Camellia shangshii (Theaceae), a new species endemic to danxia landscape from Guangdong Province, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
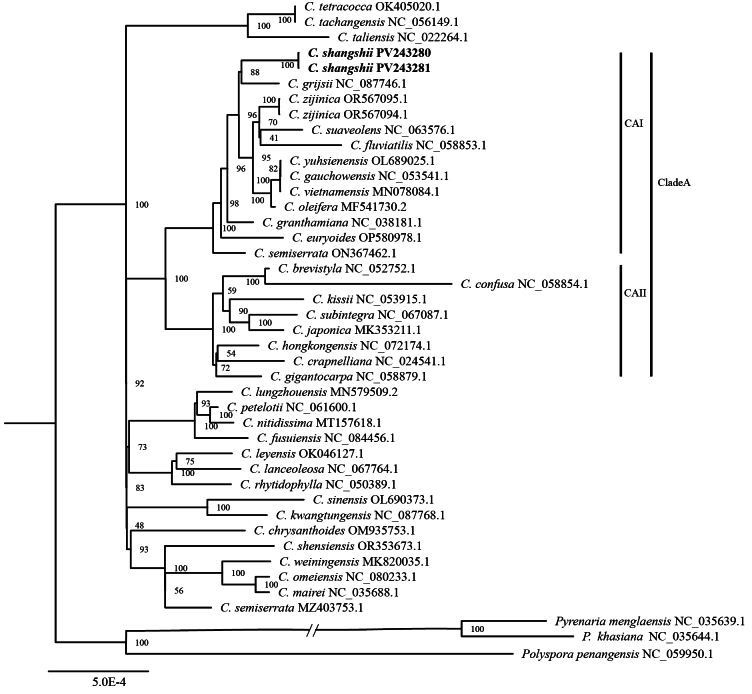 Figure 1
Figure 1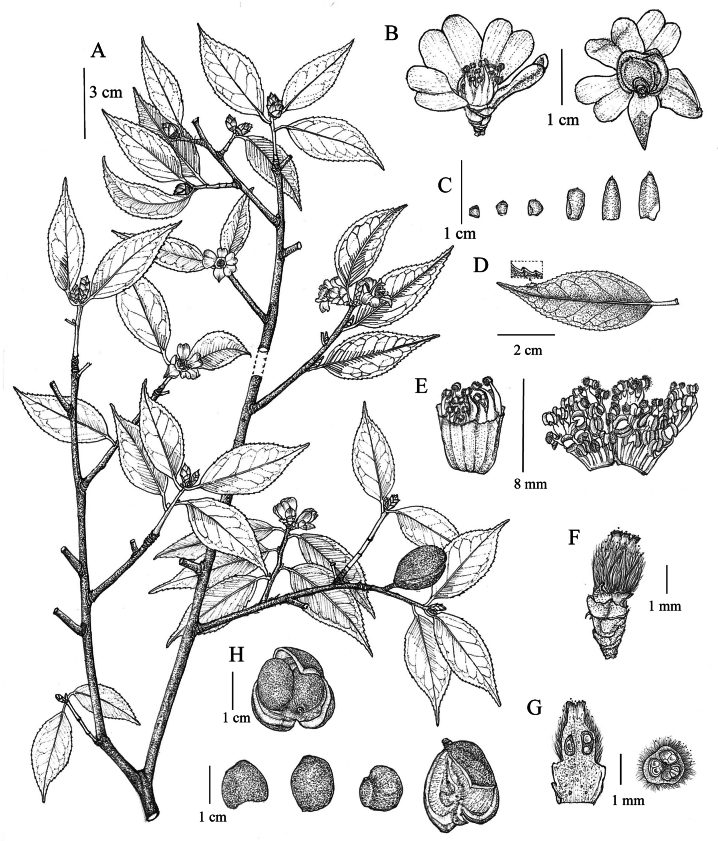 Figure 2
Figure 2 Figure 3
Figure 3| Character |
|
|
|
|---|---|---|---|
| Leaf shape | oblong-lanceolate | oblong-elliptic | elliptic, oblong-elliptic, obovate-elliptic, or oblong-lanceolate |
| Leaf size | 2.5–5.5 × 0.8–1.9 cm | 6–9 × 2.5–3.7 cm | 2–4.8 × 0.8–1.9 cm |
| Leaf apices | acute or acuminate | acuminate or caudate-acuminate | rounded, acute, or acuminate |
| Young branchlets | glabrous | puberulent | glabrous |
| Petiole length | 3–7 mm | 5–8 mm | 1–3 mm |
| Bracteoles and sepals | 6–8 | 9–10 | 4–6 |
| Flower diameter | 1.5–2 cm | 4–5 cm | 1 cm |
| Petals, shape, and size | 5–6, apex concave, connate at the base and conjunctive with stamen, 0.7–1.1 × 0.3–0.5 cm | 5–6, obovate, apex concave, base conjunctive with stamen, 2–2.5 × 1.2–2 cm | 4–6, oblong-elliptic to obovate-elliptic, apex retuse, 0.7–1.1 × 0.4–0.5 cm |
| Stamens | 7–8 mm | 7–8 mm | 4–5 mm |
| Capsule shape and size | ovoid or subglobose, 1.5–1.7 cm | ovoid, 2–2.5 cm | ovoid or subglobose, 1.2 cm |
| Styles | 3, 1–2 mm | 3, 3–4 mm | 3–4, 1–2 mm |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Marine Sponges and Natural Products
Introduction
Camellia L., one of the most primitive genera in the family Theaceae, is predominantly distributed throughout East and Southeast Asia, with southwestern China serving as its center of species diversity and harboring approximately 85% of all known Camellia species (Ming et al. 1999; Zan et al. 2023). Furthermore, a phylogenetic tree constructed from 93,212 genome-wide SNPs across 1,325 Camellia accessions indicates that southwestern China is both the origin and domestication center of these plants (Kong et al. 2025). The genus Camellia holds significant economic importance, featuring species such as C. oleifera for oil, C. japonica in horticulture, and C. sinensis as the source of tea. Furthermore, new species within this genus continue to be discovered regularly (Yu et al. 2021; Ly et al. 2022; Quach et al. 2022; Ye et al. 2022; Chen et al. 2023; Lin et al. 2024; Li et al. 2025). Most of them are critically endangered, characterized by extremely limited distributions and small population sizes. This situation highlights both the richness of biodiversity in China and the critical need to intensify species protection.
During the 2023 biodiversity survey in Danxiashan Mountain, we identified a putative new species of Camellia, which closely resembles C. grijsii in morphology but can be easily distinguished by its characteristic petals and smaller leaves. Based on comprehensive morphological comparisons and phylogenetic analyses conducted from 2023 to 2024, this species was confirmed as a novel member of Camellia sect. Paracamellia. This study provides its formal description and detailed illustrations.
