vital for proper pancreatic development. RNA-binding proteins (RBPs) are key players of post-transcriptional regulation, harboring one or more RNA recognition motifs (RRMs) that allow them to regulate RNA metabolism at multiple levels, from pre-RNA splicing to mRNA localization, polyadenylation, stability, translation, and nuclear export (Li et al., 2010). These multifaceted RBPs are integral to a diverse array of biological processes, highlighting their importance in the intricate regulation of pancreatic function (Blencowe, 2006; Grifone et al., 2020).
Several RBPs have been identified as crucial players in pancreatic development and homeostasis. Recent studies using various animal models have elucidated multiple post-transcriptional regulatory mechanisms by which they function within the pancreas. For instance, PTBP binds to the pyrimidine-rich region in the 3′-untranslated region (3′-UTR) of rat insulin mRNA, promoting its stability and translation (Tillmar et al., 2002). The translocation of PTBP1 between the nucleus and cytoplasm is also linked to the rapid post-transcriptional regulation of insulin levels in pancreatic islets, which are involved in the response to glucose levels. This regulatory process may be disrupted in type 2 diabetes, leading to impaired insulin secretion (Ehehalt et al., 2010). Furthermore, RBPs are involved in translation control, which is another essential aspect of pancreatic homeostasis. RBMS3, for example, facilitates the translation of Ptf1a mRNA by interacting with its 3′-UTR (Lu et al., 2012). HuD serves as a key regulator of insulin mRNA translation in pancreatic β-cells by binding to the 5′-UTR of pre-proinsulin (ins2) mRNA, inhibiting its translation, and consequently decreasing insulin production. Alternative splicing represents another layer of gene regulation, contributing to pancreatic development. Rbm4 modulates pancreatic cell differentiation and insulin secretion by coordinating the alternative splicing of key genes encoding pancreatic developmental transcription factors, such as Isl1 and Pax4 (Lin et al., 2013). NOVA1 plays an important role in the functionality of pancreatic β-cells, influencing insulin secretion and apoptosis by regulating alternative splicing events (Villate et al., 2014). Despite the progress made in understanding the post-transcriptional regulation of pancreatic development, there remains an urgent need for identifying additional RBPs that participate in this process in order to gain further insight into the molecular underpinnings of pancreatic function and disease.
Rbm38 contains a single RRM domain and is involved in various post-transcriptional regulatory processes. It binds to AU-rich or GU-rich regions in the 3′-UTR of target mRNAs, regulating mRNA metabolism through multiple mechanisms, including stabilization, destabilization, and/or translation (Shu et al., 2006; Zhang et al., 2010; Cho et al., 2012; Yan et al., 2012; Xu et al., 2013; Ye et al., 2018; She et al., 2020). For instance, Rbm38 stabilizes p53, p21, and p73 transcripts, thereby promoting their translation (Shu et al., 2006; Zhang et al., 2011; Yan et al., 2012). Conversely, it promotes the degradation of mRNAs encoding oncogenic proteins such as MYC and MDM2 (Xu et al., 2013; Li et al., 2017). Additionally, Rbm38 inhibits the translation of p53 mRNA by preventing eIF4E from binding to its 5′ cap region (Zhang et al., 2011). Furthermore, Rbm38 acts as a tissue-specific splicing factor, regulating the splicing of numerous genes involved in erythroid differentiation, antiviral immunity, neural differentiation, and muscle development (Anyanful et al., 2004; Heinicke et al., 2013; Du et al., 2020; Li et al., 2020). Previous studies suggested that Rbm38 is a candidate gene for the susceptibility to type 2 diabetes, and the decreased expression of Rbm38 may promote the proliferation of pancreatic cancer cells (Saxena et al., 2013; Gong et al., 2016; Lu et al., 2022). However, no causal relationship between Rbm38 and pancreatic development has been established to date.
Here, we uncover a novel function of Rbm38 in zebrafish pancreatic development. Rbm38 displays prominent expression in the pancreas at larval stages. The deficiency of Rbm38 leads to abnormal enlargement of the pancreas during early development. Mechanistically, Rbm38 regulates pancreatic development by destabilizing pdx1 transcripts and controlling the alternative splicing of key genes encoding pancreatic transcription factors, including isl1a, smad2, and nkx2.2a. These data demonstrate a critical role of Rbm38 in zebrafish pancreatic development by post-transcriptionally controlling the expression of pancreatic genes.
Results
Zebrafish Rbm38 shows restricted expression in the developing pancreas
Gene function is often closely related to its expression pattern. The expression profile of rbm38 in the online single-cell transcriptome database (Sur et al., 2023) indicates that it is expressed in endocrine and exocrine cells as well as hepato–pancreatic progenitor cells, with significantly higher expression levels in exocrine cells than in endocrine cells (Figure 1A).
Expression of rbm38 during zebrafish early development. (A) Expression pattern of zebrafish rbm38 in endoderm cell lineages. The analysis was performed using online single-cell transcriptome database (https://daniocell.nichd.nih.gov/index.html). The size of each dot reflects the relative mean expression level of the corresponding gene. (B) WISH analysis of zebrafish rbm38 expression at different developmental stages, including 2-cell stage (a), 4 hpf (b), 6 hpf (c, lateral view; d, animal pole view), 9 hpf (e, dorsal view; f, vegetal pole view), 10 hpf (g, lateral view; h, posterior view), 12 hpf (i, lateral view; j, posterior view), 18 hpf (k), 72 hpf (l; m, higher magnification), and 96 hpf (n; o, higher magnification). Scale bar, 300 μm. (C) RT-qPCR analysis of rbm38 temporal expression levels during early development. Data are normalized to gapdh and presented as mean ± SD. (D) Subcellular localization of Rbm38-GFP in SW1990 cells. Nuclei are counterstained by DAPI. Scale bar, 10 μm.
