Taxonomic revision of Conobolbitina (Bolbitiaceae, Agaricales) based on collections from Jilin Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
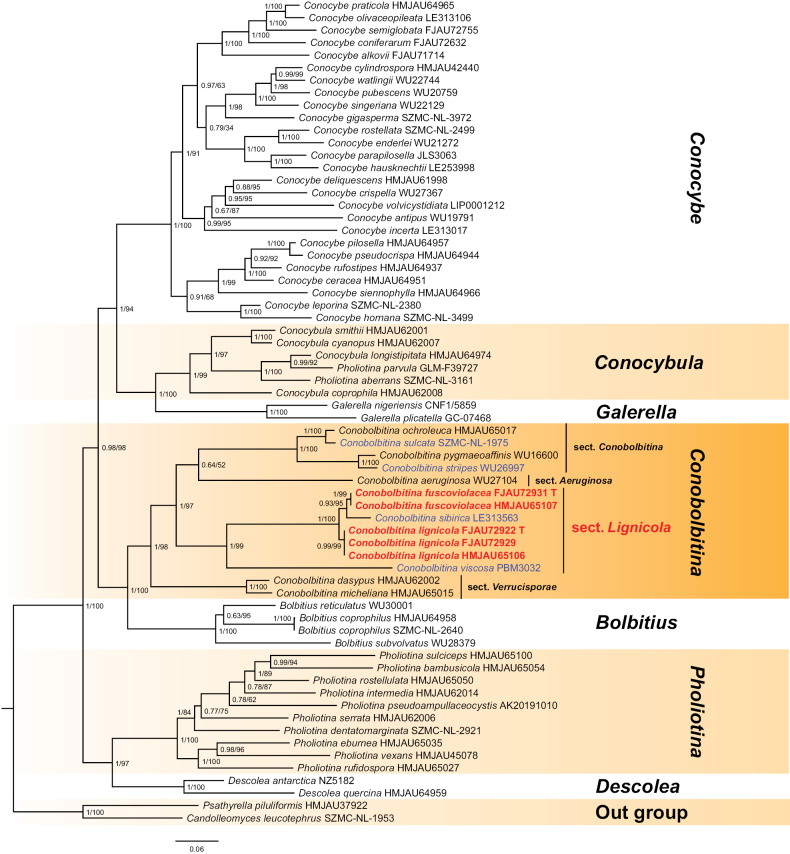 Figure 1
Figure 1 Figure 2
Figure 2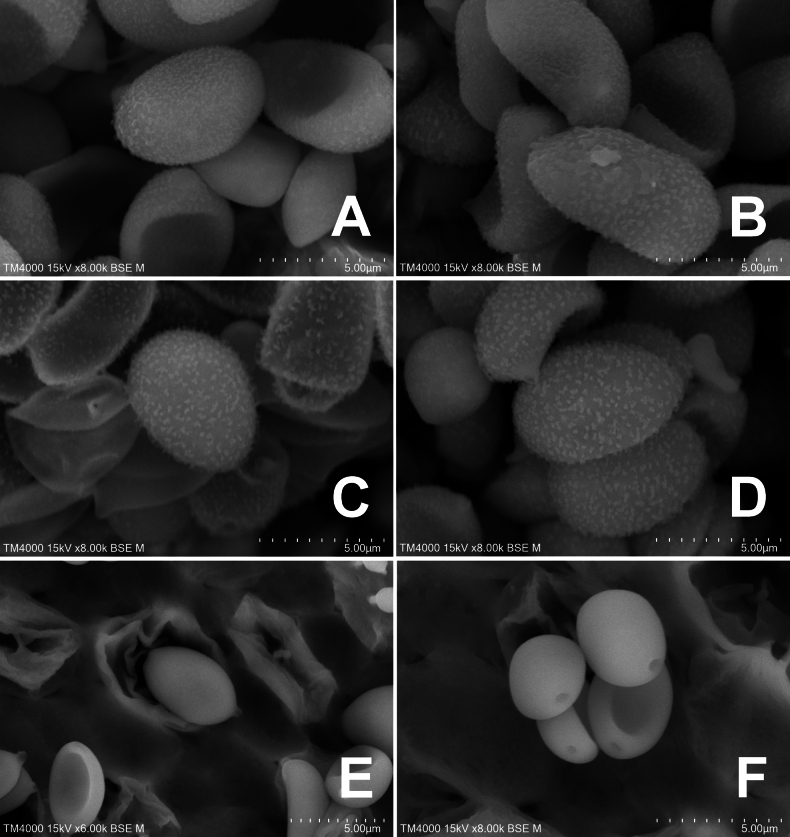 Figure 3
Figure 3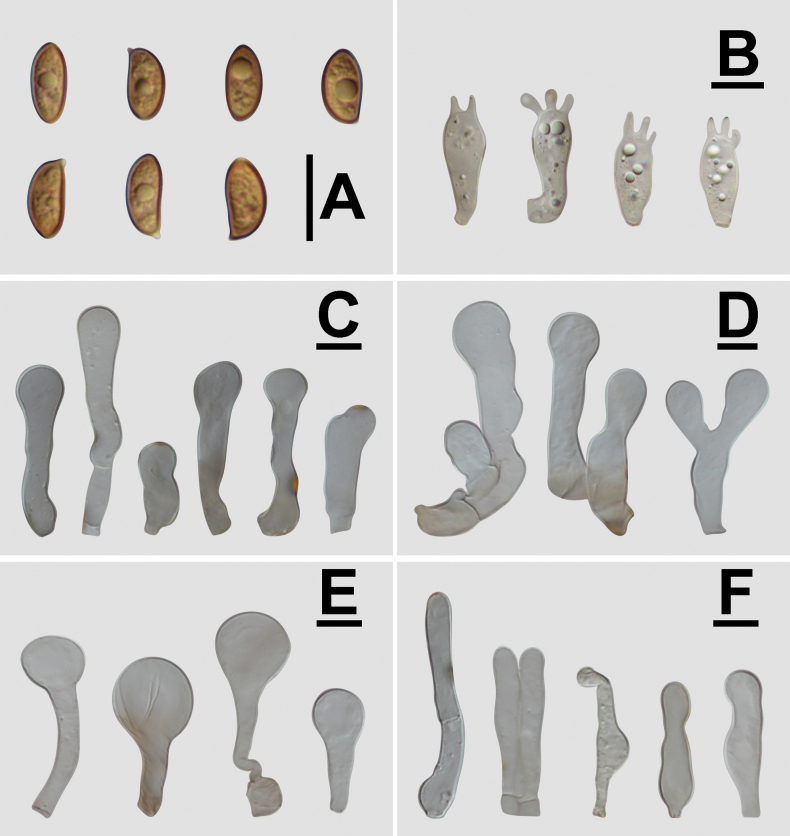 Figure 4
Figure 4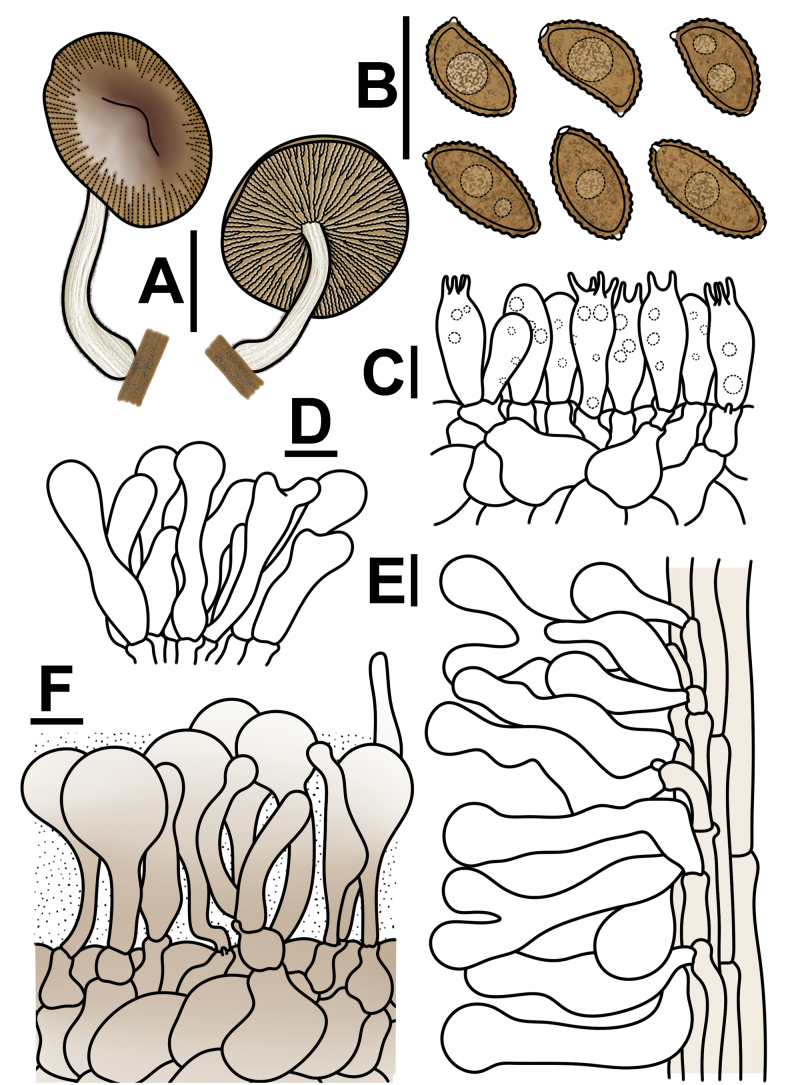 Figure 5
Figure 5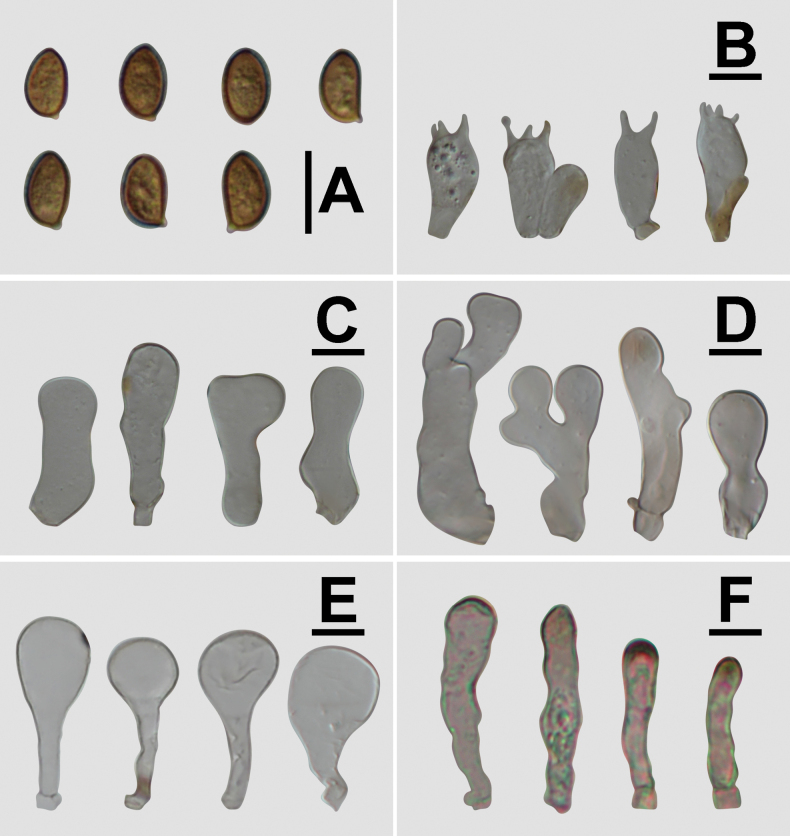 Figure 6
Figure 6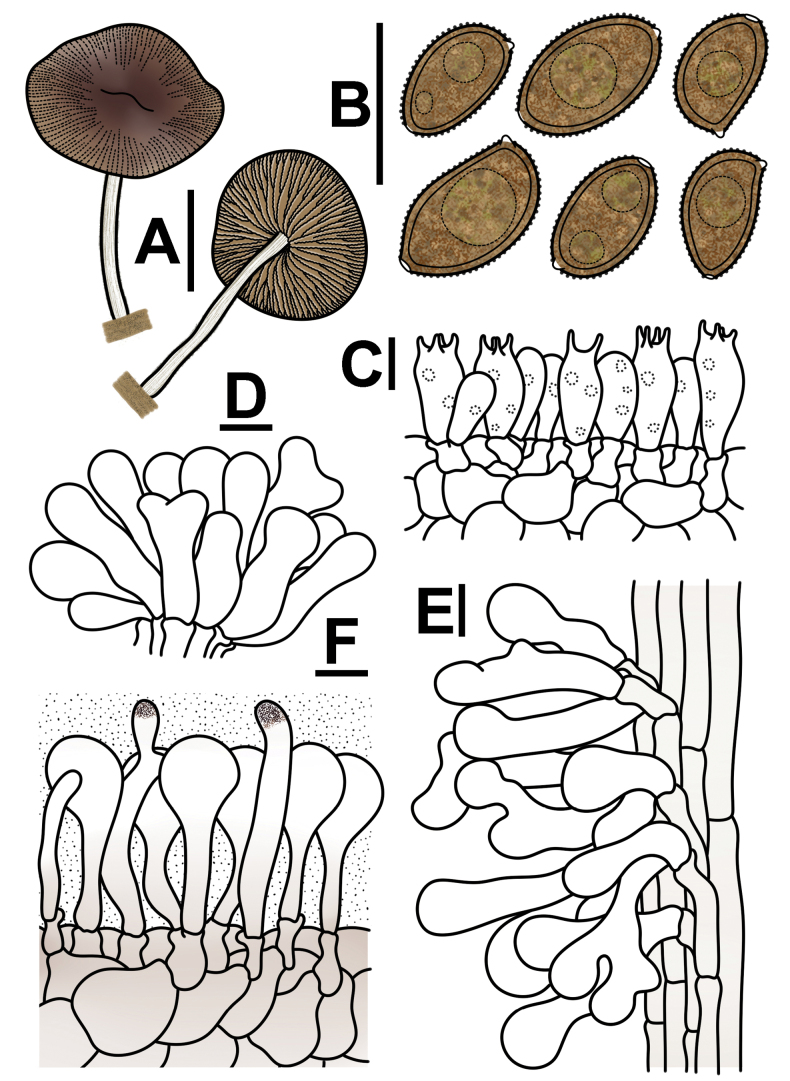 Figure 7
Figure 7| Taxon | Voucher specimen | GenBank accession numbers | Origin | References | ||
|---|---|---|---|---|---|---|
|
|
| |||||
|
| HMJAU64958 |
|
| – | China |
|
|
| SZMC-NL-2640 |
|
| – | Hungary |
|
|
| WU30001 |
|
|
| Hungary |
|
|
| WU28379 |
|
|
| Italy |
|
|
| SZMC-NL-1953 |
|
|
| Hungary |
|
|
|
|
|
|
|
|
|
|
| WU19791 |
|
|
| Austria |
|
|
| HMJAU64951 |
|
|
| China |
|
|
|
|
|
|
|
|
|
|
| WU27367 |
|
|
| Australia |
|
|
| HMJAU42440 |
|
|
| China | |
|
| HMJAU61998 |
|
|
| China |
|
|
| WU21272 |
|
| – | Italy |
|
|
| SZMC-NL-3972 |
|
|
| Slovakia |
|
|
|
| – | – | Russia |
| |
|
| SZMC-NL-3499 |
|
|
| Slovakia |
|
|
|
| – | – | Russia |
| |
|
| SZMC-NL-2380 |
|
|
| Hungary |
|
|
|
| – | – | Russia |
| |
|
| JLS3063 |
| – | – | Spain |
|
|
| HMJAU64957 |
|
|
| China |
|
|
| HMJAU64965 |
|
|
| China | |
|
| HMJAU64944 |
|
|
| China |
|
|
| WU20759 |
|
|
| Italy |
|
|
| SZMC-NL-2499 |
|
|
| Sweden |
|
|
| HMJAU64937 |
|
|
| China |
|
|
|
|
|
|
|
|
|
|
| HMJAU64966 |
|
|
| China |
|
|
| WU22129 |
|
|
| Austria |
|
|
| LIP0001212 |
| – | – | France |
|
|
| WU22744 |
|
|
| Finland |
|
|
| HMJAU62008 |
|
|
| China |
|
|
| HMJAU62007 |
|
|
| China |
|
|
| HMJAU64974 |
|
|
| China |
|
|
| HMJAU62001 |
|
|
| China | |
|
| WU27104 |
|
| – | Germany |
|
|
| HMJAU62002 |
|
|
| China |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| HMJAU65015 |
|
|
| China |
|
|
| HMJAU65017 |
|
|
| China |
|
|
| WU16600 |
|
| – | Austria |
|
|
|
|
| – | Russia | ||
|
| WU26997 |
|
|
| Austria | |
|
| SZMC-NL-1975 |
|
|
| Hungary | |
|
| PBM3032 |
|
| – | USA | Direct Submission |
|
| NZ5182 |
| – | – | USA |
|
|
| HMJAU64959 |
|
|
| China |
|
|
| CNF1/5859 |
|
|
| Nigeria |
|
|
| GC-07468 |
|
| – | Italy | Direct Submission |
|
| SZMC-NL-3161 |
|
|
| Sweden |
|
|
| HMJAU65054 |
|
|
| China |
|
|
| SZMC-NL-2921 |
|
|
| Hungary |
|
|
| HMJAU65035 |
|
|
| China |
|
|
| HMJAU62014 |
|
|
| China |
|
|
| GLM-F39727 |
| – | – | Germany | Direct Submission |
|
| AK20191010 |
| – | – | Germany |
|
|
| HMJAU65050 |
|
|
| China |
|
|
| HMJAU65027 |
|
|
| China |
|
|
| HMJAU62006 |
|
|
| China |
|
|
| HMJAU65100 |
|
|
| China |
|
|
| HMJAU45078 |
|
|
| China |
|
|
| HMJAU37922 |
|
|
| China |
|
| 1 | Basidiospores smooth |
|
| – | Basidiospores rough |
|
