Molecular detection of Ehrlichia chaffeensis in marsh deer (Blastocerus dichotomus) and their parasitic Amblyomma triste ticks in Argentina suggests a local transmission cycle
Eliana Carolina Guillemi, María Marcela Orozco, Iara Figini, Paula Blanco, Marisa Diana Farber

TL;DR
Researchers found Ehrlichia chaffeensis in marsh deer and their ticks in Argentina, suggesting a local transmission cycle.
Contribution
This study provides the first evidence of Ehrlichia chaffeensis transmission in a natural setting involving wild deer and ticks in Argentina.
Findings
Ehrlichia chaffeensis DNA was detected in two marsh deer blood samples.
The pathogen was also found in the salivary glands of three Amblyomma triste ticks.
A previously reported genotype was identified through sequencing of specific molecular markers.
Abstract
Since the first finding of Ehrlichia chaffeensis in the Argentinian marsh deer populations in 2018, we have conducted ongoing surveillance efforts to better understand the ecological and epidemiological dynamics of this zoonotic tick-borne pathogen in the region. Over a period of 7 years (2018–2024) we analyzed blood and tissue samples from 40 marsh deer (Blastocerus dichotomus) and 13 parasitic Amblyomma triste ticks in the Paraná River Delta, Argentina. We identified E. chaffeensis DNA in two deer blood samples and in the salivary glands of three A. triste ticks parasitizing one of those deer. The approach used for E. chaffeensis detection in the tick sample ensured that the identified DNA came from an active tick infection, not residual host blood in the gut. After sequencing generic genes (16S ribosomal RNA [rRNA], 23S rRNA and groEl) and the specific E. chaffeensis…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
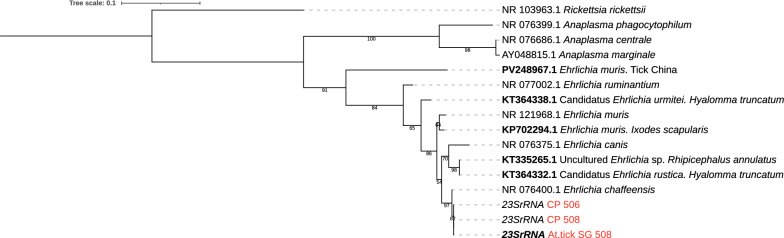 Figure 1
Figure 1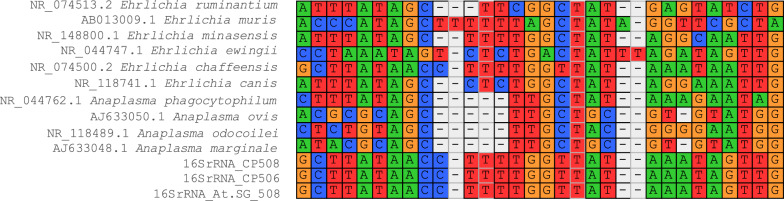 Figure 2
Figure 2 Figure 3
Figure 3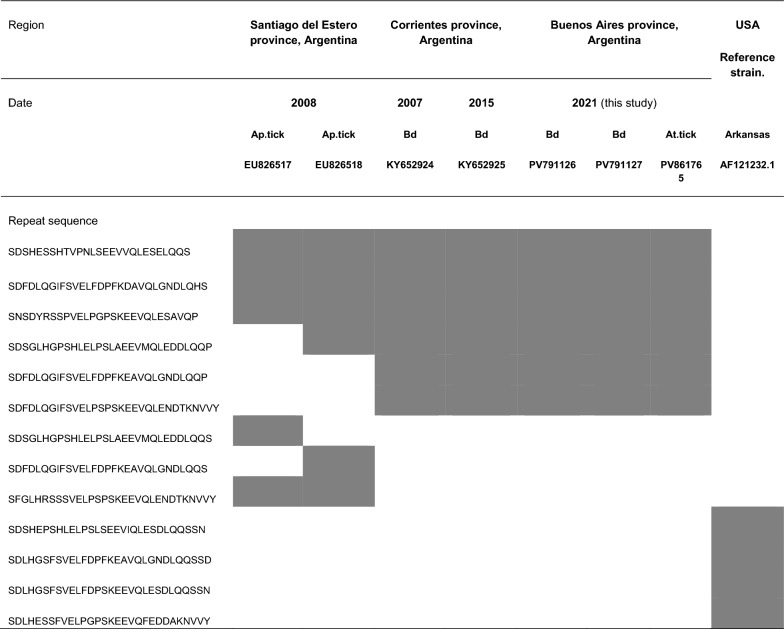 Figure 4
Figure 4- —Instituto Nacional de Tecnología Agropecuaria
- —https://doi.org/10.13039/501100003074Agencia Nacional de Promoción Científica y Tecnológica
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Zoonotic diseases and public health · Viral Infections and Vectors
Background
Emerging and re-emerging vector-borne diseases are among the major public health concerns around the world [1]. These diseases impact both human and animal populations, with increasing incidence driven by factors including climate change, habitat alteration, and urban expansion [2]. In response to this growing threat, the World Health Organization (WHO) launched the Global Vector Control Response 2017–2030 (GVCR), aiming to strengthen vector surveillance and promote integrated control strategies. In this context, the study of infectious agents at the human-animal-environment interface is a critical aspect of such efforts.