To analyze the expression pattern of rbm38 during early stages of zebrafish development, we conducted whole-mount in situ hybridization (WISH) using an antisense RNA probe (Figure 1B). The expression of rbm38 was observed from the 2-cell stage to 96 h post-fertilization (hpf). Prior to the onset of zygotic gene expression, rbm38 displayed a robust ubiquitous expression pattern (Figure 1B, a and b), indicating that it is a maternal gene. From 6 hpf to 9 hpf, rbm38 was specifically expressed in the mesoderm except for the dorsal region representing the precursor of the notochord (Figure 1B, c–f). From 10 hpf to 24 hpf, rbm38 was specifically expressed in the tail bud, eyes, and unsegmented presomitic mesoderm (Figure 1B, g–k). Between 72 hpf and 96 hpf, rbm38 was primarily detected in the pharyngeal arches and pancreas (Figure 1B, l–o). Quantitative real-time PCR (RT-qPCR) analysis revealed that the temporal expression of rbm38 exhibited a biphasic pattern, with high levels at initial stages of development, progressively decreasing to the lowest level at 48 hpf, and then rising again (Figure 1C).
We next examined the subcellular localization of Rbm38 by expressing GFP or Rbm38-GFP fusion protein in SW1990 cells. The results revealed that GFP was expressed uniformly in the cell, while Rbm38-GFP was widely distributed in both the cytoplasm and the nucleus, displaying a granular pattern in the cytoplasm (Figure 1D). This suggests that Rbm38, as an RBP, may exert function through the formation of ribonucleoprotein (RNP) granules.
Zebrafish Rbm38-specific antibody is not available. To examine the expression pattern of endogenous Rbm38 protein, we set out to generate an rbm38-GFP knock-in zebrafish line. A linear donor template comprised of the last exon and partial flanking sequences fused with the GFP coding sequence was co-injected with Cas9 protein and a specific small guide RNA (sgRNA) into 1-cell-stage embryos (Figure 2A–C). In the offspring, the expression pattern of Rbm38 protein was consistent with the data obtained by WISH. Notably, a robust expression was observed in the pancreas at 72 hpf and 96 hpf (Figure 2D). The pancreatic localization of Rbm38-GFP was further confirmed by co-staining using a pancreas-specific antibody for carboxypeptidase A (Supplementary Figure S1).
Generation of zebrafish rbm38-GFP knock-in line by CRISPR/Cas9. (A) Schematic of the intron targeting-mediated strategy for generating GFP knock-in at the zebrafish rbm38 locus. The sgRNA target sequence is shown in red and the protospacer adjacent motif (PAM) sequence in blue. (B) The GFP coding sequence was integrated into the rbm38 locus after co-injection of the donor template with the sgRNA and Cas9 protein. The left arm is 1497 bp, and the right arm is 2120 bp. (C) Genotyping of the rbm38-GFP knock-in line. Blue indicates the rbm38 exon 4 sequence, green shows the GFP sequence, and red marks the rbm38 stop codon. (D) Expression of endogenous Rbm38 at 72 hpf and 96 hpf. Lower panels are higher magnifications of the insets. Scale bar, 100 μm.
To understand the mechanism underlying rbm38 expression in the pancreas, we examined its transcriptional regulation. The full-length rbm38 promoter and its sequential deletions (P1–P6, P2–P6, P3–P6, P4–P6, P5–P6, and P6) were cloned into the pGL3 vector (Figure 3A) and transfected into HEK293T cells. Reporter gene assays indicated that two of the six regions (P3 and P6) are likely responsible for regulating rbm38 gene expression (Figure 3B). We then searched for possible pancreas-related transcription factors that might bind to promoter segments P3 and P6 using online tools and identified eight putative pancreatic developmental transcription factors, including Neurod2, Gata6, Sox9, Ptf1a, Foxa1, Foxa2, Pax6b, and Nkx6.1 (Figure 3C). To examine whether they bind to P3 and P6 regions, their coding regions were individually cloned into the pcDNA3.1 vector and co-transfected with pGL3-(P3–P6) into HEK293T cells (Supplementary Figure S2). The dual-luciferase assay revealed that Pax6b and Nkx6.1 could activate rbm38 expression, whereas Gata6 seemed to have an inhibitory effect (Figure 3D). Consistently, injection of pax6b or nkx6.1 mRNA into 1-cell-stage zebrafish embryos led to a significant upregulation of rbm38 expression (Figure 3E).
