DNA methylation landscapes in human cells and their chromatin determinants
Wei Cui, Zhijun Huang, Gerd P. Pfeifer

TL;DR
This study explores how DNA methylation patterns are established in human cells and how chromatin modifications influence these patterns.
Contribution
The study provides new insights into the chromatin determinants of DNA methylation in human bronchial epithelial cells.
Findings
Lower DNA methylation regions are associated with PRC2 marks and H3K27 trimethylation.
H3K36 methylation is depleted in PRC2-marked regions, suggesting a regulatory crosstalk.
Methylated CpG islands show distinct relationships with H3K36 methylation compared to non-CpG regions.
Abstract
DNA methylation patterns are established during development and are propagated in a cell type specific manner, but these patterns may become aberrant during aging and cancer. Regions of alternating high and moderate to low levels of DNA methylation exist along all chromosomes in human cells. It is unclear how these distinct DNA methylation blocks are established. Most of the prior work in this area has been performed with mouse embryonic stem cells. Using whole genome bisulfite sequencing and chromatin-immunoprecipitation sequencing, we have profiled DNA methylation at single base resolution and various histone modifications in human bronchial epithelial cells. We found that many regions of lower DNA methylation (< 50%) are characterized by presence of the Polycomb repressive complex 2 (PRC2) mark, histone H3K27 trimethylation, but less so by the PRC1 mark histone H2AK119…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
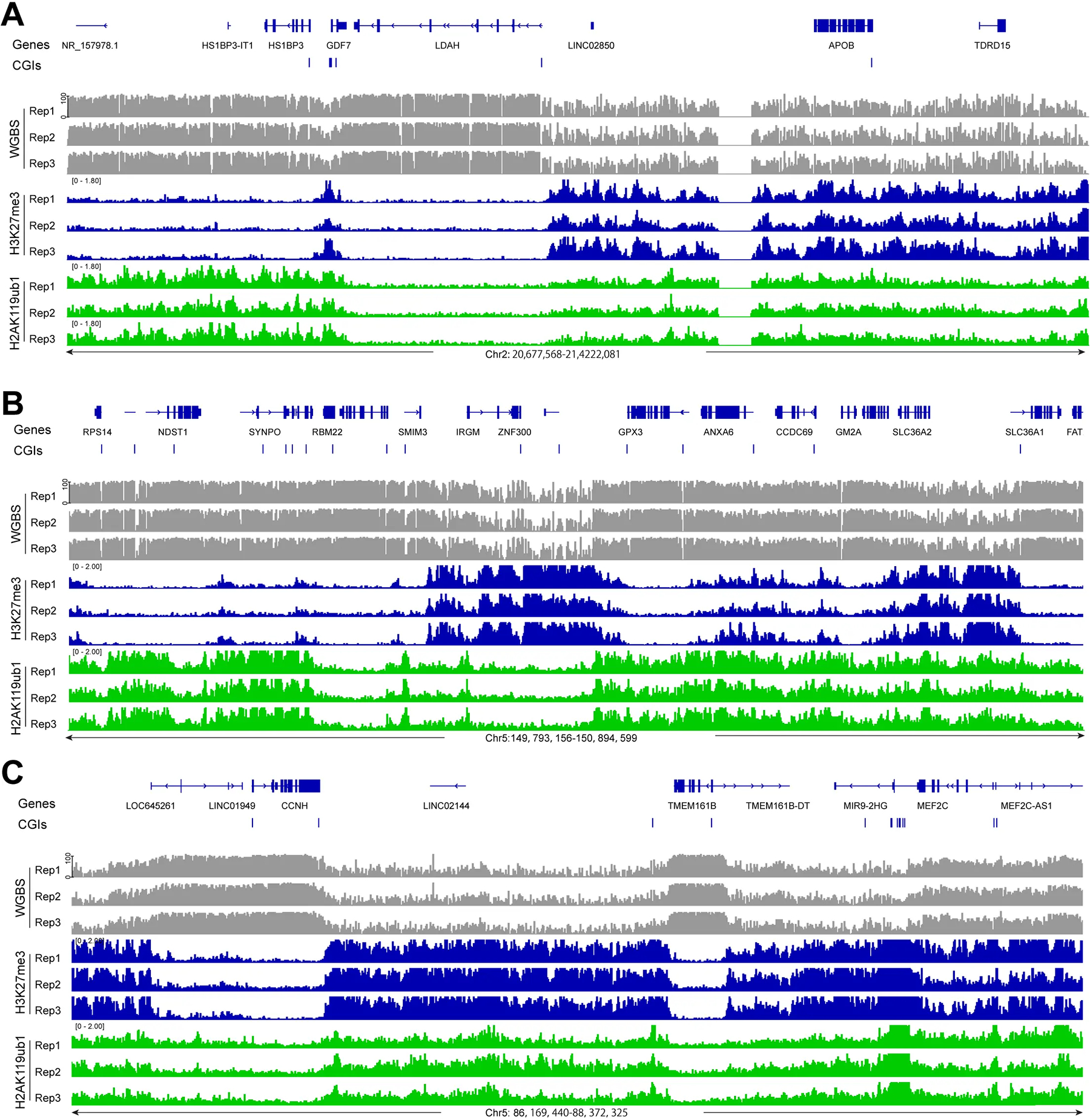 Figure 1
Figure 1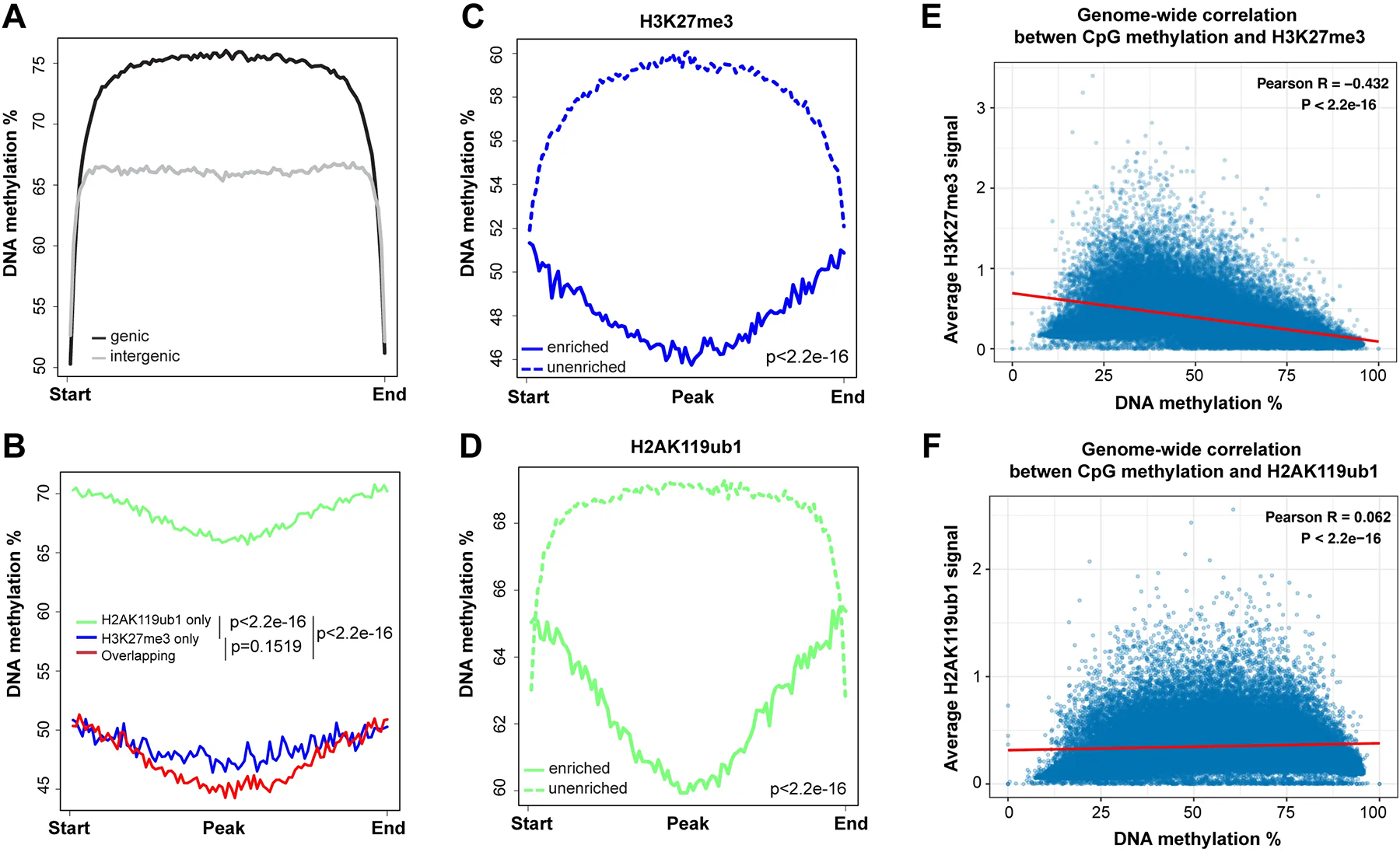 Figure 2
Figure 2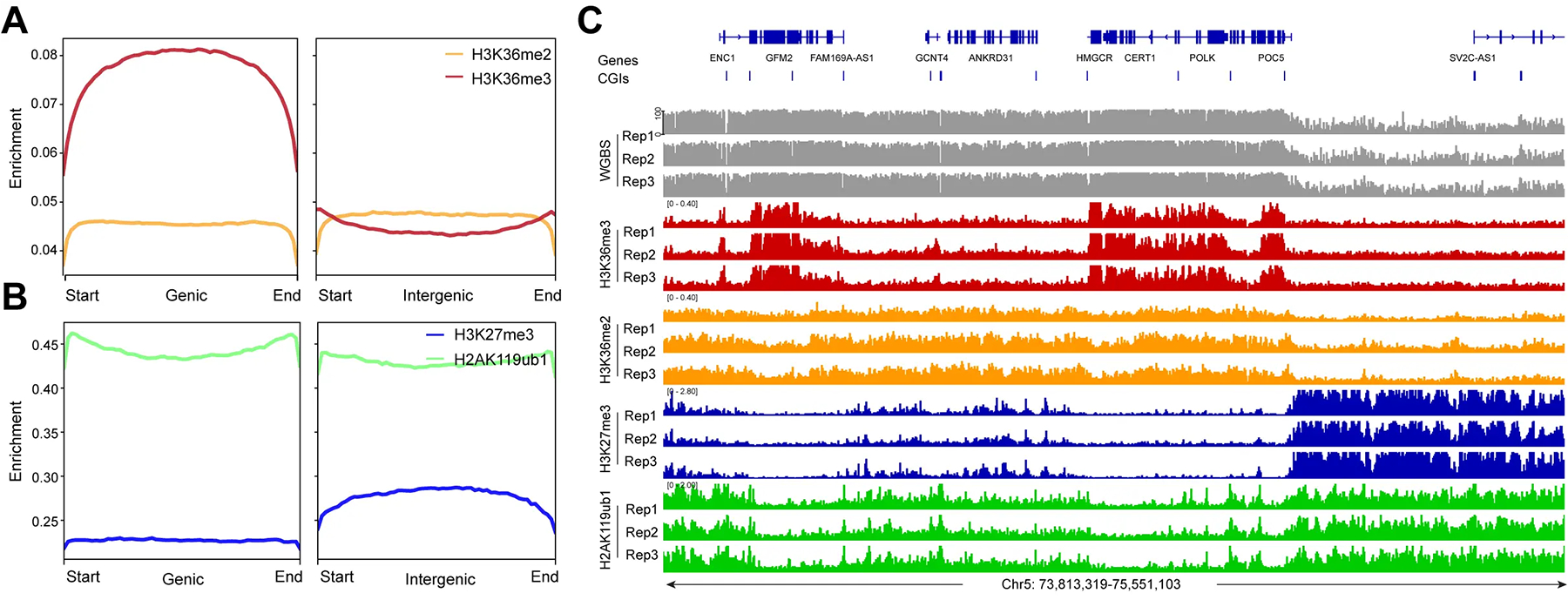 Figure 3
