Molecular surveillance of foodborne bacterial pathogens and resistome in food products from Hong Kong
Qiao Hu, Lianwei Ye, Tao Zang, Chen Yang, Xuemei Yang, Ruanyang Sun, Edward Wai Chi Chan, Sheng Chen

TL;DR
This study analyzed raw meat samples from Hong Kong markets to track foodborne bacteria and antibiotic resistance genes, finding that raw foods may spread drug-resistant bacteria.
Contribution
The study identifies novel conjugative plasmids in foodborne bacteria and highlights raw foods as potential reservoirs for antibiotic resistance gene dissemination.
Findings
Raw meat samples from Hong Kong markets were contaminated with multiple foodborne pathogens, including antibiotic-resistant strains.
Novel conjugative plasmids carrying antimicrobial resistance genes were detected in isolated bacterial strains.
Some plasmids originated from other environmental sources or bacterial species, suggesting raw foods as a reservoir for resistance elements.
Abstract
Foodborne infections pose an increasing public health challenge worldwide. The problem has been aggravated by the dissemination of antimicrobial resistance genes among zoonotic pathogens, which results in a sharp increase in antibiotic resistance rate recorded among the major foodborne pathogens. To obtain an overview of the extent to which food products purchased in the markets in Hong Kong were contaminated by foodborne pathogens, we collected 95 raw meat samples from wet markets and isolated 236 bacterial strains of various species, with Escherichia coli being the most dominant species (131 strains). Contamination of food products by multiple foodborne pathogens was commonly observed. These include both Gram-positive and Gram-negative bacteria that exhibit various levels of resistance, with some possessing multiple clinically important antibiotic resistance genes. Seventeen bacterial…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
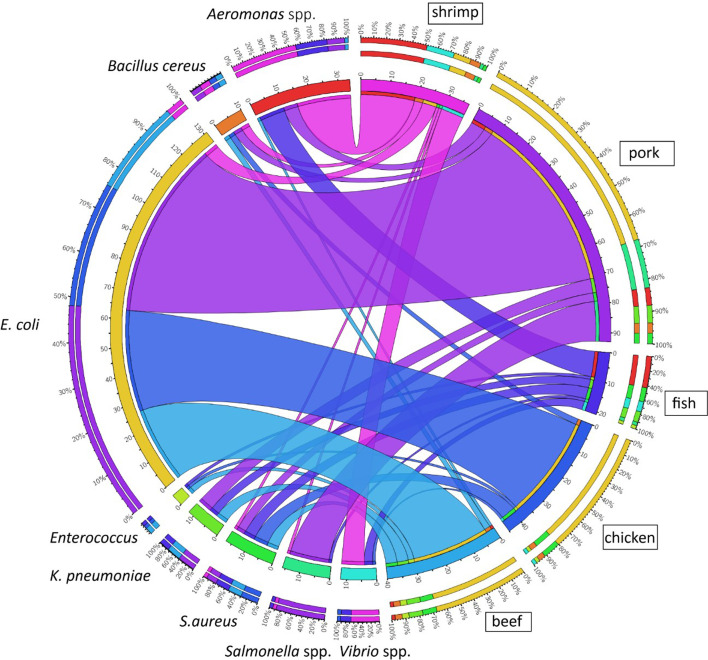 Fig. 1
Fig. 1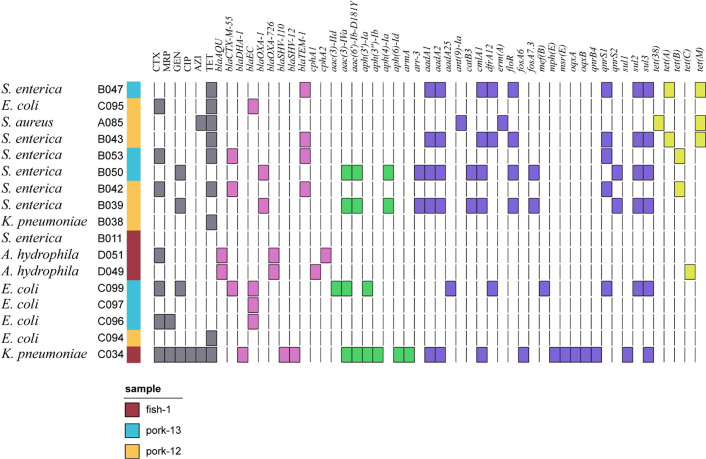 Fig. 2
Fig. 2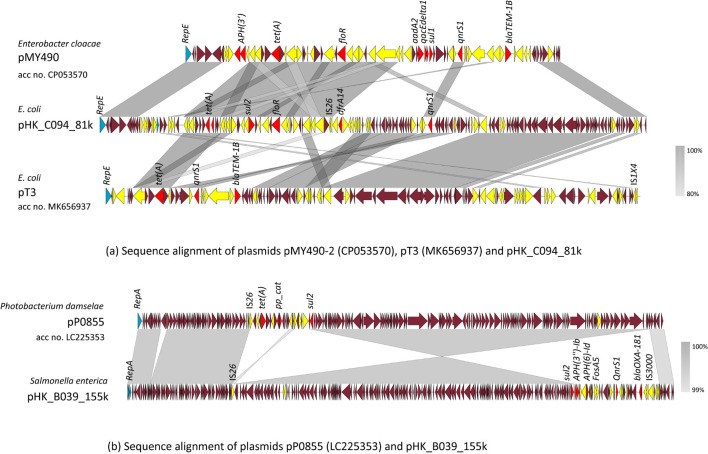 Fig. 3
Fig. 3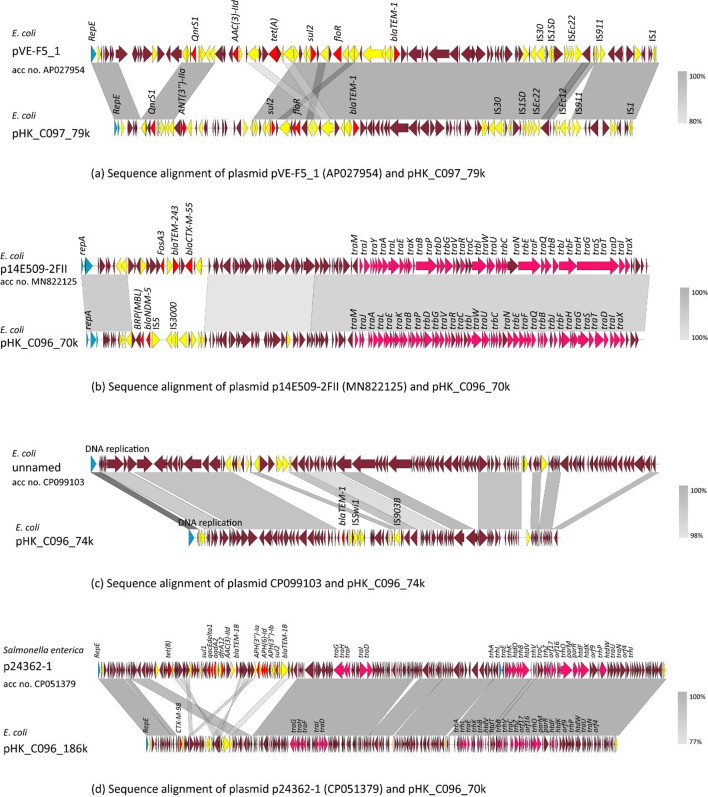 Fig. 4
Fig. 4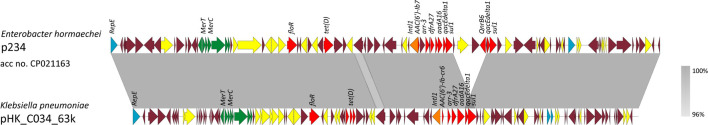 Fig. 5
Fig. 5| No. of sample | Kowloon | New Territories | Hong Kong Island | Total |
|---|---|---|---|---|
| Beef | 10 | 7 | 3 | 20 |
| Chicken | 6 | 6 | 3 | 15 |
| Pork | 9 | 11 | 5 | 25 |
| Fish | 8 | 6 | 1 | 15 |
| Shrimp | 7 | 9 | 4 | 20 |
| No. of strains | Beef | Chicken | Pork | Fish | Shrimp | Total |
|---|---|---|---|---|---|---|
|
| 27 (12) | 36 (12) | 63 (23) | 0 | 5 (3) | 131 |
|
| 1 (1) | 0 | 5 (5) | 9 (6) | 19 (12) | 34 |
|
| 0 | 0 | 14 (7) | 1 (1) | 1 (1) | 16 |
|
| 2 (1) | 2 (1) | 3 (2) | 0 | 3 (2) | 10 |
|
| 4 (4) | 5 (4) | 3 (3) | 4 (4) | 1 (1) | 17 |
|
| 4 (4) | 0 | 5 (4) | 3 (2) | 0 | 12 |
|
| 0 | 1 (1) | 0 | 3 (3) | 8 (4) | 12 |
|
| 2 (1) | 1 (1) | 0 | 1 (1) | 0 | 4 |
| Total | 40 | 45 | 93 | 21 | 37 | 236 |
| Rate of resistance | MRP | CTX | CAZ | TIG | TET | CIP | GEN | ATM | CST |
|---|---|---|---|---|---|---|---|---|---|
|
| 17/131 (13.0%) | 64/131 | 11/44 (25.0%) | 1/72 (1.4%) |
| 66/131 (50.4%) | 18/44 (40.9%) |
| 16/131 (12.2%) |
|