*Regulation of rbm38 expression by Pax6b and Nkx6.1. (A) Schematic diagram illustrating reporter constructs with different rbm38 promoter regions (P1–P6). (B) The rbm38 promoter-driven luciferase activity in HEK293T cells. The value from cells transfected with pGL3 vector was set to 1, and data were normalized to Renilla luciferase activity. (C) Mapping of putative pancreatic developmental transcription factor-binding sites within the P3 and P6 regions of the rbm38 promoter. (D) Regulation of rbm38 promoter (P3–P6) activity by different transcription factors in HEK293T cells. The value from cells transfected with pcDNA3.1 empty vector (NC) was set to 1, and data were normalized to Renilla luciferase activity. (E) Quantitative analysis of rbm38 expression in embryos overexpressing (OE) Pax6b and Nkx6.1 at 72 hpf. (F) Mapping of the putative Pax6b- or Nkx6.1-binding sites in the P3 and P6 regions of the rbm38 promoter. (G and H) Binding site 5 (GCGGCGGCGGCGC) and binding site 6 (GGTAATGA) are essential for Pax6b and Nkx6.1 to regulate luciferase activity in HEK293T cells, respectively. Values in pcDNA3.1 empty vector-transfected cells were set to 1 after normalization to Renilla luciferase activity. Data are presented as mean ± SD (one-way ANOVA; *P < 0.05; **P < 0.01; ***P < 0.001; ***P < 0.0001; ns, not significant).
Subsequently, luciferase reporter constructs with deletions of Pax6b- or Nkx6.1-binding sites were generated (pGL3-rbm38Δ1–pGL3-rbm38Δ6) (Figure 3F). The results showed that the transcriptional activity of the rbm38 promoter significantly decreased after the deletion of candidate binding site 5 (GCGGCGGCGGCGC) for Pax6b and candidate binding site 6 (GGTAATGA) for Nkx6.1, indicating that these sites may play a crucial role in activating rbm38 expression (Figure 3G and H).
Generation of rbm38 mutants
To analyze the function of Rbm38 in pancreatic development, we set out to create a zebrafish rbm38 mutant line using CRISPR/Cas9 technology by targeting the first exon, which encodes the RRM domain (Figure 4A). An 8-bp deletion was obtained, and the mutation is expected to cause a premature termination of protein translation, resulting in a truncated peptide with only 72 amino acids (Figure 4B–D). Compared to the wild-type rbm38, the level of mutant rbm38 mRNA was dramatically reduced (Figure 4E and F), likely due to nonsense-mediated RNA decay. No upregulated expression of the homologous genes rbm24a and rbm24b was observed in rbm38 mutants (Supplementary Figure S3). In addition, rbm38 maternal-zygotic mutants could develop into viable adults.
*Generation of zebrafish rbm38 mutant line by CRISPR/Cas9-mediated genome editing. (A) Location of the sgRNA target site in the zebrafish rbm38 locus. (B and C) Genotyping of rbm38 mutation. The sgRNA target sequence is highlighted in red, and the indel is indicated by dashes. (D) Protein domain and amino acid sequence of wild-type and mutant Rbm38. Residues translated after a frameshift are shown in red. (E and F) PCR and RT-qPCR analyses of rbm38 expression levels in wild-type and rbm38 mutant embryos at 72 hpf, with gapdh as a control. Data are presented as mean ± SD (Student's t-test; **P < 0.001).
Deficiency of Rbm38 causes abnormal enlargement of the pancreas
To determine whether the deficiency of Rbm38 impacts pancreatic development, we first monitored the expression of the pancreatic endocrine marker gene insulin and the exocrine marker gene prss1 at 72 hpf and 96 hpf using WISH. Both genes exhibited significantly expanded expression areas in the mutants compared to wild-type embryos at these stages (Figure 5A and B). Furthermore, fluorescence in situ hybridization (FISH) and quantitative analysis of the fluorescent signal area revealed a significant expansion of the insulin expression domain in rbm38 mutants (Figure 5C and D). We then crossed rbm38 mutants with the transgenic line Tg(fabp10a:dsRed;ela3l:GFP), which labels the liver and exocrine pancreas in red and green, respectively (Korzh et al., 2008). Consistent with the WISH results, the mutants displayed abnormal pancreatic enlargement at 72 hpf and 96 hpf (Figure 5E). Quantitative analysis of fluorescence areas at 72 hpf showed that the pancreas in rbm38 mutants was significantly larger than that of sibling controls (Figure 5F).
*Loss of Rbm38 leads to pancreas enlargement. (A and B) WISH analysis of pancreatic marker gene expression in wild-type embryos and rbm38 mutants at 72 hpf and 96 hpf. Lateral view. Scale bar, 100 μm. (C) FISH analysis of insulin expression in wild-type embryos and rbm38 mutants at 72 hpf and 96 hpf. Lateral view. Scale bar, 100 μm. (D) Dot plot compares the area of insulin expression in wild-type embryos and rbm38 mutants at 72 hpf. (E) Confocal z-stacks showing pancreas formation in wild-type embryos and rbm38 mutants under Tg(fabp10a:dsRed;ela3l:GFP) background at 72 hpf and 96 hpf. Lateral view. Scale bar, 100 μm. (F) Dot plot compares the pancreatic area in wild-type embryos and rbm38 mutants at 72 hpf. Data are presented as mean ± SD (Student's t-test; *P < 0.05; **P < 0.01; ***P < 0.0001).