Figure 3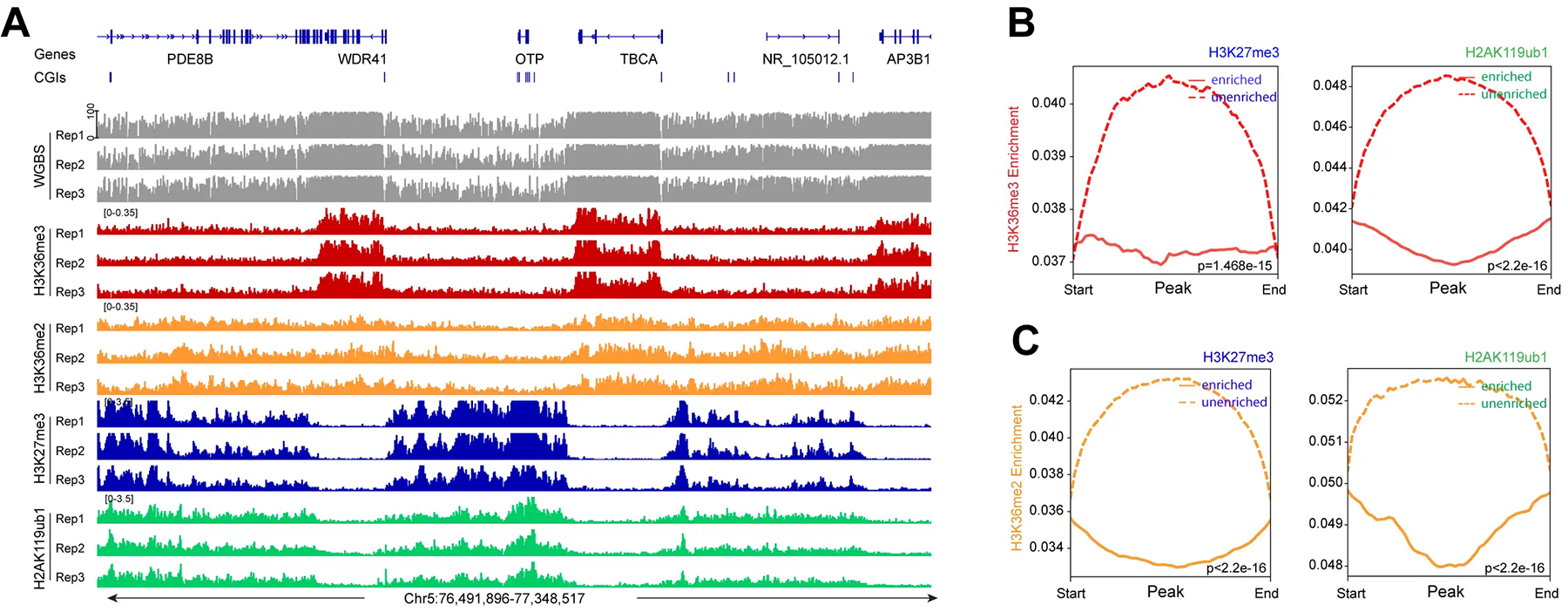 Figure 4
Figure 4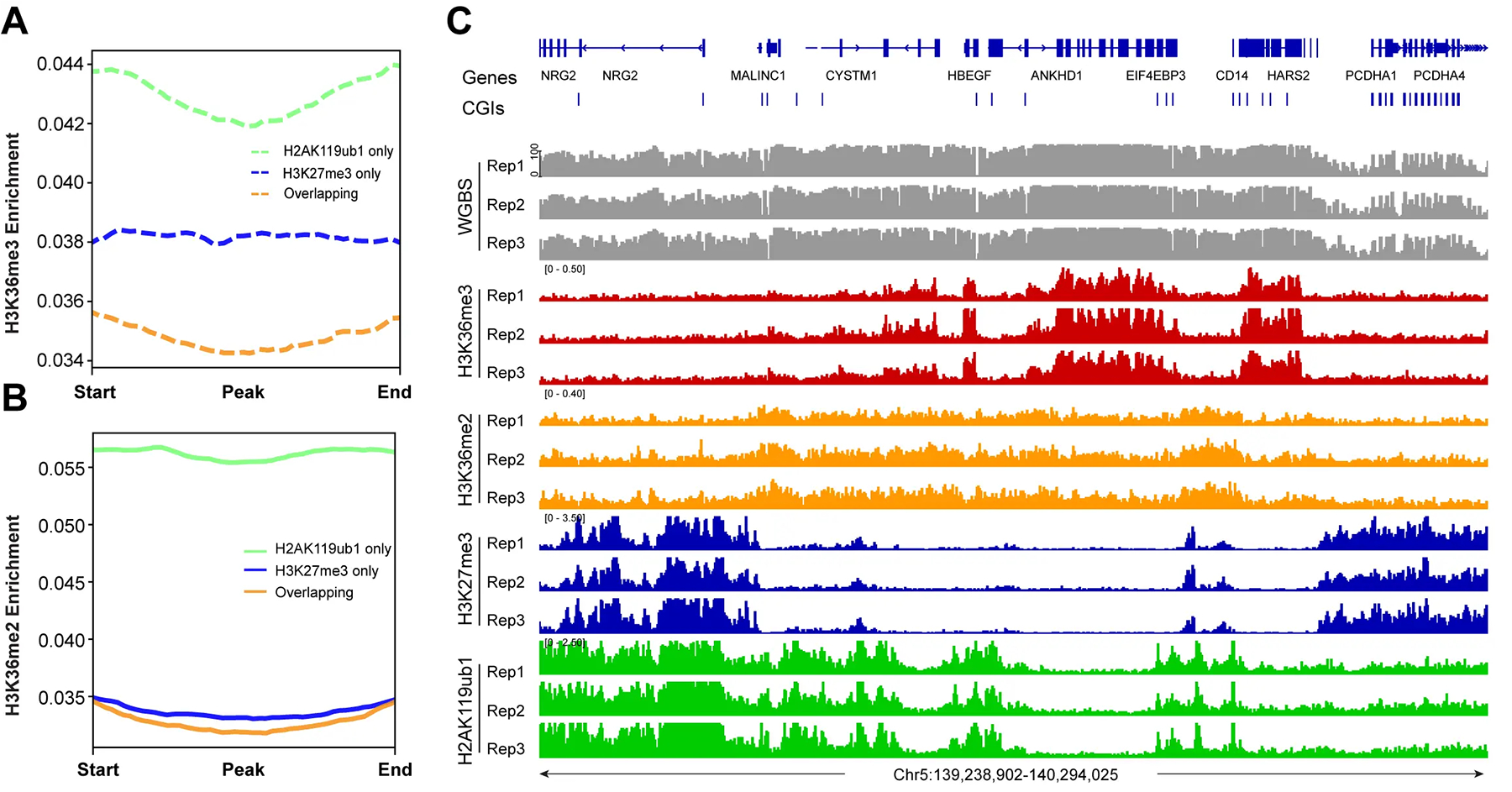 Figure 5
Figure 5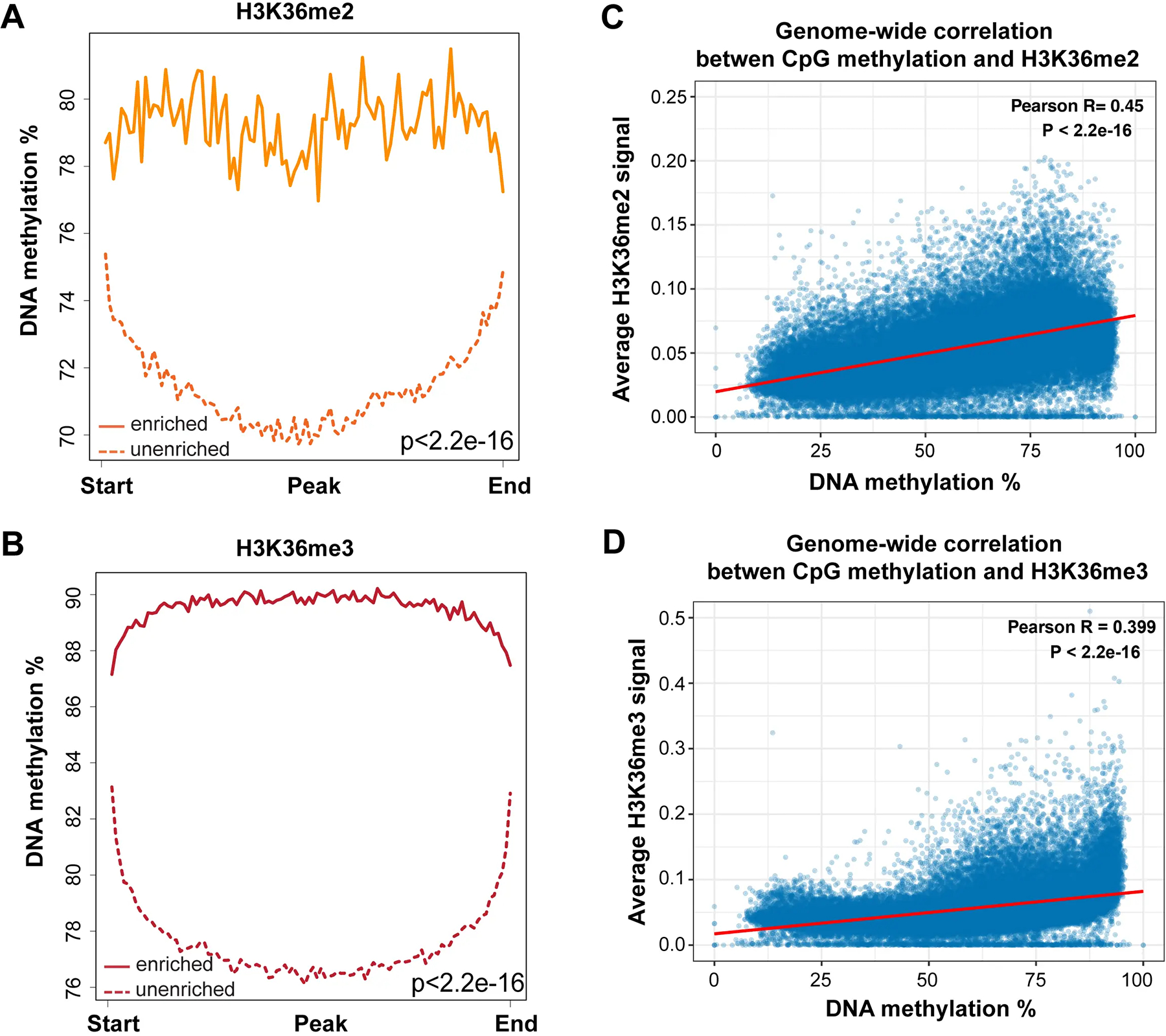 Figure 6
Figure 6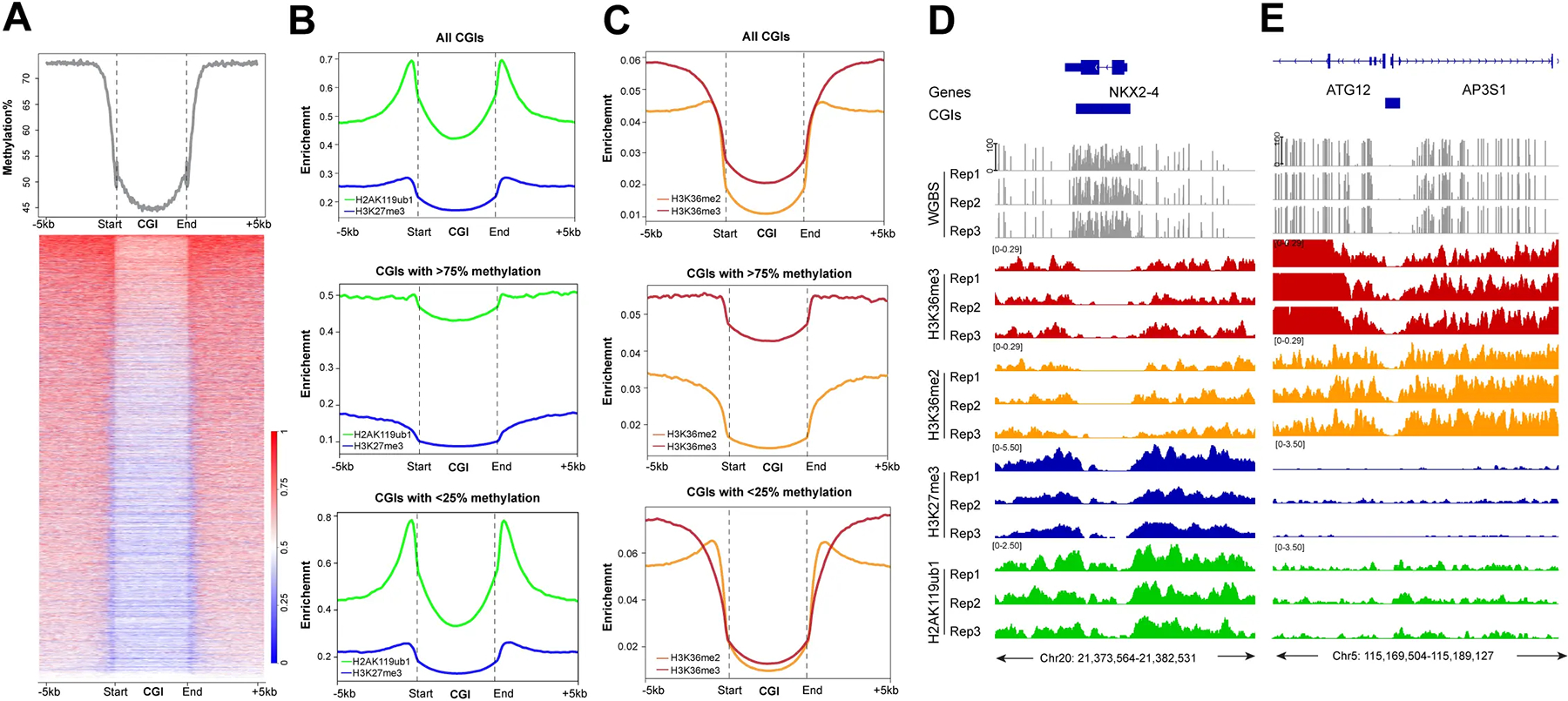 Figure 7
Figure 7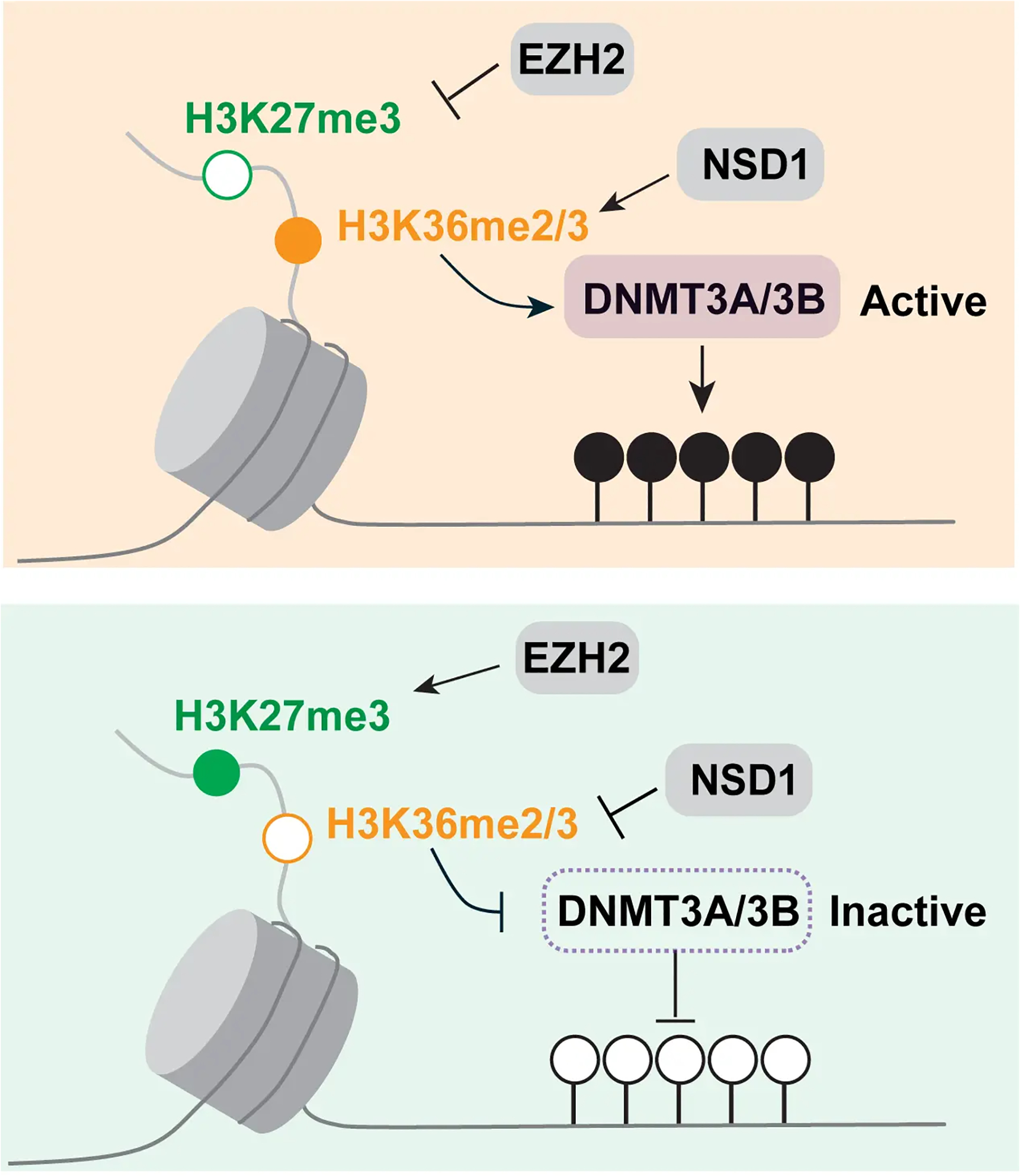 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEpigenetics and DNA Methylation · Genomics and Chromatin Dynamics · DNA Repair Mechanisms
Introduction
Somatic cells and tissues of mammalian organisms, with few exceptions, contain the same genome in terms of DNA sequence. Yet, there are hundreds of phenotypically different cell types, which raises the question of how such diversity can arise independently of the primary DNA sequence. In each cell, DNA is packaged into chromatin, a structure that consists of DNA wrapped around nucleosome core particles, which are further modified by a series of post-translational histone modifications. The DNA itself also is modified and carries methyl groups or hydroxymethyl groups, chiefly at CpG (CG) DNA sequences, where the position 5 of the cytosine ring is modified. Collectively, the assembly of modified DNA and histones, along with the three-dimensional packaging of chromosomes into defined territories, compartments, and topologically associated domains, is referred to as the epigenome. The epigenome is inherently plastic, allowing for changes during development and differentiation. Cell type identity is determined by different sets of epigenomes between each cell type. The most important readout of a specific epigenome is the establishment of cell type specific gene expression patterns.
