Antimicrobial Resistance in Selected Enterobacteriaceae from Broilers and Their Environment: ESBL, AmpC, Carbapenemases, Colistin, and Fluoroquinolone Resistance—A Systematic Review and Meta-Analysis
Julia von Kiparski, Nunzio Sarnino, Diana Vargas, Aleksandra Atanasova, Roswitha Merle

TL;DR
This study reviews antimicrobial resistance in bacteria from broiler chickens and their environment, highlighting high levels of resistance to several antibiotics.
Contribution
The study provides a comprehensive meta-analysis of AMR in Enterobacteriaceae from broilers and their environment across three continents.
Findings
ESBL-resistant E. coli was found in 41% of meat and 38% of fecal samples.
European meat samples showed significantly higher ESBL-producing E. coli rates than North African samples.
The study highlights the need for standardized surveillance systems to monitor AMR.
Abstract
Background/Objectives: Antimicrobial resistance (AMR) threatens global public health. This systematic review and meta-analysis, as part of the “ENVIRE” project (interventions to control the dynamics of antimicrobial resistance from chickens through the environment), assesses the prevalence of phenotypic and genotypic resistance, including extended-spectrum beta-lactamases (ESBLs), AmpC beta-lactamases, carbapenemases, colistin, and fluoroquinolone resistance, in broiler chickens and their environment. Methods: The analysis covers the years 2002–2022, focusing on Escherichia (E.) coli, Klebsiella spp., Enterobacter spp., and Citrobacter spp. in fecal, meat, environmental, and other-than-feces samples from observational studies published in PubMed and Web of Science. Quality assessment was performed using the Alberta Heritage Foundation criteria. Results: Data from 170 studies, conducted…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
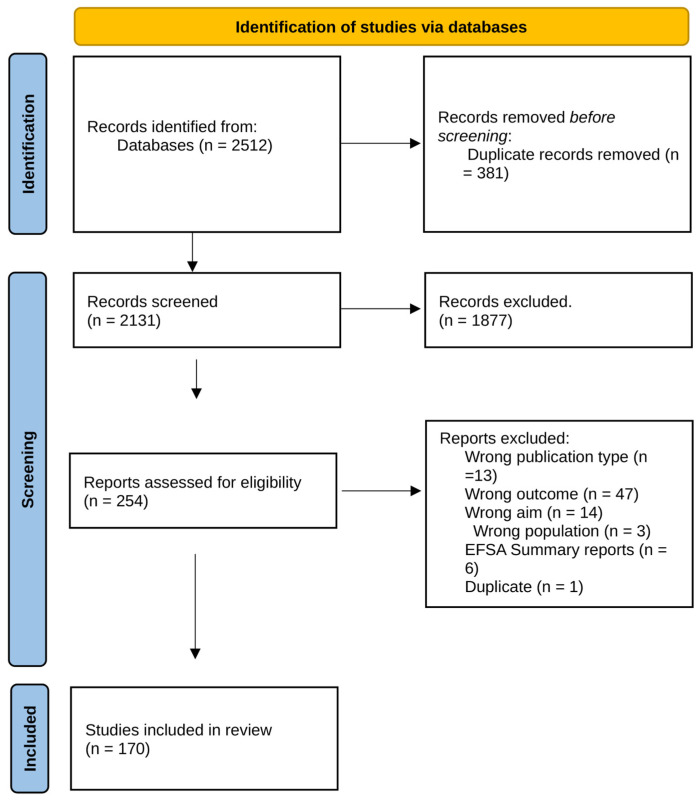 Figure 1
Figure 1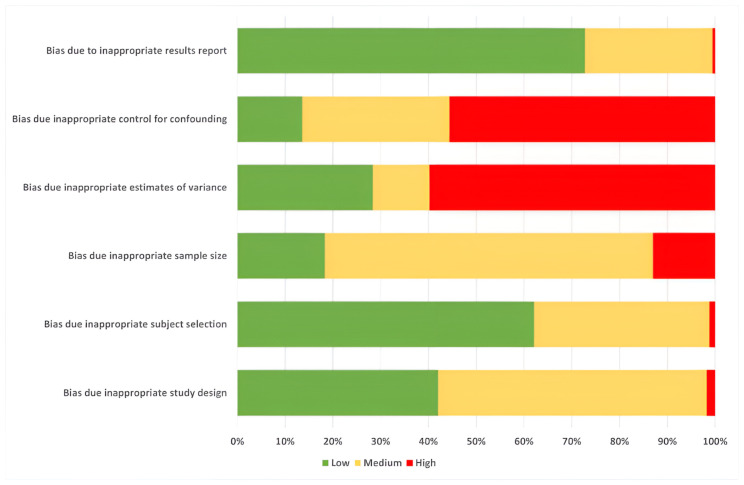 Figure 2
Figure 2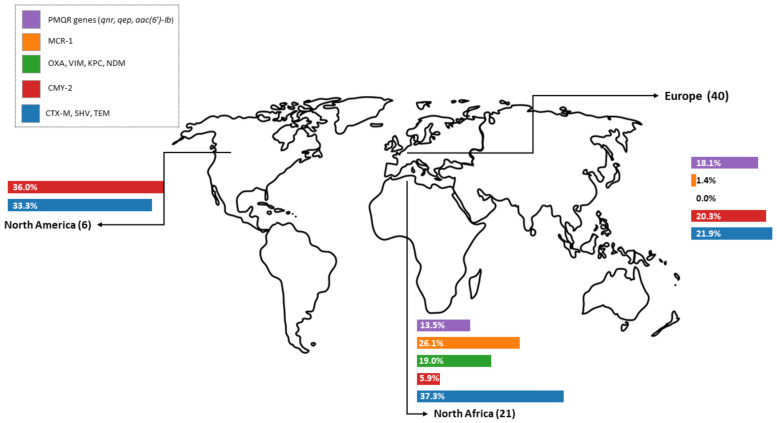 Figure 3
Figure 3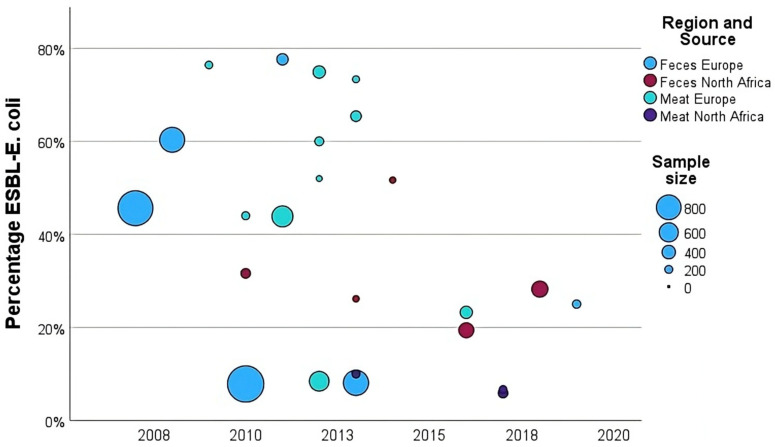 Figure 4
Figure 4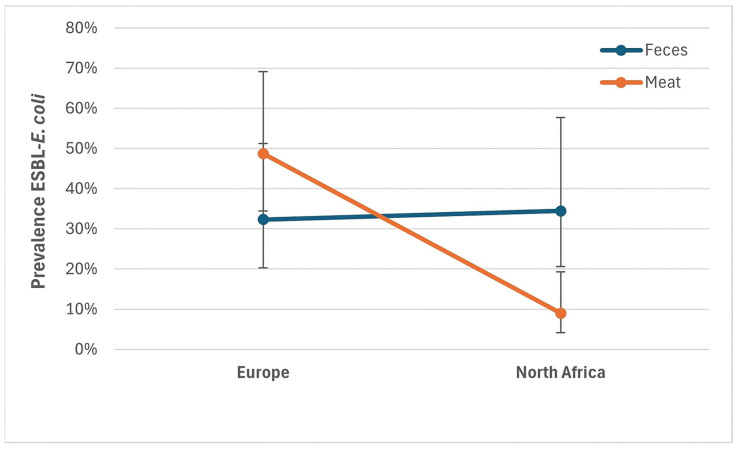 Figure 5
Figure 5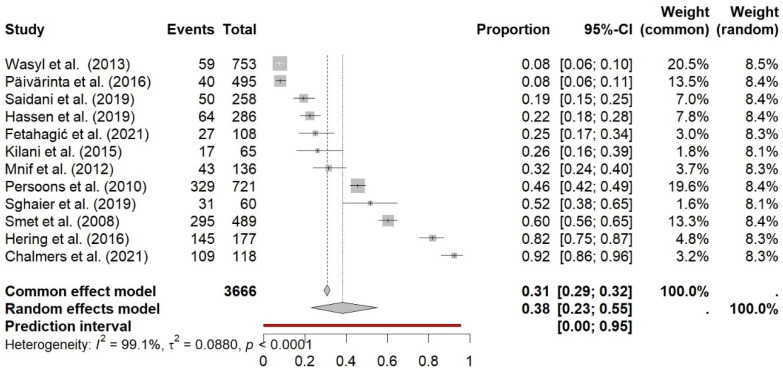 Figure 6
Figure 6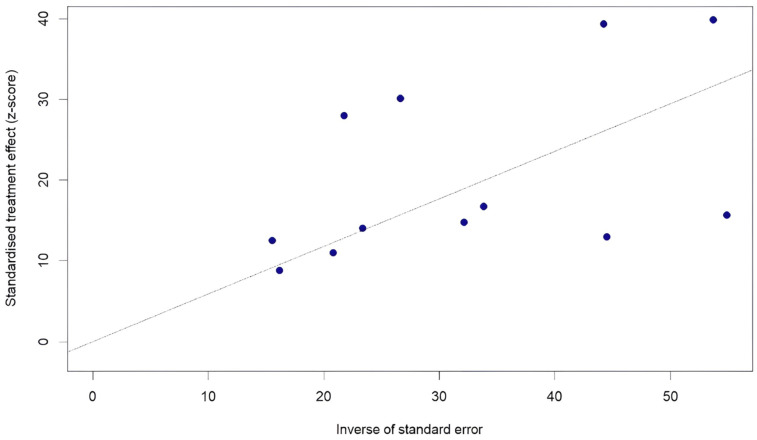 Figure 7
Figure 7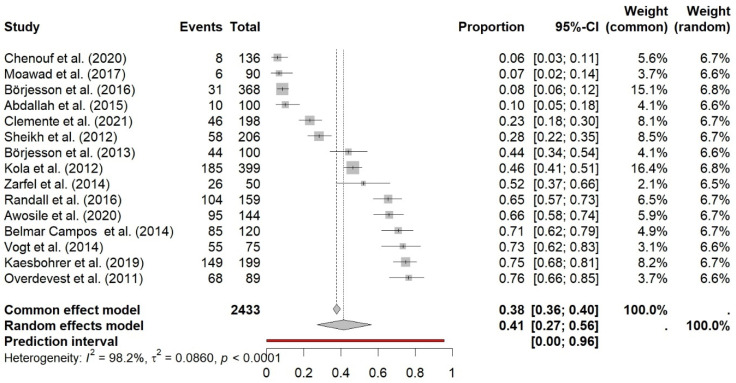 Figure 8
Figure 8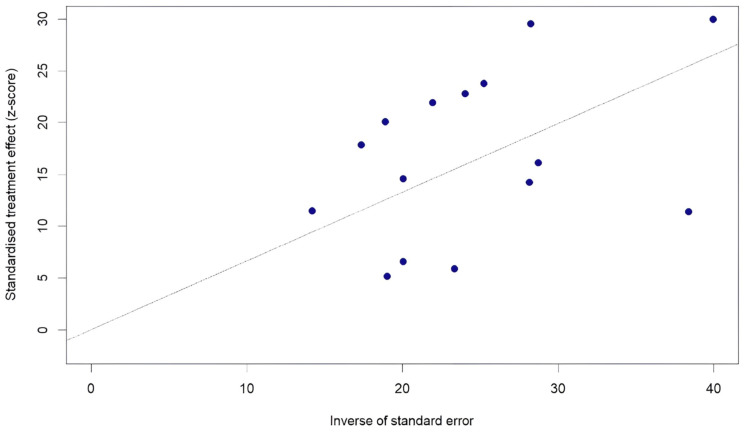 Figure 9
Figure 9| Author | DOI | Year | Country | Sample Type | Bacteria | n Samples (Isolates) | % Pheno-ESBL | % Pheno-AmpC | % Geno-ESBL | % Geno-AmpC | Denominator |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Maciuca et al. [ | 10.1089/mdr.2014.0248 | 2015 | Romania | cecal |
| 127 (90) | 70.87% | 37.79% | 70.87% | 37.79% | S |
| Reich et al. [ | 10.1016/j.fm.2015.10.020 | 2015 | Germany | neck skin/cecal |
| 1800 | neck skin: 93.30%, cecal 90.00% | S | |||
| Alba et al. [ | 10.3389/fmicb.2018.01217 | 2018 | Italy | cecal |
| 300 (300) | 81.33% | S, I | |||
| Messaili et al. [ | 10.12834/VetIt.799.3865.2 | 2019 | Algeria | intestine |
| 100 (100) | 73.00% | 70.00% | S, I | ||
| Boulianne et al. [ | n.a. | 2015 | Canada | cecal |
| 1500 (962) | 45.11% | I | |||
| Romero-Barrios et al. [ | 10.1089/fpd.2019.2776 | 2020 | Canada | carcasses |
| (1135) | 35.95% | I | |||
| Belmahdi et al. [ | 10.1016/j.jgar.2016.04.006 | 2016 | Algeria | cecal |
| 61 (61) | 32.79% | 80.00% | 20.00% | I | |
| Parker et al. [ | 10.1136/vr.103579 | 2016 | UK | cecal |
| 125 | 28.80% | 9.60% | 28.00% | 6.40% | S |
| Thorsteinsdottir et al. [ | 10.1111/j.1863-2378.2009.01256.x | 2008 | Iceland | cecal |
| 146 (185) | 0.00% | S | |||
| Randall et al. [ | 10.1093/jac/dkq396 | 2010 | UK | cecal |
| 388 | 4.38% | 3.61% | S | ||
| Dierikx et al. [ | 10.1016/j.vetmic.2010.03.019 | 2010 | The Netherlands | cecal |
| (153) | 14.38% | 95.45% | 22.73% | I | |
| Girlich et al. [ | 10.1128/AEM.02491-06 | 2007 | France | cecal |
| 112 | 10.71% | 10.71% | S | ||
| Päivärinta et al. [ | 10.1016/j.ijfoodmicro.2019.108361 | 2019 | Finland | cecal |
| 605 (605) | 18.00% | 0.83% | 18.84% | S |
| Author | DOI | Year | Country | Bacteria | n Samples (Isolates) | % Pheno-ESBL | % Pheno-AmpC | % Geno-ESBL | % Geno-AmpC | Denominator |
|---|---|---|---|---|---|---|---|---|---|---|
| Egea et al. [ | 10.1016/j.ijfoodmicro.2012.08.002 | 2012 | Spain |
| 15 | 93.33% | S | |||
| Ghodousi et al. [ | 10.1089/fpd.2015.1936 | 2015 | Italy |
| 163 (134) | 91.79% | 98.51% | 11.19% | I | |
| Casella et al. [ | 10.1016/j.ijfoodmicro.2017.07.005 | 2017 | France |
| 48 (77) | 91.67% | 4.05% | 100% | 3.90% | I |
| Clemente et al. [ | 10.3390/antibiotics10111333 | 2021 | Portugal |
| 198 (60) | 76.67% | 21.67% | 100% | 1.67% | I |
| Kaesbohrer et al. [ | 10.1016/j.vetmic.2019.03.025 | 2019 | Germany |
| 199 (192) | 74.9% | 71.87% | 26.04% | S | |
| Belmar Campos et al. [ | 10.1016/j.ijmm.2014.04.012 | 2014 | Germany | 120 (87) | 100% a | S | ||||
| Overdevest et al. [ | 10.3201/eid1707.110209 | 2011 | The Netherlands | 89 (71) | 83.15% a | S | ||||
| Vogt et al. [ | 10.1089/mdr.2013.0210 | 2014 | Switzerland |
| 75 (68) | 73.33% | 73.33% | 17.33% | S | |
| Randall et al. [ | 10.1016/j.ijfoodmicro.2016.10.036 | 2016 | UK |
| 159 | 65.41% | 65.41% | S | ||
| Zarfel et al. [ | 10.3390/ijerph111212582 | 2014 | Austria |
| 50 | 52.00% | 52.00% | S | ||
| Forward et al. [ | 10.1155/2004/695305 | 2004 | Canada |
| 75 (43) | 97.67% | 90.70% | 90.7% | I | |
| Awosile et al. [ | 10.1139/cjm-2020-0442 | 2020 | Canada |
| 144 | 65.28% | 61.81% | 21.53% | 26.39% | S |
| Mollenkopf et al. [ | 10.1089/fpd.2017.2390 | 2018 | USA |
| 497 | 11.10% | 77.80% | 17.51% | 12.88% | S |
| Author | DOI | Year | Country | Bacteria | n Samples (Isolates) | % Pheno-ESBL | % Pheno-AmpC | % Geno-ESBL | % Geno-AmpC | Denominator |
|---|---|---|---|---|---|---|---|---|---|---|
| Abdallah et al. [ | 10.1371/journal.pone.0136052 | 2015 | Egypt | KEC, | 100 (106) | 65.09% a | S | |||
| Sheikh et al. [ | 10.1089/fpd.2011.1078 | 2012 | Canada |
| 206 (193) | 28.15% | 75.13% | 25.24% | S | |
| Moawad et al. [ | 10.1186/s13099-017-0206-9 | 2017 | Egypt |
| 90 (15) | 40.00% | 66.67% | I | ||
| Kola et al. [ | 10.1093/jac/dks295 | 2012 | Germany | 399 | 46.30% a | I | ||||
