Tissue-specific microbiota dictates the competitive dynamics of listeria species colonization
Juliette Poujol de Molliens, Carla Palacios-Gorba, Jazmin Meza-Torres, Jesús Gomis, Angel Gómez-Martín, Juan J. Quereda

TL;DR
This study shows how different Listeria species interact with host tissues and microbiota, influencing their ability to colonize and spread in ruminants.
Contribution
The study reveals tissue-specific microbiota influence the competitive colonization dynamics of Listeria species.
Findings
L. monocytogenes is more resistant to gastric fluid than L. ivanovii.
Gut microbiota outcompete and eliminate L. ivanovii and L. valentina.
L. monocytogenes and L. seeligeri outcompete intestinal commensal bacteria.
Abstract
The genus Listeria is heterogeneous and contains pathogenic and nonpathogenic species. Pathogenic L. monocytogenes and L. ivanovii have different environmental distributions, infect different hosts, and cause distinct syndromes. Here, we evaluated whether responses of different Listeria species to diverse host niches contribute to virulence heterogeneity and influence their environmental distribution. We assessed resistance to gastric and intestinal fluids, gut and uterus microbiota, and semen. L. monocytogenes was more resistant than L. ivanovii in gastric fluid, whilst L. seeligeri and L. valentina showed an intermediate phenotype. All the tested Listeria species resisted the intestinal fluid. Gut microbial communities outcompeted and eliminated L. ivanovii and L. valentina. However, L. monocytogenes and L. seeligeri outcompeted intestinal commensal bacteria. Our findings suggest…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
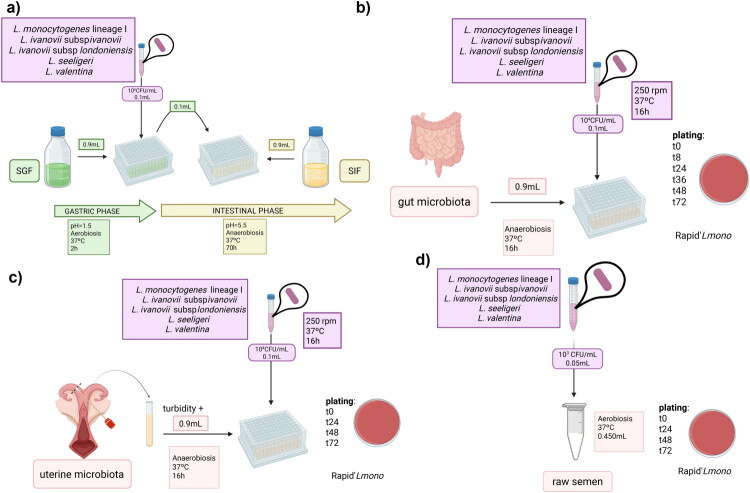 Figure 1
Figure 1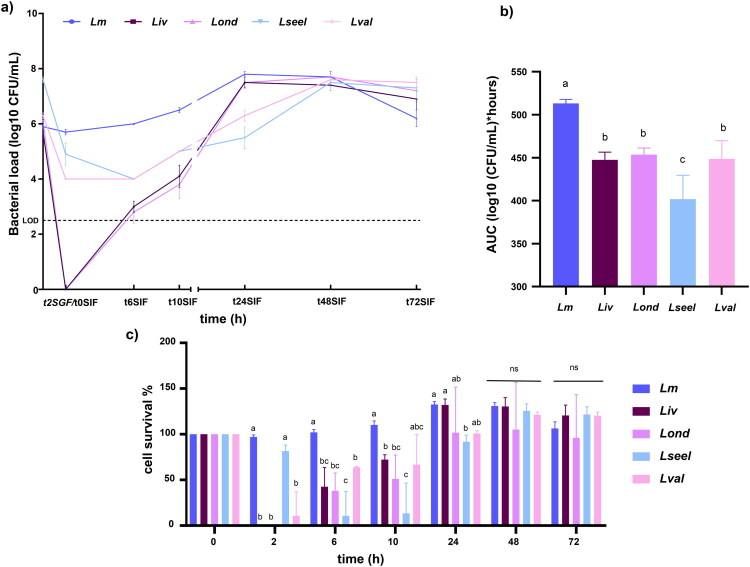 Figure 2
Figure 2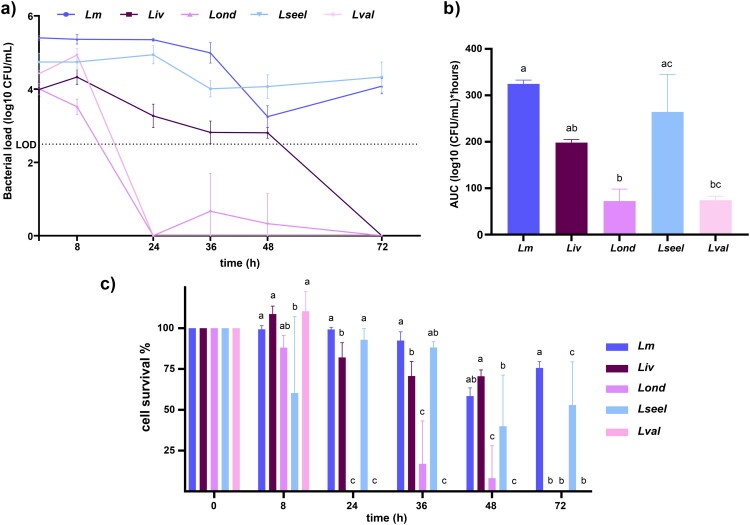 Figure 3
Figure 3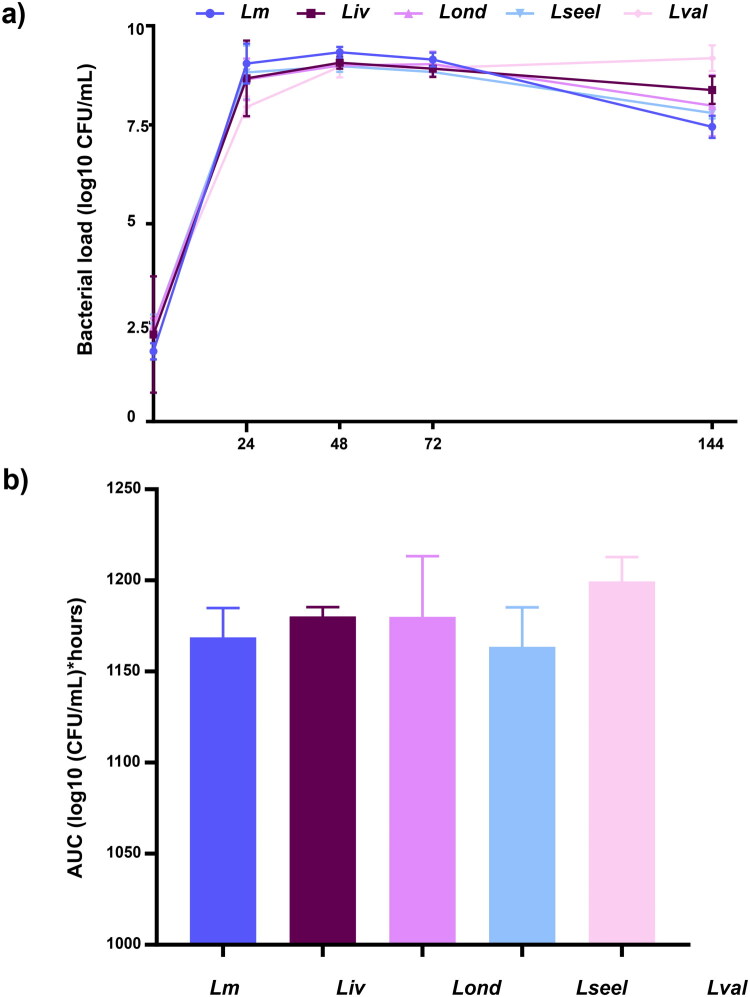 Figure 4
Figure 4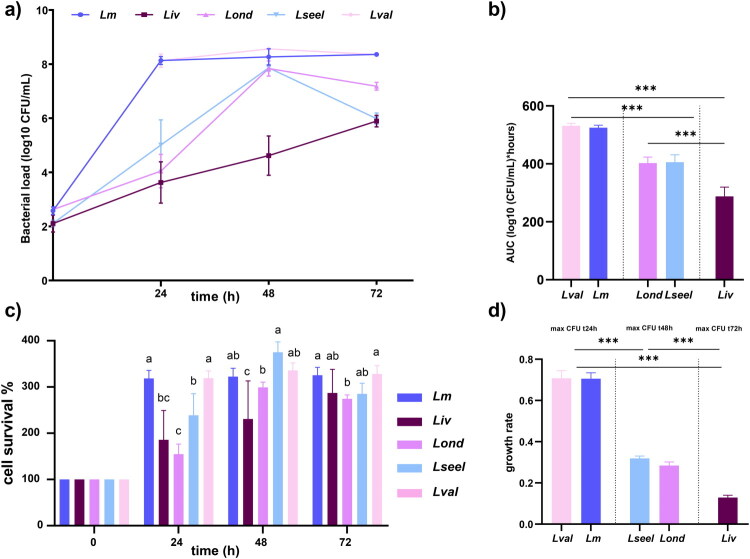 Figure 5
Figure 5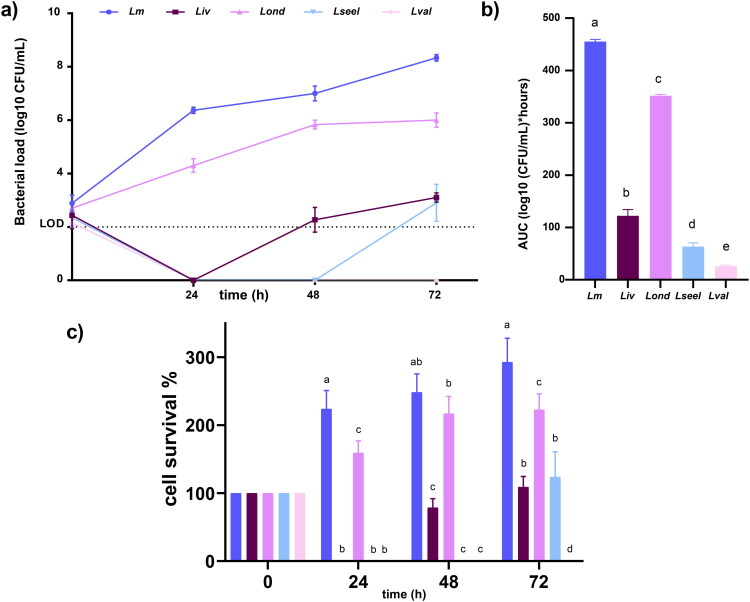 Figure 6