| 3/12 | 7/12 (58.3%) | 4/12 (33.3%) | 0/12 | 10/12 (83.3%) |
| 3/12 (25.0%) | 4/12 (33.3%) | 3/12 (25.0%) |
| 0/33 | 22/33 (66.7%) | 15/33 (45.5%) | 2/33 (6.1%) | 22/33 (66.7%) | 5/33 (15.1%) | 1/33 (3.0%) | 6/33 (18.2%) |
| |
| 1/16 (6.2%) | 3/16 (18.7%) |
| 1/16 (6.2%) |
| 6/16 (37.5%) |
|
| 4/16 (25.0%) | |
| 0/12 | 4/12 (33.3%) |
| 0/12 | 3/12 (25.0%) | 2/12 (16.7%) |
|
|
| |
|
| 1/10 (10.0%) |
|
| 0/10 |
| 2/10 (20.0%) | 0/10 |
|
|
| Samples | Strains | Species | Chromosomal ARGs | Plasmids | Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | Size | G+C (%) | AGRs | Plasmid type | Acc no. | Query cover (%) | Per. identity (%) | ||||
| Pork-12 | A085 |
|
| ||||||||
| B038 |
| pB038_15k | 15393 | 45.5 | ColRNAI_1 | 100 | 99.68 | ||||
| C094 |
|
| pHK_C094_81k | 81110 | 50.8 | IncX1_1, IncR_1 | 47/66 | 99.16/99.66 | |||
| pC094_50k | 50847 | 50.7 | IncFIA(HI1)_1_HI1, IncFIB(K)_1_Kpn3 | 100 | 99.5 | ||||||
| C095 |
|
| pC095_127k | 127742 | 51.4 | IncFIB(pHCM2)_1_pHCM2 | 89 | 99.73 | |||
| pC095_28k | 28886 | 46.8 | IncR_1 | 47 | 99.53 | ||||||
| pC095_79k | 79529 | 51.8 | IncFIA(HI1)_1_HI1, IncX1_1 | 49 | 99.72 | ||||||
| B039 | pB039_16k | 16885 | 49.4 | 58 | 92.35 | ||||||
| pHK_B039_155k | 155988 | 50.7 | IncA/C2_1 | 85 | 99.669 | ||||||
| B042 | pB042_151k | 151750 | 45.2 | IncHI2_1, IncHI2A_1 | 99 | 99.67 | |||||
| B043 | pB043_9k | 9271 | 52.2 | ColRNAI_1, Col440II_1 | 99 | 98.32 | |||||
| Pork-13 | C096 |
|
| pHK_C096_186k | 186788 | 47.4 |
| IncHI1A_1, IncHI1B(R27)_1_R27, IncFIA(HI1)_1_HI1 | 91 | 99.55 | |
| pHK_C096_74k | 73915 | 47.1 |
| IncFIB(pHCM2)_1_pHCM2 | 81 | 99.02 | |||||
| pHK_C096_70k | 70062 | 52.3 | IncFII(pHN7A8)_1_pHN7A8 | 89 | 99.7 | ||||||
| pC096_73k | 73056 | 48.4 | IncY_1 | 79 | 97.07 | ||||||
| C097 |
|
| pHK_C097_79k | 79108 | 50.0 | IncX1_1, IncFIA(HI1)_1_HI1, IncFIB(K)_1_Kpn3 | 84 | 99.73 | |||
| C099 |
| pC099_90k | 90077 | 50.5 | IncFII_1 | 93 | 99.47 | ||||
| B047 | pB047_9k | 9275 | 52.3 | ColRNAI_1, Col440II_1 | 99 | 98.28 | |||||
| B050 | pB050_156k | 156050 | 50.7 | 79 | 99.68 | ||||||
| pB050_90k | 90570 | 47.4 | 87 | 97.92 | |||||||
| B053 | pB053_151k | 151750 | 45.2 | IncHI2_1, IncHI2A_1 | 99 | 99.67 | |||||
| Fish-1 | C034 |
| pHK_C034_63k | 63038 | 53.2 | IncR_1, IncFIA(HI1)_1_HI1 | 100 | 99.69 | |||
| D049 |
| \ | |||||||||
| D051 |
| \ | |||||||||
| B011 |
| \ | |||||||||
- —General Research Fund of Research Grant Council of the Government of Hong Kong SAR
- —Key Research and Development Program of Guangdong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Antibiotic Resistance in Bacteria · Pharmaceutical and Antibiotic Environmental Impacts
Availability of Data and Materials
The whole-genome sequencing, including individual accession numbers for each strain (SAMN40576005, SAMN40576004, SAMN40576003, SAMN40576002, SAMN40576001, SAMN40576000, SAMN40575999, SAMN40575998, SAMN40575997, SAMN40575996, SAMN40575995, SAMN40575994, SAMN40575993, SAMN40575992, SAMN40575991, SAMN4057590 and SAMN40575989), has been submitted to NCBI under BioProject accession number PRJNA1090847.
Introduction