| 2 | Obvious pointed ornamentation, growing on decayed wood |
|
| – | Shallow ornamentation, growing in the humus layer |
|
| 3 | Pileocystidia with unpigmented, Qm is 1.72 |
|
| – | Pileocystidia with chocolate-colored pigment, Qm is 1.65 |
|
| 4 | Pileus golden yellow |
|
| – | Pileus beige |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Slime Mold and Myxomycetes Research · Plant Pathogens and Fungal Diseases
Introduction
The genus Conobolbitina T. Bau & H.B. Song was established by Song and Bau (2024) to resolve the polyphyletic problem of Pholiotina sensu lato. Based on morphological and phylogenetic studies, the initial circumscription of Conobolbitina included species formerly classified under Bolbitius Fr., Conocybe Fayod, and Pholiotina Fayod (Song and Bau 2024). Conobolbitina is a transitional group, sharing characteristics with all three genera, a concept also reflected in its name, derived from “Cono” (Conocybe), “bolbi” (Bolbitius), and “tina” (Pholiotina) (Song and Bau 2024).
Song and Bau (2024) divided the genus into three sections based on the presence of a blue-green tinge on the pileus and on whether the basidiospores are verrucose or smooth: Conobolbitina sect. Conobolbitina, sect. Aeruginosa (Hauskn. & Krisai) T. Bau & H.B. Song, and sect. Verrucisporae (Singer) T. Bau & H.B. Song. Section Conobolbitina was originally described as Pholiotina sect. Piliferae Hauskn. & Krisai (Hausknecht 2007). However, based on differences in cheilocystidia shape, the absence of the first intron in the tef1-α (983–2218 bp) region in Conocybula T. Bau & H.B. Song, and, critically, their distinct phylogenetic positions, some species formerly placed in sect. Piliferae were transferred to Conocybula (Song and Bau 2024). Because the type species of Conobolbitina resides within this group, the section name is nomenclaturally established as sect. Conobolbitina [= sect. Piliferae] (Song and Bau 2024). Section Aeruginosa was initially described as Pholiotina series Aeruginosa Hauskn. & Krisai under sect. Cyanopodae Singer (Singer 1973; Hausknecht 2007). This series was later elevated to sect. Aeruginosa within Conobolbitina based on phylogenetic evidence. In addition, sect. Cyanopodae (Singer) T. Bau & H.B. Song is characterized by a blue coloration at the stipe base and by sub-lecythiform cheilocystidia (Song and Bau 2024). Section Verrucisporae was established to accommodate species with verrucose basidiospores. In the phylogeny presented by Song and Bau (2024), Conobolbitina sp.1 and Conobolbitina sp.2 both possess verrucose basidiospores, fitting the morphological definition of this section. However, their phylogenetic positions resulted in a paraphyletic grouping of sect. Verrucisporae, representing a current taxonomic issue within the genus (Song and Bau 2024).