Figure 6- —Generalitat Valenciana10.13039/501100003359
- —Universidad CEU Cardenal Herrera10.13039/100031289
- —Universidad Cardenal Herrera-CEU
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsListeria monocytogenes in Food Safety · Salmonella and Campylobacter epidemiology · Vibrio bacteria research studies
Introduction
The genus Listeria consists of 29 widely distributed bacterial species, among which only Listeria monocytogenes (Lm) and Listeria ivanovii are recognized as zoonotic pathogens (Brown et al. 2025). Listeria species are diverse and display high diversity in virulence and environmental distribution (Orsi and Wiedmann 2016; Hafner et al. 2021). Pathogenic species encode, among other virulence factors, the Listeria pathogenicity island 1 (LIPI-1) and the inlA-inlB locus, which are necessary for key steps of intracellular parasitism (e.g. host cell adhesion, internalization, intracellular survival, and dissemination). Lm is the primary cause of listeriosis, affecting both ruminants and humans, particularly, the elderly, pregnant women and immunocompromised individuals. In these hosts, Lm causes septicemia, meningoencephalitis, abortion, and stillbirth (Quereda et al. 2021). In contrast, L. ivanovii, which comprises two subspecies (L. ivanovii subsp. ivanovii (Liv) and L. ivanovii subsp. londoniensis (Lond)), is mainly associated with ruminants, causing fetoplacental infections in pregnant ewes, goats, and cows. Interestingly, L. ivanovii has never been associated with neurological infections (Sergeant et al. 1991; Alexander et al. 1992; Vázquez-Boland et al. 2001; Guillet et al. 2010; Rossi et al. 2022). Moreover, only eleven human cases of gastroenteritis and septicemia caused by L. ivanovii have been reported to date (Guillet et al. 2010; Mani et al. 2025). Outbreaks of listerial encephalitis in domestic ruminants are associated with silage contamination; however, the source of infection for listerial abortion outbreaks