We next examined the expression of transcription factor genes related to pancreatic development, including pdx1, ptf1a, isl1a, pax4, nkx2.2a, and neurod1 at 72 hpf. Except for a distinctive downregulation of pax4, expression levels of the other genes were all upregulated, particularly for pdx1 and ptf1a (Figure 6A). This upregulation was also observed at 96 hpf (Supplementary Figure S4A). We also analyzed the expression of endocrine cell-related genes insulin, glucagon a, glucagon b, ghrelin, and somatostatin, as well as exocrine cell-related marker genes cpa4 and prss1. Compared with wild-type embryos, the expression of several pancreatic marker genes was dysregulated in rbm38 mutants. Except for glucagon b, which showed no significant change, the expression level of somatostatin was downregulated, whereas expression levels of the other genes, particularly insulin and ghrelin, were significantly upregulated (Figure 6B; Supplementary Figure S4B). Additionally, glucose levels were significantly reduced in mutant embryos at 72 hpf and 96 hpf compared to wild-type embryos (Figure 6C). These findings suggest that the loss of Rbm38 disrupts the normal expression patterns of key pancreatic genes, which may cause the abnormal enlargement of the pancreas and potentially affect its function.
*Loss of Rbm38 leads to pancreatic dysfunction. (A) Relative expression levels of pdx1, ptf1a, isl1a, neurod1, nkx2.2a, and pax4 in wild-type embryos and rbm38 mutants at 72 hpf. The expression level in wild-type embryos was set as 1 after normalization to gapdh. (B) Quantitative analysis of the expression levels of endocrine cell-related genes (insulin, glucagon a, glucagon b, ghrelin, somatostatin, and sst2) and exocrine cell-related marker genes (cpa4 and prss1) in wild-type embryos and rbm38 mutants at 72 hpf. The expression level in wild-type embryos was set as 1 after normalization to gapdh. (C) Relative glucose levels in wild-type and rbm38 mutant tissues at 72 hpf and 96 hpf, with glucose level in wild-type embryos normalized to 1. Data are presented as mean ± SD (Student's t-test; *P < 0.05; **P < 0.01; ***P < 0.001; ***P < 0.0001; ns, not significant ).
Rbm38 binds to the 3′-UTR of pdx1 mRNA and reduces its stability
Previous studies have shown that Rbm38 can regulate the stability of target mRNAs by binding to the 3′-UTR region (Zhang et al., 2010; Zou et al., 2021). To investigate the molecular mechanism underlying the elevated expression of key pancreatic developmental transcription factor genes pdx1 and ptf1a in rbm38 mutants, the stability of pdx1 and ptf1a mRNAs was analyzed following actinomycin D treatment. We found that the half-life of pdx1 mRNA in rbm38 mutants was significantly longer than that in wild-type embryos (Figure 7A). However, the half-life of ptf1a showed no significant differences (Supplementary Figure S5A), suggesting that Rbm38 may regulate its expression through other mechanisms. Conversely, overexpression of Rbm38 in HEK293T cells reduced the stability of pdx1 mRNA and accelerated its degradation (Figure 7B).
*Rbm38 reduces the stability of pdx1 mRNA by binding to its 3′-UTR. (A) The stability of pdx1 mRNA after actinomycin D treatment in wild-type embryos and rbm38 mutants at 72 hpf. (B) The stability of pdx1 mRNA over time following actinomycin D treatment in control and HEK293T cells overexpressing Rbm38. (C) Schematic diagram illustrating the four segments (U1–U4) of zebrafish pdx1 3′-UTR fused with luciferase sequence. (D) Mapping of the three putative Rbm38-binding sites in the U2 region of pdx1 3′-UTR. (E and F) The CAGAUU sequence is required for Rbm38 to regulate luciferase activity in HEK293T cells. Values in empty vector transfected cells were set to 1 after normalization to Renilla luciferase activity. Data are presented as mean ± SD (One-way ANOVA; **P < 0.01; ***P < 0.001; ***P < 0.0001; ns, not significant).
The full-length 3′-UTR of pdx1 mRNA and different truncated forms were cloned into the pGL3 vector to generate luciferase reporter constructs (Figure 7C). These plasmids were then co-transfected with pcDNA3.1 or pcDNA3.1-rbm38 into HEK293T cells. The results showed that Rbm38 inhibited the activity of luciferase reporters containing the U2 region (Figure 7E), suggesting that it may bind to this region within the pdx1 3′-UTR. In addition, we identified three potential Rbm38-binding sites within the U2 region using the online prediction tool and constructed luciferase reporters with deletions of these sites (pdx1 3′-UTRΔ1, pdx1 3′-UTRΔ2, and pdx1 3′-UTRΔ3) (Figure 7D). In contrast to the deletion of binding site 1 (AUAUGU) and binding site 2 (AGUGCU), the deletion of binding site 3 (CAGAUU) abolished the inhibitory effect of Rbm38 on luciferase activity (Figure 7F), suggesting that Rbm38 can bind to the CAGAUU sequence in the pdx1 3′-UTR, thereby decreasing the stability of its mRNA.
Pdx1, a key pancreatic transcription factor, not only activates the expression of Insulin and Nkx2.2 but also promotes the transcription of cell cycle-related genes such as Ccnd1 and Ccnd2, thereby facilitating pancreatic cell proliferation (Baumel-Alterzon and Scott, 2022). In rbm38 mutants, although the expression levels of ccnd1, ccnd2a, and ccnd2b showed no significant changes compared to wild-type embryos, they were relatively higher in the absence of Rbm38 (Supplementary Figure S5B).
Rbm38 regulates pancreatic development via alternative splicing
To further explore the mechanism by which Rbm38 functions in pancreatic development, we performed RNA sequencing (RNA-seq) analyses of wild-type and rbm38 mutant embryos at 72 hpf. Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment uncovered a significant number of differentially expressed genes in the glycolysis, gluconeogenesis, and citrate cycle pathways (Figure 8A). These pathways are integral to glucose metabolism and energy balance, both of which are critical for pancreatic function.