| Hadžić-Hasanović et al. [ | 10.17392/1206-20 | 2020 | Bosnia and Herzegovina |
| 100 (64) | 29.00% | 24.00% | S | ||
| Randall et al. [ | 10.1111/jam.14687 | 2020 | UK |
| 622 | 2016: 45.00%, | 11.25% | 18.48% | S | |
| Börjesson et al. [ | 10.3201/eid2204.151142 | 2016 | Sweden |
| 368 (190) | 8.42% | 40.49% | S |
| Author | DOI | Year | Country | Bacteria | n Samples (Isolates) | % Pheno-ESBL (S/I) | % Pheno-AmpC (S/I) | % Geno-ESBL (S/I) | % Geno-AmpC (S/I) | Denominator |
|---|---|---|---|---|---|---|---|---|---|---|
| Ružauskas et al. [ | n.a. | 2010 | Lithuania |
| 240 (100) | 7% | S | |||
| Agersø et al. [ | 10.1093/jac/dkr507 | 2012 | Denmark |
| 153 | 3.3% | 35.75% | 47.83% | S | |
| Chenouf et al. [ | 10.1089/mdr.2020.0024 | 2020 | Algeria | 136 (78) | 9.56% a | S | ||||
| Mollenkopf et al. [ | 10.1089/fpd.2017.2390 | 2018 | USA |
| 497 | 11.1% | 77.8% | S |
| Author | DOI | Year | Country | Sample Type | Bacteria | n Samples (Isolates) | % Pheno-R | Denominator |
|---|---|---|---|---|---|---|---|---|
| Ramadan and Awad [ | 10.1186/s12941-016-0174-9 | 2016 | Egypt | samples other than feces |
| 400 (116) | 41.38% | I |
| Ceccarelli et al. [ | 10.1099/jmm.0.001176 | 2020 | The Netherlands | feces |
| (1811) | 62.23% | I |
| Gousia et al. [ | 10.1089=fpd.2010.0577 | 2011 | Greece | meat |
| 19 (8) | 62.50% | I |
| Hanon et al. [ | 10.1016/j.prevetmed.2015.09.001 | 2015 | Belgium | samples other than feces |
| (1132) | >50.00% | I |
| Kaesbohrer et al. [ | 10.1111/j.1863-2378.2011.01451.x | 2012 | Germany | meat, feces |
| (397) | meat: 53.10%, feces: 43.10% | I |
| Mainali et al. [ | 10.4315/0362-028X.JFP-13-203 | 2013 | Canada | samples other than feces |
| (600) | 0.00% | I |
| Ružauskas et al. [ | n.a. | 2010 | Lithuania | samples other than feces |
| 240 (100) | 47.00% | I |
| Zhao et al. [ | 10.1128/AEM.07522-11 | 2012 | USA | meat |
| (2494) | 0.00% | I |
| Manageiro et al. [ | 10.1016/j.ijfoodmicro.2017.10.007 | 2017 | Portugal | samples other than feces |
| 680 (202) | 90.60% | S, I |
| Chenouf et al. [ | 10.1089/mdr.2020.0024 | 2020 | Algeria | meat | 136 (78) | 60.20% a | I | |
| Thorsteinsdottir et al. [ | 10.1111/j.1863-2378.2009.01256.x | 2010 | Iceland | cecal, meat |
| 146 (185) | cecal: 18.20%, meat: 36.00% | I |
| Author | DOI | Year | Country | Sample Type | Bacteria | n Samples (Isolates) | % Pheno-R | Denominator |
|---|---|---|---|---|---|---|---|---|
| Casella et al. [ | 10.1016/j.ijfoodmicro.2017.07.005 | 2017 | France | meat |
| 48 (77) | 20.80% | I |
| Egea et al. [ | 10.1016/j.ijfoodmicro.2012.08.002 | 2012 | Spain | meat |
| 15 | 32.26% | I |
| Fetahagić et al. [ | 10.2478/aiht-2021-72-3560 | 2021 | Bosnia and Herzegovina | feces |
| 108 | 10.67% | I |
| Geser et al. [ | 10.1186/1746-6148-8-21 | 2012 | Switzerland | feces |
| 93 | 6.45% | I |
| Gregova et al. [ | 2012 | Slovakia | processing plant of a slaughterhouse |
| (48) | 43.00% | I | |
| Hricová et al. [ | 10.21101/cejph.a4328 | 2017 | Czech Republic | bedding |
| 126 (126) | 61.11% | S, I |
| Martínez-Álvarez et al. [ | 10.3390/antibiotics11040444 | 2022 | Spain | air, manure |
| 111 (111) | 38.70% | S, I |
| Musa et al. [ | 10.3390/ani10071215 | 2020 | Italy | cloacal, skin |
| (conventional 135, ABF 131, organic 140) | conventional 44.4%, ABF 20.6%, organic 23.6% | I |
| Zarfel et al. [ | 10.3390/ijerph111212582 | 2014 | Austria | meat |
| 50 | 0.00% | S |
| De Koster et al. [ | 10.3390/antibiotics10080945 | 2021 | Belgium, The Netherlands | feces |
| 779 | 84.81% | S |
| García-Béjar et al. [ | 10.3390/ani11113197 | 2021 | Spain | meat |
| 30 (240) | ca. 25.00% | I |
| Randall et al. [ | 10.1093/jac/dkq396 | 2010 | UK | cecal |
| 388 | 4.60% | I |
| Much et al. [ | 10.1016/j.prevetmed.2019.104755 | 2019 | Austria | cecal |
| 1031 (962) | 63.83% | I |
| Pesciaroli et al. [ | 10.1016/j.ijfoodmicro.2019.108391 | 2019 | Italy | cecal, different production systems |
| 855 (854) | conventional 67.7%, ABF 42.8%, organic 45.2% | I |
| Maciuca et al. [ | 10.1089/mdr.2014.0248 | 2015 | Romania | cecal |
| 127 (90) | 87.79% | I |
| Myrenås et al. [ | 10.1016/j.vetmic.2017.11.015 | 2017 | Norway, Sweden, Iceland | cecal, meat |
| (319) | 13.50% | I |
| Pavlickova et al. [ | 10.1080/03601234.2015.1011959 | 2014 | Czech Republic | meat |
| (75) | 16.00% | I |
| Persoons et al. [ | 10.1089/mdr.2009.0062 | 2010 | Belgium | cloacal, cecal, neck skin | 2249 (2076) | 14.98% a | I | |
| Vanni et al. [ | 10.3382/ps.2013-03627 | 2014 | Italy | feces, organ |
| (235) | 24.20% | I |
| Wasyl et al. [ | 10.3389/fmicb.2013.00221 | 2013 | Poland | cloacal |
| 753 (682) | 72.24% | S |
| Galler et al. [ | 10.3390/antibiotics10040466 | 2021 | Austria | intestine |
| 100 | 0.00% | S |
- —German Federal Ministry of Research and Education
- —Open Access funds of Freie Universität Berlin
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts · Antimicrobial agents and applications
1. Introduction
The global rise of antimicrobial resistance (AMR) has become one of the most pressing challenges in public health, threatening the effective treatment of infectious diseases in both human and veterinary medicine [1,2]. The prevalence of antimicrobial resistance in broiler production is a significant concern for public health, as recent studies on commercial broilers have shown that multidrug-resistant E. coli act as potential reservoirs along the food chain and pose a risk of transmission to humans [3,4]. A recent review from China has also described diverse antimicrobial determinants in E. coli from poultry production, further underlining the global relevance of AMR in poultry and other food-producing animals [5]. Among extended-spectrum beta-lactamase (ESBL), CTX-M variants are the most detected ones in humans and poultry [6,7]. In livestock, and particularly in poultry production, antibiotics are widely used to prevent and treat infections, particularly in intensive farming systems where large numbers of broilers are raised under confined conditions [8,9]. Chicken meat represents an affordable and globally consumed source of animal protein, driving the expansion of intensive production, which often relies heavily on antibiotics to maintain flock health [10].