Food safety has become a major public health concern in recent years, prompting government policy makers, clinicians and researchers to devise effective approaches to prevent and treat foodborne microbial infections. The surveillance data in the USA estimated that 31 major pathogens caused a total of 9.4 million episodes of foodborne illness each year, resulting in 55,961 hospitalizations and 1,351 deaths [1]. Among these foodborne infections, seafood-associated bacterial gastroenteritis was mainly caused by Vibrio parahaemolyticus [23]. The ubiquitous spore-forming bacterium Bacillus cereus is known to cause widespread food poisoning incidents [4]. Detection of infectious agents such as Salmonella spp. and Staphylococcus aureus in various common food products is also increasingly being described, suggesting that food samples are a key reservoir of various opportunistic bacterial pathogens that may cause food poisoning [56]. Through the years, novel synthetic compounds, natural products, peptides and proteins have been exhaustively studied in order to identify novel agents that can be used to combat foodborne pathogens such as Escherichia coli [7]. However, the development and dissemination of phenotypic antimicrobial resistance (AMR) among bacterial pathogens that reside in food samples, different environmental niches and clinical settings have severely compromised efforts to control bacterial infections. Plasmids carrying AMR-encoding genes (ARGs) render bacteria to evolve efficiently into multidrug-resistant (MDR) strains, which pose a severe threat to animal and human health [89]. A variety of mobile resistance and/or virulence genes are now detectable in food animals, meat, manure and various environmental and wastewater samples; hence, the risk by which these genetic elements are transmitted to humans via the food chain has increased significantly. One example is carbapenem-resistant Enterobacteriaceae (CRE) which has been recognized as a cause of difficult-to-treat infections associated with high mortality, particularly those which occur in health-care facilities. Resistance development in CRE has been attributed to excessive clinical use of β-lactams of different generations including penicillin, cephalosporins, carbapenems and monobactams [1012]. Although many efforts have been devoted to the development of new antibiotics and non-traditional antibacterial compounds, few new drugs are available for treatment of nosocomial infections caused by MDR Gram-negative bacterial pathogens, a number of which have been classified as critical priority pathogens by WHO [1315].
This study aims to investigate the epidemiological characteristics of potential bacterial pathogens, as well as their resistance phenotypes, by performing a surveillance of meat products purchased from a wet market in Hong Kong, followed by analysis of their phenotypic and genetic traits with the focus on the potential transmission of different AMR bacterial pathogens and their resistance determinants in the same food sample. Data obtained in this work should facilitate the development of effective food safety policy and intervention strategies to minimize the chance of occurrence of foodborne infections.