Ehrlichia chaffeensis (Anaplasmataceae family), is a Gram-negative obligate intracellular bacterium transmitted by hard ticks mainly among humans, deer, and canids [3]. When infecting humans, this tick-borne pathogen is responsible for a life-threatening disease known as human monocytic ehrlichiosis (HME) [4]. This flu-like illness usually requires hospitalization in more than half of the cases [5]. In the USA the HME is maintained in an epidemiological cycle involving white-tailed deer (Odocoileus virginianus) as the main vertebrate reservoir and the lone star tick (Amblyomma americanum) as its principal vector. In South America, E. chaffeensis has been detected in ticks (Amblyomma parvum and Riphicephalus microplus) and cervids (Blastocerus dichotomus and Mazama gouazoubira) through specific DNA target fragments [6–9]. Since HME is a flu-like neglected and underdiagnosed disease, few reports and confirmed cases are found in the bibliography. In the context of a cluster of tick-borne disease clinical cases in northern Argentina, Ripoll et al. [10] demonstrated for the first time in Argentina human serologic responses to E. chaffeensis or an antigenically related Ehrlichia species. Later, in 2016, Halac [11] described a case report of a 12-year-old child who, after being bitten by ticks, suffered unspecific symptoms and tested positive for E. chaffeensis or a related species, through serology (immunofluorescence assay [IFA]) and molecular detection (Ehrlichia sp.). Other South American countries have reported positive serosurves for Ehrlichia sp., including Brazil [12–15], Chile [16], and Peru [17]. Additionally, novel Ehrlichia species, different from but closely related to the E. chaffeensis, have been detected through molecular methods in Amblyomma species ticks in Argentina [18–20].
Even with these scarce reports, to the best of our knowledge, no studies have shown any evidence of a connection between a vector and vertebrate reservoirs for maintaining E. chaffeensis in these environments. In Argentina, our group reported the molecular detection of E. chaffeensis in A. parvum ticks from semiarid southern Chaco [7], and later in blood samples collected from marsh deer (Blastocerus dichotomus) in the alluvial plain of the Paraná River [9]. In this last study, we also detected E. chaffeensis in fully engorged R. microplus ticks collected from the same infected marsh deer, though we could not infer a vector role, as the blood meal was included in the analyzed specimens. Particularly, the marsh deer is a threatened native cervid from South America, primarily inhabiting wetland ecosystems across Argentina, Bolivia, Peru, Brazil, Uruguay, and Paraguay, with populations undergoing a marked decline due to multiple anthropogenic pressures, including habitat loss and degradation, illegal hunting, and diseases [21, 22]. Since this first finding of E. chaffeensis in the Argentinian marsh deer populations, we have conducted ongoing surveillance efforts to better understand the ecological and epidemiological dynamics of this zoonotic tick-borne pathogen in the region [23, 24].