Rbm38-regulated pancreas-specific alternative splicing events. (A) KEGG pathway analysis of differentially expressed genes in rbm38 mutants. (B) GO analysis shows enrichment of alternative splicing events in the pancreas. (C–E) Validation of altered splicing in rbm38 mutants at 72 hpf. Boxes highlight exons that are differentially spliced between wild-type embryos and rbm38 mutants. (F) Schematic diagram of the molecular mechanism underlying Rbm38 function in pancreatic development. The upper panel shows wild-type embryos, where Rbm38 maintains normal pancreatic development and tissue homeostasis by regulating the stability of pdx1 mRNA and modulating the alternative splicing of isl1a, nkx2.2a, and smad2, thereby fine-tuning the expression of pancreas-related genes. The lower panel shows rbm38 mutants, in which the loss of Rbm38 leads to increased stability of pdx1 and dysregulated splicing of several key transcription factor genes, resulting in pancreatic enlargement.
Rbm38 is a multifaceted RBP, regulating the stability of target mRNAs and also acting as a splicing factor. We employed rMATS to analyze the occurrence of five major types of alternative splicing events from RNA-seq data, including skipped exon, retained intron, mutually exclusive exon, alternative 5′ splice site, and alternative 3′ splice site. Given that the transcriptomic analysis was conducted on whole embryos at 72 hpf, we performed a Gene Ontology (GO) functional enrichment analysis on pancreas-specific genes (Figure 8B). The results revealed significantly altered alternative splicing events, suggesting that Rbm38 also plays a pivotal role in regulating the alternative splicing during pancreatic development.
We next experimentally validated the splicing defects of several key genes related to pancreatic development in rbm38 mutants at 72 hpf. In line with the results from the rMATS analysis, there was a skipped second exon in a transcript variant of isl1a (Figure 8C), a skipped fourth exon in smad2 (Figure 8D), and a skipped second exon in nkx2.2a (Figure 8E). These findings underscore the importance of Rbm38 in alternative splicing that is crucial for pancreatic development.
In summary, Rbm38 plays a crucial role in maintaining pancreatic homeostasis by destabilizing pdx1 mRNA and regulating the alternative splicing of key transcription factors, such as isl1a, nkx2.2a, and smad2, thereby fine-tuning the expression of genes involved in pancreatic development. In the absence of Rbm38, the stability of pdx1 mRNA is increased, and the splicing of several critical transcription factors is disrupted, leading to pancreatic enlargement and dysfunction (Figure 8F).
Discussion
Pancreatic development is a complex process that relies on the coordinated regulation of gene expression at both transcriptional and post-transcriptional levels (Apelqvist et al., 1999; Hebrok, 2003; Fujitani et al., 2006; Glauser and Schlegel, 2009; Leung, 2010; Jennings et al., 2015; Mi et al., 2024). Recently, an increasing number of RBPs have been demonstrated to play critical roles in pancreatic development through post-transcriptional regulation. In this study, we found that Rbm38 is predominantly expressed in the pancreas during zebrafish development and contributes to pancreatic development by regulating alternative splicing and mRNA stability of pancreatic genes. The loss of its function results in abnormal pancreatic enlargement.
Rbm38 displays highly restricted yet diverse expression patterns in early embryos, highlighting its involvement in a range of developmental processes. Furthermore, rbm38 exhibits dynamic expression within various tissues, including mesoderm, eyes, brain, spinal cord, neural plate, duodenum, liver, and hematopoietic tissues, as shown in Xenopus (Boy et al., 2004). Importantly, our WISH analyses, knock-in studies, and promoter reporter assays have uncovered that Rbm38 is highly expressed in the pancreas. This finding underscores the potential for Rbm38 in regulating pancreatic development. Indeed, we found that the loss of Rbm38 leads to abnormal enlargement of the pancreas during zebrafish development, accompanied by the upregulation of marker genes associated with both endocrine and exocrine glands. It has been shown that Rbm38-deficient mice exhibit symptoms such as hematopoietic disorders, splenomegaly, and cardiac enlargement (van den Hoogenhof et al., 2017). Furthermore, Rbm38 deficiency accelerates aging in mice and predisposes them to spontaneous tumor formation (Zhang et al., 2014). Although there is no definitive conclusion on whether the loss of Rbm38 affects pancreatic development in mice, the currently distinct phenotypes observed in zebrafish and mice implies that Rbm38 plays diversified roles in a tissue-specific and stage-specific manner.