Methods
Bacterial isolation
The sampling locations represented typical wet markets in Kowloon, the New Territories (including both village-based markets and urban town centres) and Hong Kong Island. Samples were collected between May and November 2022, covering late spring to autumn, when ambient temperature may influence bacterial contamination levels. Five types of fresh meat, including beef (20), chicken (15), pork (25), freshwater fish (15) and shrimp (20), were collected and transported to the laboratory immediately (Table 1). The sample size (95 raw meat samples) provides a snapshot rather than population-level estimates. The different numbers of food types reflected availability and sales volume in local wet markets during sampling, rather than deliberate stratified design. Briefly, 5 g of each sample was aseptically transferred to 45 ml of PBS buffer, followed by homogenization and incubation in enriching broth for 18–24 h; the culture was then inoculated onto LB agar plates. A total of five bacterial species were studied: S. aureus, B. cereus, Salmonella spp., E. coli and Vibrio spp. Enrichment broths for different pathogens were processed in parallel rather than sequentially to avoid competitive exclusion. Selective media containing antibiotics of breakpoint concentration were applied following standard workflows (XLT4 for Salmonella, Baird-Parker agar for S. aureus, TCBS for Vibrio and MYP for B. cereus). Only one colony of typical morphology was picked from each plate for downstream analyses; therefore, within-sample strain diversity may be underestimated. After purification, the strains were identified by MALDI-TOF MS using a Bruker UltrafleXtreme MALDI-TOF/TOF MS. Bacterial strains were preserved in glycerol stock solution (20% v/v) and stored at −80 °C for subsequent genetic and phenotypic characterization.
Antimicrobial susceptibility test
The minimal inhibitory concentrations (MICs) of the foodborne isolates were determined by the agar dilution, as well as the broth microdilution method according to protocols of the Clinical and Laboratory Standards Institute (CLSI). Representative antibiotics of β-lactams, aminoglycosides and fluoroquinolones were tested: cefotaxime, meropenem, gentamicin, tigecycline, ciprofloxacin, vancomycin, erythromycin, colistin, azithromycin, clindamycin, aztreonam, ceftazidime and tetracycline. Resistance breakpoints were interpreted according to CLSI recommendations. E. coli ATCC 25922 and S. aureus ATCC 29213 were used as the quality control strains.
Nanopore sequencing and bioinformatic analyses
Three samples (pork-12, pork-13 and fish-1) were selected as they contained the highest number of co-existing bacterial species and exhibited multidrug resistance, making them suitable for investigating plasmid structures and potential ARG exchange. The selection aimed to maximize genomic diversity rather than represent all food types. A total of 17 strains of various species recovered from 3 food samples (pork-12, pork-13 and fish-1) were subjected to nanopore sequencing. Genomic DNAs were extracted by using QIAamp DNA Kits (QIAGEN, Inc., Netherlands) according to the manufacturer’s protocols. The DNA concentration and purity were quantified with the Qubit fluorometer and Thermo Scientific Nanodrop spectrophotometer, respectively.
For sequencing in the MinION platform (Oxford Nanopore Technologies, Oxford, UK)[16], the library was prepared using the Rapid Barcoding Kit 96 V14 (SQK-RBK114.96), following the manufacturer’s protocols. Sequencing was then performed on the MinION platform using the R10.4.1 flow cell, and the procedure was allowed to run for 72 h. The MinKNOW software was applied to collect raw sequencing data from MinION. POD5 files were base-called using Dorado (Oxford Nanopore Technologies) and then subjected to barcode trimming with Guppy. Alignment of the plasmids and nanopore long reads was visualized by the Easyfig tools [17]. ARGs were identified using Abricate (version 1.0.0) (Seemann)[18] against the Comprehensive Antibiotic Resistance Database [19] and NCBI AMRFinderPlus [20].
Results
Prevalence of foodborne opportunistic pathogens in 95 raw meat samples
A total of 236 bacterial strains were isolated from 95 raw meat samples (Table 1). There were 131 E. coli strains detectable in 50 samples except fish; 16 Salmonella spp. isolates were recovered from 9 samples, 17 S. aureus strains were isolated from 16 samples, and 12 Vibrio spp. isolates were recovered from 8 samples (Table 2, Fig. 1). The rate of recovery of B. cereus was relatively low, as only 10 strains could be isolated from 6 of the 95 meat samples. In addition to the target organisms that we intended to investigate in the original study plan, other bacterial species including 34 Aeromonas spp., 12 Klebsiella pneumoniae and 4 Enterococcus faecalis strains were persistently recoverable from various selective media. E. coli was the most abundant species of all the isolates, with almost half (63 out of 131) originating from pork. Notably, pork was the food product from which the majority of Salmonella enterica were isolated. On the other hand, although it is commonly regarded as a poultry-associated bacteria, Salmonella could not be recovered in the 15 chicken samples. Of all the 236 meat-borne strains, pork was the most predominant source of bacterial contamination with a total of 93 bacterial strains being isolated from 24 pork samples. Among them, E. coli isolates were found in 23 pork samples, and 1 such sample, pork-12, was found to be contaminated with 4 different species of bacteria, namely S. aureus (1), K. pneumoniae (1), E. coli (2) and S. enterica (4). Another sample, pork-13, was found to be contaminated with three E. coli and three S. enterica strains. Moreover, the 15 raw chicken samples were found to harbour a total of 45 bacterial isolates, the majority of which were E. coli (36) and S. aureus (5). Furthermore, 40 strains were recovered from beef samples, 37 from shrimp samples and 21 from 7 freshwater fish samples. Like pork and chicken, the main organism recovered from beef was E. coli (27), but other bacterial species were also detectable, including four S. aureus strains, four K. pneumoniae strains and two each of B. cereus and E. faecalis. In aquatic products such as shrimps and freshwater fishes, Aeromonas spp. (19 and 9, respectively) was the most prevalent species, followed by strains of the Vibrio spp. (8 and 3, respectively).