Among the tick species parasitizing marsh deer, Amblyomma triste is one of the most frequent and widely distributed in the Southern Cone of South America. This three-host tick parasitizes a broad spectrum of wild and domestic mammals across wetland and riparian habitats [25]. Because Ehrlichia spp. are not transmitted transovarially, at least one of the developmental stages of the tick (larva, nymph, adult) must feed on competent reservoirs to maintain the bacterium in nature. Understanding the ecology, host associations, and seasonal dynamics of A. triste in the Paraná River Delta is therefore critical to interpret its potential involvement in E. chaffeensis transmission cycles. In the present study we analyzed the occurrence of E. chaffeensis in marsh deer from wetlands along the Paraná River, testing both deer samples and their Amblyomma triste parasitizing ticks.
Methods
Study area and sample collection
As part of a participatory surveillance, we studied morbidity and mortality events in the marsh deer population at the Paraná River Delta region in the Buenos Aires province of Argentina (located between 60° 39′ W, 32° 60′ S and 58° 30′ W, 34° 30′ S) [23, 24]. Over a period of 7 years (2018–2024) the members of the collaborative network remained alert for any reports of disease or mortality events affecting marsh deer populations. Upon notification of such events, a field team was mobilized to the affected site to conduct field investigations. Complete necropsies were performed on succumbed marsh deer, and biological samples were systematically collected. When possible, each deer was sexed and assigned to an age category (fawn, yearling, or adult) based on tooth eruption and wear patterns [26]. The body condition of the deer was determined using the Body Condition Scoring Chart [27]. Tick examination was conducted by visual inspection, and in those cases where the deer were parasitized, tick burden was estimated based on the method described by Orozco et al. [23]. Ticks were collected from deer using fine-point tweezers and preserved in tubes containing 70% ethanol until further identification. Full blood samples were collected from the heart of recently succumbed animals (within the hour) via cardiac puncture (2 mL). Aliquots were stored at 4 °C in tubes containing Ethylenediaminetetraacetic acid (EDTA), while additional aliquots were frozen at −20 °C and −80 °C without additives. For those dead deer that presented signs of autolysis, we sampled the ear rim by performing a tissue incision (1 × 1 cm) using a scalpel and conserved this at −20 °C until further analysis.
The study adhered to the ethical and biosafety standards of the Faculty of Exact and Natural Sciences, University of Buenos Aires, with approval from the Institutional Animal Care and Use Committee (Protocol N° 2021/164). Capture and transit permits were obtained from the National Parks Administration (NEA 513 Rnv 1).
Ticks taxonomic identification and processing
Ticks were identified under a stereoscopic magnifier (SMZ-2 T Nikon, Sendai, Natori, Japón) using taxonomic keys [28]. After identification (Electronic Supplementary Material [ESM] Fig. S1A–B), ticks were subjected to dissection to recover the salivary glands (SG). The specimens were first rinsed in 1 mL of sterile 1× phosphate-buffered saline (PBS; Sigma-Aldrich, Saint Louis, MO, USA), and then, the ventral cuticle was released using a scalpel blade (ESM Fig. S1C). The SG were identified, removed from the tick body (ESM Fig. 1D), washed with sterile 1× PBS, and conserved at −20 °C until further processing [29, 30]. To identify the contaminants potentially incorporated during sample processing [31], we systematically collected an aliquot of 1× PBS from each Petri dish after dissection (dissection controls).
Detection of E. chaffeensis from deer and tick samples
DNA extraction from deer blood and ear samples was performed using the DNeasy Blood and Tissue Mini Kit (QIAGEN. Hilden, Germany) following the protocols recommended by the manufacturer. DNA from the dissected tick’s SG was extracted using the Nucleospin Tissue Kit (Macherey–Nagel. Düren, Germany) following the manufacturer’s instructions. In all cases DNA was stored at −20 °C until further use.