RBPs typically associate with proteins and RNA to form RNP granules, which are involved in various cellular processes such as RNA transport, translation, and degradation (Pereira et al., 2017; Corbet and Parker, 2019). This is consistent with our observation that Rbm38 exhibits a granule-like distribution in SW1990 cells. Previous studies have shown that Rbm38 typically binds to AU-rich or GU-rich regions, which can regulate the degradation rate of mRNAs, thus affecting their half-life (Xu et al., 2013; Qian et al., 2020). We found that Rbm38 can recognize the CAGAUU sequence in the 3′-UTR of pdx1 mRNA, thereby decreasing its stability. The transcription factor PDX1 is the earliest expressed protein during pancreatic development, playing a crucial role in early pancreas formation and maintaining the function of mature β-cells (Jonsson et al., 1994; Gao et al., 2014). In zebrafish and mice, pdx1 exhibits highly similar expression patterns and functions, with predominant expression in pancreatic progenitor cells and mature β-cells. The loss of pdx1 in zebrafish severely disrupts normal pancreatic development, leading to a significant decrease in the β-cell number and insulin secretion, resulting in the increased blood glucose levels and metabolic disorders (Kimmel et al., 2015; Bastidas-Ponce et al., 2017). In early pancreatic progenitors, Pdx1 is essential for driving proliferation and pancreatic growth, while at later stages, it plays a crucial role in lineage specification, proliferation, and maintenance of selected cell types (Babu et al., 2007). Elevated Pdx1 expression stimulates pancreatic β-cell proliferation and enhances insulin secretion (Regue et al., 2021). This is consistent with our findings in rbm38 mutants, where increased pdx1 mRNA stability leads to upregulated insulin expression and a concomitant decrease in blood glucose levels. The loss of Rbm38 results in the upregulation of Pdx1 expression level, which may partially account for the abnormal enlargement of the pancreas.
Accumulating evidence suggests that Rbm38 participates in various biological processes through multiple post-transcriptional mechanisms (Shu et al., 2006; Zhang et al., 2010; Lucchesi et al., 2019, 2023; Li et al., 2020). Previous studies suggested an essential role of Rbm38 in alternative splicing during late erythroid differentiation, neurogenesis, and myogenesis (Anyanful et al., 2004; Heinicke et al., 2013; Amrane et al., 2014; Ganaie et al., 2018; Li et al., 2020; Han et al., 2022). Consistently, we found that several important pancreas transcription factor genes, including isl1a, nkx2.2a, and smad2, were aberrantly spliced in Rbm38-deficient pancreas. The LIM-homeodomain transcription factor Isl1 plays a critical role in regulating the expression of numerous genes, including Pdx1, Pax6, and Mafb, thereby influencing pancreas morphology and endocrine cell differentiation (Du et al., 2009; Ediger et al., 2014; Bastidas-Ponce et al., 2017; Scavuzzo et al., 2018; Bakhti et al., 2019; Bohuslavova et al., 2023). In vertebrates, Isl1 exhibits a highly conserved expression pattern and is continuously expressed in all endocrine cells. In isl1 mutant zebrafish, the initial specification of endocrine cells occurs normally, but more than half of these cells fail to successfully initiate endocrine hormone expression (Wilfinger et al., 2013). Interestingly, the Isl1 gene has two splicing variants, Isl1α and Isl1β, whose abundance differs between tissues (Ando et al., 2003; Bejarano-Escobar et al., 2015). Specifically, Isl1β is the predominant form in the pancreas (Ando et al., 2003; Lin et al., 2013; Jung et al., 2018). In addition, in vitro studies have shown that Isl1 directly regulates cyclin D1, suggesting a potential link between Isl1 and islet cell proliferation (Liu et al., 2012). Nkx2.2 governs crucial cell fate decisions in several developing organs, including the central nervous system, pancreas, and intestine (Seymour and Serup, 2023). Smad2 is an intracellular signaling factor activated by TGF-β signals and plays a vital role in regulating pancreatic islet function and maintaining β-cell mass (Harmon et al., 2004; Nomura et al., 2014). Pdx1 and Isl1a promote pancreatic cell proliferation by activating the expression of cell cycle-related genes such as* ccnd1*, while Smad2 indirectly influences this process through its involvement in the TGF-β signaling pathway (Harmon et al., 2004; Liu et al., 2012; Nomura et al., 2014). Whether these regulatory effects converge on a common signaling cascade remains to be elucidated through further investigation.
Our findings suggest that Rbm38 plays a pivotal role in pancreatic development by regulating alternative splicing and mRNA stability. Previous studies have demonstrated that Rbm38 is involved in regulating cell cycle and apoptosis through multiple post-transcriptional mechanisms, including pre-mRNA alternative splicing and mRNA localization, stability, polyadenylation, and translation (Zhang et al., 2010, 2011; Xu et al., 2013; Zhang et al., 2013; Lucchesi et al., 2016). Rbm38 is expressed in skeletal muscle cells and contributes to the differentiation process, at least in part, by post-transcriptionally regulating muscle gene expression. A similar function has been reported for its nematode ortholog, SUP-12 (Miyamoto et al., 2009; Grifone et al., 2020). Rbm24 is another homolog of the Caenorhabditis elegans SUP-12 (Miyamoto et al., 2009; Ray et al., 2009; Heinicke et al., 2013; Grifone et al., 2020). Both Rbm24 and Rbm38 display highly restricted yet diverse expression patterns during early development across various species (Fetka et al., 2000; Boy et al., 2004; Miller et al., 2008; Maragh et al., 2011, 2014; Poon et al., 2012; Grifone et al., 2014). Their expression patterns show remarkable similarity, which is consistent with their redundant functions (Miyamoto et al., 2009; Sun et al., 2016; van den Hoogenhof et al., 2017; Grifone et al., 2020). Interestingly, they also exhibit distinct expression profiles, indicating cell type-specific functions. For example, Rbm24 is essential for lens differentiation by regulating poly(A) tail length of crystallin mRNAs (Shao et al., 2020), whereas Rbm38 is crucial for differentiation of retinal neurons in Xenopus laevis (Boy et al., 2004). Therefore, Rbm38 and Rbm24 display both specific and redundant functions during vertebrate development.