The relative abundance of bacterial strains of various species recovered from different types of meat samples. Visualization by Circos [42]: the right half consists of five categories of meat samples, and the left half consists of strains of eight groups of bacterial species recovered from each food type. The degree of association between each food sample and different bacterial species is shown.
Phenotypic AMR characteristics of meat-borne bacterial strains
Analysis of MICs of 131 E. coli strains showed that the AMR rate of these strains was very high, with about half of the strains (64 out of 131 and 66 out of 131) being resistant towards cefotaxime and ciprofloxacin, respectively. Resistance to meropenem and tigecycline was relatively low, accounting for 13% (17 out of 131) and 2% (3 out of 131) of the E. coli strains, respectively. Colistin resistance (MIC ≥4 mg l^−1^, broth microdilution) was detected in 12.2% (16 out of 131) of E. coli isolates. However, another 108 strains also exhibited intermediate resistance to this antibiotic (Table 3). Among the five types of meats, chicken-borne E. coli strains exhibited the highest rate of resistance to cefotaxime, meropenem and ciprofloxacin. The 27 beef-derived E. coli strains exhibited a higher rate of resistance towards ciprofloxacin than the other 2 β-lactam drugs (CTX, MRP), as did the 63 strains recovered from pork samples. On the other hand, the 16 Salmonella strains displayed the highest rate of resistance towards ciprofloxacin [37.5% (6 out of 16)], followed by 18.8% (3 out of 16), 6.25% (1 out of 16) and 6.25% (1 out of 16), to tigecycline, meropenem and cefotaxime, respectively. Furthermore, Aeromonas spp. displayed the highest rate of resistance to tetracycline and cefotaxime (22 out of 33 or 66.7%). A lower level of AMR was observed among the Vibrio spp. and B. cereus strains, with 33.3% (4 out of 12) of Vibrio strains being cefotaxime resistant, and 20% (2 out of 10) B. cereus isolates exhibiting ciprofloxacin resistance. Importantly, the 12 K. pneumoniae strains recovered in this work were resistant to multiple antibiotics, with cefotaxime and tetracycline being the least effective drugs in case these K. pneumoniae strains cause foodborne infections. Furthermore, one strain recovered from a fish sample was resistant to all test antibiotics except tigecycline (Table 3). All in all, our data showed that the foodborne bacteria were mainly susceptible to meropenem and tigecycline but exhibited different levels of resistance to other antimicrobial agents.
Genetic characteristics of bacterial strains of bacteria isolated from the same food sample
To investigate the genetic relationship and transmission characteristics of AMR genes among different bacterial strains isolated from the same food sample, we conducted complete genome and plasmid sequence analysis of all bacterial strains isolated from three representative samples. A total of 17 strains were isolated from these 3 meat samples, namely pork-12, pork-13 and fish-1. Seven bacterial strains belonging to five different species were isolated from pork-12; six strains of two species were isolated from pork-13, and four strains that belonged to three species were isolated from fish-1. Phenotypically, all the 17 strains exhibited resistance towards tetracycline and cefotaxime, and among them, the K. pneumoniae strain C034 (acc no. SAMN40576002) was resistant to almost all the test drugs (Fig. 2). Results of the pairwise comparison between strains with or without plasmids suggested no sign of plasmid dissemination among strains of different species isolated from one or different samples. Apart from the resistance genes located in mobile elements, ARGs in the chromosomal regions of various isolates were identified by matching with the NCBI database; such chromosomal ARGs were found to encode resistance to different classes of antibiotics, such as beta-lactam, carbapenem, cephalosporin, quinolones and kanamycin (Table 4).
AMR phenotypes and ARGs located in the chromosome of meat-borne isolates. A total of 17 isolates recovered from three samples represented by three different colours are shown on the left. The corresponding AMR phenotypes and genotypes detectable in each bacterial chromosome are shown on the right. CTX, cefotaxime; MRP, meropenem; GEN, gentamicin; CIP, ciprofloxacin; AZI, azithromycin; TET, tetracycline.
Bacterial strains isolated from three representative samples were analysed in detail. The first sample, pork-12, was collected from a wet market located in northern Hong Kong. Nine bacterial strains were isolated in the raw meat, including one Gram-positive isolate (S. aureus A085, acc no. SAMN40575989) and eight Gram-negative strains, of which seven strains, except one Aeromonas hydrophila, were sequenced. Alignment of plasmids showed that each of the seven Gram-negative bacterial strains carried at least one plasmid, most of which were highly analogous with previously reported data, but a few were recognized as novel (Fig. 3). The S. aureus strain A085 was found to carry a number of ARGs in the chromosome, but no plasmid was detectable. One of the Gram-negative strains, K. pneumoniae strain B038 (acc no. SAMN40575990), carried a few intrinsic ARGs and a small ColRNAI_1 plasmid that did not contain any ARGs and was structurally similar to a plasmid pAZS099-2 (GenBank: CP086763). E. coli strain C094 (acc no. SAMN40575991) was sensitive to most antimicrobials including cefotaxime and meropenem. This strain carried the bla_EC-15_ gene and a new plasmid, pHK_C094_81 k, which appears to be a fusion plasmid formed by integration of specific segments of the plasmids pMY490 and pT3, with a low coverage of, respectively, 47 and 66%, but >99% identity to these two plasmids. The resistance gene dfrA14 found in this plasmid was probably derived from a newly inserted genetic fragment as neither of the two reference plasmids bears this gene. The E. coli strain C095 (acc no. SAMN40575992) was found to harbour a blaEC gene located on chromosome, as well as three plasmids, namely pC095_127k, pC095_28k and pC095_79k, and the plasmids pC095_127k and pC095_28k did not carry any ARG and exhibited a certain degree of sequence homology with plasmids pB379_2 (89%, GenBank: CP090457) and pYLPM4b (47%, GenBank: CP074003), respectively. Plasmid, pC095_79k, was found to carry multiple ARGs including blaCTX-M-55; it is likely that this plasmid formed by fusion between the IncFIA(HI1)1_HI1 plasmid and an IncX1_1 plasmid. In the genome of S. Derby B039 (acc no. SAMN40575993), there were a variety of chromosomal genes including *bla_OXA-1*, qnrS2, arr-3, catB3, aac(6')-Ib-cr, aph(4)-Ia, aac(3)-IVa and cmlA1. This strain also carried an MDR plasmid, pHK_B039_155k, which exhibited 85% homology with the plasmid pP0855 from a Photobacterium damselae strain but differed dramatically in the MDR region that carried different types of ARGs, as very few ARGs were found in pHK_B039_155k. In another two Salmonella strains, B042 (acc no. SAMN40575994) and B043 (acc no. SAMN40575995), multiple ARGs including blaCTX-M-55 were detectable in the chromosome. Each of these two strains also carried one plasmid that was highly analogous with (KPC-2)_IncHI2 (GenBank: CP050163) and pN18S0406-4 (GenBank: CP082690), respectively.