This widespread use of antibiotics has raised concerns that broilers could serve as reservoirs of antimicrobial-resistant bacteria, potentially transmitting resistance to humans through food, direct contact, or environmental pathways [11]. Moreover, E. coli has been described as particularly capable of persisting in hosts and the environment due to virulence factors and biofilm formation, which promote adhesion and reduced susceptibility to antimicrobial treatment [12]. Despite the growing body of literature on AMR, the research landscape remains fragmented. Existing studies often have limited geographical coverage, inconsistent methodologies, and a lack of epidemiological focus, hindering the accurate assessment of resistance prevalence and comparability across regions [13]. These gaps hinder reliable prevalence estimates and complicate cross-study comparisons. Furthermore, low- and middle-income countries often face gaps in surveillance and regulatory frameworks, leaving critical resistance data underrepresented [14]. A systematic approach is needed to address these gaps and identify methodological challenges [13]. The project “ENVIRE” (interventions to control the dynamics of antimicrobial resistance from chickens through the environment), part of the European Transnational Programme JPIAMR-ACTION (Joint Programming Initiative on Antimicrobial Resistance, https://www.jpiamr.eu/), focuses on interventions to minimize the selection and dissemination of antimicrobial-resistant bacteria in broiler chickens and their surroundings, ultimately addressing risks to human health [15].
The systematic review and meta-analysis presented here, conducted as part of the ENVIRE project, aims to quantify and summarize the reported occurrence of phenotypic resistance patterns and resistance genes in Enterobacteriaceae, specifically Escherichia (E.) coli, Klebsiella spp., Enterobacter spp., and Citrobacter spp. (KEC), over the past two decades. It focuses on resistance mechanisms such as beta-lactam resistance mechanisms, including ESBLs and AmpC (Ampicillinase C) beta-lactamases. Colistin resistance, mediated by mcr-1 (mobilized colistin resistance gene 1), and fluoroquinolone resistance, including plasmid-mediated quinolone resistance (PMQR), are also addressed. Additionally, this study examines carbapenem resistance. Data were collected from broiler chickens, covering various sample types during the fattening period. By synthesizing data from feces and meat samples across multiple regions through a meta-analysis, this approach provides a quantitative assessment of resistance patterns and highlights critical knowledge gaps. Ultimately, this review contributes to a better understanding of the global dynamics of antimicrobial resistance.
2. Results
The systematic literature search revealed 2512 publications; 381 were duplicates and removed. Overall, 2131 were screened for eligibility during the title–abstract screening, and 1961 publications were excluded because they did not meet all of the inclusion criteria (e.g., obtained from hatcheries, parent stock, breeding stock, wrong population, wrong study type, geographical limit). Finally, 170 publications (6.7%) were considered for the systematic review (Figure 1).
Regarding publication date, 87% (n = 148/170) of the included publications were from 2012 to July 2022, and 12.9% (n = 22/170) were from 2002 to 2011. Overall, 66.4% (n = 113/170) of the studies were distributed over 25 European countries, followed by North African studies, amounting to 21.8% (n = 37/170), with 19 from Egypt, 9 from Algeria, and 9 from Tunisia. Lastly, 11.8% (n = 20/170) of the studies originated from North America (Canada, USA).
2.1. Quality Assessment
Overall, 85 publications were regarded as high quality, 55 were acceptable (n = 55), and 30 publications were of critical quality (minimum score 2). Their data are described in this review, but they were excluded from any meta-analysis. Figure 2 shows the distribution of scores of the most essential criteria for the quality assessment. Concerning the sample sizes, we recorded 22 articles with a score of 0 (“obviously inadequate”), 117 articles in which the “samples seemed small or there was no mention of power/sample size/effect size of interest”, and 31 articles in which the sample size seemed appropriate. Only 48 publications provided appropriate variance estimates. A sufficient control for confounding took place in 22 studies.
2.2. Beta-Lactam Resistance (ESBL- and AmpC-Producing Enterobacteriaceae)
Most studies reported on beta-lactam resistance (147 articles). A total of 100 publications were published from 2014 to 2022, but only 47 studies were published from 2004 to 2013. Samples from 99 studies were collected in European countries, 28 in North African countries, and 20 in North American countries.
2.2.1. Enterobacteriaceae—Prevalence in Feces
We identified 20 studies for a prevalence calculation that investigated fecal samples. Except for four studies—three from North Africa and one from Canada—all the studies originated from Europe. Overall, fecal samples predominantly showed high ESBL/AmpC prevalence in the European studies, with more variable results in North Africa.
Eight studies from Europe and one from North Africa reported high percentages of phenotypic resistant ESBL-Enterobacteriaceae (50–100%) in broiler production (Table 1). A subsequent study by Mesa et al. [16] reported the detection of various genetic variants, including 25.56% ESBL genes and 6.77% AmpC genes (Blanc et al. [17]). Five European studies reported moderate percentages of resistant bacteria (between 20% and 50%) among broiler isolates (Table 2). Another five studies reported a relatively low prevalence of resistance, all below 20% (Table 3).
2.2.2. Enterobacteriaceae—Prevalence in Slaughterhouses
A total of fifteen studies—two from North Africa (Algeria and Tunisia), two from Canada, and eleven from Europe—investigated samples other than feces (cecal, intestinal, or neck skin samples collected at slaughter). The prevalence data from 13 of these studies are presented in Table 4. Internal slaughterhouse samples (cecal/intestine) from Europe covered the full range from low to high ESBL/AmpC prevalences, but high prevalence categories (50–100%) were more frequently reported than in North Africa or Canada, where studies more often fell into low (<20%) or moderate ranges (20–50%), with only occasionally high-prevalence findings. The two studies that investigated neck skin or carcasses as external slaughterhouse samples showed a high prevalence in Germany on neck skin, in contrast to a moderate range in Canada [36,37]. Two studies could not be included in the quantitative analysis but still provided relevant trends: in their trend analysis from 2011 to 2014, Hanon et al. [38] reported a significant decreasing trend of cefotaxime resistance in 1132 chicken cecal samples in Belgium. Pacholewicz et al. [39] investigated 17 batches in two slaughterhouses in The Netherlands and reported a significant reduction in ESBL E. coli along the slaughter process.
2.2.3. Enterobacteriaceae—Prevalence in Meat
We summarized 24 studies concerning beta-lactam resistance in meat samples. Four studies each originated from North America and North Africa. The remaining 17 studies belonged to Europe.
High prevalence ranges were mostly observed in southern, central, and northwestern Europe, as well as in several North American studies, whereas moderate and low prevalences were more sporadically reported from all three regions. In detail, thirteen studies reported the prevalence of beta-lactam resistance between 50% and 100%, with ten of these from Europe, three from North America, and one from North Africa (Table 5). Notably, Mollenkopf et al. [40] reported 77.8% AmpC and only 11.1% ESBL phenotypes in E. coli from retail meat in the United States, highlighting a clear predominance of AmpC. Seven studies reported moderate levels of resistant Enterobacteriaceae between 20% and 50% (Table 6), while four studies reported lower prevalences of resistance (<20%; Table 7).
2.2.4. Clinical Samples
Across the ten studies that investigated isolates from diseased or clinically affected chicken broilers, resistance prevalences were highest in North Africa, while the studies from Europe and North America generally showed much lower levels, mostly below 20%. Notably, three studies from Egypt showed a higher prevalence of resistance, namely, 55% by Amer et al. [41], over 59% by Awad et al. [42], and 76.8% by Badr et al. [43], when analyzing strains from diseased animals with colibacillosis. Dhaouadi et al. [44] reported 30% resistant Enterobacteriaceae in Tunisia, while the remaining studies showed lower resistances (below 20%). Another study from Egypt demonstrated phenotypic ESBL and AmpC resistance in 18% of isolates [45]. In Canada, Varga et al. [46] found that 15% of isolates from chickens with suspected colibacillosis were resistant to ceftiofur. In the United States, Huang et al. [47] reported a ceftiofur resistance rate of 4.8%. In Germany, resistances ranged from 0.8% [48] to 4.9% [49]. Additionally, Niero et al. [50] reported genotypic evidence of ESBL/AmpC genes in 9% of broiler samples in Italy.