Active genes generally carry a distinct set of defining epigenomic marks. They include an unmethylated, often GC-rich promoter, referred to as a CpG island^[1]^. At the chromatin level, active promoters are characterized by histone H3 lysine 4 (H3K4) methylation, with H3K4 trimethylation (H3K4me3) being a typical promoter mark^[2]^, and histone acetylation, for example, H3K27 acetylation. Downstream of transcription start sites, along the gene body, we find other specific marks, DNA CpG hypermethylation, H3K36 trimethylation (H3K36me3), and H3K79 trimethylation^[3,4]^.
Genomic regions that lack active genes are often associated with repressive histone marks. They have much lower levels of histone acetylation and carry the mark H3K9 trimethylation^[5]^. Such regions are referred to as constitutive heterochromatin and are often found in nuclear lamina associated domains (LADs)^[6,7]^. Some genes are repressed by a more flexible system known as facultative heterochromatin^[8]^. This means that these regions may be active during specific phases of development or in specific cell types but are otherwise repressed. A typical component of facultative heterochromatin is the Polycomb system. It consists of two sub-pathways, Polycomb repressive complex 1 (PRC1) and Polycomb repressive complex 2 (PRC2)^[9,10]^. These multi-protein complexes contain enzymatic activities that install histone modifications that interfere with gene activity. The most widely studied such activity is the EZH2 (and EZH1) histone methyltransferase activity of the PRC2 complex, which methylates lysine 27 at the N-terminal tail of histone H3 to produce H3K27me3^[9]^. The major enzymatic activity of the PRC1 complex is the transfer of one ubiquitin moiety onto lysine 119 of histone H2A to produce H2AK119ub1^[11–13]^. PRC1 and PRC2 complexes interact structurally and functionally^[9]^. They promote the compaction of chromatin to make it less accessible to transcription factors and RNA polymerase enzymes.