Detection of E. chaffeensis DNA from deer and tick samples was conducted by amplifying family- and species-specific gene targets. We first screened all samples by targeting the 16S rRNA, 23S rRNA and groEl genes common to Anaplasmataceae [32–34]. For those positive samples we further used two sets of primers specific to E. chaffeensis, one of them targeting a fragment of the 16S rRNA gene [35] and the other targeting the variable-length polymerase chain reaction (PCR) target (VLPT) region [36]. This last protocol was also used for E. chaffeensis genotyping by sequencing the PCR fragment amplified from this size variation gene [36]. All PCR reactions were performed in 50 µL reaction mixture (0.2 mM of each deoxyribonucleotide triphosphate, 1.25 U of GoTaq DNA polymerase [Promega Madison, WI, USA], 10 μL of 5× PCR buffer). The primer final concentration was 0.4 μmol of each oligonucleotide except for the groEl gene reaction mixture. In this last PCR reaction, as primers are degenerate, we optimized final concentrations to 0.56 μmol (oligo_1236) and 0.8 μmol (oligo_643). Positive (Arkansas reference strain) and negative (pure water) controls were included in each PCR reaction. For visualization of the amplicons, 5 μL of mixture were visualized through electrophoresis in 1% agarose gel stained with ethidium bromide. A molecular size marker (1 Kb Plus DNA Ladder, Invitrogen) was used to determine PCR product size. For positive reactions, the remaining 45 μL of mixture were purified using a commercial kit (Monarch Spin PCR & DNA Cleanup Kit, NEB, Whitby, ON, Canada) according to the manufacturer’s instructions.
Both strands from 16S rRNA (Anaplasmataceae), 23S rRNA, groEL and VLPT fragments were sequenced with a Big Dye Terminator v3.1 kit from Applied Biosystems and analyzed on an ABI 3130XL genetic analyzer from the same supplier (Unidad de Genómica y Bioinformática, IABIMO INTA-CONICET). For each PCR fragment, forward and reverse chromatograms (ab1 files) were used for assembling, and the outcome contig (FASTA file) was used for further phylogenetic and characterization analysis.
Phylogenetic analysis and molecular characterization
Evolutionary analyses were conducted in MEGA6 [37]. Sequences from the 16S rRNA, 23S rRNA, and groEl genes (three gene targets common to Anaplasmataceae) were used for constructing phylogenetic trees. The evolutionary relation among sequences was inferred by using the maximum likelihood method based on the Kimura two-parameter model [38]. A discrete gamma distribution was used to model evolutionary rate differences among sites. Node robustness was assessed with 1000 bootstrap replicates. Apart from the sequences obtained in this study, we also included sequences from reference species in the family Anaplasmataceae and a sequence from Rickettsia species as outgroup. For tree visualization and labels edition, we employed the iTOL.v7 program [39].
For molecular characterization through VLPT target, we first employed an online bioinformatic tool available at Expasy website [40] for the translation of the nucleotide VLPT sequences into proteins. After translation, we searched for tandemly repeated amino acid sequences in each sample. The number of repeats and amino acid sequence type defined the genotypes present in each sample.