Research on Conobolbitina in China remains limited. As of December 2025, only three confirmed species and two undetermined species have been reported from the country, with records prior to 2024 restricted to a single species (Liu 2018; Song et al. 2023; Song and Bau 2024). For instance, Con. dasypus (Romagn.) T. Bau & H.B. Song was initially reported from China by Liu (2018), who treated it separately as P. utriformis (P.D. Orton) Bon and P. dasypus (Romagn.) P.-A. Moreau (Bon 1991; Moreau 2005). Subsequent taxonomic revisions have demonstrated that both names are synonyms of Con. dasypus (Song and Bau 2024). This synonymy is supported by Arnolds (2005), who recognized Conocybe subnuda Kühner as a synonym of C. utriformis P.D. Orton, and by Hausknecht (2009), who further synonymized both C. utriformis and C. subnuda with P. dasypus (Romagn.) P.-A. Moreau. During his revision of Romagnesi’s Naucoria specimens, Moreau (2005) determined that the earliest valid description of this taxon was made by Romagnesi (1937), thereby establishing dasypus as the correct epithet. The transfer of this species to Conobolbitina was formally completed by Song and Bau (2024). The other two confirmed species in China are Con. ochroleuca T. Bau & H.B. Song, which has been found in both Northeast and Southwest China (e.g., collected by the authors in Dali, Yunnan, in November 2025, confirming its distribution in Southwest China; see Fig. 2), and Con. micheliana T. Bau & H.B. Song, which is known only from Yunnan Province (Song and Bau 2024).
The phylogenetic relationships of Conobolbitina within the Bolbitiaceae family were analyzed using Bayesian inference and maximum likelihood methods based on a multi-marker dataset (ITS, nrLSU, and tef1-α). In the phylogenetic tree, newly proposed species are highlighted in bold red, and newly combined species are shown in blue. T = holotype.
Basidiomata of Conobolbitina species. A, B. Con. lignicola (FJAU72922 T); C, D. Con. lignicola (FJAU72929); E, F. Con. fuscoviolacea (FJAU72931 T); G. Con. ochroleuca (FJAU72937); H. Con. ochroleuca (FJAU72951). T = holotype. Habitat photos of Con. lignicola (HMJAU65106) and Con. fuscoviolacea (HMJAU65107) are shown in fig. 7 of Song and Bau (2024). Scale bars: 1 cm.
The rarity of these fungi means that Conobolbitina sp.1 and Conobolbitina sp.2, which contributed to the paraphyly of sect. Verrucisporae were each known from only one specimen (Song and Bau 2024). Repeated annual collections were conducted at their original sites, and both taxa were successfully recollected between June and July 2025. Based on these new collections, this study aims to re-evaluate and resolve the paraphyly of sect. Verrucisporae propose new combinations for relevant species of Conobolbitina and clarify species diversity within the genus.