With the common use of genomic mapping techniques beginning about two decades ago^[14–16]^, much knowledge has been assembled about the specific features of histone modifications and DNA methylation patterns in many different cell types. Through numerous uncoordinated and coordinated (e.g., by ENCODE) efforts, the epigenome of commonly used cell types and mammalian tissues has been mapped comprehensively. Of great interest has always been the question if the epigenome is aberrant during the initiation of human diseases or during the aging process. For example, how would an altered epigenome that is found in one cell type promote a tissue-specific disease, and why would this alteration be tissue-specific? Through the work of countless laboratories, we now know that cancer cells are characterized by widespread epigenomic alterations. This field was initiated over four decades ago when it was reported that cancer cells from animals or humans have altered DNA methylation patterns, either genome-wide^[17,18]^ or at specific gene loci^[19]^. The methylation changes include genome-scale DNA hypomethylation, which affects many parts of the genome in different cancer types^[20]^, as well as more localized DNA hypermethylation events, which target mostly CpG islands^[21,22]^. In some cases, methylation of promoter-based CpG islands may silence critical genes, for instance, tumor suppressor genes or genes essential for maintaining the differentiated state of a somatic cell^[23–26]^. Although studied less widely and less systematically, alterations in chromatin structure are also found in tumors. Most notably, mutations in many cancer types are observed in genes that encode enzymes of the epigenetic machinery, for example, histone methyltransferases and acetyltransferases, components of Polycomb complexes, and chromatin remodeling factors^[27,28]^.