Table 7: ESBL and AmpC resistance in meat samples (<20%; four studies).
2.2.5. Comparison of Different Farming Conditions
In nine studies, prevalence was determined as part of comparisons between broiler groups on different farming conditions (antibiotic-free, organic, conventional) or medical treatment management (treated versus non-treated). Conventional broiler systems generally showed higher resistance levels than organic or antibiotic-free farms, although some studies still found substantial resistance in all production types. In particular, two studies detected high percentages of resistant Enterobacteriaceae (50–100%): In The Netherlands, Stuart et al. [84] compared 38 organic and 60 conventional retail chicken breast samples and found that the prevalence of ESBL-producing microorganisms was 100% (95% CI: 94–100%) in the conventional samples, compared to 84% (95% CI: 70–93%) in the organic samples (p value < 0.001). The distribution of ESBL genes in the conventional and organic samples was 42% vs. 56% for CTX-M-1 (not significant), respectively, 20% vs. 42% for TEM-52 (not significant), and 23% vs. 3% for SHV-12 (p < 0.001). Additionally, CTX-M-2 (7%), SHV-2 (5%), and TEM-20 (3%) were detected exclusively in the conventional samples. In Canada, Chalmers et al. [85] investigated fecal samples from six antibiotic-free farms (60 samples) and seven conventional farms (58 samples), finding extended-spectrum cephalosporin (ESC)-resistant E. coli in 90% of the antibiotic-free samples and 95% of the conventional samples. Overall, both groups showed similar resistance levels. Among these ESC-resistant E. coli, CTX-M was identified in 25% of the isolates from conventional farms and 11% from antibiotic-free farms. CMY was more frequent in 76% of the conventional isolates and 89% of the antibiotic-free isolates. Two studies by Musa et al., both originating from Italy and comparing samples from antibiotic-free, organic, and conventional farms, showed results with prevalences of resistance between 20 and 50%. One investigated cloacal, environmental, cecal, and skin samples from farms and slaughterhouses [86], where cefotaxime resistance was detected in 36.2% of the samples from antibiotic-free farms, 29.3% from organic farms, and 51.7% from conventional farms. The odds of resistance were significantly lower on organic and antibiotic-free farms than on conventional farms and significantly higher in the samples collected at slaughterhouses than in those taken on the farms. The second study by Musa et al. [87] investigated cloacal swabs and skin samples from a slaughterhouse. Cefotaxime resistance was found in 23.7% of samples from antibiotic-free farms, 24.3% from organic farms, and 43.7% from conventional farms, with a statistically significant difference between farm types (p = 0.001). No significant difference was observed between the cloacal swabs and skin samples (p = 0.520).
Another two studies resulted in lower percentages of resistant isolates (<20%). Much et al. [88] compared 962 cecal isolates of organic and conventional farms in Austria (2010–2016) and revealed 2.0% ESBL producers in the organic samples and 1.9% in the conventional samples. Pesciaroli et al. [89] collected cecal samples from antibiotic-free, organic, and conventional farms, with prevalences of ESBL-producing Enterobacteriaceae of 4.8% in the antibiotic-free samples, 1.8% in the organic samples, and 3.4% in the conventional samples. Among these, 76% carried CTX-M-15, 12% TEM, and 8% SHV genes.
Three additional studies focused on different objectives. Voets et al. [90] compared plasmid-mediated Ampicillinase C beta-lactamase (pAmpC) genes between chicken meat and human clinical isolates in The Netherlands, finding that 12% of the meat isolates carried pAmpC, specifically CMY-2. Baron et al. [91] analyzed fecal samples from French broilers treated with third-generation cephalosporins and non-treated broilers, finding that the treated group had a significantly higher phenotypic prevalence of ESBL-producing bacteria than the non-treated group. Genotypic analysis revealed the presence of CTX-M in 14.3% of the isolates, TEM in 4.9%, and CMY-2 in 5.6%. In an experimental study in Canada, Merchant et al. [92] investigated the effect of feed supplementation on E. coli in litter across six different diets. The results showed significant treatment effects, with ceftiofur resistance ranging from 11.1% to 44.6%.
2.2.6. Wastewater
Among the four European studies on poultry slaughterhouse wastewater, the extractable data were limited but indicated substantial ESBL resistance, ranging from a moderate range in Spanish wastewater to very high cefotaxime in German process and wastewater samples. In particular, Sabaté et al. [93] analyzed four wastewater samples in Spain and found that 21% were ESBL-producing. Facciola et al. [94] investigated wastewater from 18 Italian slaughterhouses, identifying beta-lactam resistance in E. coli, Citrobacter spp., and Enterobacter spp. without specifying the number of poultry-related samples.
A comprehensive study by Savin et al. [95] in Germany examined the process and wastewater from two slaughterhouses. Among 186 E. coli isolates, 99% in slaughterhouse 1 and 86% in slaughterhouse 2 were resistant to cefotaxime. Additionally, over 90% of the 71 isolates from the KEC group were cefotaxime-resistant. Two further publications from the study included smaller datasets and are only briefly mentioned [96,97]. Homeier-Bachmann et al. [98] studied wastewater from three poultry slaughterhouses in Germany, but no extractable results were available.
2.2.7. Studies Investigating Only Klebsiella spp.
Across the five studies focused on resistant Klebsiella spp., ESBL-positive strains were very common, and several studies represented nearly all recovered isolates, even though only one study aimed to determine prevalence. Specifically, Ramadan and Awad [99] aimed to calculate prevalence and found 48.8% ESBL-producing Klebsiella spp., 84.6% carried blaTEM, and 30.8% blaCTX-M. Daehre et al. [100] did not aim to determine prevalence, but were able to isolate 41 Klebsiella spp. strains from various stages of the broiler production chain, all of which carried the ESBL gene blaSHV-2. One study by Savin et al. [101,102] examined process and wastewater, identifying 63 PCR-confirmed ESBL-producing Klebsiella spp., representing 88.8% of the isolates. Two studies from Egypt reported the presence of beta-lactamase-producing Klebsiella spp. Elmonir et al. [103] investigated diseased chickens and detected 19 PCR-confirmed resistant Klebsiella spp. out of 160 organ and environmental samples. Abdallah et al. [74] found that all Klebsiella spp. isolates from retail meat were ESBL producers.
2.2.8. Other Targets
Four studies reported genotypic findings following PCR confirmation of only ESBL-positive isolates, showing moderate to high carriage of ESBL genes. In Canada, Ghosh et al. [104] identified ESBL-positive E. coli in 12.7% of broiler flocks, with all positive isolates carrying CTX-M-1. In Tunisia, 24% of cloacal samples from five farms contained ESC-resistant Enterobacteriaceae, with considerable variation between farms (ranging from 4% to 67%). Among these, 98% harbored CTX-M, 10% carried SHV-12, and 2% CMY-2 [105]. While another study by Hassen et al. [106] in Tunisia showed that 31.5% of isolates harbored ESBL genes. In Germany, Irrgang et al. [107] reported that 25.9% of 567 E. coli isolates from broiler production carried resistance genes, with 22.2% testing positive for SHV.
Across nine studies with other primary objectives, ESBL/AmpC-producing E. coli were repeatedly detected in various environmental compartments around broiler production (air, slurry, surfaces, flies, manure) and even in farm workers. In more detail, in Slovakia, Gregova et al. [108] examined bioaerosols and surface swabs in a poultry slaughterhouse and found that 43% were ESBL-producing E. coli. Among 19 selected isolates, 16 carried CMY-2, while no CTX-M was detected. In Spain, Solá-Ginés et al. [109] collected 682 flies from five poultry farms and detected ESBL-producing E. coli in 6.2%. Among the 42 positive isolates, 54.8% carried CTX-M-1, 43% CTX-M-14, and 2.4% CTX-M-9. Additionally, 78.6% carried TEM; SHV and CMY-2 were undetected. In Germany, Laube et al. [110] collected samples from the environment in seven broiler farms. Slurry samples were positive in 12 of 14 cases, and 28.8% of the surrounding boot swabs contained resistant E. coli. Three of the forty air samples outside the barns tested positive. Of 51 ESBL/AmpC-producing E. coli, 82% carried ESBL or CMY. Genetic analysis suggested transmission of resistant strains from the barn to the surrounding environment. In The Netherlands, Huijbers et al. [111] monitored cloacal and environmental samples from a broiler flock raised without antibiotics. A susceptible–infectious–susceptible (SIS) model demonstrated a rapid increase in ESBL-E. coli prevalence, reaching nearly 100% by day three. Martínez-Álvarez et al. [112] sampled manure and indoor air on a broiler farm in Spain, where 20.7% ESBL-producing E. coli were PCR-confirmed with SHV-12. Wadepohl et al. [113] studied stool samples from chicken farm workers and found that 5.1% carried ESBL-producing Enterobacteriaceae.
A total of 25 additional studies were reviewed and are presented in the Supplementary Material (Table S2). However, as these studies were assessed to be of lower methodological quality or lacked sufficient data, their findings were not considered in the main results section.
2.3. Carbapenem Resistance
Fifty-four studies investigated carbapenem resistance in the period from 2009 to 2022. Three studies came from North America, and eighteen were from North Africa. Samples from 33 studies were collected in Europe. Carbapenem resistance in broiler-related Enterobacteriaceae was overall very rare, with only a few findings in meat from Egypt, Algeria, and Spain.
Eleven studies analyzed samples from retail meat sources and rarely detected carbapenem-resistant bacteria. For instance, carbapenem-resistant Klebsiella spp. were found in chicken carcasses from Egyptian butcheries (26.1% phenotypically carbapenem-resistant Klebsiella spp. [74]) and in chicken meat from Algeria (16.0%). In Spain, a study reported that approximately 70% of E. coli isolates from chicken meat were carbapenem-resistant [114]. In contrast, several studies from North Africa and Europe did not detect carbapenem-resistant E. coli in meat samples (Randall et al. [70], Zarfel et al. [71], Chenouf et al. [83], Ramadan et al. [115], Gousia et al. [116], Pavlickova et al. [117]). Two additional investigations (Ojer-Usoz et al. [118], Sgariglia et al. [119]) addressed the same question but did not provide conclusive resistance data. Six of the seven studies that investigated samples collected from slaughterhouses did not show evidence of carbapenem resistance [46,51,55,61,65,78,89]. Concerning the studies that investigated fecal samples, only one study out of eight detected carbapenem-resistant E. coli (1.8%) in Egypt [32]. Regarding samples from diseased animals—mainly from North Africa—carbapenem-resistant Klebsiella spp. were found in two Egyptian studies (Elmonir et al. [103] with 40%; Hamza et al. [120] with 43%), while no such resistant pathogens were identified in the remaining three studies [43,45,121]. Comparison of conventional, organic, and antibiotic-free chicken production samples revealed only carbapenem-susceptible isolates [87,88]. Three studies investigated carbapenemases in wastewater; in two of these, no resistance was detected [95,98]. In the third, Facciola et al. [94] examined samples from various species; no carbapenem resistance was observed. Similarly, thirteen studies that focused exclusively on ESBL-positive isolates did not detect carbapenem resistance [24,27,31,48,63,64,67,69,73,105,122,123,124]. Two studies were excluded from the main results due to insufficient methodological quality [125,126].