remains elusive. Lm population is classified into four distinct lineages, with lineage I being strongly associated with clinical cases and considered hypervirulent (Maury et al. 2016). Notably, lineage I is predominantly linked to ruminants. The frequency of Lm fecal carriage ranges between 3.8% and 60% in domestic ruminants (Esteban et al. 2009; Hurtado et al. 2017; Palacios-Gorba et al. 2021a). Furthermore, Lm and Liv/Lond differ in prevalence and isolation sources. Lm is more prevalent than Liv/Lond in soil and host-associated environments (Hafner et al. 2021). Liv/Lond prevalence in feces is generally inferior to 2% in all the sampled niches, with some notable exceptions in poultry and wild rodents (Gaya et al. 1996; Gwida et al. 2011; Alvarez-Ordóñez et al. 2015; Cao et al. 2019; Palacios-Gorba et al. 2021a, 2021b, 2023). A tendency for L. ivanovii to be more easily isolated from aborted fetuses, milk, udders, tonsils, and vaginal swabs—particularly from sheep—rather than in fecal samples has been observed in some cases (Alexander et al. 1992; Gaya et al. 1996; Palacios-Gorba et al. 2021b, 2023). Different authors have suggested that L. ivanovii may be a venereal pathogen in small ruminants rather than a foodborne pathogen (Gray 1963.; McDonald 1967; Smith et al. 1967; Alexander et al. 1992; Wiedmann et al. 1999).
Some L. seeligeri (Lseel) isolates also possess LIPI-1 (as L. monocytogenes and L. ivanovii) and show hemolytic capabilities. Since an isolated L. seeligeri human meningitis case has been reported (Rocourt et al. 1986), it remains to be elucidated if this species is pathogenic in a specific, yet to be revealed, host species (Orsi and Wiedmann 2016). L. valentina (Lval) is a recently discovered species isolated from the feces of healthy sheep. It does not possess pathogenicity islands or virulence factors and differentially to Lm, Liv, Lond, and Lseel, belongs to the sensu lato group, and does not grow at low temperatures (Quereda et al. 2020).