In addition to cancer and a few other diseases, the aging process also leads to a disruption of epigenomic patterns. This has most widely (in fact, almost exclusively) been studied for DNA methylation. Similar to what happens in cancer, aging leads to a slow general loss of DNA methylation and, at the same time, to an encroachment of DNA methylation into CpG islands^[29–31]^. Often, related groups of genes are affected in cancer and aging. Most notably, CpG islands targeted by the Polycomb complexes, such as developmental transcription factor genes, are methylated during carcinogenic processes^[16,32–34]^, but also during the aging process^[35,36]^ and during inflammation^[37]^, a condition that may predispose to cancer and aging. Since the discovery of an active DNA demethylation process about 15 years ago^[38]^, it has now become clear that the establishment, maintenance, and erosion of DNA methylation patterns are much more dynamic and complicated than previously thought. DNA methylation is established by DNA methyltransferases that methylate CpG sequences^[39]^. The active enzymes include the maintenance methyltransferase DNMT1 and the de novo methyltransferases DNMT3A and DNMT3B, which methylate previously unmethylated sites preferentially. The DNA demethylation process is initiated by oxidation of the methyl group of 5-methylcytosine by the Ten-Eleven-Translocation (TET) enzymes, TET1, TET2, and TET3^[38]^. The demethylation process is often defective in human cancer, either by mutation of TET2 or by defects in TET activity^[40]^.