Schematic representation of the structure of two plasmids recovered from pork-12-derived strains. Results of sequence alignments between (a) Enterobacter cloacae plasmid pMY490-2 (CP053570), E. coli plasmid pT3 (MK656937) and pHK_C094_81k and (b) P. damselae plasmid pP0855 (LC225353) and pHK_B039_155k are shown. CDSs without labels represent hypothetical proteins. The shadow parallelograms denote genetic regions that exhibit sequence homology. Light shadow denotes regions with a lower level of sequence identity. Arrows indicate CDSs, with arrowheads indicating the direction of transcription: red, antibiotic resistance-encoding genes; yellow, mobile elements; blue, replication protein-encoding genes; deep burgundy, maintenance/stability genes, or genes that encode hypothetical proteins.
The second sample, pork-13, was collected from an eastern Hong Kong market, from which three E. coli strains and three Salmonella strains were isolated. E. coli strain C097 (acc no. SAMN40575997) remained sensitive to most antimicrobial drugs including the cephalosporins, except that low-level resistance to ciprofloxacin was detected. The gene bla_EC_ could be detected in the genome of C097. A plasmid – pHK_C097_79k – was recovered from this strain and found to exhibit a coverage rate of 84% with the plasmid pVE-F5_1 (Fig. 4), which was recovered from a colistin-resistant E. coli isolated from Vietnam; pHK_C097_79k also carried five AMR genes including bla_TEM-1_, as well as several mobile elements, such as IS1 and ISEc22. With a size of 79,108 bp, the plasmid type of pHK_C097_79k was presented as IncX1_1/IncFIA(HI1)_1_HI1/IncFIB(K)_1_Kpn3 (Table 4).
Schematic representation of the structure of four plasmids recovered from pork-13-derived isolates. Results of sequence alignments between (a) E. coli plasmid pVE-F5_1 (AP027954) and pHK_C097_79k, (b) E. coli plasmid p14E509-2FII (MN822125) and pHK_C096_70k, (c) E. coli plasmid CP099103 and pHK_C096_74k and (d) S. enterica plasmid p24362-1 (CP051379) and pHK_C096_186k are shown. CDSs without labels represent hypothetical proteins. The shadow parallelograms denote genetic regions that exhibit sequence homology. Light shadow denotes regions with a lower level of sequence identity. Arrows indicate CDSs, with arrowheads indicating the direction of transcription: red, antibiotic resistance-encoding genes; yellow, mobile elements; blue, replication protein-encoding genes; pink, genes associated with the tra loci; deep burgundy, maintenance/stability genes, or genes that encode hypothetical proteins.
Another E. coli isolate, C096 (acc no. SAMN40575996), also carried the beta-lactamase-encoding gene blaEC-13 in the chromosome. C096 exhibited broad-spectrum resistance to the third-generation cephalosporins cefotaxime and ceftazidime, and the carbapenem antibiotic of meropenem. Consistently, three MDR plasmids which carried the ARGs blaCTX-M-98, bla_TEM-1_, and bla_NDM-5_/ble_MBL_, respectively, could be detected in this strain. The size of these three plasmids, namely pHK_C096_186k, pHK_C096_74k and pHK_C096_70k, was 186,788, 73,915 and 70,062 bp, with a G+C content of 47.4 mol% and 47.1–52.3 mol%, respectively. Plasmid pHK_C096_186k was shown to be homologous to plasmid p24362-1 [21], which was recovered from an S. enterica strain previously. However, the MDR region in pHK_C096_186k was structurally very different from that of p24362-1 [21], suggesting that evolution of this type of plasmid occurs in different organisms. A bla_CTX-M-98_ gene was found located in the MDR region of pHK_C096_186k. Plasmid pHK_C096_70k was an IncFII plasmid containing a blaNDM-5 gene-bearing IS3000 mobile element. It exhibited a high degree of sequence homology with plasmid p14E509-2FII from an E. coli strain from China, except for the MDR region. The MDR region in plasmid p14E509-2FII contained a blaCTX-M-55 gene rather than a blaNDM-5 gene, suggesting that this plasmid actively evolves. It is also predicted to be a conjugative plasmid which carried the tra locus, thereby enabling it to transfer the carbapenemase gene bla_NDM-5_ to other antibiotic-susceptible organisms. Plasmid pHK_C096_74k exhibited a relatively high degree of homology (81%) to an unnamed plasmid (CP099103.1) from E. coli isolated from a pooled sediment sample collected from the floor of a pig farm in the UK. Unlike the unnamed plasmid that did not carry any AMR gene, pHK_C096_74k carried a blaTEM-1 gene. S. Rissen strain B047 (acc no. SAMN40575999) carried multiple ARGs which were located in the chromosome, as well as a small 9k plasmid; S. Derby strain B050 (acc no. SAMN40576000) carried multiple ARGs that were located in the chromosome, as well as two plasmids, namely pB050_156k and pB050_90k, both of which exhibited over 80% homology with previously reported plasmids; S. enterica (4,5,12:i:-) strain B053 (acc no. SAMN40576001) carried multiple ARGs, including a blaCTX-M-55 gene in the chromosome and a 151k plasmid which has been reported previously (Table 4).