Materials and methods
Abbreviations
For Latin names: B . = Bolbitius; C. = Conocybe; Ca. = Candolleomyces; Co. = Conocybula; Con. = Conobolbitina; D. = Descolea; G. = Galerella; P. = Pholiotina; Po. = Populus; Ps. = Psathyrella.
Samplings and morphological analyses
The specimens used in this study were collected in Jilin Province, China, between 2021 and 2025. Fresh basidiomata were photographed in situ, and macroscopic data were recorded, as well as geographic and ecological data, including collection site, geographic coordinates, elevation, date, habitat, and substrate. Given their typically small and delicate habitus, the basidiomata were dried rapidly using silica gel to preserve microscopic structures. Each basidioma was individually wrapped in single-ply tissue paper or corn fiber bags, placed in zip-lock bags with silica gel, and accompanied by a herbarium label. The dried specimens are deposited in the Fungarium of Jilin Agricultural University (HMJAU/FJAU).
For microscopic examination, the specimens were rehydrated in distilled water or a 5% KOH solution. When necessary, a 1% Congo red solution was applied to stain structures for improved visualization (Hausknecht 2009). Microscopic characteristics, including basidia, basidiospores, cystidia, and pileipellis elements, were observed and photographed using a Nikon Eclipse 80i microscope under oil immersion at magnifications of up to ×1000. For scanning electron microscopy (SEM) analysis, fragments of the lamellae were taken from dried specimens. These fragments were sputter-coated with gold and examined using a Hitachi TM4000 Plus II tabletop microscope at the Dali University Analysis and Testing Center. The colors of both fresh and dried basidiomata were described using the color-coding system of the German Institute for Quality Assurance and Certification (Reichs-Ausschuss für Lieferbedingungen und Gütesicherung, RAL; https://www.ral-guetezeichen.de/), abbreviated as RAL in the text.
In this study, basidiospore measurements exclude the apiculus. Measurements are expressed in the format “(a)b–c(d),” where “b–c” represents a minimum of 90% of the measured values, and “a” and “d” indicate the extremes. The dimensions of basidia, cheilocystidia, caulocystidia, and pileipellis (excluding sterigmata or excrescences) were measured when present. At least 20 structures were measured for each noted feature. The notation “(n/m/p)” indicates that measurements were taken from “n” randomly selected basidiospores obtained from “m” basidiomata belonging to “p” different collections. Twenty or 30 basidiospores were measured from each basidioma. The length-to-width ratio is referred to as Q. The mean quotient (Qm), representing the average Q, was calculated together with the standard deviation (Li et al. 2024).
DNA extraction, PCR amplification, and sequencing
Total genomic DNA was extracted following the protocol of Song and Bau (2025). Polymerase chain reaction (PCR) amplifications were performed on a Bio-Rad T100™ Thermal Cycler (Bio-Rad Inc., Hercules, CA, USA) using the primer pairs ITS1F/ITS4 (White et al. 1990; Gardes and Bruns 1993), LR0R/LR7 (Vilgalys and Hester 1990; Moncalvo et al. 2000), and EF1-983F/EF1-2218R (Rehner and Buckley 2005) to amplify the ITS, nrLSU, and tef1-α gene regions, respectively. The reaction mixture composition and thermal cycling conditions followed Song and Bau (2025).
PCR products were separated by 1% agarose gel electrophoresis. Electrophoresis was conducted at 80–120 V for 30 minutes using a 2000 bp DNA ladder for size reference, and the gels were visualized under UV light. The purified PCR products were sent to Sangon Biotech (Shanghai) Co., Ltd. for sequencing. The ITS region was sequenced in one direction, whereas the nrLSU and tef1-α regions were sequenced in both directions and subsequently assembled.
Raw sequence chromatograms (.ab1 files) were inspected and edited using BioEdit v7.2.5 (Hall 1999). For the ITS sequences, the defined region from the 3’ end of the 18S rRNA gene (GGAAGGATCATTA) to the 5’ end of the 28S rRNA gene (TGACCTCAAA) was selected. For nrLSU and tef1-α sequences, low-quality terminal regions with noisy signals were trimmed. Polymorphic sites within sequences were encoded using standard IUPAC ambiguity codes (e.g., R = A/G, Y = C/T). In addition, BLAST searches were conducted against the National Center for Biotechnology Information (NCBI) database to verify the taxonomic identity of the sequences and to exclude potential contaminants. The final sequences were deposited in GenBank.
Phylogenetic analyses
Sequence data for phylogenetic analyses were obtained from GenBank (Table 1) and supplemented with newly generated sequences from this study. The ITS, nrLSU, and tef1-α sequences were aligned separately using the G-INS-i algorithm implemented in the online MAFFT tool (Katoh et al. 2019; https://mafft.cbrc.jp/alignment/server/), with the maximum number of iterative refinements set to two. The resulting alignments were manually adjusted and trimmed at the ends to remove poorly aligned regions using MEGA7 (Kumar et al. 2016). A concatenated dataset was then assembled using the “Concatenate Sequence” function in PhyloSuite v2 (Zhao et al. 2025), with gaps treated as missing data (?).
The best-fit partitioning scheme and substitution models (edge-linked) were selected under the Akaike and Bayesian information criteria using ModelFinder v2.2.0 (Kalyaanamoorthy et al. 2017). Maximum likelihood (ML) analysis was performed using IQ-TREE 3 (Nguyen et al. 2015; Wong et al. 2025) with the selected models, and branch support was assessed using 1000 standard bootstrap replicates and the Shimodaira–Hasegawa–like approximate likelihood ratio test. Bayesian inference (BI) was conducted using MrBayes v3.2.7a (Ronquist et al. 2012) under the partitioned model, running four simultaneous Markov Chain Monte Carlo (MCMC) chains for 1,000,000 generations and sampling every 1000^th^ generation. The first 25% of trees were discarded as burn-in after confirming that the average standard deviation of split frequencies had fallen below 0.004. Phylogenetic trees were visualized and finalized using iTOL (Letunic and Bork 2019), Adobe Photoshop 2021, and Adobe Illustrator 2021. Species of Psathyrella and Candolleomyces (Song and Bau 2023) were selected as the outgroup.
Results
Phylogenetic analyses
The phylogenetic tree was reconstructed based on a combined dataset of ITS, nrLSU, and tef1-α sequences using Bayesian inference (BI). The maximum likelihood (ML) tree is not shown because it exhibited a topology consistent with the Bayesian tree. Bayesian posterior probabilities (PP) and ML bootstrap values (MLbs) are indicated at the tree nodes (Fig. 1). The multi-marker dataset (ITS + nrLSU + tef1-α) comprised 827 bp of ITS, 1345 bp of nrLSU, and 1102 bp of tef1-α. The final alignment included 66 sequences with 3274 sites, comprising 1445 distinct patterns, 1048 parsimony-informative sites, 244 singleton sites, and 1982 constant sites. For the ML analysis, the best-fit substitution models selected under the Akaike Information Criterion (AIC) were GTR+F+R4 for ITS, TIM3+F+R8 for nrLSU, and SYM+I+G4 for tef1-α. For the Bayesian analysis, the best-fit models selected under the Bayesian Information Criterion (BIC) were GTR+F+I+G4 for ITS and nrLSU and SYM+I+G4 for tef1-α.
In the phylogenetic tree (Fig. 1), new species and the new section are highlighted in bold red, whereas some newly combined species are shown in blue. With Ps. piluliformis (Bull.) P.D. Orton and Ca. leucotephrus (Berk. & Broome) D. Wächt. & A. Melzer as outgroups, all genera within Bolbitiaceae Singer formed a monophyletic group in a well-resolved tree. Among them, Conobolbitina and Bolbitius were recovered as sister groups with strong support (PP = 1, MLbs = 100).
Within Conobolbitina, specimens FJAU72922, FJAU72929, and HMJAU65106 formed a distinct clade, which was sister to a clade comprising Con. sibirica (LE313563), FJAU72931, and HMJAU65107, also with full support (PP = 1, MLbs = 100). Furthermore, FJAU72931 and HMJAU65107 formed a well-supported clade (PP = 0.93, MLbs = 95) that was sister to Con. sibirica (LE313563).