The acidity of the gastric juice is the first barrier against pathogens ingested with food or water (Smith 2003). After passing through the stomach and reaching the small intestine, ingested Listeria cells must survive bile, pancreatic enzymes, and high osmolarity conditions. The use of artificial gastrointestinal systems that replicate the dynamics of gastrointestinal transit provides valuable information about factors affecting the survival of Listeria in the gastrointestinal tract (Ramalheira et al. 2010). Once these barriers have been endured, gut commensal bacteria can directly protect against pathogen colonization by competing for nutrients or producing bacteriocins (Oliveira et al. 2025). Similarly, uterine microbiota might provide colonization resistance against venereal pathogens (Benner et al. 2018; Wang et al. 2021).
To the best of our knowledge, no previous study has focused on using in vitro and ex vivo gastrointestinal systems of gut and reproductive microbiota models to decipher Listeria species virulence heterogeneity. Here, to gain insights into the ecological preferences of Listeria species described in the previous paragraphs, we examined differences in gastrointestinal survival among Lm, Liv, Lond, Lseel, and Lval using an in vitro simulation model of the stomach and small intestine. Next, we investigated their survival in ex vivo gut and uterine microbiota models and in semen.
Material and methods
Bacterial strains and CFU enumeration
In all the experiments, we used two L. ivanovii, one Liv and one Lond, one Lm belonging to CC1 from lineage I, one L. seel and one Lval, a specie isolated for the first time by our group in the south-east of Spain (Quereda et al. 2020). Their genetic characteristics and isolation sources are detailed in Table S1.
Colonies were obtained from −80 °C frozen aliquots that were plated on BHI agar plates and grown at 37 °C. A colony was then inoculated into BHI and grown overnight at 37 °C at 250 rpm for all Listeria species to OD = 0.9–1.2.
Simulated gastrointestinal fluids preparation
To prepare simulated gastrointestinal fluid (SGF) and simulated intestinal fluid (SIF), the following stock solutions were used: SGF consisted of sodium chloride (175.3 g/L), sodium dihydrogen phosphate (88.8 g/L), potassium chloride (89.6 g/L), calcium chloride (22.2 g/L), ammonium chloride (30.6 g/L), glucose (65.0 g/L), glucuronic acid (2.0 g/L), urea (25.0 g/L), glucosamine (33.0 g/L), bovine serum albumin (1.0 g/L), mucin type II from porcine stomach (3.0 g/L) and pepsin (1.3 g/L). The pH was adjusted to 2.5 with hydrochloride acid (1.0 mol/L). The volumes used of each solution are indicated in Table S2.
SIF-complete was prepared by mixing a duodenal juice and a bile solution. The stock solution used to make the duodenal juice consisted of sodium chloride (175.3 g/L), sodium bicarbonate (84.7 g/L), potassium dihydrogen phosphate (8.0 g/L), potassium chloride (89.6 g/L), magnesium chloride (5.0 g/L), urea (25.0 g/L), calcium chloride (22.2 g/L), bovine serum albumin (1.0 g/L), lipase (0.5 g/L), and pancreatin (3.0 g/L). The pH was adjusted to 7.5 with hydrochloride acid (1.0 mol/L). The bile solution was made with stock solutions of sodium chloride (175.3 g/L), sodium bicarbonate (84.7 g/L), potassium chloride (89.6 g/L), urea (25.0 g/L), calcium chloride (22.2 g/L), bovine serum albumin (1.7 g/L) and bile (40 g/L), which is equivalent to a bile concentration of ± 1%, as reported in the duodenum (Shah and Bergholz 2020). The pH was adjusted at 8.0 with sodium hydroxyde (1.0 mol/L). Three parts of duodenal solution and one part of the bile solution were mixed at room temperature to concoct the SIF-complete. The volumes of each solution are indicated in Table S2.
All the reagents and water were autoclaved for 15 min at 121 °C, and the enzyme solutions were prepared aseptically with sterile water and sterile filtered. The solutions were prepared and stored at 4 °C, except the enzyme solutions, which were made just before use. This protocol was previously published by Melo et al. (2013).
Growth experiments in simulated gastrointestinal fluids
The experimental process is adapted from Melo et al. (2013) and summarized in Figure 1a. Briefly, the overnight culture was adjusted to 10^6^ CFU/mL. Then 100 μL from the culture was mixed with 900 μL of SGF in a 96 deep-well plate, resulting in a final concentration of 10^5^ Listeria spp. CFU/mL. The inoculated SGF was then incubated for two hours at 37 °C in aerobiosis. After that, the culture was plated again, and 100 μL of the previously inoculated SGF was mixed with 900 μL of SIF-complete in a new deep-well plate and grown at 37 °C in anaerobiosis for an additional 70 h. Aliquots were withdrawn, diluted in PBS, and plated on BHI agar plates to enumerate the surviving Listeria at the subsequent times: 0, 2, 6, 10, 24, 48, and 72 h. The experiment was conducted, and the resulting data analyzed, between November and December 2024.