Methods
Morphological study
Field observations and collections of the new species were carried out from 2022 to 2024 in Renhua County, Guangdong Province, China. Morphological comparisons were conducted using living plants, herbarium specimens, and published literature (Chang 1981; Chang and Ren 1998; Ming and Bartholomew 2007). Additional specimen data were obtained from the Chinese Virtual Herbarium (https://www.cvh.ac.cn/). Finally, eleven key morphological characters were utilized to differentiate the species. All character states were measured and described using a dissecting microscope.
Molecular analysis
Fresh leaf materials of individuals were collected and stored in silica gel for subsequent molecular experiments. Whole genomic DNA for each sample was extracted using the modified CTAB method (Doyle and Doyle 1987) and then purified with magnetic beads. A library was constructed for each sample using the True Prep DNA Library Prep Kit, which was then sent for Illumina sequencing on the NovaSeq 6000 platform under standard operating procedures. Raw sequencing data were filtered with fastp v0.23.4 (Chen et al. 2018) to obtain clean data. Chloroplast genomes were assembled using GetOrganelle v1.7.7.0 (Jin et al. 2020) and annotated with cpGAVAS (Liu et al. 2012). The complete chloroplast sequences of C. shangshii were submitted to NCBI (https://www.ncbi.nlm.nih.gov/) and deposited under the accession numbers PV243280–PV243281. Voucher specimens were deposited in the herbarium of Sun Yat-sen University (SYS). For phylogenetic analysis, 41 chloroplast genome sequences, representing 37 Camellia species and three outgroup species, were obtained from the NCBI database for subsequent tree reconstruction.
The chloroplast genome sequences were aligned using MAFFT v7 (Katoh and Standley 2013). TrimAl v1.2 was applied to trim the alignment using the “gap out” model setting (Capella-Gutiérrez et al. 2009). Maximum likelihood (ML) and Bayesian inference (BI) were used to derive and construct the phylogenies, respectively. The ML tree was inferred using IQ-TREE v2.2.6 (Nguyen et al. 2015), with branch support assessed by 1,000 replicates of the SH approximate likelihood ratio test (SH-aLRT). BI was performed using MrBayes v3.2.7a (Ronquist et al. 2012). Markov chain Monte Carlo (MCMC) simulations were run for 2 million generations with one cold chain and three heated chains, starting from random trees, sampling every 100 generations, and continuing until the average standard deviation of split frequencies was below 0.01, after which the first 25% of samples were discarded as burn-in and posterior probabilities were estimated.