Results
Ticks taxonomic identification
All parasitic ticks studied were taxonomically identified as Amblyomma triste. We collected and analyzed 13 ticks; these specimens were all adults, corresponding to females (n = 2) and males (n = 11). These ticks were collected from two deer, CP_499 and CP_508, with low and medium tick burdens, respectively (Table 1).Table 1. Summary of ticks collected and analyzed in the present study from deer hosts CP_499 and CP_508Tick sourceAmblyomma tristeAdult femaleAdult maleCP_49923CP_508—8Total211CP_499 and CP_508 are the deer hosts’ tags
Detection of E. chaffeensis from deer and tick samples
We assessed the E. chaffeensis presence in blood or ear samples from B. dichotomus (n = 40) as well as in SG from parasitic ticks (n = 13) collected from two succumbed deer. From the 40 tested deer samples, 11 were positive to the generic Anaplasmataceae reactions (16S rRNA, 23S rRNA, and groEl genes), and 2 of them (CP_506 and CP_508) also tested positive for the E. chaffeensis-specific PCR reactions (16S rRNA and VLPT) (Table 2). Both E. chaffeensis–positive marsh deer were juveniles, with CP506 exhibiting a regular body condition and CP508 showing a poor condition. Additionally, three male A. triste ticks (from a total of eight collected) from a positive deer (CP_508) tested positive for the Anaplasmataceae generic (16S rRNA, 23S rRNA, and groEl), and E. chaffeensis-specific (16S rRNA and VLPT) reactions. The other deer hosting A. triste ticks (CP_499) and their SG tested negative to the PCR reactions. Those samples that tested positive for Anaplasmataceae PCR reactions but were negative for E. chaffeensis protocols (n = 9) were purified and sequenced for identification. These samples corresponded to Anaplasma species (data not shown). All tick dissection controls tested negative to the Anaplasmataceae 16S rRNA protocol.Table 2. Summary of the gene targets studied for each sampleSampleGene targets16S rRNA(Anaplasmataceae)23S rRNA(Anaplasmataceae)groEl(Anaplasmataceae)16S rRNA(E. chaffeensis)VLPT(E. chaffeensis)CP_506Sequenced(PV785953)Sequenced(PV786002)Sequenced(PV861766)PCR positiveSequenced(PV791126)CP_508Sequenced(PV785954)Sequenced(PV786003)Sequenced(PV868190)PCR positiveSequenced(PV791127)SG.At_508Sequenced(PV785955)Sequenced(PV786004)Sequenced(PV861764)PCR positiveSequenced(PV861765)CP: marsh deer samples; SG.At: salivary glands from Amblyomma triste tick. For those gene targets that were sequenced, accession numbers are mentioned in brackets
Ehrlichia chaffeensis population variability analysis
We used the obtained nucleotide sequences from the 16S rRNA (Anaplasmataceae), 23S rRNA, groEL, and VLPT fragments from both deer samples (CP_506 and CP_508) and from one of the positive ticks (SG.At_508) for further molecular characterization. All retrieved sequences have been deposited in GenBank (Table 2).
We first performed a local alignment using the BLAST bioinformatic tool available at the National Center for Biotechnology Information (NCBI) website (www.ncbi.nlm.nih.gov). Using the longest sequences obtained for each gene in our samples, the results of this alignment for the analyzed fragments retrieved the following sequences as first hits: for the 16S rRNA gene, 99.56% identity (CP_508 sequence) with E. chaffeensis reference strains (CP007476.1, CP007473.1, CP007477.1, CP007475.1, CP007478.1, CP007480.1, CP000236.1, CP007479.1); for the 23S rRNA gene, 99.81% identity (CP_508 sequence) with E. chaffeensis reference strains (CP007476.1, CP007473.1, AF416765.1, CP007475.1, CP007477.1, CP007478.1, CP007479.1, CP007480.1); and for the groEl gene, we found 99.83% identity (At.SG_508) with a recently proposed Ehrlichia sp. strain San Luis (KY425415.1) from A. tigrinum ticks and 99.16% identity with an E. chaffeensis identified in Blastocerus dichotomus from Brazil (JQ085941.1). To confirm the specific assignment and intraspecific variability, nucleotide sequences from 16S rRNA, 23S rRNA and groEl gene fragments were used for phylogenetic analysis. The tree with the highest log likelihood (−1655,2686) corresponding to a fragment of the 23S rRNA gene is shown in Fig. 1. We included reference sequences from the Anaplasmataceae family (AY048815.1, NR_076399.1, NR_076686.1, NR_077002.1, NR_076400.1, NR_076375.1, NR_121968.1) and a reference sequence from Rickettsia rickettsii (NR_103963.1) as an outgroup. Phylogenetic trees for 16S rRNA and groEl genes retrieved equivalent evolutionary associations (ESM). However, through the alignment of the 16S rRNA sequences with those from reference Anaplasmataceae species, we identified that the hypervariable V1 region in our samples corresponded to E. chaffeensis (Fig. 2).Fig. 1. Phylogeny for 23S rRNA gene based on 458 bp. Maximum likelihood tree. Substitution model: Kimura two-parameter model with discrete Gamma distribution. Phylogeny was tested with 1000 replications of bootstrap. Sequences corresponding to the samples under study (CP_506, CP_508, and SG.At_508) are labeled in red (bold font was used for the tick sample). Accession numbers from other Ehrlichia sp. found in tick samples are labeled with bold font. iTOL.v7 program was used for tree visualizationFig. 2Alignment of the hypervariable V1 region in the 16S rRNA gene from reference Ehrlichia and Anaplasma strains and those amplified in the present study (16SrRNA_CP506, 16SrRNA_CP508, and 16SrRNA_At.SG_508). Image obtained with the alignment tool from the Mega 6.06 program