Experimental design for the study of listeria spp. growth abilities in a) SGF and SIF, b) an ex vivo gut microbiota, c) an ex vivo uterine microbiota, and d) raw semen.
Ethic statement
The sampling of the animals were approuved by the ethic of animal experimentation committee (CEEA: Comité de ética de experimentación animal) of the CEU Cardenal Herrera University. Moreover, the samplings were carried out on animals belonging to CEU Cardenal Herrera University during regulated practical classes held at the Faculty of Veterinary Medicine. The sampling of the animal is considered a routine academic and/or veterinary procedure; therefore, no further permit was required. For the same reason, no animal license numbers are available.
Ex vivo gut model coculture experiments
The experimental process is schematized in Figure 1b. Fresh stools were collected from goats and processed within 1 h. The collected samples were pooled and treated as single biological replicates to minimize variation between individuals (Kauer et al. 2025). The goats used as donors in this study belonged to the farm of the CEU Cardenal Herrera University and shared housing, limiting the impact of a different environment on the microbiota. All the goats used were healthy, and none had shown listeriosis symptoms previously. The animals were kept and fed with their standard balanced diet, and fresh water was provided ad libitum. Animal handling and health measures were those recommended for this species under current legislation. The sample was collected on April 9, 2025, and the following experiment was made immediately after. Three g. of stool were mixed with 30 mL of reduced PBS and vortexed for 30 s. The supernatants were collected after stool sedimentation and mixed with AF medium (18 g/L brain-heart infusion (Oxoid), 15 g/L trypticase soy broth (Oxoid), 5 g/L yeast extract, 2.5 g/L K_2_HPO_4_, 1 mg/L hemin, 0.5 g./L D-glucose, 0.5 mg/L menadione, 3% heat-inactivated fetal calf serum, 0.25 g./L cysteine HCl‧H_2_O) supplemented with sodium carbonate (0.4 g/L) and mucin type II from porcine stomach (2.5 mg/L) at a 1:5 ratio (Weiss et al. 2022). The microbiota in AF broth was then incubated at 37 °C overnight in an anaerobic workstation (Whitley A25). The resulting ex vivo microbiota was used for the following experiment.
For the assessment of Listeria spp. survival, each strain was cultured overnight at 37 °C and 250 rpm. The OD of each strain was measured at 600 nm, and the overnight diluted 1:10 in PBS to reach 10^6^ CFU/mL. Then, 900 µL of ex vivo microbiota were inoculated with 100 µL of Listeria spp. overnight culture (Listeria final concentration: 10^5^ CFU/mL) in 96-deep well plates, one strain per column, following the method proposed by Li et al. (2019). After 0, 8, 20, 32, 48, and 72 h, six drops of 10 µL were plated in Rapid’L.mono agar plates (Bio-Rad) to assess Listeria spp. survival when cultured with an intestinal microbiota ex vivo. All the data was analyzed in the following month.
Ex vivo uterine microbiota model coculture experiments
To explore the growth potential of Listeria species in the urogenital tract, we used a model replicating the uterine microbiota from an ewe. To do so, we sampled recently euthanised ewes culled for educational purposes from the authors’ university. The samples were collected in the first two weeks of May 2025 and processed immediately after (one sample was collected on May 1, and two samples on May 8). To euthanize the animals, an intravenous injection of barbiturate was used. Then, complete necropsies were carried out. Once the ewe was stabilized on the necropsy table, we checked for obvious reproductive lesions in the genital tract and for possible pregnancy. Only non-pregnant and without visible genital alterations ewes were selected for this study. We then disinfected the necessary material and one uterus horn with alcohol and opened a small window in its wall. We inserted one sterile standard calcium alginate swab (Puritan CalgiSwab) in the uterus horn, rubbing the uterine cavity firmly from the horn to the uterine body. The tip of the swab was cut to be introduced into a sterile Falcon tube filled with 30 mL of AF medium. The falcon tube was incubated at 37 °C in anaerobiosis for 16 h. The culture was considered successful if turbidity was observed the following day.
For Listeria spp. survival assessment, each strain was cultured overnight at 37 °C and 250 rpm. Overnight cultures were measured at 600 nm, diluted 1:10 in PBS 1x to reach 10^3^ CFU/mL. Then, 900 µL of ex vivo microbiota per well in 96-deep well plates were inoculated with 100 µL of Listeria spp. (Listeria final concentration: 10^2^ CFU/mL), one Listeria strain per column, following the method proposed by Li et al. (2019). After 0, 24, 48, and 72 h, six drops of 10 µL were plated in Rapid’L.mono agar plates (Bio-Rad) to assess Listeria spp. survival when cultured with a uterine microbiota ex vivo. This experimental process is resumed in Figure 1c. All the data was analyzed in the following month.