2.4. Colistin Resistance
A total of 48 studies investigated colistin resistance. All of the studies were published within the timeframe of 2014 to 2022. Most of them (n = 32) were published between 2019 and 2022, while sixteen studies were published between 2014 and 2018. The majority of the studies were published in European countries (n = 30). The remaining studies originated from countries in North Africa (Egypt, Tunisia, Algeria).
2.4.1. Prevalence Studies
Ten studies focused on determining prevalence, all of which reported prevalences of resistance below 20%. In more detail, five studies from different European countries were all conducted within a similar timeframe. These studies focused primarily on cecal or fecal samples and reported comparable percentages of resistant Enterobacteriaceae, consistently ranging between 1% and 5% [38,52,127,128,129]. In contrast, three separate studies, one from the UK and two from Algeria, reported negative results. Randall et al. [79] analyzed meat samples over a comparable period and did not detect colistin-resistant isolates. Similarly, Agabou et al. [126] and Messaili et al. [53], who investigated fecal and intestinal samples, also found no positive cases.
Moawad et al. conducted two studies in Egypt: In 2016, they analyzed 576 cloacal swab samples from poultry flocks, isolating 72 Enterobacteriaceae strains, of which 63 were E. coli. Five isolates (7.9%) were phenotypically resistant to colistin and carried the mcr-1 gene [32]. In contrast, their 2017 study on 90 chicken meat samples yielded 15 E. coli isolates, all susceptible to colistin [76].
Two European studies by Casella et al. [64] and Päivärinta et al. [61] investigated meat samples, whereby both found no colistin resistance phenotypically and genotypically.
Three studies examined colistin resistance using cecal and intestinal swabs collected at slaughterhouses, none of which found any colistin resistance. Belmahdi et al. [55] tested 61 intestinal swabs in Algeria in 2014 and found that all isolates were susceptible to colistin. Similarly, Much et al. [88] reported no colistin resistance among 962 cecal isolates from Austria in 2010–2016. Likewise, Päivärinta et al. [61] investigated cecal and meat samples in Finland in 2015 but detected no colistin resistance genes.
2.4.2. Comparison of Different Farming Conditions
Five studies compared resistance patterns between different treatment groups in healthy animals. Musa et al. [87] (compared conventional, organic, and antibiotic-free farms) and Much et al. [88] (investigated organic and conventional farms) found no resistant isolates, while Pesciaroli et al. [89] (samples from conventional, organic, and antibiotic-free farms) and Majewski et al. [130] (who investigated colistin-treated and non-treated fecal samples) reported a higher prevalence of resistance on conventional farms than in organic or antibiotic-free systems. Additionally, Ribeiro et al. [131] demonstrated that the number of PCR-positive Enterobacteriaceae decreased as the withdrawal period from colistin use increased.
2.4.3. Clinical Samples
Six studies examined colistin resistance in diseased broiler chickens, yielding a lower prevalence of resistance (<20%) in four studies (two from Europe and North Africa). However, two studies found higher resistance levels (between 20 and 40%) in North Africa. More precisely, Mesa-Varona et al. [49] observed a colistin resistance of 5.7% among clinical E. coli isolates from cecal samples in Germany. In Poland, Majewski et al. [132] documented 15.3% colistin-resistant E. coli isolates and 15.5% in Klebsiella spp. isolates collected from various organs of affected birds. In Algeria, Halfaoui et al. [121] examined organ samples from 180 diseased chickens diagnosed with colibacillosis and identified 6.54% colistin-resistant E. coli. In Egypt, Elmonir et al. [103] examined 160 samples (100 organ and 60 environmental samples) and isolated 1.25% resistant Klebsiella spp. In contrast, Dhaouadi et al. [44] found 24% colistin-resistant isolates in broilers with colibacillosis from three farms, with more than half of the resistant isolates carrying the mcr-1 gene. Even higher levels of resistant E. coli were reported by Badr et al. [133], who found that 41.1% of isolates from pooled organ samples across 120 farms were resistant, with all isolates carrying the mcr-1 gene.
2.4.4. Wastewater
Results from wastewater investigations were published in three studies conducted in Germany, where colistin-resistant E. coli and Klebsiella spp. were repeatedly found in wastewater from poultry slaughterhouses. In more detail, two separate studies by Savin et al. focused on poultry slaughterhouses. The first study [95] showed 9.8% phenotypic resistance to colistin, and PCR analysis confirmed resistance genes in 72%. Additionally, the study investigated isolates of a KEC group (Klebsiella spp., Enterobacter spp., Citrobacter spp.), detecting colistin resistance in 14.1%, with PCR performed on a subset of resistant Klebsiella spp. isolates, of which 50% were confirmed. The second study by Savin et al. [101], using samples from the same timeframe and locations, specifically analyzed Klebsiella spp. isolates and reported higher percentages of colistin resistance: 32.4% exhibited phenotypic colistin resistance. A third German study by Homeier-Bachmann et al. [98] investigated wastewater from three additional poultry slaughterhouses. Although the study reported colistin-resistant E. coli isolates, the lack of sufficient detail in the study design prevented the reliable extraction of specific prevalence data.
2.4.5. Other Targets
Six studies examined only ESBL-positive isolates for colistin resistance, whereas five studies—three from Europe and two from North Africa—reported low prevalences of resistance (<20%). One Tunisian study detected higher percentages pheno- and genotypically. In detail, Vogt et al. [58] could not detect any colistin resistance in meat in Switzerland, followed by Kluytmans-van den Bergh et al. [134], who found 1.1% phenotypically resistant bacteria and 1.6% harboring mcr-1 in meat samples in The Netherlands. Balázs et al. [28] detected 2.6% colistin-resistant ESBLs in feces from Hungary. The North African studies showed a 100% susceptibility in Algeria [126], compared to Saidani et al. [105], who reported 10% resistant E. coli and 0% in Klebsiella spp. In Tunisia, Hassen et al. [106] analyzed fecal and meat samples, detecting positivity of 53.1% and 66.7%, respectively, with 96.6% of the positive isolates carrying the mcr-1 gene.
Four studies (three European and one North African) reported colistin resistance as an additional finding, although their primary focus was on other resistance patterns or targets, showing very low or no colistin resistance. Myrenås et al. [135] investigated colistin and cephalosporin resistance in cecal and meat samples and detected only 0.3% colistin-resistant E. coli isolates in Scandinavia. In Portugal, Manageiro et al. [124] analyzed ESBL-E. coli in cecal samples from poultry slaughterhouses and identified 3% reduced susceptibility to colistin. Schwaiger et al. [136] analyzed 438 isolates obtained from fecal samples of six broiler chicken flocks in Germany and reported a decrease in colistin resistance from 4.8% in week three to 0.9% in week five. Chaalal et al. [137], who studied meat samples in Algeria primarily for carbapenemase-producing bacteria, detected no colistin-resistant isolates at all.
Eight studies that reported colistin resistance were reviewed and are presented in the Supplementary Material (Table S4). However, their findings were not included in the main Results Section due to lower methodological quality or incomplete data reporting.
2.5. Fluoroquinolone Resistance
Ninety-two studies investigated fluoroquinolone resistance, which were published in the period from 2006 to 2022 (fifty-three studies from 2016 onward). In 57 studies, samples were collected from Europe, 24 from North Africa (Egypt, Algeria, and Tunisia), and 11 from North America.
2.5.1. Prevalence Studies
Eleven studies aimed at determining the phenotypic prevalence of ciprofloxacin- and enrofloxacin-resistant Enterobacteriaceae, with seven from Europe, two from North Africa, and two from North America (Table 8). Overall, the Europe and Norboulth Africa studies showed moderate to high prevalences in fecal, cecal, liver, and meat samples, whereas an Iceland study presented lower levels, and the North American studies detected no resistance in comparable samples.
2.5.2. Studies with Non-Prevalence Primary Objectives
Most studies (65 studies) focused on aims other than prevalence estimation, examining ciprofloxacin- and enrofloxacin-resistant Enterobacteriaceae across Europe, North America, and North Africa.
Europe
Across Europe, fluoroquinolone resistance in broiler-related Enterobacteriaceae in 21 studies was highly variable (Table 9), ranging from absent or low levels in some countries and different sample types to very high resistance levels in others (e.g., Poland and Romania), with several studies indicating higher resistance in conventional compared with organic or antibiotic-free (ABF) production systems.
North America
Fluoroquinolone resistance in seven North American studies was absent or extremely low. E.g., no resistance was found in two studies that investigated fecal samples [33,54], 1% was reported by Vounba et al. [138], and 0.28% was reported by Varga et al. [139]. Meat samples from Canada showed similar results, with no observed ciprofloxacin resistance [75]; three ciprofloxacin-resistant isolates reported by Awosile et al. [73], without mentioning the number of analyzed isolates; and just 0.2% in carcass samples [37].
North Africa
Six North African studies reported consistently high fluoroquinolone resistances in several sample types from Egypt and Algeria besides variable results from Tunisia. In particular, 43.3% of ciprofloxacin-resistant E. coli out of 120 isolates of meat samples could be observed in Egypt [115]. No resistance was found in one Tunisian study that investigated fecal samples [140], but another Tunisian study by Abbassi et al. [31] found 32.5% ciprofloxacin-resistant E. coli in fecal samples, analyzing only multi-resistant isolates. An Algerian study showed higher prevalences of resistance, with 89% in ciprofloxacin in fecal samples [141]. Hamed et al. [142] observed 71% in organ samples and up to 58.6% in cloacal, skin, and environmental samples. Similar to the findings of Hamed et al. [142], another Egyptian study revealed 72.9% ciprofloxacin-resistant Klebsiella spp. in organ and environmental samples [103].
Wastewater
Three studies investigated wastewater from poultry slaughterhouses. Two of these studies originated from Germany, reporting 53% ciprofloxacin-resistant E. coli and 15.5% ciprofloxacin-resistant Klebsiella spp./Enterobacter spp./Citrobacter spp. (KEC) in one study [98] and 1.1% ciprofloxacin-resistant E. coli in the second German study [98]. Sabaté et al. [93] found a ciprofloxacin resistance of 56% in E. coli isolates from Spain.
Clinical Samples
Eight studies investigated isolates of diseased broilers (mostly colibacillosis), whereby the studies from Europe and the USA reported very low to modest levels of fluoroquinolone resistance, whereas the North African studies consistently showed very high prevalences. In detail, one study from Germany detected 20% of 20 ESBL-positive isolates as enrofloxacin resistant [48]. Huang et al. [47] revealed 3.4% enrofloxacin-resistant E. coli from clinically ill animals out of laboratories in several US states. In North Africa, two Egyptian studies showed 82% ciprofloxacin-resistant E. coli [45] and 75% enrofloxacin-resistant E. coli [83]. Another Egyptian study investigated clinical and non-clinical isolates (100% enrofloxacin-resistant non-clinical E. coli) [143]. Dhaouadi et al. [44] analyzed 100 organ samples from colibacillosus-diseased animals and found 68% fluoroquinolone-resistant E. coli. The highest prevalence of enrofloxacin resistance was observed in Algeria, at 86.3% [121]. Badr et al. [43] found 66.1% phenotypic ciprofloxacin resistance in organ samples.