RBPs also act in concert and/or mutually interact to post-transcriptionally regulate tissue-specific gene expression (Montanes-Agudo et al., 2023; Shi, 2024). Studies have demonstrated that Rbm38 collaborates with HuR to modulate the stability of p21 mRNA (Cho et al., 2010), while Rbm24, Rbm38, and PTBP1 function collectively to control the alternative splicing of exon 68 in Cdh23 (Li et al., 2020). Based on the String database, Rbm38 can interact with several other RBPs, many of which are important for pancreatic function (Bennett et al., 2010; Kuwasako et al., 2014; Moss and Sussel, 2020). For example, Rbfox1 and Rbfox2 are required for insulin secretion and blood glucose homeostasis (Juan-Mateu et al., 2017). Additionally, conditional mutation of Rbfox2 in the mouse pancreas leads to reduced insulin secretion and compromised blood glucose regulation (Moss et al., 2023). Whether Rbm38 interacts with Rbfox1, Rbfox2, or other RBPs during pancreatic development awaits future investigation.
In conclusion, we identified a specific expression of zebrafish Rbm38 in the pancreas and a novel role of this protein in pancreatic development. Our study also revealed an essential role of Rbm38 in regulating pre-mRNA alternative splicing and mRNA stability of key pancreatic genes.
Materials and methods
Zebrafish, embryo, and cell culture
The zebrafish AB strain was raised at 28.5°C in the standard zebrafish culture system (Haisheng, China). The Tg(fabp10a:dsRed;ela3l:GFP) transgenic line was purchased from the China Zebrafish Resource Center. Embryos were maintained at 28.5°C in a temperature-controlled incubator. To inhibit pigment formation, embryos were cultured in medium containing 0.0045% 1-phenyl-2-thiourea starting from 24 hpf.
SW1990 and HEK293T cells were cultured in Dulbecco's modified Eagle medium (DMEM) supplemented with 10% (v/v) fetal bovine serum (FBS; YEASEN) at 37°C in a humidified incubator with 5% CO_2_.
This study was performed according to the standard animal guidelines and approved by the Animal Care Committee of Ocean University of China (OUC-AE-2018108).
Expression profile analysis and transcription factor-binding site prediction
The expression profile of zebrafish rbm38 was retrieved from online single-cell transcriptome database (https://daniocell.nichd.nih.gov/index.html). Putative transcription factors binding to the rbm38 promoter were identified via AnimalTFDB 4.0 (https://guolab.wchscu.cn/AnimalTFDB4/). Potential Rbm38-binding sites on pdx1 mRNA were predicted using RBPmap (https://rbpmap.technion.ac.il/) and RegRNA 2.0 (http://regrna2.mbc.nctu.edu.tw/).
WISH and FISH
WISH experiments were conducted according to the standard protocol (Thisse and Thisse, 2008). Digoxigenin-labeled anti-sense probes were in vitro synthesized using SP6 RNA polymerase (Thermo Fisher) or T7 RNA polymerase (Promega). Stained embryos were imaged using a stereomicroscope (Nikon, SMZ1000). FISH experiments were performed by following the standard protocol (He et al., 2020), and images were acquired using confocal microscopy (Andor Dragonfly 600). The fluorescence area was quantified by ImageJ.
RT-qPCR
Embryonic tissues were homogenized in TRIzol, and total RNAs were extracted using the Total RNA Kit I (Omega). The cDNAs were synthesized using M-MLV reverse transcriptase (TaKaRa) in the presence of random primers following the manufacturer's instructions. RT-qPCR was performed to examine the expression profiles of rbm38 at the various development stages using gapdh as an input control.
Analysis of Rbm38 subcellular localization
The pcDNA3.1/V5/GFP vector was generated by inserting the GFP gene into the eukaryotic expression vector pcDNA3.1/V5-His A. The amplified rbm38 open reading frame was cloned into the pcDNA3.1/V5/GFP vector. Plasmids were transfected into SW1990 cells using the HieffTrans™ Liposomal Transfection Reagent (YEASEN) according to the manufacturer's instructions. Transfected cells were cultured in DMEM supplemented with 5% (v/v) FBS without penicillin and streptomycin for 24 h and then stained with DAPI, followed by washes with phosphate-buffered saline (PBS). Fluorescent images were acquired using a confocal microscope (Leica Sp8).
Generation of zebrafish rbm38 mutant and rbm38-GFP knock-in lines
The zebrafish rbm38 mutant line was generated using the CRISPR/Cas9 method by targeting the first exon. The sgRNA (5′-GGGCGACATCGATGAAGCGG-3′) was synthesized by in vitro transcription, and 200 pg of this sgRNA along with 320 pg of Cas9 protein (NEB) were injected into zebrafish embryos at the 1-cell stage. At 24 hpf, pooled genomic DNAs from 6–8 embryos were extracted using 25 μl of NaOH (50 mM), followed by neutralization with 25 μl of Tris–HCl (40 mM). The DNAs were then amplified using specific genotyping primers (Supplementary Table S1), and the resulting PCR products were subjected to Sanger sequencing. Upon confirmation of successful targeting, the remaining embryos were raised to adulthood and screened for the mutation. The rbm38-GFP knock-in zebrafish line was generated by following the established protocol (Levic et al., 2021), with some modifications. Specifically, the primers used for template amplification were modified with a phosphorothioate bond at the 5′ end of the first five bases. This modification has been shown to protect the template from degradation by exonuclease activity (Zhang et al., 2023). An sgRNA targeting the last intron of rbm38 (5′-TGAGGACTCGCAAAGTA-3′) was synthesized by in vitro transcription.