Results
Comparative morphology revealed that C. shangshii is similar to C. grijsii among the examined Camellia species. C. shangshii differs from C. grijsii in its smaller flowers (1.5–2 cm vs. 4–5 cm in diameter), leaves (2.5–5.5 × 0.8–1.9 cm vs. 6–9 × 2.5–3.7 cm), and fruits (1.5–1.7 cm vs. 2–2.5 cm). Additionally, the petals are connate at the base for 24 mm, whereas those of C. grijsii are only basally conjunctive with stamens. C. grijsii also has larger petals (2–2.5 × 1.2–2 cm) and puberulent young branchlets (vs. glabrous). Furthermore, the new species differs from C. zijinica in its distinct staminal column, in contrast to the latter, in which only the outer filament whorl is basally connate (1–1.5 mm) (Table 1).
Table 1.: Morphological comparison of Camellia shangshii, C. grijsii, and C. zijinica.
Both maximum likelihood (ML) and Bayesian inference (BI) analyses of the chloroplast genomes yielded highly congruent and well-supported phylogenetic trees. The results strongly supported the new species as sister to C. grijsii (BS = 88; Fig. 1) and placed it within clade CAI together with C. zijinica, C. fluviatilis, C. yuhsienensis, C. oleifera, and other species, with full branch support (BS = 100). The species C. brevistyla, C. japonica, and C. kissii were nested within clade CAII, which was sister to clade CAI, together forming CladeA with highly supported values (PP = 1.00; BS = 100)(Appendix 1).
Maximum likelihood phylogenetic tree of C. shangshii and 41 other related species based on chloroplast genomes. Above the nodes of the tree, maximum likelihood ultrafast bootstrap support values are shown. The new species is highlighted in bold. “CAI” and “CAII” refer to the two clades within CladeA.
Taxonomic treatment
Camellia
shangshii
Taxon classificationFungiEricalesTheaceae
Shi Y.Wang & Q.Fan sp. nov.
660D7181-66C6-59E9-B935-9CCD88209BF1
urn:lsid:ipni.org:names:77375587-1
Type.
China. • Guangdong: Danxiashan Mountain, Renhua County, in mixed forests, 23°29'N, 114°44'E, 294 m a.s.l., 5 November 2023, Z.X.Chen JXZ231101 (holotype: SYS!; isotype:SYS!).
Camellia shangshii sp. nov. A. Flowering and fruiting branch; B. Flowers; C. Bracteoles and sepals; D. Leaf; E. Stamens; F. Pistil; G. Pistil in longitudinal section and ovary in transverse section; H. Fruits and seeds. Illustrated by Rong-En Wu.
Diagnosis.
Camellia shangshii resembles C. grijsii, but it can be distinguished from the latter species by its petals connate at the base, smaller leaves, fewer bracteoles/sepals (6–8 vs. 9–10), and glabrous young branchlets (vs. puberulent).
Camellia shangshii. A. Flowering individual and habitat; B. Flowering branch; C. Fruiting branches; D. Leaves; E. Flower; F. Petals, bracteoles, and sepals, pubescent; G. Flower buds; H. Androecium; I. Pistil; J. Fruit and seed. Photographed by Shiyang Wang, Yinyu Wu, and Xiaowei Yi.
Distribution and ecology.
Presently, the new species is only known from its type locality, Danxiashan Mountain, Guangdong.
Phenology.
Flowering from October to November, fruiting from November to December.
Etymology.
The specific epithet “shangshii” is dedicated in honor of Dr. Shangshi Wu (1904–1947), whose seminal work on the danxia landscape established its theoretical framework.
Conservation status.
The new species is currently known only from Danxiashan Mountain, and most individuals are located within the Danxiashan Natural Reserve, where they are well protected. According to the Guidelines for Using the IUCN Red List Categories and Criteria, v. 16 (IUCN Standards and Petitions Committee 2024), we suggest classifying C. shangshii as Least Concern (LC).
Description.