However, what remains largely unclear to date is how these epigenomic patterns are established mechanistically in normal cells during development and how these patterns become aberrant in cancer. The extent of crosstalk between different epigenomic states and modification patterns along the genome is only partially understood. Most of the prior work has been done with mouse cells, preferentially mouse embryonic stem cells. One study analyzed the Polycomb-DNA methylation crosstalk in human cells^[41]^. However, it did not include data on H3K36 methylation. Another paper showed a correlation between highly methylated domains and H3K36me2, but it did not analyze H3K27me3^[42]^. In our own previous work, we reported that dysfunction of the Polycomb protein RYBP, which is a critical component of PRC1 complexes, along with loss of TET proteins, leads to widespread hypermethylation of thousands of CpG islands in human bronchial epithelial cells^[43]^. In that publication, we focused on CpG islands. From this published work^[43]^ and from additional chromatin modification mapping experiments, we have now obtained a detailed picture of how the different epigenomic parameters of DNA and histone modification overlap or antagonize each other. Here, we provide a genome-wide assessment of this crosstalk and show that a tripartite relationship between DNA methylation, H3K27me3, and H3K36 methylation largely correlates with long-range epigenomic states along the genome of human bronchial epithelial cells.
Materials and Methods
Cell culture
2.1
Human bronchial epithelial cells (HBEC-3KT, CRL-4051; RRID: CVCL_X491)^[44]^ were obtained from ATCC. Cell lines were authenticated by ATCC by short tandem repeat profiling. The cells were cultured in keratinocyte serum-free medium (K-SFM; Thermo, 17005042) containing 50 μg/ml bovine pituitary extract and 5 ng/ml human recombinant epidermal growth factor and were incubated at 37 °C with 5% CO_2_. The media were changed every 3 to 4 days.
DNA methylation analysis
2.2
Genome-wide DNA methylation patterns at single base resolution were determined by whole genome bisulfite sequencing (WGBS). The analysis pipeline for WGBS and the data obtained have been published previously^[43,45,46]^. The correlation analysis between DNA methylation and histone modifications based on the genome-wide signal of 50 kb bins was calculated by Pearson’s coefficients using R (v4.1.2).
ChIP-seq
2.3
ChIP-seq was performed as previously described^[43]^. H2AK119ub1 (CST, 8240), H3K27me3 (CST, 9733), H3K36me2 (CST, 2901S), and H3K36me3 (Abcam, ab9050) antibodies were used for immunoprecipitation. Briefly, cells were cross-linked with 1% formaldehyde for 10 min at room temperature, and the reaction was quenched with glycine. Cross-linked cells were lysed in lysis buffer and incubated on ice for another 10 min. The washed cell pellets were resuspended in shearing buffer and sonicated to shear the DNA into 300 to 500 bp long fragments. An optimal amount of chromatin, antibodies, and protein-G bead complexes was incubated overnight on a rotator at 4 °C. Purified DNA was quantified for library preparation with the Qubit sensitivity dsDNA HS Kit. Libraries were prepared using the TruSeq ChIP Sample Preparation Kit (Illumina, IP202–1012, IP-202–1024) according to the manufacturer’s instructions. The libraries were pooled and sequenced with an Illumina NextSeq500 or NovaSeq6000 instrument. A 5% spike-in of PhiX DNA was added to the runs to increase diversity and for quality control purposes. Each library had at least 50 million reads. Three replicates were generated in this study. The triplicates for each mapping experiment were highly correlated (Figure S1).
ChIP-seq data analysis
2.4
ChIP-seq data analysis was performed as previously described^[43,45]^. Briefly, the adapter and low-quality sequences were trimmed with Trimgalore (v 0.4.0) with default parameters, then the trimmed reads were mapped to the human genome hg19 with Bowtie2 (v 2.3.5). The deduplicated reads were used for peak calling by HOMER (v 4.10). Density plots of ChIP-seq enrichment at peak center regions were made with the R package genomation (version 1.28.0). For all overlap analysis of ChIP peaks, we used the bedtools intersection function, as well as for all other region overlap analysis in this study. Since regions enriched for a specific chromatin mark always alternate with unenriched regions (regions with no peaks) along the genome, their total numbers used in the analysis were almost identical. Pearson’s correlation coefficients between samples and groups were calculated using deepTools plotCorrelations, based on the genome-wide signal of 10 kb bins (Figure S1). Statistical testing (t-test) was used to compare the means of two groups and was performed using R (v4.1.2).
Results
DNA methylation is segregated into highly methylated and partially methylated regions along the genome
3.1
DNA methylation was analyzed at single base resolution by the WGBS method as described previously^[43,45]^. Triplicate samples of bronchial epithelial cell clones of the telomerase- and CDK4-immortalized cell line HBEC3 were used. Broader genomic regions at the scale of one to several megabases were inspected, as shown in Figure 1 for several chromosomal segments. Most genomic regions can be subdivided into segments of high DNA methylation (more than 75% methylation at most CpG sites) and intermediate or partial methylation (30–50% methylation). Using genome-wide analysis of genic and intergenic regions, we observed the highest DNA methylation levels in genic regions, also referred to as gene bodies (Figure 2A). This is consistent with prior work in other cell types^[47–49]^. Methylation levels are known to correlate with gene expression levels^[47,50]^.
We also find that many of the 30 to 50% methylated domains fall into gene-poor areas of the genome. Genome segments that are depleted of active genes tend to be associated with the nuclear lamina^[51]^. Although we lack data on LADs in this bronchial cell line, constitutive LADs are often common between different human cell types. The reason why LADs have lower DNA methylation levels has been debated. Suggestions include late DNA replication of these domains allowing less time for DNA methyltransferases to complete their reactions before mitosis^[52]^, a sequence preference for DNMTs that disfavors A/T-CG-A/T DNA sequences^[53]^, and as we proposed recently, a limited access of DNMT enzymes to regions attached to the nuclear lamina^[45]^. Despite the clear segregation of the DNA methylation patterns into highly methylated (> 75%) and partially methylated (30–50%) domains, it has remained unknown how these patterns are established and maintained along the genome.
Lower DNA methylation is best correlated with the PRC2 mark, H3K27me3
3.2
In parallel with DNA methylation, we mapped the distribution of various histone marks in the same cell clones. We focused here on the Polycomb marks H3K27me3, installed by the EZH1/2 histone methyltransferases of the PRC2 complex, and on H2AK119ub1, installed by the PRC1 complex. As can be seen in Figure 1 and several subsequent figures, the presence of regions strongly enriched in H3K27me3 generally coincides with regions of lower DNA methylation. When we compared DNA methylation levels over regions covered by H3K27me3, H2AK119ub1 or regions containing both Polycomb marks, we observed that DNA methylation levels are higher at H2AK119ub1 marked regions (65–70%) but much lower at genomic segments marked by H3K27me3 or by both marks (45–50%) (Figure 1, Figure 2B,C,D).
A genome-wide scatter plot analysis shows that DNA methylation levels are inversely correlated with H3K27me3 levels but show little overall correlation with the level of the PRC1 mark H2AK119Ub1 (Figure 2E,F).