Growth experiments in semen
We selected four adult and healthy rams with proven fertility belonging to the flock kept at the CEU Cardenal Herrera University farm. The animals were fed with their standard balanced diet, and fresh water was provided ad libitum. Animal handling and health measures were those recommended for this species under current legislation. Rams belonging to this ovine experimental herd have never shown symptoms of listeriosis before. The raw ejaculate samples were collected with an artificial vagina, brought back to the lab, and pooled (Bahadi et al. 2023). They were aliquoted in five sterile eppendorfs tubes and kept in the dark at 37 °C. As previously detailed, Listeria spp. overnight cultures were diluted and inoculated in the semen at an inoculum level of 10^2^ CFU/mL. At each time point (0, 24, 48, and 72 h), 6 drops of 10 µL were plated in Rapid’L.mono agar plates (Bio-Rad) to assess Listeria spp. survival, as shown in Figure 1d. The semen samples were extracted the 15^th^ of May 2025 and inoculated with Listeria spp. within the hour. All the data was analyzed in the following month.
Statistical analysis
To examine the statistical differences between species, the analysis of variance test (ANOVA) and Kruskal-wallis tests were performed by using SPSS (IBM, version 27.01.0). The tests were applied to logarithmically transformed counts, calculated area under the curve (AUC), calculated growth rate, and cell survival percentages. In the SGF and SIF experiments, as well as.in the co-culture of Listeria with microbiota, 6 technical replicates of each isolate were made. For the validation of the culture of uterine microbiota (Listeria spp. in AF medium) and the inoculation of Listeria spp. in semen, 4 technical replicates were made. Differences were considered significant when P ≤ 0.05.
Results
Survival in simulated gastrointestinal fluids
The survival of Listeria species cells in simulated gastrointestinal conditions is illustrated in Figure 2. SGF exposure for two hours had variable inhibitory activity but reduced Listeria species viable CFU in all cases. The kinetics of SGF clearance varied significantly across the different Listeria species tested, indicating that acidic conditions of the digestive system do not contribute to the same extent to eliminating the Listeria species. As previously reported, a lack of substantial sublethal injury in Lm was detected (Barmpalia-Davis et al. 2008; Ramalheira et al. 2010; Rahman et al. 2020), probably due to acid adaptation mechanisms. Both clones of the subspecies of L. ivanovii displayed the highest acid sensitivity (P < 0.05), followed by Lseel and Lval. All the tested Listeria species recovered in the intestinal fluid (Figure 2a and c). Lm reached the stationary phase (7.8 Log_10_ CFU/ml) in SIF at 24 h (Figure 2a). Upon virtual clearance after SGF contact, SIF exposure resulted in a marked bloom of Liv and Lond. Thus, despite negative SGF cultures, residual Liv and Lond bacteria can undergo expansion in SIF when commensal microbes are absent. Even so, the AUC of L. monocytogenes was higher than Liv and Lond AUC in simulated gastrointestinal fluids (Figure 2b). Lseel and Lval demonstrated reduced growth in SGF and SIF compared to L. monocytogenes during the first 24 h. Lseel and Lval reached the stationary phase within 48 h of incubation in SIF medium at 37 °C and showed similar CFU counts to Lm, Liv, and Lond. Altogether, these data show that all the clones of the Listeria species tested grew exponentially in SIF without commensal bacteria, indicating that neither bile acids nor pancreatic enzymes provide bactericidal effects.
a) Growth curve, b) AUC, and c) % of cell survival of Lm, Liv, Lond, Lseel, and Lval in SGF and SIF (n = 6 replicates). Error bars represent the standard deviation. Different letters represent statistical differences (P < 0.05). ns stands for not significantly different.
Ex vivo competition with gut microbiota
To investigate the ruminant gut microbiota resistance against oral Listeria species infection, we used an ex vivo model, which has been previously shown to be a good model for intestinal infection (Becattini et al. 2017). Ex vivo experiments showed that commensals mediated Listeria species clearance at different proportions (Figure 3a–c). Gut microbiota inhibited Lm and Lseel CFUs by over two orders of magnitude within 72 h, reaching a final concentration of 4 Log_10_ CFU/mL (Figure 3a). In contrast, Lond and Lval were rendered undetectable by the gut microbiota within 24 h, and Liv within 72 h. Altogether, these results showed that gut commensals displayed less inhibitory activity against the tested isolate of Lm than against the isolates of Liv and Lond, although in all cases, they reduced viable CFUs over 72 h.
a) Growth curve, b) AUC, and c) % of cell survival of Lm, Liv, Lond, Lseel, and Lval in gut microbiota ex vivo (n = 6 replicates). Error bars represent the standard deviation. Different letters represent statistical differences (P< 0.05).