Other Targets
Across thirteen studies restricted to ESBL-Enterobacteriaceae, fluoroquinolone resistance varied widely in Europe, ranging from very low to about 50%. In contrast, the North African studies showed very high levels. In detail, Vogt et al. [69] found a low prevalence of 1.4% in ESBL-positive isolates from retail chicken meat in Switzerland. Kola et al. [77] and Belmar Campos et al. [67] reported 7.6% and 23% ciprofloxacin resistance among ESBL-producing Enterobacteriaceae from chicken meat in Germany. Smet et al. [22] found 8.7% enrofloxacin resistance in cloacal samples from broilers on five Belgian farms. Stuart et al. [84] reported 14% ESBL-producing E. coli. Blanc et al. [17] found 31.2% ciprofloxacin resistance among 192 ESBL-producing isolates from fecal samples in Spain, while Machado et al. [144] observed 50% resistance among 14 ESBL-producing Klebsiella spp. from meat and feces in Portugal; however, both of these studies were assessed as having poor methodological quality. Among the North African studies, Saidani et al. [105] and Hassen et al. [106] reported similarly high levels of resistance, with 86.0% of cloacal samples and 86.1% of ESBL-producing isolates from meat samples resistant to enrofloxacin or ciprofloxacin in Tunisia, respectively.
2.5.3. Comparison of Phenotypic and Genotypic Fluoroquinolone-Resistant Enterobacteriaceae
The findings revealed significant differences between the samples and phenotypic and genotypic prevalence rates.
In Europe, phenotypic and genotypic resistances were generally lower than in North Africa, though certain sample types exhibited higher resistances, particularly in bedding, skin, fecal, and manure samples (Table 10). Only one study from Italy reported similarly high phenotypic and genotypic resistance.
In North Africa, the studies consistently indicated higher levels of fluoroquinolone resistance (Table 11), often in the moderate to very high range. Genotypic confirmation was often lower than phenotypic resistance.
2.5.4. PCR-Confirmed Fluoroquinolone-Resistant Enterobacteriaceae
Two studies conducted only genotypic detection (qnr, qep, aac(6′)-Ib) or reported only genotypic results. In the study by Savin et al. [102], water samples from two poultry slaughterhouses in Germany were tested for resistance in Klebsiella spp., including phenotypic detection. Although absolute numbers for phenotypic resistance were not provided in the publication, 3 out of 71 Klebsiella isolates were identified as carriers of PMQR genes (4%). Niero et al. [50] examined lesion swabs from broilers with colisepticemia and found PMQR genes in 4 of 98 E. coli isolates (4%).
2.6. Regional and Overall Occurrence of Genotypic Resistance Patterns
Across all regions, 67 studies (40 studies from Europe, 21 from North Africa, and only six from North America) provided data on resistance genes at the isolate level. Out of these, 49 articles reported ESBL resistance proteins (CTX-M, SHV, and TEM), with an overall occurrence of 24.7% (n = 2277/9210) positive isolates, while at the same time, 26 studies investigated the AmpC resistance protein CMY-2, including 3701 investigated isolates, which resulted in an overall prevalence of 26.6% (n = 616/3701). Six studies investigated carbapenem resistance (OXA, VIM, KPC, and NDM), with an overall resistance of 4.4% (n = 32/730). Seventeen articles performed molecular detection of colistin resistance to mcr-1 in environmental and other sample types and found 4% (n = 258/8674) of isolates to be positive. Ten studies tested 974 isolates for PMQR proteins (qnr, qep, aac(6′)-Ib) and yielded an overall prevalence of 17.3% (n = 168/974).
As shown in Figure 3, the highest occurrence of beta-lactam resistance genes was observed in North Africa, where 37.3% of isolates carried CTX-M, SHV, or TEM across 16 studies involving different sample types. In contrast, North America reported a higher prevalence of AmpC resistances (CMY-2), with 36% across six studies. Notably, carbapenem resistance genes were primarily reported from North Africa, where five studies identified them in 19% of Enterobacteriaceae, while one European study detected no carbapenemase genes. A similar pattern was seen for colistin resistance genes, with 26.1% in North Africa (six studies), no reports from North America, and only 1.4% in Europe (11 studies). Regarding PMQR proteins (qnr, qep, aac(6′)-Ib), six European studies found a prevalence of 18.1%, compared to 13.5% reported in four studies from North Africa.
2.7. Prevalence in Meat and Feces
2.7.1. Negative Binomial Regression Model
To explore how resistance patterns varied over time, between regions, and by sample types, Figure 4 shows the ESBL-E. coli prevalence reported in the studies included in the analyses of prevalence data. Studies reporting high prevalences mainly involved meat samples from Europe, before 2015, whereas studies from North Africa showed lower prevalences.
The regression model revealed that region and sample type had a significant influence on prevalence (global p-values: region p = 0.0118; sample type p = 0.0914; interaction p = 0.0030), while prevalence declined over time (IRR 0.93), but this effect was not statistically significant (p = 0.1228, Table 12). While there was no difference between Europe and North Africa in feces samples, meat samples in Europe had a significantly higher prevalence than in North Africa (p = 0.0029, Figure 5).
2.7.2. Meta-Analyses
The pooled prevalence of twelve publications about ESBL-resistant E. coli in fecal samples was estimated at 38% (95% CI 23–55%; I^2^ = 99.1%) (Figure 6). Neither the sample size nor the publication year seemed to impact prevalence. Six studies originated from Europe, five from North Africa, and one from North America. Between-study variability was high (between-study variance τ2 = 0.088, p < 0.0001), which corresponds to the Galbraith plot (Figure 7).
Concerning the meat samples, the pooled prevalence was 41% (95% CI 27–56%; I^2^ = 98.2%) (Figure 8). The Galbraith plot also displays a large variability between the studies (Figure 9), which was statistically significant (τ2 = 0.086, p < 0.0001). Most studies originated from Europe (ten), followed by three from North Africa and two from North America.
3. Discussion
3.1. General
Our review included 6.7% of the originally selected articles. The data from 170 studies enabled us to calculate and compare the occurrence of phenotypic resistance and resistance genes in beta-lactams (ESBL and AmpC), colistin, carbapenemases, and fluoroquinolones in certain Enterobacteriaceae over a period of 21 years and three global regions. To our knowledge, this is the first study to use a systematic review and meta-analyses of prevalence data on ESBL-E. coli in fecal and meat samples, as well as the occurrence of resistance genes. In recent years, several reviews on specific resistances or on specific sample types were published (e.g., [160,161,162,163,164]), but no review covered the whole range of bacteria and resistance patterns covered in our publication.
As expected, this review comprised numerous studies including different sample types, different laboratory methods, articles from various countries, and a wide period of published articles. Corresponding to our results, Luz et al. [2] reported an increase in the number of publications on antimicrobial resistance of 450% between 1997 and 2018, reflecting the increased awareness of this topic. The most frequently used sample type in this review was fecal samples because they are easy to obtain and contain Enterobacteriaceae, which were our focus and represent the One Health aspect.
The detection method was a crucial factor contributing to the heterogeneity between studies. As mentioned by Silley et al. [165] in 2011, it is essential to distinguish between clinical and epidemiological cut-offs and to take both into account even in the context of monitoring. Clinical breakpoints guide antimicrobial therapy, but an isolate with reduced susceptibility may have sufficiently low minimum inhibitory concentration (MIC) to ensure clinical therapeutic success. Therefore, isolates with low susceptibility to an antimicrobial substance should be distinguished from those with clinical resistance.
A notable observation is the number of studies on the topic, yet very few focus on prevalence. Many studies appear to prioritize methods designed to maximize the detection of isolates rather than accurately assessing prevalence. Consequently, the methodologies employed in these studies are not well-suited for providing reliable prevalence estimates. The limited data availability in many countries can often be attributed to resource constraints, the absence of active surveillance programs, or a lack of transparency surrounding antibiotic use in livestock industries, further hindering a comprehensive understanding of resistance patterns. The lack of focus on epidemiological aspects is evident in many studies, as insufficient information was provided on sample sizes or the number of positive samples. In most cases, this limitation significantly hindered our ability to reliably calculate percentages of resistant bacteria, reflected in poor quality assessment scores and limited comparability across studies.
3.2. Beta-Lactam Resistance (ESBL- and AmpC-Producing Enterobacteriaceae)
Most studies focused on beta-lactam resistance, with a rise in publications after 2014, reflecting the increased attention to AMR. In our review, both fecal samples collected on farms and meat samples often showed high resistance patterns, with several studies reporting values between 50% and 100%. According to the latest European Union (EU) summary report, the presumptive prevalence of ESBL-/AmpC-producing E. coli in broilers and broiler meat was 34.9% and 29.2% at the Member state-group level in 2022 [166]. These surveillance data suggest lower average prevalences of resistance than many of the included studies, highlighting the importance of harmonized monitoring to provide representative estimates. However, when looking at individual countries, the EU data revealed heterogeneity, ranging from 3.4% in Denmark to 61.3% in Hungary [166]. This variation mirrors the broad range of resistance levels observed across the included studies in our review. This suggests that national differences in antimicrobial usage, biosecurity measures, and slaughterhouse hygiene may contribute to this variation.
In contrast, the results from slaughterhouse samples were more heterogeneous: half of the included studies reported resistance levels below 20%, while the other half ranged up to 100%, suggesting that factors such as sampling context, cross-contamination, and methodological differences may play a role. Samples obtained at slaughterhouses or from broiler meat are subject to additional influences, including cross-contamination during scalding and defeathering [167]. Foyle et al. [162] reported a median of 75% Gram-negative isolates resistant to cefuroxime in wastewater in 27 studies, which is higher than the results in our review. Regarding non-E. coli ESBLs, ESBL-producing Enterobacter spp. were most often detected in German wastewater, consistent with their ability to colonize the gastrointestinal tract. As other authors have described, clonally related strains were detected in the parent flock and later in the fattening flock, indicating pseudo-vertical transmission and/or cross-contamination [95,100].
Studies on diseased animals reported lower resistance rates than those in healthy broilers. One explanation could be that these studies were more targeted, with smaller sample sizes or restricted to specific clinical conditions, which may not represent the true extent of resistance in the general population.
Our review also highlights the importance of phenotypic testing and molecular methods. While many studies reported resistance based on phenotypic detection methods, fewer provided detailed genotypic data. The EU summary report emphasized that molecular characterization is required to determine whether transferable genes encoding extended-spectrum cephalosporin resistance are present [166]. Such information is essential to assess the potential for horizontal gene transfer and the dissemination of resistance patterns.
Overall, the findings highlight the persistence of ESBL- and AmpC-producing Enterobacteriaceae throughout broiler production and support the importance of coordinated monitoring strategies. Given the potential for transmission to humans through meat consumption and environmental exposure, reducing beta-lactam resistance in poultry remains a crucial target for global One Health approaches.