A BLAST search of the ITS sequence from FJAU72931 against the NCBI database revealed similarity values of 95.2% with Con. sibirica (LE313563), 94.1% with FJAU72922, 79.9% with Con. viscosa (PBM3032), and 79.37% with Con. aeruginosa (WU27104). For FJAU72922, the ITS sequence showed 92.8% similarity with Con. sibirica (LE313563), 80.2% with Con. viscosa (PBM3032), and 79.21% with Con. aeruginosa (WU27104).
Based on their distinct phylogenetic placements and morphological characteristics, two new species are proposed: Con. lignicola (represented by FJAU72922, FJAU72929, and HMJAU65106) and Con. fuscoviolacea (represented by FJAU72931 and HMJAU65107). To resolve the paraphyly of sect. Verrucisporae, a new section, sect. Lignicola, is established to accommodate the monophyletic clade comprising Con. fuscoviolacea, Con. lignicola, Con. sibirica, and Con. viscosa. This new section is sister to the clade comprising sect. Conobolbitina and sect. Aeruginosa, with strong support (PP = 1, MLbs = 97). Furthermore, seven new combinations are proposed based on their morphology or phylogenetic relationships with other species within Conobolbitina.
Taxonomy
Conobolbitina
Taxon classificationFungiAgaricalesBolbitiaceae
T. Bau & H.B. Song, in Song & Bau, Mycosphere 15(1): 1613 (2024)
45BECCC4-0D55-5657-8C13-C9E143FC752E
Type species.
Conobolbitina pygmaeoaffinis (Fr.) T. Bau & H.B. Song.
Notes.
Conobolbitina is characterized by the absence of an annulus, a Bolbitius-like but non-deliquescent habit, and the presence of diverse, non-lecythiform cheilocystidia. These fungi typically grow in meadows, as well as in forested habitats on forest litter or on decaying wood. The genus has a wide global distribution (Song and Bau 2024).
Conobolbitina
section Conobolbitina
Taxon classificationFungiAgaricalesBolbitiaceae
92D54298-8077-58B2-8BC0-3E29ED421E31
Synonymy.
Pholiotina section Piliferae Hauskn. & Krisai, Öst. Z. Pilzk. 16: 136 (2007).
Notes.
This section is characterized by the absence of a veil and any blue or blue-green coloration, smooth basidiospores, and lageniform to utriform or subcylindrical cheilocystidia with elongated necks that lack a well-defined capitulum. These fungi grow in diverse habitats, including forests, meadows, grassy roadsides, dung, and compost. The section has a broad global distribution (Song and Bau 2024).
Conobolbitina
sulcata
Taxon classificationFungiAgaricalesBolbitiaceae
(Arnolds & Hausknecht) T. Bau & H.B. Song comb. nov.
43C39BCC-AA36-56CF-8579-79A3C6286E81
MB861426
Basionym.
Pholiotina sulcata Arnolds & Hauskn., Persoonia 18(2): 248 (2003).
Synonymy.
Pholiotina sulcata var. oreina Hauskn., Österr. Z. Pilzk. 16: 105 (2007).
Notes.
This species is characterized by smooth basidiospores and lageniform to long-necked lageniform cheilocystidia. Phylogenetic analysis confirms its placement within sect. Conobolbitina, based on sequence data from the specimen SZMC-NL, which is deposited in the Department of Microbiology, University of Szeged (Arnolds and Hausknecht 2003; Tóth et al. 2013).
Conobolbitina
striipes
Taxon classificationFungiAgaricalesBolbitiaceae
(Cooke) T. Bau & H.B. Song comb. nov.
E6C0174A-6DAA-51D9-B833-93C31511A5EE
MB861429
Basionym.
Agaricus striipes Cooke [as ‘striaepes’], Ill. Brit. Fung. (London) 4(29/30): pl. 478 (1885).
Synonymy.
Pholiotina striipes (Cooke) M.M. Moser [as ‘striaepes’], in Gams, Kl. Krypt.-Fl., Edn 3 (Stuttgart) 2b/2: 229 (1967). Conocybe striipes (Cooke) S. Lundell [as ‘striaepes’], Fungi Exsiccati Suecici 41–42: 2049 (1953). Naucoria striipes (Cooke) Sacc. [as ‘striaepes’], Syll. fung. (Abellini) 5: 839 (1887).
Notes.
Both the morphological description by Hausknecht (2009) and the phylogenetic analysis by Tóth et al. (2013) support the placement of this species within sect. Conobolbitina.
Conobolbitina
section Aeruginosa
Taxon classificationFungiAgaricalesBolbitiaceae
(Hauskn. & Krisai) T. Bau & H.B. Song, in Song & Bau, Mycosphere 15(1): 1616 (2024)
28240D49-FF25-50A6-963D-FC197BDD5F3B
Type species.
Conobolbitina aeruginosa (Romagn.) T. Bau & H.B. Song.
Basionym.
Pholiotina series Aeruginosa Hauskn. & Krisai, Öst. Z. Pilzk. 16: 135 (2007).
Notes.
This section is characterized by a pileus with a blue, pale greyish-blue to dark blue disc, the absence of a veil, smooth basidiospores, and lageniform to long-necked lageniform cheilocystidia (Hausknecht 2007; Song and Bau 2024).
Conobolbitina
atrocyanea
Taxon classificationFungiAgaricalesBolbitiaceae
(Esteve-Raventós, Hausknecht & Rejos) T. Bau & H.B. Song comb. nov.
080FA383-0483-5405-A064-EAD4BF7B8094
MB861434
Basionym.
Pholiotina atrocyanea Esteve-Rav., Hauskn. & Rejos, Österr. Z. Pilzk. 16: 118 (2007).
Notes.
This species possesses a dark blue-green pileus, smooth basidiospores, and lageniform to sublageniform cheilocystidia, all of which align with the diagnostic characteristics of sect. Aeruginosa (Esteve-Raventós et al. 2007; Song and Bau 2024). This placement is consistent with its classification in the series Aeruginosa by Hausknecht (2009). It is worth noting that no sequence data are available to confirm this placement.
Conobolbitina
section Verrucisporae
Taxon classificationFungiAgaricalesBolbitiaceae
(Singer) T. Bau & H.B. Song, in Song & Bau, Mycosphere 15(1): 1614 (2024)
CE90BA8F-26E0-5A17-9C8D-612C96F3BD0E
Type species.
Conobolbitina verrucispora (Singer) T. Bau & H.B. Song.
Basionym.
Pholiotina section Verrucisporae Singer, Beih. Sydowia 7: 79 (1973).
Synonymy.
Pholiotina subsection Verrucisporae (Singer) Arnolds, Persoonia 18(2): 229 (2003).
Description.
Based on the description in Hausknecht and Krisai-Greilhuber (2007) with revisions: Basidiomata small to medium. Pileus smooth to slightly rugulose, dry, slightly glutinous to distinctly viscid. Veil absents or weakly developed, soon vanishing. Basidiospores in light microscope almost smooth, with shallow ornamentation, slightly irregularly ridged to punctate, small to medium, with a distinct germ-pore present or absent. Cheilocystidia utriform to clavate-subcapitate. Pileocystidia present, versiform. Found in forests, on soil or litter, rarely in grassland, and almost never on wood.
Notes.
To resolve the paraphyly of sect. Verrucisporae, we have excluded those taxa characterized by a strongly viscid pileus, basidiospores with prominent, acute ornamentation, and a lignicolous habit. This taxonomic revision is supported by integrated morphological and phylogenetic evidence.
Conobolbitina
australis
Taxon classificationFungiAgaricalesBolbitiaceae
(Singer) T. Bau & H.B. Song comb. nov.
F84C6249-09A2-5ED3-ADFF-812CE60FA5F9
MB861433
Basionym.
Pholiotina australis Singer, Beih. Nova Hedwigia 29: 214 (1969).
Synonymy.
Conocybe australis (Singer) Watling, in Watling & Gregory, Biblthca Mycol. 82: 96 (1981).
Notes.