Immunofluorescence and pancreas size quantification
Zebrafish embryos at 72 hpf and 96 hpf were fixed using 4% paraformaldehyde at 4°C overnight. Immunofluorescence staining was performed according to the standard protocol (Li et al., 2025). The following antibodies were used: rabbit anti‐carboxypeptidase A (1:400, MCE, YA2486), mouse anti-GFP (1:400, Roche, 11814460001), Alexa Fluor™ 488-conjugated goat anti-mouse IgG (H+L) cross-adsorbed secondary antibody (1:400, Invitrogen, A-11001), and Alexa Fluor™ 555-conjugated goat anti-rabbit IgG (H+L) cross-adsorbed secondary antibody (1:400, Invitrogen, A-21428). Nuclei were counterstained with DAPI, and embryos were imaged using a confocal microscopy (Andor, Dragonfly 600).
Embryos derived from crossing between rbm38 mutants and the transgenic line Tg(fabp10a:dsRed;ela3l:GFP) were imaged using a fluorescence microscope (Leica, M205 FA). The fluorescence area was quantified by ImageJ.
Glucose quantification
Glucose quantification experiments were conducted according to the standard protocol (Sancho et al., 2010), with some modifications. Zebrafish embryos (30 per group) were collected at 72 hpf and 96 hpf, respectively. After washing three times with PBS, embryos were homogenized in 50 μl of PBS. Glucose levels were then quantified using a Glucose (GOD-POD) Assay Kit (Yeasen, 60408ES60) following the manufacturer's instructions.
Analysis of mRNA stability
The mRNA stability was analyzed by following a standard protocol with minor modifications (Zhang et al., 2017). Zebrafish embryos (30 for each condition) at 72 hpf were treated with 20 μg/ml of actinomycin D (MCE) or DMSO for 4 h and 8 h. HEK293T cells were treated with 10 μg/ml of actinomycin D or DMSO for 2 h, 4 h, 6 h, and 8 h. After treatment, total RNAs were extracted from the tails of zebrafish embryos or HEK293T cells for reverse transcription and RT-qPCR analysis.
Luciferase assays
HEK293T cells were transfected with plasmids using the HieffTrans™ Liposomal Transfection Reagent. After 24 h, cells were lysed, and the activity of firefly luciferase and renilla luciferase was measured using the Dual-Luciferase Reporter Assay System according to the manufacturer's instructions (YEASEN). Renilla luciferase was used as an internal control.
Statistical analysis
Quantitative data were expressed as mean ± standard deviation (SD) from at least three independent experiments. Statistical analysis was analyzed by one-way analysis of variance (ANOVA) using the software GraphPad Prism. P < 0.05 was considered as statistically significant.
Supplementary Material
mjaf025_Supplemental_File
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amrane S., Rebora K., Zniber I. et al. (2014). Backbone-independent nucleic acid binding by splicing factor SUP-12 reveals key aspects of molecular recognition. Nat. Commun. 5, 4595.25183497 10.1038/ncomms 5595 · doi ↗ · pubmed ↗
- 2Ando K., Shioda S., Handa H. et al. (2003). Isolation and characterization of an alternatively spliced variant of transcription factor islet-1. J. Mol. Endocrinol. 31, 419–425.14664703 10.1677/jme.0.0310419 · doi ↗ · pubmed ↗
- 3Anyanful A., Ono K., Johnsen R.C. et al. (2004). The RNA-binding protein SUP-12 controls muscle-specific splicing of the ADF/cofilin pre-m RNA in C. elegans. J. Cell Biol. 167, 639–647.15545320 10.1083/jcb.200407085 PMC 1781344 · doi ↗ · pubmed ↗
- 4Apelqvist A., Li H., Sommer L. et al. (1999). Notch signalling controls pancreatic cell differentiation. Nature 400, 877–881.10476967 10.1038/23716 · doi ↗ · pubmed ↗
- 5Babu D.A., Deering T.G., Mirmira R.G. (2007). A feat of metabolic proportions: pdx 1 orchestrates islet development and function in the maintenance of glucose homeostasis. Mol. Genet. Metab. 92, 43–55.17659992 10.1016/j.ymgme.2007.06.008PMC 2042521 · doi ↗ · pubmed ↗
- 6Bakhti M., Bottcher A., Lickert H. (2019). Modelling the endocrine pancreas in health and disease. Nat. Rev. Endocrinol. 15, 155–171.30504925 10.1038/s 41574-018-0132-z · doi ↗ · pubmed ↗
- 7Bastidas-Ponce A., Scheibner K., Lickert H. et al. (2017). Cellular and molecular mechanisms coordinating pancreas development. Development 144, 2873–2888.28811309 10.1242/dev.140756 · doi ↗ · pubmed ↗
- 8Baumel-Alterzon S., Scott D.K. (2022). Regulation of Pdx 1 by oxidative stress and Nrf 2 in pancreatic beta-cells. Front. Endocrinol. 13, 1011187.10.3389/fendo.2022.1011187 PMC 952130836187092 · doi ↗ · pubmed ↗