3.3. Carbapenem Resistance
The limited number of studies on carbapenem resistance highlights a significant gap in research on this critical antibiotic class. Fifty-four studies were identified, with a notable geographic bias: thirty-three were conducted in Europe, eighteen in North Africa, and just three in North America. This uneven distribution suggests an underrepresentation of data from many regions, notably where carbapenem use may differ significantly. Carbapenem resistance was rarely detected in chicken-associated Enterobacteriaceae, yet isolated studies reported alarmingly high percentages of carbapenem-resistance, particularly in Klebsiella spp. from North Africa and in one European study from Spain. While these findings may indicate true resistance hotspots, they could also reflect methodological differences or limited sample sizes. This highlights the need for harmonized surveillance systems to distinguish sporadic detections from emerging regional trends reliably. These findings may reflect differences in antibiotic usage practices. In North Africa, unregulated or excessive use of carbapenems in both human and veterinary medicine could drive the emergence and spread of resistance. The higher prevalence of resistance observed in North Africa may also be linked to inadequate surveillance systems, weak regulatory enforcement, and a lack of standardized guidelines for antimicrobial use [168]. This contrasts with Europe and the USA, where stricter antibiotic stewardship programs and bans on certain antibiotics exist. Our findings on carbapenem-resistant Enterobacteriaceae (CRE) align with those of Köck et al. [163], who also reported CRE in Algeria. In contrast, their review included two extensive European studies indicating that CRE were not highly prevalent in livestock. This supports the notion of significant regional differences in CRE prevalence, likely influenced by antibiotic usage patterns and regulatory practices. In addition, carbapenem resistance has also been reported in other bacterial species and production settings, such as carbapenem-resistant Acinetobacter baumanii in raw milk from dairy farm animals, highlighting that multiple livestock sectors may contribute to its emergence and spread [169]. The limited data, regional disparities in prevalence, and varying detection methods underscore the need for more comprehensive and standardized research on carbapenem resistance.
3.4. Colistin Resistance
Research on colistin resistance in broiler production has intensified in recent years; the first study included in this review was conducted in 2014. This timing matches the global discovery of the transferable mcr-1 gene, which led to more monitoring [170]. Interestingly, we could not include studies on colistin resistance from the USA and Canada. One reason might be that the oral administration of colistin in livestock was never authorized in these countries [160]. In contrast, in North Africa, colistin is used widely even without a prescription [137], which is reflected by the high prevalence of colistin resistance at the phenotypic and molecular levels in our review. In Europe, the overall occurrence of colistin resistance was relatively low, reflecting the restricted or minimal veterinary use of colistin [161]. Some exceptions exist, such as the study from Poland that reported resistance rates around 15.8% [130], which may be linked to more frequent usage in that country. The studies by Badr et al. [133] and Dhaouadi et al. [44] suggest that diseased animals have higher resistance rates than healthy ones. The studies that compared production systems indicated that conventional farms sometimes showed higher resistance than organic or antibiotic-free systems [86,88,89,139]. While the number of such studies is limited, these observations indicate that farming practices and antibiotic usage can shape the occurrence and dissemination of colistin resistance [170].
3.5. Fluoroquinolone Resistance
Fluoroquinolones were classified as “highest priority critically important” antibiotics in human medicine besides third- and fourth-generation cephalosporins, macrolides, glycopeptides, and polymyxins [171]. Research on fluoroquinolone resistance in broilers shows an increase in the number of publications since 2016. Roth et al. [172] reported a rise in ciprofloxacin resistance in Spain from 17% in 2001 to 91% in 2016, potentially linked to the continued use of enrofloxacin in poultry, as its use is not entirely prohibited under EU directives [145].
In Europe, fluoroquinolones remain approved for treating bacterial infections in poultry, despite the 2006 ban on antibiotic growth promoters. In contrast, the Food and Drug Administration banned enrofloxacin use in the US as early as 2005 [172]. These policy differences may explain the significant disparities in resistance patterns between regions. North American studies consistently reported low or no resistance, whereas the prevalence in Europe reached up to 90%. This is consistent with EU monitoring data, which reported very high median ciprofloxacin and nalidixic acid resistance in broilers and high resistances in turkeys [167]. In North Africa, resistance data varied widely, which may reflect inconsistent antimicrobial use and surveillance systems in the region. Factors such as insufficient regulatory enforcement, lack of standardized treatment guidelines, and inadequate border controls exacerbate the problem [173]. Our findings indicate substantial variability in resistant isolates, with high numbers observed in the studies from Egypt and no resistance reported in the studies from Tunisia.
The prevalence of resistance observed in the studies should be viewed critically due to differences in detection methods and the number of isolates tested. Prevalences also showed wide variation across sample types. Notably, Italian meat samples [63] and Egyptian organ samples [42] showed close alignment between phenotypic and genotypic resistance, suggesting that PMQR genes might significantly contribute to phenotypic resistance in certain sample types. These differences may be shaped by alternative mechanisms (e.g., efflux pumps, target mutations) or environmental factors. In North Africa, high phenotypic resistance appears frequently even without associated PMQR genes, implying that non-genotypic factors could play a substantial role.
3.6. Regional and Overall Occurrence (Genotypic)
The fact that so many studies have been conducted in North African countries, in particular, may be because the prevalence of resistance is so high there. One of the reasons for this might be that even critically important substances are still used there as growth promoters [173]. Our genotypic findings underline this pattern, with the highest prevalence of ESBLs, carbapenemases, and colistin resistance genes reported from North Africa, in contrast to lower levels in Europe and North America, represented by only a few studies. Overall, ESBL and AmpC genes were the most frequently detected resistance determinants (approximately 25–27% of isolates), carbapenemase and colistin genes remained rare (around 4% each), and PMQR genes accounted for 17%. Notably, North Africa showed consistently higher prevalence across multiple gene classes, including ESBLs, carbapenemases, and colistin genes. At the same time, AmpC (CMY-2) was most frequent in North America and Europe, showing moderate ESBL/AmpC but more PMQR genes. However, these results should not be interpreted as representative prevalence estimates for each region, but rather as a synthesis of the available study data, which may be influenced by sampling strategies, isolate selection, and publication bias.
3.7. Analyses of Prevalence Data
As shown in Figure 4, most European studies before 2015 reported high levels of ESBL-producing E. coli in broiler fecal and meat samples, frequently exceeding 40–60%. After 2015, fewer studies were available, and the reported resistance appeared more heterogeneous, with some dropping below 20%. This shift may reflect the impact of EU measures restricting the off-label use of extended-spectrum cephalosporins in poultry production in 2012 [174]. Our regression model revealed that region and sample type significantly influenced the prevalence of ESBL-producing E. coli, while no consistent temporal decline could be confirmed. This shows that regional and methodological differences may substantially affect more than time trends. Notably, a higher prevalence was observed in European meat samples than in North African meat samples, while no regional differences were observed in fecal samples. Here, the possibility of publication bias must be considered (studies with higher resistance levels are more likely to be published), which plays a role in the variability [175].
Meta-Analyses
Through the meta-analysis of meat samples, the highest estimated ESBL-E. coli prevalence was 41%. The meta-analysis revealed that the average prevalence in meat samples was slightly higher than in fecal samples (38%). As reported by Projahn et al. [167], the effect of specific intervention steps in a slaughterhouse cannot prevent the spread of Enterobacteriaceae between carcasses and thus leads to an increase in prevalence. It has also been described several times that the structure of chicken skin has an influence on the adhesion of bacteria and that firmly attached bacteria are more difficult to remove during the plucking process [176]. However, the extremely high I^2^ values in both meta-analyses indicate considerable heterogeneity between studies and limit the interpretability of these pooled estimates. Our analysis of prevalence data, combined with Figure 4 and the generalized linear regression model, as described above, provides more nuanced insights into how prevalence varies over time, by region, and by sample type than the pooled estimates alone.
Only studies analyzing one colony per sample from healthy animals without preselection for multidrug resistance were included for fecal samples to avoid overestimating prevalence. Studies with more than one colony per sample were also considered for meat samples, because otherwise, too few studies would have been available. Nevertheless, analyzing only one colony per sample is most suitable for epidemiological prevalence studies, while testing multiple colonies can be helpful in laboratory diagnostics to show a broader picture of resistance. The lack of studies applying such epidemiological criteria suggests a possible publication bias, focusing on reporting higher resistance levels instead of representative prevalence estimates. In this context, classical statistical tests for publication bias, such as funnel plots and Egger’s test, are designed for comparative effect measures. In our single-group, prevalence-based meta-analyses with only a few studies and considerable heterogeneity, the results would be difficult to interpret and potentially misleading.
It was also surprising that even current studies reported high resistance levels, although the EU summary report on antimicrobial resistance shows a continuous decline i ESBL-E. coli prevalence in all European countries for many years [166], which may at least partly be explained by publication bias. Apart from that, the variance between the study results in both meta-analyses was surprising, considering the strict inclusion criteria of the studies. Publications in peer-reviewed journals focus on objectives other than representative prevalence estimates. This is a pity because national monitoring systems are not always available in English, and thus (except for the EU summary report), comparability of results from different nations is limited. We recommend publishing the results of (national) monitoring systems in peer-reviewed journals, thus making them available to the international research community.
For instance, the 2013 study by Koga et al. showed a prevalence of 65.4% in chicken carcasses [177] from Brazil, the world’s largest exporter of chicken meat and second-largest producer. This is similar to our results, but there is a lack of further studies in this area from Brazil. The fact that neither country, year, nor sample size affected the prevalence in feces and meat samples was surprising. Despite thorough inclusion criteria concerning high-quality scores and the study objective, homogeneity in the study results could not be achieved. The interpretation of study results should therefore be performed carefully. One possible explanation is that many studies covered the inclusion criteria but did not focus on prevalence. This leads us to conclude that the methods are designed to find many isolates, but this does not result in a reliable prevalence statement.
3.8. Conclusions
In conclusion, this systematic review and meta-analysis revealed substantial variation in prevalence and underscores the need for standardized surveillance systems and robust study designs. In addition, potential drivers, such as climate change and its impact on AMR in broiler production systems, should be considered. Future studies should adopt a clear epidemiological focus and comprehensive reporting of all baseline data, including sample sizes and the number of positive samples. This would allow for accurate prevalence calculations and improve the overall quality and comparability of research in this field.
4. Materials and Methods
This systematic review followed the guidelines of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) [178] and was registered in PROSPERO (International Prospective Register of Systematic Reviews) under the registration number CRD42024423479. The review question was “What is the extent of antimicrobial resistance in poultry farms and their environment?”
4.1. Inclusion and Exclusion
We included observational studies that reported the occurrence of antimicrobial resistance in broiler chicken production. We focused on resistance caused by beta-lactamases (ESBLs and AmpC) and carbapenemases, as well as colistin resistance and fluoroquinolone resistance in E. coli, Klebsiella spp., Enterobacter spp., and Citrobacter spp. The initial search string included Shigella spp., but no articles that examined Shigella spp. met the inclusion criteria. Therefore, Shigella spp. was not considered further in this review. This study was limited to publications between January 2002 and July 2022 and included studies from Europe, defined geographically, North America (USA and Canada), and North Africa. Accordingly, we excluded studies with samples from hatcheries and hatching eggs and samples from parent birds, breeders, and laying hens. Only peer-reviewed original research articles were included, while short communications, reviews, and other articles were excluded.