Ex vivo competition with uterine microbiota
As the literature concerning the culture of uterine microbiota from ruminants is lacking, we first validated the model used in this study. We successfully cultivated the ewe’s uterine microbiota to 9.0 and 11.8 Log_10_ CFU/mL (Supplementary Figure 1). We then compared the growth of Listeria species in AF medium with and without microbiota to evaluate whether the potential differences observed between species could be attributed to their specific ability in competing against the uterine microbiota (Figures 4a and b and 5a–d). In AF medium, all the strains reached maximum counts in the first 24h, with no difference between them. At the end of the experiment at 144 h, no statistical differences were found between the five Listeria species when grown in AF medium without microbiota (Figure 4a and b).
a) Growth curve and b) AUC of Lm, Liv, Lond, Lseel, and Lval in AF medium (n = 6 replicates). Error bars indicate the standard deviation.
Next, we cultivated the same five Listeria species in AF medium with uterine microbiota and observed different growth patterns (Figure 5d). As the only difference between these two experimental setups is the presence or absence of the microbiota, these results suggest that the presence of uterine commensals influences the growth of our five strains. Uterine microbiota restricted the growth of Listeria species compared to the AF medium alone (Figures 4a and b and 5a–d). In contact with uterine microbiota, nonpathogenic Lval and pathogenic Lm reached higher counts than the other species, reaching their highest concentration in 24 h (Figure 5a, c, and d). Although Lond and Lseel growth was slower, they reached similar values to Lm and Lval after 48h. Finally, although Liv presented the slowest growth, it was able to survive, multiply, and finally reach a concentration of 5.9 Log_10_ CFU/ml at 72 h (Figure 5a). In contrast with the previous results where the tested isolates of Liv, Lond, and Lval could not survive to gut microbiota exposure, here all could compete with the uterine microbiota and grow.
a) Growth curve, b) AUC, c) cell survival percentage, and d) growth rate of Lm, Liv, Lond, Lseel, and Lval in uterine microbiota ex vivo (n = 6 replicates). Error bars represent the standard deviation. In figures 5b and d, statistical differences are represented by three stars (P< 0.001). In Figure 5c), different letters represent statistical differences ( P< 0.05).
Listeria species growth in semen
The survival and growth of Listeria species cells in semen are illustrated in Figure 6. Semen exposure had variable inhibitory activity. Lval could not replicate in this environment and was undetectable during the experiment. Liv and Lseel CFUs decreased in the first 24 h and 48 h, respectively, although they finally grew to 10^3^ CFU/mL after 72 h. Lm and Lond constantly grew up to 72 h and expanded to 8.3 and 6 Log_10_ CFU/ml, respectively. Altogether, these data show that the isolates of pathogenic Listeria species can grow in semen.
a) Growth curve, b) AUC, and c) cell survival percentage of Lm, Liv, Lond, Lseel, and Lval grown in semen at 37 °C (n = 4 replicates). Error bars represent the standard deviation. Different letters represent a significant difference (P < 0.05).
Discussion
Studies about the ecological preferences of different Listeria species are necessary to understand their distribution in nature. Although Lm and Liv/Lond are pathogenic species, two remarkable differences exist between them: i) while Lm causes septicemia, encephalitis, and reproductive disorders in humans and ruminants, Liv/Lond only causes reproductive disorders in ruminants (Sergeant et al. 1991; Alexander et al. 1992; Gill et al. 1997; Vázquez-Boland et al. 2001); ii) Lm is significantly more abundant than Liv/Lond in hosts, soils and food (MacGowan et al. 1994; Hafner et al. 2021; Palacios-Gorba et al. 2021a). The present study helps to understand why Lm and Liv/Lond have distinct host niche preferences. Our results and previous studies suggest that establishing an asymptomatic fecal carriage plays a fundamental role in Listeria spp. ecology since survival to gastrointestinal conditions may favor host intestinal colonization, replication, and fecal shedding. Fecal excretion finally helps Listeria to transit between the host intestine, soil, and food (Nightingale et al. 2004; Hafner et al. 2021; Palacios-Gorba et al. 2021a). Extrapolating the present data to a species level, Lm and Lseel resistance against gastrointestinal fluids, as well as competitiveness against gut microbiota, appear to ensure their higher distribution in urban, ruminant farm, and natural environments than L. ivanovii (MacGowan et al. 1994; Sauders et al. 2012; Chapin et al. 2014; Linke et al. 2014; Orsi and Wiedmann 2016; Palacios-Gorba et al. 2021a; Liao et al. 2023). In contrast, our results demonstrate that bacteria inhabiting the intestine allow the inhibition of the tested clones of Liv and Lond growth. Possible mechanisms may include nutrient competition (Maltby et al. 2013), production of antibacterial molecules (Zhu et al. 2000; Corr et al. 2007; Lakshminarayanan et al. 2013; Vijayakumar and Muriana 2015; Egan et al. 2016; Saraoui et al. 2016), as well as contact-dependent inhibition (Ruhe et al. 2013). Altogether, presuming that the behavior of the tested isolates is representative of the species, our results show that acidic conditions of the digestive system and the intestinal microbiota provide the first-line defense against orally acquired Liv and Lond infection, preventing gut colonization, replication, and fecal shedding. The different environmental distribution of Lm versus Liv/Lond could also be due to differences in environmental persistence, transmission dynamics and host immune evasion. These latter hypotheses reserve further investigation.