H3K27me3 is reverse to H3K36me2 and H3K36me3
3.3
Next, we determined the genomic distribution of the H3K36me2 and H3K36me3 histone modifications (Figure 3). These marks are established by the NSD family of histone methyltransferases, NSD1, NSD2, and NSD3^[54]^. H3K36me3, produced by SETD2, is a mark preferentially found in gene bodies, where it likely accumulates in conjunction with RNA polymerase II progression^[55]^. It is thought that H3K36me3 promotes gene activity by suppressing intragenic initiation and antisense transcription. Genomic profile plots in Figure 3A show that H3K36me3 is indeed accumulating in gene bodies genome-wide, as opposed to intergenic regions. In contrast, H3K36me2 is more broadly distributed and is found along genes and in non-genic regions (Figure 3A). In Figure 3B, we establish the profiles of the Polycomb marks H3K27me3 and H2AK119ub1 along genic and intergenic regions. Genome-wide, the H2AK119 ubiquitin mark occurs in genes and in non-genic regions, but H3K27me3 is more abundant in intergenic segments compared to genes (Figure 3B). An example of this is shown in Figure 3C. Figure 3C shows a part of chromosome 5 with the distribution of these chromatin marks and that of DNA methylation determined by WGBS. The preferred distribution of H3K36me3 along genes can easily be recognized (Figure 3A,C), whereas H3K36me2 is found at genes and in between genes. This Figure also shows a noticeable reduction of histone H3 K36 modifications in an area with strong marking by the Polycomb system, as shown by the strong enrichment patterns of H3K27me3 and H2AK119ub1 (right side of Figure 3C).
We next investigated the crosstalk between the Polycomb marks and the H3K36 methylation system (Figure 4, Figure 5). It is easily visible that regions with high levels of H3K36me3 have extremely low levels of H3K27me3 or H2AK119ub1 (Figure 4A). When there are regions of H3K36me2, the Polycomb marks are also reduced. Conversely, genomic areas that are strongly marked by H3K27me3 and H2AK119ub1 show low levels of the H3K36 modifications. Figure 4B,C show a correlation profile analysis. At the genome-wide level, regions bound by the Polycomb marks show lower levels of H3K36me3 (Figure 4B) and H3K36me2 (Figure 4C). Figure 5 shows additional examples of this relationship. Profile plots of H2AK119ub1 show some enrichment for H3K36me3 (Figure 5A) and H3K36me2 (Figure 5B). However, when H3K27me3 is present alone or when it is overlapping with H2AK119ub1, the enrichment of the H3K36 modifications is much lower.
High DNA methylation levels are linked to H3K36me2 genome-wide
3.4
When we integrated the distribution of DNA methylation versus the different histone modification profiles, we found that high DNA methylation levels (78 to 90%) best correlated with H3K36me3 levels, and then also correlated with H3K36me2, the latter not being limited to gene bodies but more broadly distributed (Figure 3C, Figure 5C, Figure 6).
A genome-wide scatter plot analysis shows that DNA methylation levels are positively correlated with H3K36me2 and H3K36me3 levels (Figure 6C,D).
DNA methylation at CpG islands
3.5
In the last part of our analysis, we focused on CpG islands, CpG-rich genomic regions, commonly found at promoters that are often unmethylated when the gene is active. Indeed, most CpG islands of the genome have relatively low DNA methylation levels (Figure 7A), even though some of these regions are at least partially methylated, for example, when they occur in gene bodies or in intergenic regions. We then profiled the histone marks over all CpG islands. Both Polycomb marks can occur at CpG islands, with higher enrichment for H2AK119ub1 (Figure 7B). Interestingly, these Polycomb marks show higher levels at CGI borders. In contrast, H3K36me2/3 are quite strongly depleted at CpG islands with the lowest levels found for H3K36me2 (Figure 7C). To score differences between CpG islands that have low versus high DNA methylation levels, we separated the CGIs into groups containing either CGIs with less than 25% methylation or CGIs with more than 75% methylation (Figure 7B,C), and then profiled the histone marks. This analysis showed again very low levels of H3K36me2 in both categories of CpG islands, but higher levels of H3K36me3 at the more highly methylated CGIs. Most of these H3K36me3-marked CGIs were localized in transcribed gene bodies. These regions are preferentially targeted by de novo DNA methylation activities^[56]^. We show two examples: one is for a methylated CpG island at the NKX2–4 gene (Figure 7D), which is not transcribed in HBEC3 cells, and one for an unmethylated CpG island adjacent to the gene ATG12 (Figure 7E). Although there is some H2AK119ub1 modification over these regions, these CpG islands are generally well depleted of H3K27me3, H3K36me2, and H3K36me3. These data suggest that methylated CpG islands show a different relationship between DNA methylation and H3K36me2 deposition compared to non-CpG island regions.
Discussion
In this study, we established links between genome-wide DNA methylation profiles and four prominent histone modifications in a human bronchial epithelial cell line. We did not focus here on the well-known relationship between the promoter mark H3K4me3, which interferes with de novo methylation by DNA methyltransferases^[57]^ and is critical for keeping active and poised promoter CpG islands free of DNA methylation. The antagonistic relationship between DNA methylation and the Polycomb mark H3K27me3 has previously been noted^[30,41,58–62]^. In some experimental systems, for example, during the progression of normal cells towards malignancy, there can be a switch of the two repressive marks. Loss of the Polycomb mark at target genes may occur in cancer when it is replaced with DNA methylation^[37,63]^. In the opposite direction, when DNA methylation is reduced, for example, artificially by treatment of cells with a DNA methylation inhibitor, the demethylated regions may become occupied and modified by the Polycomb system^[64]^. In these situations, DNA methylation and Polycomb marking appear as opposite events, similar to what we have observed in our experimental system (Figure 1, Figure 2). The reason why this exclusion occurs is unclear. It is possible that highly CpG-methylated DNA interferes with Polycomb deposition since PRC complexes contain subunits (for example, KDM2B) that prefer binding to unmethylated CpG-rich DNA through their CXXC zinc finger domains^[65–68]^. On the other hand, once a region is occupied by Polycomb and the histones are marked by these activities, these complexes and modifications, perhaps through chromatin compaction, may not allow access of DNA methyltransferases. In our previous study, we found that depletion of the PRC1 component RYBP (RING1 and YY1 binding protein) in combination with the loss of TET proteins leads to DNA hypermethylation of several thousand CpG islands^[43]^.
However, our data suggest that the situation is probably more complicated because of additional crosstalk between specific histone modifications and between DNMTs and those histone modifications. First, the EZH2-catalyzed H3K27 methylation activity of PRC2 is sensitive to the methylation state of the H3K36 lysine residue, at least when present on the same histone tail^[69]^. The activity of EZH2 is much higher on H3 histone tails that are unmodified at lysine 36 compared to dimethylated or trimethylated H3K36^[70,71]^. Cryo electron microscopy studies have shown that histone H3 tails containing H3K36me3 interact poorly with PRC2^[72]^.
The methylation marks at H3K36 are dimethylation established primarily by NSD1, NSD2 and NSD3, as well as trimethylation installed by the SETD2 methyltransferase^[73]^. H3K27me3 is not known to block NSD or SETD2 activity, although their activity may be blocked by PRC1-induced H2AK119Ub1^[74]^. Loss of NSD1 leads to genome-wide expansion of H3K27me3 domains^[75]^. A mutually exclusive domain model may explain the stability of histone H3K27 and H3K36 histone methylation landscapes^[76]^. These inhibition models suggest that once an active domain with H3K36me2, and even more so with H3K36me3, has been established, it is difficult to overcome this active state by PRC-mediated Polycomb repression.