4.2. Search Strategy
PubMed (National Library of Medicine, Bethesda, MD, USA) and Web of Science (Clarivate Analytics, Philadelphia, PA, USA) were searched for studies published within the mentioned time frame. The literature search in the databases was carried out on the 4th of August 2022. The databases were searched using the following search strategy: “((chicken or poultry or broiler) AND (“E. coli” or Escherichia or Enterobacter or Klebsiella or Shigella or Citrobacter) AND (ESBL or AmpC or pAmpC or “plasmid-mediated AmpC β-lactamase” or “plasmid mediated AmpC β-lactamase” or “plasmid-mediated AmpC beta lactamase” or “plasmid mediated AmpC beta lactamase” or “plasmid mediated AmpC beta lactamase” or “extended-spectrum β lactamase” or “extended-spectrum β-lactamase” or “extended-spectrum beta lactamase” or “extended spectrum beta lactamase” or “colistin -resistance” or “colistin resistance” or “mcr-1” or mcr or “fluoroquinolone resistance” or “Fluoroquinolone-resistance” or “quinolone resistance” or “quinolone-resistance” or “carbapenem resistance” or carbapenemase or “carbapenemase producing” or PMQR or “Plasmid-mediated quinolone resistance” or “Antibiotic resistance” or “Antimicrobial resistance” or AMR or susceptibility or “Multi-drug resistance” or ABR or “Inhibitory effect” or “Multidrug resistance” or ARB or “Antimicrobial resistance bacteria” or MDR))”.
4.3. Study Selection
The software Rayyan (Rayyan Systems Inc., Doha, Qatar) was used for the screening process. For the study selection process, three reviewers (JK, DV, AA) screened the publications found through the title and abstract search strategy, following the inclusion criteria in Table 13. Possible duplicates were removed after manual confirmation. After the first screening, the reviewers’ decisions were unblinded. Reviewer disagreements were resolved by reaching a consensus.
4.4. Data Extraction from Eligible Studies
The publications that met the inclusion criteria by screening the titles and abstracts were collected in Citavi (Swiss Academic Software GmbH, Wädenswil, Switzerland). The data were extracted from the eligible studies selected by full-text reading and recorded in Microsoft Excel^®^ (Microsoft Corporation, Redmond, WA, USA). Full-text reading to assess eligibility and data extraction was conducted jointly by two reviewers (JK, RM). Studies were considered as eligible if the aim of the study was fulfilled by testing relevant chicken sample types (fecal, meat, environmental, or other) for the resistance types of interest (ESBL, AmpC, carbapenemases, colistin, fluoroquinolone) and reporting phenotypic and/or genotypic outcomes. Only observational studies were eligible.
Automation tools were not used, and contacting the study authors was not necessary. We collected general data concerning the continent and country from which the samples were collected. The samples were categorized as described above, and the bacteria and resistance patterns were noted. We recorded the laboratory methods and documented whether diseased or healthy animals were sampled.
The outcomes were the number of samples per sample type, the number of isolates, the number of phenotypically resistant isolates, the number of PCR-confirmed isolates, and the number of different detected genes. We considered the following genes: blaCTX-M, blaSHV, and blaTEM for ESBL, blaCMY-2 for AmpC, blaOXA, blaKPC, blaNDM, and blaVIM for carbapenemases, mcr-1 for colistin resistance, and qnr, qep, and aac(6′)-Ib for PMQR.
4.5. Data Analysis
For all resistance types, the resistance patterns reported in the original publications were extracted when available; if not explicitly stated, the percentage of resistant bacteria was calculated based on the number of tested samples and, if given, the number of isolates, according to the study design. When genotypic results were reported, they were included alongside phenotypic findings. Calculations could not be performed if the data were unavailable or multiple sample types were included in the results and not presented separately. Such studies were excluded from the data analysis but were still mentioned. Supplementary Tables S1–S6 summarize the relevant studies for each resistance type included in this review and the studies that are mentioned but whose results were not reported due to poor methodological quality. These tables provide details on study characteristics (year, country, sample type, bacterial species, number of samples, and isolates) and phenotypic and genotypic resistance patterns.
The studies with similar objectives were grouped (e.g., sample type, aim of prevalence determination, study design, diseased animals). The results were further stratified within each region (Europe, North America, North Africa) into resistance level categories of high (50–100%), moderate (20–50%), or low (<20%). For fluoroquinolone resistance, the studies were analyzed in three groups: those reporting only phenotypic results, those presenting phenotypic and genotypic results in comparison, and those reporting exclusively genotypic results.
4.6. Regional and Overall Occurrence of Genotypic Resistance Patterns
Regional and overall occurrence of genotypic resistance patterns were calculated using only those studies that reported both the total number of isolates tested for specific resistance genes and the number of positive detections. Supplementary Table S5 summarizes the included studies. The studies were assigned to three geographic regions (Europe, North Africa, North America) for each resistance type: beta-lactam (ESBL and AmpC), carbapenemases, colistin resistance, and plasmid-mediated quinolone resistance (PMQR). A base world map was obtained from the Pixabay image repository (https://pixabay.com, image ID 160811, accessed on 11 August 2025) and edited in Microsoft PowerPoint^®^ (Microsoft Corporation, Redmond, WA, USA) to highlight the study regions and to insert genotypic resistance charts.
4.7. Analyses of Prevalence Data
To compare study results between regions and over time, we selected studies that investigated ESBL-resistant E. coli isolates in two sample types. Studies reporting on fecal or cloacal samples were included if they (1) investigated only one colony per sample, (2) examined only healthy animals, (3) had a minimum sample size of 50, and (4) did not restrict their analysis to multi-resistant isolates. The inclusion criteria for the analysis of meat samples were analogous; however, due to the limited number of eligible studies, those that examined more than one colony per sample were also included. This resulted in 12 (fecal) and 15 (meat) studies (Figure 6 and Figure 8). A figure to display the raw prevalences per study, taking the sample size into account, was constructed using IBM SPSS Statistics (Version 29.0, IBM Corp., Armonk, NY, USA).
We used the statistical program SAS 9.4 (SAS Institute Inc. 2013, Cary, NC, USA) to build a generalized linear regression model with a negative binomial distribution and a logit link using the studies from Europe (n = 16) or North Africa (n = 8). The year of sampling, region, and sample type, including the interaction between region and sample type, were used as independent variables, and the prevalence of ESBL-resistant E. coli (number of resistant samples out of all samples) was the dependent variable. The level of significance was set to 0.05. Incidence risk ratios (IRRs) were also calculated, and model diagnostics included visual inspection of residuals for normality and homoscedasticity. Cook’s distance and leverage were used to investigate outliers. To illustrate the interaction between region and sample type, a figure displaying the estimates and 95% confidence intervals (95% CI) was created using MS Excel^®^ (Microsoft Corporation, Redmond, WA, USA). North America was excluded because only three studies fulfilled the criteria.
4.8. Meta-Analyses
Since we assumed that the distribution of prevalences across the studies was heterogeneous, we used random-effects models to determine a pooled prevalence estimate, including a 95% CI using the Clopper–Pearson method for individual studies. The results are presented as forest plots displaying each study’s point estimates and confidence intervals, including summary and confidence intervals.
Heterogeneity was assessed by Cochran’s Q test and the inverse variance index I2. The I^2^ was 99.1% for feces and 98.2% for meat, indicating a very high level of heterogeneity in the studies (p < 0.0001 in both models). Due to the small number of studies included in the analyses, we limited our evaluation of heterogeneity to a Galbraith plot. Analyses and visualization were carried out using the meta package in R (version 4.3.1) [179].
4.9. Quality Assessment
Three people independently assessed quality (JK, RM, NS). We used the checklist for assessing the quality of quantitative studies based on the Standard Quality Assessment Criteria for evaluating primary research publications from a variety of fields from the Alberta Heritage Foundation for Medical Research [180].
Scores from 0 to 2 were given to each of the 11 questions concerning the sufficient description of the objectives; the appropriateness of the study design; the method and subject characteristics; whether the outcome and measures were well defined and robust to errors; the appropriateness of the samples size; whether analysis was described and appropriate; if estimates of variances in the main results were reported; if confounding was controlled; if the results were reported in a sufficient detail; and if the results supported the conclusion. The scores were summed to a total score ranging from 0 to 18. Articles with values of 8 to 11 were reassessed. The studies were divided into three groups depending on the total score (low risk of bias, medium risk of bias, and high risk of bias), and these were displayed using a bias plot in R. Publications with a score > 11 were regarded as high-quality and marked as low risk of bias in the bias plot. Publications were regarded as acceptable with scores of between 8 and 11 and shown as medium risk of bias, while those with scores of less than 8 were assessed as critical and displayed as high risk of bias in the bias plot. The studies grouped into these three categories (low, medium, or high risk of bias) were displayed in a bias plot generated with robvis (version 0.3.0; R Foundation for Statistical Computing, Vienna, Austria; DOI: 10.32614/CRAN.package.robvis). The approach addressed the quality of individual studies. No additional method, such as GRADE (Grading of Recommendations Assessment, Development and Evaluation), was applied to assess certainty in the body of evidence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ferri M. Ranucci E. Romagnoli P. Giaccone V. Antimicrobial resistance: A global emerging threat to public health systems Crit. Rev. Food Sci. Nutr.2017572857287610.1080/10408398.2015.107719226464037 · doi ↗ · pubmed ↗
- 2Luz C.F. van Niekerk J.M. Keizer J. Jong N.B. Braakman-Jansen L. Stein A. Sinha B. van Gemert-Pijnen J. Glasner C. Mapping Twenty Years of Antimicrobial Resistance Research Trends Artif. Intell. Med.202212310221610.1016/j.artmed.2021.10221634998519 · doi ↗ · pubmed ↗
- 3Abreu R. Semedo-Lemsaddek T. Cunha E. Tavares L. Oliveira M. Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control Microorganisms 20231195310.3390/microorganisms 1104095337110376 PMC 10141167 · doi ↗ · pubmed ↗
- 4Madni W.A. Zahoor M.A. Nawaz Z. Khurshid M. Prevalence and Sequence Analysis of Escherichia coli Harboring Colistin, Gentamicin, Streptomycin, Tetracycline and Quinolones Resistant Genes from Commercial Broilers Pak. Vet. J.20254539039510.29261/pakvetj/2025.010 · doi ↗
- 5Shoaib M. Hameed M.F. Aqib A.I. Wang W. Wang Q. Wang S. Pu W. Emerging threat of antimicrobial resistance determinants and plasmid replicon types acquisition by Escherichia coli of poultry and other food-producing animal origin in China: Local findings with global implications Poult. Sci.202510410605510.1016/j.psj.2025.10605541232214 PMC 12662103 · doi ↗ · pubmed ↗
- 6Laube H. Friese A. von Salviati C. Guerra B. Käsbohrer A. Kreienbrock L. Roesler U. Longitudinal monitoring of extended-spectrum-beta-lactamase/Amp C-producing Escherichia coli at German broiler chicken fattening farms Appl. Environ. Microbiol.2013794815482010.1128/AEM.00856-1323747697 PMC 3754693 · doi ↗ · pubmed ↗
- 7Dierikx C. van der Goot J. Fabri T. van Essen-Zandbergen A. Smith H. Mevius D. Extended-spectrum-β-lactamase- and Amp C-β-lactamase-producing Escherichia coli in Dutch broilers and broiler farmers J. Antimicrob. Chemother.201368606710.1093/jac/dks 34922949623 · doi ↗ · pubmed ↗
- 8Landoni M.F. Albarellos G. The use of antimicrobial agents in broiler chickens Vet. J.2015205212710.1016/j.tvjl.2015.04.01625981931 · doi ↗ · pubmed ↗