This species, originally discovered in Patagonia (Argentina), was treated by Singer (1969) as a taxon closely related to Con. dasypus (originally described as P. dasypus). Horak and Hausknecht (2002) described Con. australis as having a vividly ochre-brown pileus, a white to pallid stipe, a fugacious veil, a subviscid pileipellis with scattered pileocystidia, and, most critically, basidiospores that are finely roughened (with insignificant ornamentation). Based on this morphological concept, we classify this species within sect. Verrucisporae. It is worth noting that no sequence data are available to confirm this placement.
Conobolbitina
section Lignicola
Taxon classificationFungiAgaricalesBolbitiaceae
T. Bau & H.B. Song sect. nov.
C860C699-307E-50AA-9294-83D087508390
MB861435
Etymology.
“Lignicola” refers to the species in this section that inhabit decaying wood in forests.
Type species.
Conobolbitina lignicola T. Bau & H.B. Song.
Description.
Basidiomata are small to medium in size, resembling Bolbitius but non-deliquescent. The pileus is smooth to slightly rugulose and distinctly viscid. The veil is absent or indistinct. Basidiospores appear smooth to slightly rough under a light microscope, while scanning electron microscopy reveals prominent pointed ornamentation and irregular punctation. Basidiospores are small to medium in size, with an indistinct germ pore that may be present or absent. Cheilocystidia are diverse, non-lecythiform, and some exhibit excrescences. Caulocystidia are similar to cheilocystidia but slightly larger. Pileocystidia are present, and the species has a gelatinous layer.
Habitat.
Grows on decaying wood in forests.
Known distribution.
Asia (China, Russia), Oceania (Papua New Guinea), and North America (United States of America).
Notes.
This section is distinguished from sect. Verrucisporae by the latter’s basidiospores with shallow ornamentation and its non-lignicolous habit.
Conobolbitina
glutinosa
Taxon classificationFungiAgaricalesBolbitiaceae
(E. Horak & Hausknecht) T. Bau & H.B. Song comb. nov.
36F8FC7E-CE6C-5887-BD12-F8582CF599CD
MB861436
Basionym.
Pholiotina glutinosa E. Horak & Hauskn., Österr. Z. Pilzk. 11: 250 (2002).
Notes.
Based on its lignicolous habit, viscid pileus, and basidiospores with distinct pointed ornamentation, this species is placed in sect. Lignicola (Horak and Hausknecht 2002).
Conobolbitina
viscosa
Taxon classificationFungiAgaricalesBolbitiaceae
(Watling) T. Bau & H.B. Song comb. nov.
89CB540E-FFAE-5BCA-BA1C-A0BBD329CDCD
MB861453
Basionym.
Bolbitius viscosus Watling, Notes R. bot. Gdn Edinb. 34(2): 242 (1975).
Notes.
This species is characterized by basidiospores with distinct ornamentation and a lignicolous habit. Both its morphology and phylogenetic placement support its classification within Conobolbitina sect. Lignicola (Watling 1975).
Conobolbitina
sibirica
Taxon classificationFungiAgaricalesBolbitiaceae
(Bulyonk., E.F. Malysheva & L.B. Kalinina) T. Bau & H.B. Song comb. nov.
91C4CA51-5462-5D03-A1B7-15836939B108
MB861454
Basionym.
Bolbitius sibiricus Bulyonk., E.F. Malysheva & L.B. Kalinina, in Crous et al., Persoonia 46: 419 (2021).
Notes.
The combination of morphological and phylogenetic evidence supports the placement of this species in Conobolbitina sect. Lignicola (Crous et al. 2021).
Conobolbitina
lignicola
Taxon classificationFungiAgaricalesBolbitiaceae
T. Bau & H.B. Song sp. nov.
26CBE719-827A-5739-88E1-F008025B383B
MB861437
Etymology.
“lignicola” refers to the species that inhabits decaying wood in forests.
Holotypus.
China • Jilin Province, Huadian City, Hongshi National Forest Park, 14 June 2025, 42°49'46"N, 127°07'48"E, alt. 454 m, T.Y. Zhang, ZTY2561407 (FJAU72922).
Diagnosis.
Conobolbitina lignicola is characterized by a subbulbous stipe base, subellipsoid to oblong basidiospores with an inconspicuous germ pore, and hyaline pileocystidia. It is further distinguished from Con. glutinosa by its occurrence in temperate broadleaf forests (vs. tropical rainforests of Oceania).
Basidiospores under electron microscopy of Conobolbitina species. A, B. Con. lignicola (FJAU72922 T); C, D. Con. fuscoviolacea (FJAU72931 T); E, F. Con. ochroleuca (FJAU72937). T = holotype. SEM images of basidiospores of Con. lignicola (HMJAU65106) and Con. fuscoviolacea (HMJAU65107) are shown in fig. 8 of Song and Bau (2024). Scale bars: 5 μm.
Description.
Basidiomata small-sized, pileus 1–2 cm in diameter, initially broadly conical to campanulate, later straight, subumbonate, and slightly plano-concave. The pileus center is mahogany brown (RAL 8016) to chocolate brown (RAL 8017), with the margin beige (RAL 1001), sand yellow (RAL 1002) to ochre yellow (RAL 1024). The pileus is hygrophanous, smooth, viscid, with distinct striations extending up to 1/2 of the center, the margin is even to slightly undulate. Context thin, sand yellow (RAL 1002) to ochre yellow (RAL 1024), with no specific odor or taste. Lamellae adnexed to narrowly adnate, ventricose, moderately crowded, unequal in length, light ivory (RAL 1015), sand yellow (RAL 1002) to ochre brown (RAL 8001), with serrulate edges. Stipe 1–3 cm long, 2–3 mm thick, cylindrical, ivory (RAL 1014) to sand yellow (RAL 1002), covered with a powdery pubescence and longitudinally striate. The base is slightly enlarged.
Microscopic structure images of Conobolbitina lignicola (FJAU72922). A. Basidiospores; B. Basidia; C. Cheilocystidia; D. Caulocystidia; E. Pileipellis elements; F. Pileocystidia. Scale bars: 10 μm.
Basidiospores (60/3/3) (7–)7.5–10.3(–11) × (4.1–)4.3–5.6(–6) μm, Q = (1.52–)1.58–2(–2.44), Q_m_ = 1.72(±0.15), nearly ellipsoid to oblong, lemon-shaped to amygdaliform in frontal view, with a slight suprahilar depression in side view, appear smooth to slightly rough under a light microscope, with pointed ornamentation visible under a scanning electron microscope, wall thick, containing oil droplets, germ pore diameter is less than 1 μm or inconspicuous. Basidiospores are ochre brown (RAL 8001) to clay brown (RAL 8003) in KOH solution. Basidia (16–)17–23(–24) × 7–10(–11) μm, clavate, 4-spored, occasionally 2-spored, sterigmata 3–5 μm long, basidia with vacuolar contents. Cheilocystidia (15–)19–41(–42) × 6–11(–12) μm, variable in shape, cylindrical and constricted at the center, broadly capitate, narrowly utriform, broadly clavate, subcylindrical, with slightly bifurcated apices, nearly femoral head-shaped, some with excrescences, and sterile margins. Pleurocystidia absent. Caulocystidia (15–)16–61(–65) × 6–14 μm, variable in shape, similar to cheilocystidia but slightly larger, spheropedunculate, utriform, subcapitate, subclavate, subcylindrical, femoral head-shaped, with swollen and bifurcated apices, some with excrescences. Pileipellis epithelioid hymeniderm, composed of (21–)23–42(–43) × (11–)12–24(–26) μm sphaeropedunculate or broadly clavate elements, with fawn brown (RAL 8007) pigment observed in the base when viewed in KOH solution. Pileocystidia (23–)24–54(–60) × 4–10(–12) μm, variable in shape, including capitate, narrowly utriform, clavate, subcylindrical, and capilliform forms. A gelatinous layer is present. All structures have clamp connections. A weakly positive reaction with ammonia solution is observed, resulting in the formation of rhomboid crystals.