For studying the subspecific variability of the VLPT target gene, the nucleotide sequences obtained were translated into protein sequences to further detect tandemly repeated amino acid blocks. VLPT sequences from both marsh deer samples (CP_506 and CP_508) and from the studied tick SG (At.SG_508) yielded identical amino acid sequences and thus the same tandem repeat profile. The VLPT tandem repeat profile detected for the three samples was also identical to the one previously identified for marsh deer samples in Argentina in previous years and shared some repeat units with VLPT genotypes detected in A. parvum ticks from the same country (Table 3).Table 3. Amino acid sequences for VLPT tandem repeats and profiles for previously reported E. chaffeensis in Argentina (in Santiago del Estero and Corrientes provinces), samples in the present study (Buenos Aires province) and the E. chaffeensis reference strain Arkansas (used as positive control)Ap.tick: Amblyomma parvum tick, Bd: Blastocerus dichotomus, At.tick: Amblyomma triste tick
Discussion
In the present study, we report the molecular identification of E. chaffeensis DNA in marsh deer blood samples and in salivary glands from its parasitic A. triste ticks. Through molecular detection we were able to amplify fragments from different target genes (16S rRNA, 23S rRNA, groEL, and VLPT) in blood samples from two deer and in salivary glands from three A. triste ticks. Additionally, we analyzed the sequences corresponding to these four genes from both deer samples (CP_506 and CP_508) and from one of the positive tick’s salivary glands (SG.At_508).
Through the phylogenetic analysis using reference sequences from the Anaplasmataceae family, we observed a clear association between our samples and the reference E. chaffeensis sequences, both for non-coding 16S rRNA and 23S rRNA genes and the protein coding groEl gene. Also, we incorporated into the groEl phylogenetic analysis, available sequences from novel proposed Ehrlichia species. These two strains, named Ehrlichia sp. strain San Luis [18] and Ehrlichia sp. strain delta [41], have been detected in A. tigrinum and A. triste ticks, respectively. One of them (Ehrlichia sp. strain San Luis) conformed a clade together with E. chaffeensis reference strain, an E. chaffeensis sequence retrieved from a B. dichotomus in Brazil [8] and our samples. We could not compare our 16S rRNA nor 23S rRNA sequences with those from Ehrlichia sp. strain delta and Ehrlichia sp. strain San Luis because sequences were not obtained for these strains or not available at GenBank. After species identification, further analysis enabled us to genotypically characterize the strains found. The VLPT gene encodes the surface-expressed TRP32 antigen. This protein has variable numbers of amino acid tandem repeats that are useful for E. chaffeensis genotypes identification [36, 42, 43]. In the present study we were able to amplify and sequence the corresponding VLPT fragments from both marsh deer samples (CP_506 and CP_508) and from the A. triste salivary glands (SG.At_508). The unique VLPT genotype identified for these samples was identical to previous genotypes detected in marsh deer from Argentina [9] and shared repeat blocks with previously reported E. chaffeensis in A. parvum tick samples from the same country [7]. The first identification of E. chaffeensis in Argentina in those A. parvum ticks took place 1000 km away from the area in which, years later, the pathogen was detected for the first time in marsh deer. It is worth noting that, while A. parvum ticks were sampled from a semiarid area, marsh deer and the A. triste ticks in the present study inhabit wetland areas thus highlighting the ubiquity of these findings despite the low detection frequency of E. chaffeensis in this country. The recurrence of these genotypes over a period of 17 years in Argentina suggests the circulation of a stable E. chaffeensis genotype in the country, different from the genotype corresponding to the North American Arkansas reference strain, originally described in the USA [44].