On the contrary, the growth of Lond -and at a lower rate, Liv - in contact with the uterine microbiota indicates that the female reproductive tract may be a more permissive niche than the gastrointestinal tract. This niche preference could explain why Liv/Lond are associated with abortion outbreaks in ruminants (Macleod et al. 1974; Dennis 1975; Sergeant et al. 1991; Alexander et al. 1992; Gill et al. 1997; Chand and Sadana 1999; Şahi̇n and Beytut 2006).
The hypothesis of sexual transmission for Listeria has been previously discussed by other authors in humans as well as in other mammals (Osebold and Inouye 1954; Rappaport et al. 1960; Toaff et al. 1962; Gray 1963; Gray and Killinger 1966; McDonald 1967; Smith et al. 1967; Wiedmann et al. 1999). This hypothesis was based on the isolation of pathogenic Listeria from the genital tract of healthy humans and animals, as well as during abortion cases. Documented cases, such as the one described by Toaff et al. Rappaport et al. and Gray’s experiments in the 60’s showed the chronic presence of Listeria in the uterus, cervix, or vagina of aborted women and does, and in the semen of their interacting males. L. ivanovii (also called Lm serotype 5) was also suspected to be of venereal transmission in some sheep’s abortion cases (McDonald 1967; Smith et al. 1967; Alexander et al. 1992). As proposed by Lennon et al. (1984), more investigation on the possible venereal transmission of L. ivanovii and Lm should be done, especially considering that MacPherson and Fish (1954) demonstrated that pathogenic Listeria could survive in frozen bovine semen.
In conclusion, the present study contributes to the understanding of the different Listeria species’ preferences for distinct ecological niches and clinical manifestations of L. monocytogenes and L. ivanovii.
Supplementary Material
Supplementary Table 2.pdf
CEEA 25_17_QUEREDA_Informe CEEA.pdf
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alexander AV, Walker RL, Johnson BJ, Charlton BR, Woods LW. 1992. Bovine abortion attributable to Listeria ivanovii: four cases (1988-1990). J Am Vet Med Assoc. 200(5):711–714. 10.2460/javma.1992.200.05.7111568917 · doi ↗ · pubmed ↗
- 2Alvarez-Ordóñez A et al. 2015. Occurrence, persistence, and virulence potential of Listeria ivanovii in foods and food processing environments in the republic of ireland. Biomed Res Int. 2015:350526–350510. 10.1155/2015/35052626543856 PMC 4620271 · doi ↗ · pubmed ↗
- 3Bahadi MA et al. 2023. Group-training of rams at puberty for artificial vagina-mediated semen collection and its influence on semen quality and sexual behavior. Anim Reprod. 20(1):e 20220051. 10.1590/1984-3143-ar 2022-005137101423 PMC 10124154 · doi ↗ · pubmed ↗
- 4Barmpalia-Davis IM, Geornaras I, Kendall PA, Sofos JN. 2008. Differences in survival among 13 Listeria monocytogenes strains in a dynamic model of the stomach and small intestine. Appl Environ Microbiol. 74(17):5563–5567. 10.1128/AEM.00319-0818586963 PMC 2546618 · doi ↗ · pubmed ↗
- 5Becattini S et al. 2017. Commensal microbes provide first line defense against Listeria monocytogenes infection. J Exp Med. 214(7):1973–1989. 10.1084/jem.2017049528588016 PMC 5502438 · doi ↗ · pubmed ↗
- 6Benner M, Ferwerda G, Joosten I, van der Molen RG. 2018. How uterine microbiota might be responsible for a receptive, fertile endometrium. Hum Reprod Update. 24(4):393–415. 10.1093/humupd/dmy 01229668899 · doi ↗ · pubmed ↗
- 7Brown P et al. 2025. Listeria tempestatis sp. nov. and Listeria rocourtiae subsp. hofi subsp. nov. Int J Syst Evol Microbiol. 75(5):006774 10.1099/ijsem.0.006774 PMC 1237842940358009 · doi ↗ · pubmed ↗
- 8Cao X et al. 2019. Prevalence and characteristics of Listeria ivanovii Strains in wild rodents in China. Vector Borne Zoonotic Dis. 19(1):8–15. 10.1089/vbz.2018.231730570448 · doi ↗ · pubmed ↗