Conobolbitinalignicola (FJAU72922). A. Basidiomata; B. Basidiospores in KOH; C. Hymenium and subhymenium; D. Cheilocystidia; E. Stipitipellis; F. Pileipellis. Scale bars: 1 cm (A); 10 μm (B–F).
Habitat.
It occurs singly or scattered on decaying wood in broad-leaved forests during summer.
Known distribution.
Jilin Province, China.
Additional specimens measured.
China • Jilin Province, Jilin City, Jiaohe City, Qianjin Forest Farm, 24 June 2021, 43°57'10"N, 127°42'7"E, alt. 650 m, Q. Q. Ye, Y2408 (HMJAU65106). China • Jilin Province, Huadian City, Hongshi National Forest Park, 31 July 2025, 42°50'02"N, 127°07'58"E, alt. 487 m, H.B. Song, S2573110 (FJAU72929).
Notes.
This species is distinguished from Con. glutinosa by its occurrence in temperate broadleaf forests (vs. tropical rainforests of Oceania), a grey-brown to deep date-brown pileus, a terete stipe, basidiospores with a distinct germ pore, and smooth cystidia (Horak and Hausknecht 2002). It differs from Con. viscosa by the latter’s olive-black pileus with an involute margin, stipe base with pale yellow mycelium, and indistinct veil (Watling 1975). It is distinguished from Con. sibirica by the latter’s non-subumbonate, olive-black pileus and its weakly to strongly pigmented pileocystidia with intracellular greyish-brown granular pigment (Crous et al. 2021). Furthermore, it differs from all members of the sect. Verrucisporae by its Bolbitius-like habit, lignicolous ecology, and distinct spore ornamentation. Phylogenetically, it forms a distinct clade sister to a cluster containing Con. sibirica and Con. fuscoviolacea, with strong nodal support, and can be readily distinguished.
Conobolbitina
fuscoviolacea
Taxon classificationFungiAgaricalesBolbitiaceae
T. Bau & H.B. Song sp. nov.
B16351CE-2C73-5336-81CA-BC318EA4AC28
MB861444
Etymology.
“fuscoviolacea” refers to the presence of dark violet pigmentation in the pileipellis.
Holotypus.
China • Jilin Province, Huadian City, Hongshi National Forest Park, 31 July 2025, 42°50'01"N, 127°07'57"E, alt. 487 m, T.Y. Zhang, ZTY2573131 (FJAU72931).
Diagnosis.
This species is characterized by a non-bulbous stipe base, ellipsoid to oblong basidiospores, a pileipellis becoming slightly pastel violet in KOH, and pileocystidia with apical, chocolate brown to terra brown pigmentation. The combination of these features distinguishes it from other Conobolbitina species.
Microscopic structure images of Conobolbitina fuscoviolacea (FJAU72931). A. Basidiospores; B. Basidia; C. Cheilocystidia; D. Caulocystidia; E. Pileipellis elements; F. Pileocystidia. Scale bars: 10 μm.
Description.
Basidiomata small-sized. Pileus 2–3 cm in diameter, initially lentiform, later becoming straight, slightly subumbonate, plano-concave. Pileus center nut brown (RAL 8011) to sepia brown (RAL 8014), margin ivory (RAL 1014) to brown beige (RAL 1011). Pileus hygrophanous, smooth, viscid, with distinct striations extending up to 1/2 of the center; the margin even to slightly undulate. Context thin, light ivory (RAL 1015) to ivory (RAL 1014), with no specific odor or taste. Lamellae adnexed to narrowly adnate, ventricose, moderately crowded, unequal in length, beige red (RAL 3012), beige (RAL 1001) to ochre brown (RAL 8001), with serrulate edges. Stipe 2–3 cm long, 2–3 mm thick, cylindrical, light ivory (RAL 1015) to ivory (RAL 1014), surface covered with pruinose pubescence, with longitudinal fibrous striations, base is not swollen and is equal in diameter throughout.
Conobolbitinafuscoviolacea (FJAU72931). A. Basidiomata; B. Basidiospores in KOH; C. Hymenium and subhymenium; D. Cheilocystidia; E. Stipitipellis; F. Pileipellis. Scale bars: 1 cm (A); 10 μm (B–F).
Basidiospores (60/2/2) (6.9–)7.8–10.4(–11.2) × (4.4–)4.8–6.1(–6.6) μm, Q = (1.47–)1.54–1.76(–1.78), Q_m_ = 1.65(±0.07), ellipsoid to oblong, slightly lemon-shaped in frontal view, with a slight suprahilar depression in lateral view. Appears smooth to slightly rough under a light microscope, with pointed ornamentation visible under a scanning electron microscope. Spore wall thick, containing oil droplets. Germ pore diameter less than 1.5 μm or inconspicuous. Basidiospores appear brown beige (RAL 1011) to clay brown (RAL 8003) in KOH solution. Basidia (17–)18–25 × (7–)8–11 μm, clavate, 4-spored, occasionally 2-spored, sterigmata 2–6 μm long, with vacuolar contents in basidia. Cheilocystidia (18–)22–43(–45) × 9–14(–15) μm, variable in shape, cylindrical and constricted at the center, broadly capitate, utriform, clavate, cylindrical, with bifurcated apices, nearly femoral head-shaped, some with excrescences, and sterile margins. Pleurocystidia absent. Caulocystidia (22–)23–60(–63) × 9–16 μm, variable in shape, similar to cheilocystidia but slightly larger, clavate, spheropedunculate, utriform, subcapitate, with bifurcated apices, nearly femoral head-shaped, some with excrescences. Pileipellis epithelioid hymeniderm, composed of (20–)21–42(–45) × 13–20(–21) μm sphaeropedunculate or broadly clavate elements, exhibiting red brown (RAL 8012) to pastel violet (RAL 4009) pigments. Pileocystidia (22–)24–50(–52) × 5–8 μm, clavate, subcylindrical, capilliform, some with apices pigmented chocolate brown (RAL 8017) to terra brown (RAL 8028). A gelatinous layer is present. All structures have clamp connections. A weakly positive reaction with ammonia solution is observed, resulting in the formation of rhomboid crystals.
Habitat.
It grows solitarily on decaying fir wood in summer.
Known distribution.
Jilin Province, China.
Additional specimens measured.
China • Jilin Province, Jilin City, Huadian City, 27 August 2023, 42°49'30"N, 127°08'17"E, alt. 483 m, H. Cheng, C2382702 (HMJAU65107).
Notes.
This species is distinguished from Con. lignicola and Con. glutinosa by the pastel violet reaction of its pileipellis in KOH and the chocolate brown to terra brown apical pigmentation on some pileocystidia (Horak and Hausknecht 2002). It differs from Con. viscosa and Con. sibirica by the latter two having distinctly olive-black pilei (Watling 1975; Crous et al. 2021). Phylogenetically, this species forms a sister group with Con. sibirica, but the latter originates from a log of Populus tremula L. and is phylogenetically distinct (Crous et al. 2021).
Key to Chinese species of Conobolbitina
**: **
Discussion
Within the phylogenetic framework established by Tóth et al. (2013) and Song and Bau (2024), our analyses reveal that Conobolbitina forms a monophyletic group and is sister to Bolbitius. Furthermore, Conobolbitina comprises four monophyletic sections, whose phylogenetic relationships are highly consistent with their morphological characteristics.
Section Conobolbitina is characterized by smooth basidiospores, as observed in Con. ochroleuca (Fig. 3E, F). Within this section, we propose a new combination for Con. sulcata, a species previously classified in Pholiotina, a genus now restricted to taxa with a distinct annulus, a feature lacking in Con. sulcata. This species typically inhabits well-lit environments such as dry grasslands or nutrient-poor meadows (Arnolds and Hausknecht 2003). Its morphology conforms to sect. Conobolbitina, and phylogenetic analysis of the sequence Con. sulcata SZMC-NL-1975 confirms its placement within this section (Tóth et al. 2013). Similarly, we propose a new combination for Con. striipes within sect. Conobolbitina. This species possesses smooth basidiospores and lageniform to long-necked lageniform cheilocystidia, grows in grasslands, parks, or mixed forests, and is phylogenetically placed within this section (Moser 1967).