Lastly, the third sample, fish-1, which was purchased from a market in the northwestern part of Hong Kong, was found to harbour one K. pneumoniae strain (C034), two Aeromonas strains, one S. enterica Stanley strain (B011, acc no. SAMN40576005) and one E. faecalis strain (A049). K. pneumoniae strain C034 exhibited the widest resistance spectrum, with a large number of ARGs being identified in the chromosome [bla_DHA-1_, bla_SHV-110_, bla_SHV-12_, aph(3')-Ia, aph(3'')-Ib, aph(6)-Id, aph(4)-Ia and aac(3)-Iva]. In addition, it also carried a plasmid, pHK_C034_63k, with a size of 63,038 bp and 53.2 mol% of G+C content. This plasmid was found to contain multiple aminoglycoside resistance genes [aac(6')-Ib-cr6, aadA16] (Fig. 5). Several types of mobile genetic elements, including MGE IS1006, ISVsa3, IS26, IS6100, IS15DI and IS1A, were identified in pHK_C034_63k, the plasmid type of which was likely to be IncR_1/IncFIA(HI1)_1_HI1 (Table 4). The large number of ARGs detected is consistent with the multidrug resistance phenotype observable in this strain. Alignment between pHK_C034_63k and plasmid p234 harboured by the organism Enterobacter hormaechei (query coverage 100%, per. identity 99.7%) showed near-complete synteny. The only divergent region was a ~4 kb segment absent in pHK_C034_63k, corresponding to a cluster of hypothetical proteins and a partial mobile element in p234. No ARGs or known functional modules were located in this missing region. The other three strains, namely A. hydrophila D049 (acc no. SAMN40576003), A. hydrophila D051 (acc no. SAMN40576004) and S. Stanley B011, did not carry any plasmid, but a few ARGs were found in the chromosome.
Schematic representation of the structure of plasmid pHK_C034_63K of a fish-1-derived K. pneumoniae strain. The result of sequence alignments between E. hormaechei plasmid p234 (CP021163) and pHK_C034_63k is shown. CDSs without labels represent hypothetical proteins. The shadow parallelograms denote genetic regions that exhibit sequence homology. Light shadow denotes regions with a lower level of sequence identity. Arrows indicate CDSs, with arrowheads indicating the direction of transcription: red, antibiotic resistance-encoding genes; yellow, mobile elements; blue, replication protein-encoding genes; orange, integrase gene; green, genes associated with silver and heavy metal resistance; deep burgundy, maintenance/stability genes, or genes that encode hypothetical proteins.
Discussion
This study provides a focused characterization of plasmid structures and ARG contexts in Hong Kong wet-market meat using long-read sequencing. The detection of several previously unreported plasmid fusion structures and distinct ARG combinations highlights regional variation in AMR dissemination dynamics and contributes new genomic evidence specific to Hong Kong’s retail food environment. Among the 236 isolates recovered from 95 raw meats in this survey, E. coli was the most commonly isolated bacterial species in most samples, and pork was the most favourable habitat of this species. Other species generally exhibited low-level prevalence in most of the food samples. B. cereus strains were identified in 6 out of the 95 samples as they habitually contaminate rice and dairy products and may cause diarrhoea or emetic syndrome [2223]. The main source of 16 S. enterica strains was also pork (14 out of 16) rather than chicken. From a global viewpoint, reports from the CDC and EFSA have confirmed that pig meat was most frequently associated with S. enterica serovar Typhimurium [2426]. Several K. pneumoniae and E. faecalis isolates, accompanied by several other unique species, colonized a small proportion of meats, which is likely underrepresented as they were not intended to be included in the isolation process. Nonetheless, previous reports have illustrated the potential by which E. faecalis transfers genetic elements to human strains following consumption of contaminated meat, and that E. faecalis and E. faecium were found in 69.5 and 11.3% of the red meat samples (fresh beef and pork), respectively [2728]. The prevalence of Aeromonas spp. in fish and shrimps (40 and 60%) was quite high when compared to another study focused on Norwegian ready-to-eat seafood, where the maximum rate appeared to be 17% in retail sushi [29]. Although the sample size in this study was not large enough, it is reasonable to speculate that this discrepancy is due to the fact that the warm weather in Hong Kong and its location in the sub-tropical zone favour the survival and circulation of various pathogenic organisms. The sample size (95 raw meat samples) provides a snapshot rather than population-level estimates. The different numbers of food types reflected availability and sales volume in local wet markets during sampling, rather than deliberate stratified design. As the study was exploratory, results should not be generalized to the entire Hong Kong food supply. A larger, longitudinal dataset would be needed to assess temporal or territory-wide epidemiological trends.