The identification of E. chaffeensis in the A. triste salivary glands, represents relevant information regarding the epidemiology of ehrlichiosis in Argentina. Since salivary glands and saliva play an important role in transmission of most tick-borne pathogens to the vertebrate host [45–47], the detection of E. chaffeensis in salivary glands suggests that A. triste ticks may be responsible for the pathogen transmission to marsh deer. This finding provides key evidence of a link that connects E. chaffeensis-positive marsh deer with a tick transmission cycle in the Argentinian wetlands, even though further studies including microscopy or culture among others, are needed to deepen the understanding of E. chaffeensis replication dynamics in this tick species. In a previous study we identified, for the first time in Argentina, E. chaffeensis in blood from marsh deer [9]. In that study, we also detected E. chaffeensis DNA in Rhipicephalus microplus ticks collected from positive deer, but due to methodological issues, we could not confirm that those ticks were actually infected and not harboring E. chaffeensis in their blood meal. In the present work we improved our methodological approach and studied the tick samples at an organ scale, thus analyzing individual salivary glands. This methodology guarantees that the detection of E. chaffeensis DNA in the tick sample corresponds to the presence of the Ehrlichia in the arthropod and not to an infected blood meal residue from the tick gut. The incorporation of dissection negative controls validated these PCR results.
From an ecological perspective, the results in the present study deepen our understanding of the pathogen’s natural cycle within a biodiverse and dynamic wetland ecosystem. The Paraná River Delta is not only a critical habitat for marsh deer, a vulnerable and declining species, but also a mosaic of ecological interfaces where human activity (e.g., tourism, livestock grazing, land conversion) increasingly overlaps with wildlife. Amblyomma triste is a tick species strongly associated with wetlands and environment prone to flooding in Argentina, Brazil, and Uruguay [28]. While primarily associated with marsh deer, adult stages of A. triste also parasitize a wide range of wild and domestic mammals, including humans [25, 48, 49], highlighting their potential as bridge vectors for zoonotic transmission. Additionally, small and medium-size rodents of the families Cricetidae and Caviidae are the main hosts for immature stages of A. triste [25, 50] and could be part of a potential enzootic cycle of E. chaffeensis in the area. Recently, an Ehrlichia species closely related to E. chaffeensis has been detected in an adult questing A. triste in the Paraná River Delta [41], suggesting a potential role of this tick in the transmission of Ehrlichia agents. Since Ehrlichia species circulate among tick stages through transtadial transmission (not transovarially), further studies in the area should focus on the identification of Ehrlichia species in immature stages of A. triste and their hosts in order to deepen the understanding of the whole enzootic transmission cycle. In the present work, the simultaneous detection of E. chaffeensis in both a wild vertebrate host (Blastocerus dichotomus) and its parasitic adult A. triste ticks in a natural setting brings new evidence regarding the plausible role of A. triste in the transmission of E. chaffeensis.