Section Aeruginosa and sect. Conobolbitina are sister groups. They are distinguished by the bluish-green hue of the pileus in species of sect. Aeruginosa (Song and Bau 2024). However, the phylogenetic support for the relationship between these two sections is relatively low (PP = 0.64, MLbs = 52). This low support may be due to the absence of a tef1-α sequence in the Con. aeruginosa dataset used for phylogenetic reconstruction and the fact that other species within this section either lack molecular data or represent undiscovered diversity. Within sect. Aeruginosa, we propose a new combination only for Con. atrocyanea. This species has a distinctly bluish-green pileus and occurs primarily in deciduous forests of the Mediterranean region, with other morphological characteristics fully conforming to the sectional definition (Esteve-Raventós et al. 2007). Its classification is further supported by Hausknecht (2009), who placed it in series Aeruginosa. Unfortunately, no molecular sequence data are currently available for Con. atrocyanea. This case highlights the critical role of morphological classification, particularly for historical species lacking sequence data. For such taxa, where type specimens are unavailable and recollection at the type locality is not feasible, taxonomic placement must rely on careful analysis of original morphological descriptions, geographical distribution, and ecological data.
Section Lignicola was established to resolve the paraphyly of sect. Verrucisporae. Species in this new section share several key characteristics that clearly distinguish them from other sections, including a Bolbitius-like but non-deliquescent habit, a distinctly viscid pileus, basidiospores with prominent, pointed ornamentation, and a lignicolous habit. Although species in sect. Verrucisporae also possess ornamented basidiospores, the ornamentation is less prominent than in sect. Lignicola, as illustrated in fig. 8 of Song and Bau (2024). Furthermore, species in sect. Verrucisporae are rarely lignicolous. Within the sect. Lignicola, the taxa previously designated as Conobolbitina sp.1 and Conobolbitina sp.2 in Song and Bau (2024), are here formally described as the new species Con. lignicola and Con. fuscoviolacea, respectively. Conobolbitina lignicola occurs on decaying wood in broad-leaved forests, whereas Con. fuscoviolacea is found on decaying fir wood. Several new combinations are also proposed for this section. For example, Con. glutinosa was transferred based on its morphological and habitat characteristics (Horak and Hausknecht 2002), although sequence data were not obtained. Its type locality is Papua New Guinea, where it was found on decaying wood in a tropical montane rainforest dominated by Fagaceae. This species was originally described in Pholiotina, and SEM images of its basidiospores were provided in plate 1 of Horak and Hausknecht (2002). Conobolbitina viscosa was originally described as a Bolbitius species by Watling (1975). The type specimen was collected in Emmet County, Michigan, USA, on decaying wood of Acer L., with SEM images of its basidiospores shown in plate 8 of Watling (1975). Conobolbitina sibirica was described as B. sibiricus by Bulyonk., Malysheva, and Kalinina in 2021 (Crous et al. 2021). Its type locality is near Novosibirsk, Russia, on a log of Po. tremula; SEM images of its basidiospores are included in the entry for B. sibiricus in Crous et al. (2021). The morphological traits and habitats of these three species are consistent with the definition of sect. Lignicola. Among them, Con. viscosa has reliable molecular sequences, and Con. sibirica has sequence data from the type material; phylogenetically, both are placed within the monophyletic sect. Lignicola clade. Accordingly, we have proposed the corresponding new combinations.
We revised the circumscription of sect. Verrucisporae by removing the taxa now assigned to sect. Lignicola. The redefined sect. Verrucisporae is characterized by basidiospores with relatively subtle ornamentation (see fig. 8 in Song and Bau 2024). The type species of the section is Con. verrucispora. Although no molecular sequence data are available for this species, its morphology, as described by Horak and Hausknecht (2002) and Hausknecht and Krisai-Greilhuber (2007), includes a dry, tomentose, and weakly translucent-striate pileus, large projecting pileocystidia, and a non-lignicolous habit, occurring on soil or litter in forests and, rarely, in grasslands. These characteristics differ markedly from those of the excluded taxa, supporting the revised sectional concept. Within this revised section, we propose a new combination for Con. australis. This species exhibits subtle basidiospore ornamentation and a non-lignicolous habit, consistent with the definition of sect. Verrucisporae (Singer 1969). Moreover, Singer (1969) considered Con. australis to be closely related to Con. dasypus. Based on this evidence, we have transferred it to sect. Verrucisporae, although sequence data were not obtained.
This study was prompted by the paraphyly of sect. Verrucisporae, which became evident through the discovery of Conobolbitina spp. (Song and Bau 2024). To address this issue, we first clarified the species composition of the section and proposed new combinations for several Conobolbitina species previously classified under other genera. Building on this framework, we revised the circumscription of sect. Verrucisporae by excluding the taxa responsible for the paraphyly and established a new section, sect. Lignicola, to accommodate them. That this taxonomic clarification was achieved despite Conobolbitina being a transitional genus represented by extremely scarce and rarely collected specimens further underscores the importance of morphology in fungal taxonomy.
Supplementary Material
XML Treatment for Conobolbitina
XML Treatment for Conobolbitina section Conobolbitina
XML Treatment for Conobolbitina sulcata
XML Treatment for Conobolbitina striipes
XML Treatment for Conobolbitina section Aeruginosa
XML Treatment for Conobolbitina atrocyanea
XML Treatment for Conobolbitina section Verrucisporae
XML Treatment for Conobolbitina australis
XML Treatment for Conobolbitina section Lignicola
XML Treatment for Conobolbitina glutinosa
XML Treatment for Conobolbitina viscosa
XML Treatment for Conobolbitina sibirica
XML Treatment for Conobolbitina lignicola
XML Treatment for Conobolbitina fuscoviolacea
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arnolds E (2005) Bolbitiaceae. In: Noordeloos ME, Kuyper TW, Vellinga EC (Eds) Flora Agaricina Neerlandica 6. Taylor & Francis, Boca Raton, London, New York, Singapore, 120–180.
- 2Arnolds E, Hausknecht A (2003) Notulae ad Floram agaricinam neerlandicam-XLI. Conocybe and Pholiotina. Persoonia 18(2): 239–252.
- 3Crous PW, Cowan DA, Maggs-Kolling G, Yilmaz N, Thangavel R, Wingfield MJ, Noordeloos ME, Dima B, Brandrud TE, Jansen GM, Morozova OV, Vila J, Shivas RG, Tan YP, Bishop-Hurley S, Lacey E, Marney TS, Larsson E, Le Floch G, Lombard L, Nodet P, Hubka V, Alvarado P, Berraf-Tebbal A, Reyes JD, Delgado G, Eichmeier A, Jordal JB, Kachalkin AV, Kubatova A, Macia-Vicente JG, Malysheva EF, Papp V, Rajeshkumar KC, Sharma A, Spetik M, Szaboova D, Tomashevskaya MA, Abad JA, Abad ZG, Alexandrova AV, Anand G, A · doi ↗ · pubmed ↗
- 4Esteve-Raventós F, Hausknecht A, Rejos F (2007) Pholiotina atrocyanea, spec. nova, and three other rare Pholiotina species (Bolbitiaceae, Agaricales) from Spain, Österr. Z. Pilzk 16: 117–126.
- 5Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes‐application to the identification of mycorrhizae and rusts. Molecular Ecology 2(2): 113–118. 10.1111/j.1365-294X.1993.tb 00005.x 8180733 · doi ↗ · pubmed ↗
- 6Hall TA (1999) Bio Edit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41(41): 95–98. 10.1021/bk-1999-0734.ch 008 · doi ↗
- 7Hausknecht A (2007) Beiträge zur Kenntnis der Bolbitiaceae 11. Unberingte Arten der Gattung Pholiotina. Österreichische Zeitschrift für Pilzkunde 16(1): 35–116.
- 8Hausknecht A (2009) A monograph of the genera Conocybe Fayod, Pholiotina Fayod in Europe. Edizioni Candusso, Italy, 1–968.