AMR is one of the most important concerns in current efforts to combat foodborne bacterial infections. A Canadian study showed that E. coli strains recovered from broiler chicken exhibited a high rate of resistance to aminoglycosides, β-lactams and tetracyclines, yet only 0.2% of such strains were resistant to ciprofloxacin. The percentage of Salmonella isolates that were resistant to β-lactams, macrolides, quinolones and tetracyclines was similar but generally lower than that of E. coli [30]. In a study in TN, USA, bulk tank milk, faeces and soil samples were collected in four dairy cattle farms, and antimicrobial tests of bacteria isolated from such samples confirmed that E. coli exhibited the highest rate of resistance (73.9%) to TET, followed by TGC (cefotaxime) (20.5%) and NAL (nalidixic acid) (8%), although the proportion of AMR E. coli varied by farms and samples [31]. In our study, meat-borne E. coli generally exhibited higher rates of resistance to cefotaxime (48.9%, 64 out of 131) but a comparatively modest rate of resistance to the other two β-lactams, meropenem and ceftazidime. Previously, K. pneumoniae was detected in raw chicken and pork liver at a frequency of 27% (16 of 60) in Singapore; in the same study, half of the K. pneumoniae strains that exhibited the MDR phenotype isolates were isolated from meat samples (5 out of 10) [32]. An even higher rate of isolation (81.8%) of K. pneumoniae strains in Greek meat products was reported, with bla_NDM_ (61.5%) and bla_OXA-48_-like (30.8%) genes being the most frequently detected resistance genes [33]. In this project, we also studied the genetic features of 17 bacterial isolates recovered from 3 different meat samples, with a focus on the genetic context of the plasmids that harboured resistance genes and mobile elements recoverable from these strains. Bioinformatic analysis suggested that, among the strains recovered from samples gathered from distinct local markets and organisms isolated from the same food sample, evidence of direct plasmid transmission could not be obtained. Instead, six new plasmids with unique structures were detectable among such strains. Among the three E. coli strains, we recovered a total of five plasmids, among which the bla_TEM-1_-bearing pHK_C097_79k did not match with any plasmid replicon type in the PlasmidFinder database [34]. A number of insertion elements including IS30, IS1, IS66 and IS3 were also found in this plasmid; such elements were found to have acquired a known copy-paste mechanism that generates a transient double‐strand circular DNA intermediate [3538]. Moreover, another three plasmids carried by the same E. coli strain (C096) were found to belong to the IncHI1A, IncFIB and IncFII types, respectively; these plasmids contained various β-lactamase genes. Similarly, another study in China described a plasmid, pHeBE7, which carried the bla_CTX-M-98b_, bla_TEM-1_, rmtB and traT genes and could spread to geographically diverse regions through human or food animals [39]. In addition, the bla_NDM-5_ and tet(X4) genes co-harboured by the clinically isolated E. coli ST648 strain were found to encode resistance to carbapenem and tigecycline, respectively, indicating that plasmid-borne resistance genes in various superbugs can pose a serious threat to public health [40]. In Singapore, researchers reported the complete nucleotide sequence of a transferable plasmid harbouring the bla_TEM-176_ and mcr-5.1 genes in an E. coli strain isolated from ready-to-eat chicken rice [41]. The Salmonella plasmid pHK_B039_155k identified in this work was categorized as the IncA/C2 type and found to carry an array of ARGs [blaOXA-181, qnrS1, fosA5, aph(6)-Id, aph(3'')-Ib and sul2] that encode resistance to five classes of antimicrobials, but phenotypically, the strain was resistant to gentamicin only. Factors that regulate the expression of plasmid-borne antibiotic resistance genes in bacterial pathogens need to be identified in future experiments. Also, because only isolates from Hong Kong Island were subjected to long-read sequencing, territorial comparison (HK vs. Kowloon vs. NT) is limited. Inclusion of additional sequenced isolates from other districts would improve the geographic resolution of plasmid and ARG distribution.
In conclusion, we have conducted a general surveillance of foodborne pathogens collected in Hong Kong markets and depicted the AMR profiles of selected bacterial strains. Several plasmids harboured ARGs and mobile elements capable of mediating horizontal transfer. Although this study did not experimentally assess transferability, the structures identified suggest plausible vehicles for ARG dissemination, warranting future functional validation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Scallan E Hoekstra RM Angulo FJ Tauxe RV Widdowson MA et al Foodborne illness acquired in the United States—major pathogens Emerg Infect Dis 20111771510.3201/eid 1701.P 1110121192848 PMC 3375761 · doi ↗ · pubmed ↗
- 2Dutta D Kaushik A Kumar D Bag S Foodborne pathogenic vibrios: antimicrobial resistance Front Microbiol 20211263833110.3389/fmicb.2021.63833134276582 PMC 8278402 · doi ↗ · pubmed ↗
- 3Newton A Kendall M Vugia DJ Henao OL Mahon BE Increasing rates of Vibriosis in the United States, 1996–2010: review of surveillance data from 2 systems Clin Infect Dis 201254 S 391S 39510.1093/cid/cis 24322572659 PMC 4604744 · doi ↗ · pubmed ↗
- 4Enosi Tuipulotu D Mathur A Ngo C Man SM Bacillus cereus: epidemiology, virulence factors, and host-pathogen interactions Trends Microbiol 20212945847110.1016/j.tim.2020.09.00333004259 · doi ↗ · pubmed ↗
- 5Chintagari S Hazard N Edwards G Jadeja R Janes M Risks associated with fish and seafood Microbiol Spectr 2017510.1128/microbiolspec.pfs-0013-2016 PMC 1168744428185612 · doi ↗ · pubmed ↗
- 6Kadariya J Smith TC Thapaliya D Staphylococcus aureus and staphylococcal food-borne disease: an ongoing challenge in public health Biomed Res Int 2014201482796510.1155/2014/82796524804250 PMC 3988705 · doi ↗ · pubmed ↗
- 7Lai EPC Iqbal Z Avis TJ Combating antimicrobial resistance in foodborne microorganisms J Food Prot 20167932133610.4315/0362-028X.JFP-15-02326818996 · doi ↗ · pubmed ↗
- 8Laxminarayan R Duse A Wattal C Zaidi AKM Wertheim HFL et al Antibiotic resistance-the need for global solutions Lancet Infect Dis 2013131057109810.1016/S 1473-3099(13)70318-924252483 · doi ↗ · pubmed ↗