Shrubs or small trees, 1–3 m tall; branchlets relatively slender, glabrous. Leaves coriaceous; blade oblong-lanceolate, 2.5–5.5 × 0.8–1.9 cm; apex acuminate to caudate-acuminate; base broadly cuneate to nearly rounded; adaxial surface dark green and glossy; abaxial surface light green, both surfaces glabrous; midrib inconspicuous on both surfaces; margin sharply serrate, teeth 1.5–3 mm apart, each with a black glandular tip; petiole 3–7 mm, glabrous. Flowers terminal, white, 1.5–2 cm in diameter; pedicel very short. Bracts 6–8, semicircular to suborbicular, outermost 1.5–2 mm, innermost 8–9 mm, coriaceous, glabrous. Petals 5–6, obovate, 0.7–1.1 × 0.3–0.5 cm, apex emarginate, base connate with stamens for 2–4 mm. Stamens 7–8 mm long, basally connate, glabrous; anthers basifixed. Ovary with transparent, long-coarse hairs. Style 1–2 mm long, glabrous, apex deeply 3-lobed. Capsule subglobose, 1.5–1.7 cm in diameter, 1–3-loculed; pericarp 1–2 mm thick.
Discussion
Phylogenomic analysis based on complete chloroplast genomes strongly supports the reciprocal monophyly of C. shangshii and C. grijsii, resolving them as sister lineages. Despite their close phylogenetic relationship and overall morphological similarity, the two species can be distinguished by several diagnostic morphological traits, most notably petal morphology, leaf size, and flower size (Table 1). Geographically, the new species has been found only in the Danxiashan Mountain region, whereas C. grijsii predominantly occurs in mountainous or hilly terrain in regions including Fujian, Hubei, Sichuan, and Guangxi Provinces. The classification of C. shensiensis as a variety of C. grijsii in “Flora of China” is contradicted by phylogenetic evidence (Fig. 1), which reveals a distant relationship between the two taxa, thus meriting further study. Meanwhile, C. zijinica, another closely related species distributed in the danxia landscape of Guangdong, also exhibits distinct morphological differences, particularly in the structure of its stamen filaments.
Camellia sect. Paracamellia was originally established by Sealy in 1958. Subsequently, based on morphological comparisons, Chang and Ren (1998) proposed its division into sect. Paracamellia and sect. Oleifera, whereas Ming et al. (2000) retained Sealy’s original classification. Recent studies, however, consistently support merging sect. Oleifera back into sect. Paracamellia (Pang et al. 2022; Wu et al. 2022; Zhao et al. 2023; Zhang et al. 2023). Accordingly, C. shangshii is classified under Camellia sect. Paracamellia.
In recent years, over 16 new plant species have been identified in Danxiashan Mountain, such as Pyrus zhaoxuanii (Yi et al. 2025), Diospyros danxiaensis (Tong and Xia 2019), and Primulina danxiaensis (Shen et al. 2010). The unique microclimates, specialized soils, and stark environmental contrasts characteristic of the danxia landscape potentially drive speciation processes and enhance species diversity. However, their precise mechanisms and relative contributions require further investigation and evidentiary support.
Supplementary Material
XML Treatment for Camellia shangshii
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T (2009) trim Al: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25(15): 1972–1973. 10.1093/bioinformatics/btp 348PMC 271234419505945 · doi ↗ · pubmed ↗
- 2Chang HT (1981) A Taxonomy of the Genus Camellia. The Editorial Staff of the Journal of Sun Yat-sen University, Guangzhou, 180 pp.
- 3Chang HT, Ren SX (1998) Theaceae. In: Chang HT (Ed.) Flora Reipublicae Popularis Sinicae (Vol. 49(3)). Science Press, Beijing, 3–34.
- 4Chen SF, Zhou YQ, Chen YR, Gu J (2018) fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34(17): 884–890. 10.1093/bioinformatics/bty 560PMC 612928130423086 · doi ↗ · pubmed ↗
- 5Chen HL, Wei SJ, Tang SQ, Liu Y (2023) Camellia wumingensis (Theaceae), a neglected species from Guangxi, China. Taiwania 68(4): 472–479. 10.6165/tai.2023.68.472 · doi ↗
- 6IUCN Standards and Petitions Committee (2024) Guidelines for Using the IUCN Red List Categories and Criteria. Version 16. Prepared by the Standards and Petitions Committee. https://www.iucnredlist.org/documents/Red List Guidelines.pdf
- 7Jin JJ, Yu WB, Yang JB, Song Y, de Pamphilis CW, Yi TS, Li DZ (2020) Get Organelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biology 21(1): e 241. 10.1186/s 13059-020-02154-5PMC 748811632912315 · doi ↗ · pubmed ↗
- 8Katoh K, Standley DM (2013) MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Molecular Biology and Evolution 30(4): 772–780. 10.1093/molbev/mst 010PMC 360331823329690 · doi ↗ · pubmed ↗