The H3K36 histone modifications explain not only the absence of H3K27 repressive marks at these regions but also the antagonism between H3K27 methylation and DNA methylation. This triangular relationship (Figure 8) exists because the H3K36 methylation marks positively induce DNA methylation. This was first shown for the de novo DNA methyltransferase DNMT3A, which interacts through its PWWP reader domain with methylated H3K36^[77,78]^. DNMT3B, which preferentially resides in gene bodies, also binds through its PWWP domain to H3K36-modified histones and prefers H3K36me3^[79–82]^. This finding may explain the high DNA methylation levels present in gene bodies, where this histone mark is prevalent. Later, it was shown that H3K36me2 is preferentially recognized by the PWWP domain of DNMT3A^[83]^. Since H3K36me2 is broadly present along many parts of the genome, this modification can explain much of the genomic DNA methylation patterns. Knockout of H3K36me2 methylation led to globally reduced levels of DNA methylation^[83]^. Interestingly, levels of H3K36me2 are also low in LAD regions, which provides yet another explanation why LADs contain only partially methylated domains^[45]^. As we showed recently, switching of LAD-associated genomic B compartments that contain heterochromatin to active euchromatic A compartments leads to an enhancement of H3K36me2 levels in the switched regions and to the acquisition of long stretches of DNA hypermethylation^[45]^.
The question may be asked: in which order these genome modifications are established and what it takes to overcome and shift these landscapes. The issues of cause or consequence are often difficult to discern in biological systems with steady state observations, as reported here. However, the available evidence from in vitro systems and from genetic manipulation experiments would suggest that the NSD- and SETD2-catalyzed marks are the primary determinants of both the Polycomb PRC2 mark setting and of the DNA methylation setting (Figure 8). Therefore, interference with the H3K36 methylation system may be an attractive approach to manipulate either DNA methylation—causing DNA hypomethylation—or to affect the Polycomb system—causing increased Polycomb-mediated repression. However, these two outcomes may neutralize each other in terms of their ultimate effect on gene expression. Furthermore, the exclusion of Polycomb complexes by DNA methylation and the exclusion of DNA methylation by Polycomb complexes may be an additional self-reinforcing mechanism for the propagation of these two epigenomic states.
One phenomenon that is widely prevalent in most cancer types and is found during the aging process as well is the methylation of Polycomb target genes. In this process, hundreds or even thousands of genes that are marked by the Polycomb system in normal cells, either in embryonic stem cells or in the corresponding normal tissue cells, undergo DNA hypermethylation at CpG islands. Even though this process has been observed almost two decades ago^[16,32,33,84,85]^, the mechanisms leading to this widespread DNA hypermethylation have remained elusive. The same process operates during tissue inflammation^[37,86]^ and during aging^[30,35,36,87,88]^. Our hypothesis is that Polycomb-marked CpG islands are normally protected from the DNA methylation machinery, but that this protection is inherently unstable and deteriorates over time, i.e., during aging^[30]^. One such pathway that protects from DNA methylation is oxidation of 5-methylcytosine by the 5-methylcytosine oxidases, the TET proteins. TET proteins are often dysfunctional in human cancer due to either mutation or reduced enzymatic activity^[40,89,90]^. This phenomenon can easily be observed by the almost complete absence of the primary TET oxidation product, 5-hydroxymethylcytosine, in human tumors of many different tissue origins^[91]^. Another protective mechanism against DNA methylation may be the presence of Polycomb complexes themselves at CpG islands. Support for a protection mechanism came from a recent study in which we inactivated all three TET genes (TET1, TET2, and TET3) along with the critical Polycomb component RYBP in human bronchial epithelial cells^[43]^. This manipulation led to DNA hypermethylation at thousands of CpG islands in the gene-modified cells and conferred a tumorigenic phenotype onto the cells. A large fraction of the hypermethylated regions corresponded to the hypermethylated CpG islands found in human squamous cell lung carcinomas^[43]^.
Given the widespread crosstalk between H3K27 methylation, H3K36 methylation, and DNA methylation, one wonders if the H3K36 methylation system plays any role in the methylation of Polycomb target genes. CpG islands are bound by the H3K36 demethylase enzymes, KDM2A and KDM2B^[65,66]^, which should reduce these modifications at unmethylated CpG islands, consistent with our findings (Figure 7). The H3K36 modifications are also largely absent from methylated CpG islands (Figure 7C,D), except for H3K36me3 at the bodies of active genes. It is possible, however, that NSD enzyme activities could forcefully introduce H3K36 methylation at Polycomb-marked CpG islands, which would interfere with PRC2 but promote de novo DNA methylation. In fact, NSD2 and NSD3 are upregulated at the transcript levels or carry activating mutations in human tumors^[92–96]^. Further work is required to better understand how these mutually dependent genome modification systems operate in different cell types and how they break down during aging and cancer.
Limitations of the study: The data in our study are mostly correlative. Therefore, we cannot infer that there is causation in one direction or the other. Only systematic perturbation experiments, in which the H3K27, the H3K36 modifications, or DNA methylation are removed, would allow us to establish mechanistically the order of events and the mutual dependency of these modifications. Doing that would represent a considerable effort, as multiple genes (e.g., NSD1, NSD2, NSD3, or EZH1, EZH2) would need to be targeted and the manipulated cells then analyzed for epigenomic patterns.
Supplementary Material
Supplementary material.
The supplementary material for this article is available at: Supplementary materials.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bird AP. Cp G-rich islands and the function of DNA methylation. Nature. 1986;321(6067):209–213.2423876 10.1038/321209 a 0 · doi ↗ · pubmed ↗
- 2Wang H, Helin K. Roles of H 3K 4 methylation in biology and disease. Trends Cell Biol. 2025;35(2):115–128.38909006 10.1016/j.tcb.2024.06.001 · doi ↗ · pubmed ↗
- 3Lam UTF, Tan BKY, Poh JJX and Chen ES. Structural and functional specificity of H 3K 36 methylation. Epigenetics Chromatin. 2022;15(1):17.35581654 10.1186/s 13072-022-00446-7PMC 9116022 · doi ↗ · pubmed ↗
- 4Ljungman M, Parks L, Hulbatte R, Bedi K. The role of H 3K 79 methylation in transcription and the DNA damage response. Mutat Res Rev Mutat Res. 2019;780:48–54.31395348 10.1016/j.mrrev.2017.11.001 · doi ↗ · pubmed ↗
- 5Padeken J, Methot SP, Gasser SM. Establishment of H 3K 9-methylated heterochromatin and its functions in tissue differentiation and maintenance. Nat Rev Mol Cell Biol. 2022;23(9):623–640.35562425 10.1038/s 41580-022-00483-w PMC 9099300 · doi ↗ · pubmed ↗
- 6Alagna NS, Thomas TI, Wilson KL, Reddy KL. Choreography of lamina-associated domains: Structure meets dynamics. FEBS Lett. 2023;597(22):2806–2822.37953467 10.1002/1873-3468.14771 PMC 10858991 · doi ↗ · pubmed ↗
- 7Manzo SG, Dauban L, van Steensel B. Lamina-associated domains: Tethers and looseners. Curr Opin Cell Biol. 2022;74:80–87.35189475 10.1016/j.ceb.2022.01.004 · doi ↗ · pubmed ↗
- 8Beck DB, Bonasio R, Kaneko S, Li G, Margueron R, Oda H, Chromatin in the nuclear landscape. Cold Spring Harb Symp Quant Biol. 2010;75:11–22.21502408 10.1101/sqb.2010.75.052PMC 3832107 · doi ↗ · pubmed ↗