Taking into consideration the previous records of seroreaction to E. chaffeensis in humans in the country [10], the report of a clinical case [11], and the neglected and underdiagnosed condition of this pathogen in our country, more in-depth study of E. chaffeensis epidemiology in the area is required. This study represents the first direct evidence of a putative transmission cycle of this agent in Argentina with critical implications within a One Health framework. The convergence of threatened native fauna, generalist tick species, and expanding human activity in shared landscapes generate conditions conducive to pathogen emergence and spillover.
Conclusions
The detection of E. chaffeensis, the etiological agent of human monocytic ehrlichiosis in a biodiverse wetland ecosystem increasingly exposed to anthropogenic pressure, highlights the need of being aware of potential tick borne Ehrlichia species. This underscores the urgency of implementing integrated, multidisciplinary surveillance systems that monitor environmental, animal, and human health as interconnected components. Such an approach is particularly relevant in ecotonal regions, where ecological disturbance, biodiversity conservation, and public health risks converge.
Supplementary Information
Additional file 1. Figure S1. Amblyomma triste tick. (a) Dorsal view of an adult male specimen, (b) ventral view of an adult male, (c) ventral cuticle released from an adult male, and (d) salivary glands extracted from an adult maleAdditional file 2. Figure S2. Phylogeny for 16S rRNA gene based on 315 bp. Maximum likelihood tree. Substitution model: Kimura two-parameter model with discrete gamma distribution. Phylogeny was tested with 1000 replications of bootstrap. Sequences corresponding to the samples under study (CP_506, CP_508 and SG.At_508) are labeled with blue fonts. iTOL.v7 program was used for tree visualizationAdditional file 3. Figure S3. Phylogeny for groEL gene based on 307 bp. Maximum likelihood tree. Substitution model: Kimura two-parameter model with discrete gamma distribution. Phylogeny was tested with 1000 replications of bootstrap. Sequences corresponding to the samples under study (CP_506, CP_508 and SG.At_508) are labeled with green font (bold font was used for the tick sample). Accession numbers from other Ehrlichia sp. found in tick samples are labeled with bold font. iTOL.v7 program was used for tree visualization
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Duarte JMB, Varela D, Piovezan U, Beccaceci MD, Garcia JE. “Blastocerus dichotomus.” The IUCN Red list of threatened species. e.T 2828 A 22160916. 2016. Accessed 16 Jul 2025.
- 2Pereira J, Varela D, Aprile G, Cirignoli S, Orozco M, Lartigau B, et al. “Blastocerus dichotomus.” Categorization of Mammals in Argentina According to Their Risk of Extinction. Red List of mammals from Argentina. https://cma.sarem.org.ar/es/especie-nativa/blastocerus-dichotomus. Accessed 16 Jul 2025.
- 3Edwards KT, Goddard J, Varela-Stokes AS. Examination of the internal morphology of the Ixodid Tick, Amblyomma maculatum Koch, (Acari: Ixodidae); a “How-to” pictorial dissection guide. midsouth entomologist. 2009;2:28–39. www.midsouthentomologist.org.msstate.edu.
- 4Patton TG, Dietrich G, Brandt K, Dolan MC, Piesman J, Gilmore RD. Saliva, salivary gland, and hemolymph collection from Ixodes scapularis ticks. J Visual Exper. 2012.10.3791/3894 PMC 391258422371172 · doi ↗ · pubmed ↗
- 5Des Vignes F, Piesman J, Heffernan R, Schulze TL, Stafford KC, Fish D. Effect of tick removal on transmission of Borrelia burgdorferi and Ehrlichia phagocytophila by Ixodes scapularis Nymphs. J Infect Dis. 2001;183:773–8. http://jid.oxfordjournals.org/.10.1086/31881811181154 · doi ↗ · pubmed ↗
- 6Guglielmone AA, Nava S. Rodents of the subfamily caviinae (Hystricognathi, caviidae) as hosts for hard ticks (Acari: Ixodidae). Mastozool Neotrop. 2010;17:279–86. http://www.sarem.org.ar.