A secreted C-type lectin in Periplaneta americana functioning in antibacterial activity, innate immune signaling and leg regeneration
Xiaoxuan Liu, Nan Sun, Shuang Geng, Shuqi Xian, Xiaojuan Wu, Ying Huang, Yechun Pei

TL;DR
A secreted C-type lectin in American cockroaches helps fight bacteria, supports immune signaling, and aids in leg regeneration.
Contribution
The study identifies and characterizes two PaSCLec isoforms in Periplaneta americana, revealing their roles in immunity and regeneration.
Findings
PaSCLec is highly expressed in hemolymph and upregulated after bacterial infection.
PaSCLec proteins bind to microbial components and agglutinate bacteria, enhancing immune responses.
PaSCLec knockdown impairs leg regeneration and reduces immune gene expression.
Abstract
C-type lectins are important pattern-recognition receptors that play essential roles in innate immune responses by recognizing pathogen-associated molecular patterns. However, their biological functions in Periplaneta americana have not been systematically investigated. Two PaSCLec isoforms predicted by RNA-seq, designated PaSCL-Ad and PaSCL-Reg, were identified using Rapid Amplification of cDNA Ends (RACE) and Nanopore sequencing. Tissue-specific expression and inducible expression following bacterial challenge were analyzed. Recombinant PaSCL-Ad and PaSCL-Reg proteins were produced to examine their binding activities to microbial polysaccharides, bacterial agglutination, antimicrobial effects, and membrane-disruptive activity. Hemocyte phagocytosis was evaluated by immunocytochemical analysis. RNA interference was employed to assess the role of PaSCLec in immune regulation and leg…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
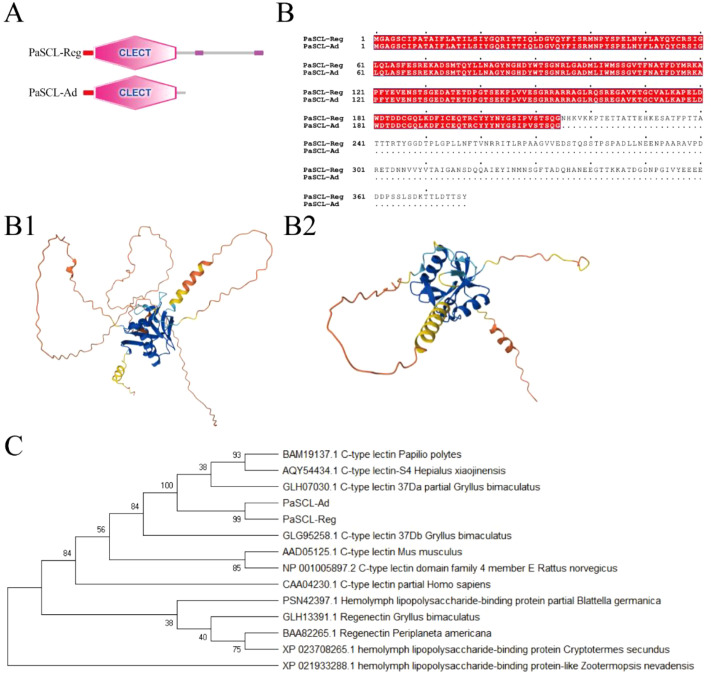 Figure 1
Figure 1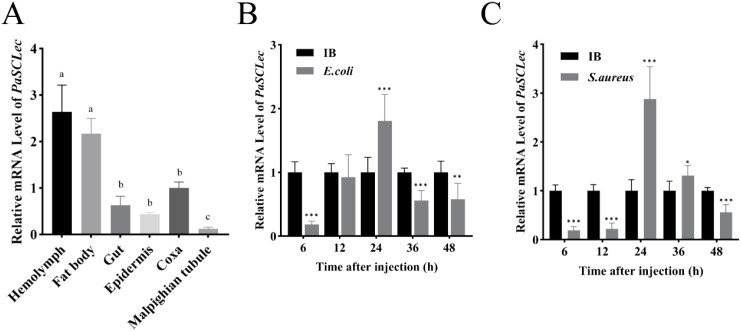 Figure 2
Figure 2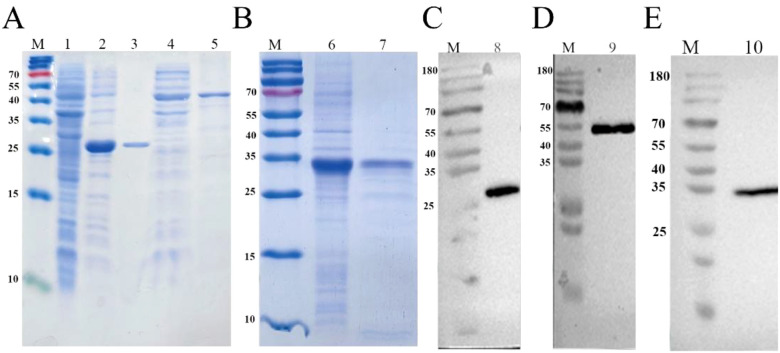 Figure 3
Figure 3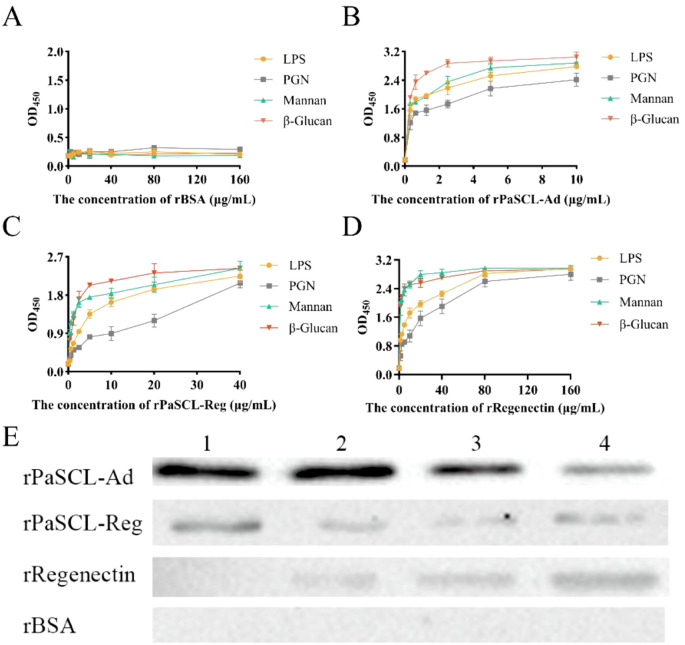 Figure 4
Figure 4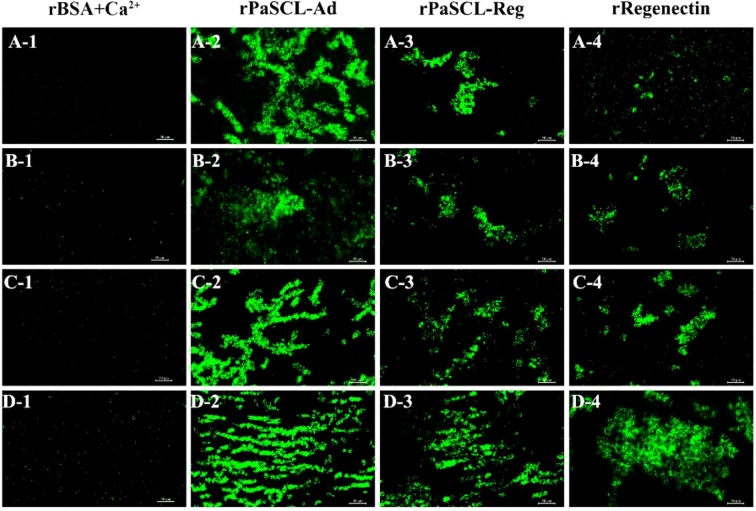 Figure 5
Figure 5 Figure 6
Figure 6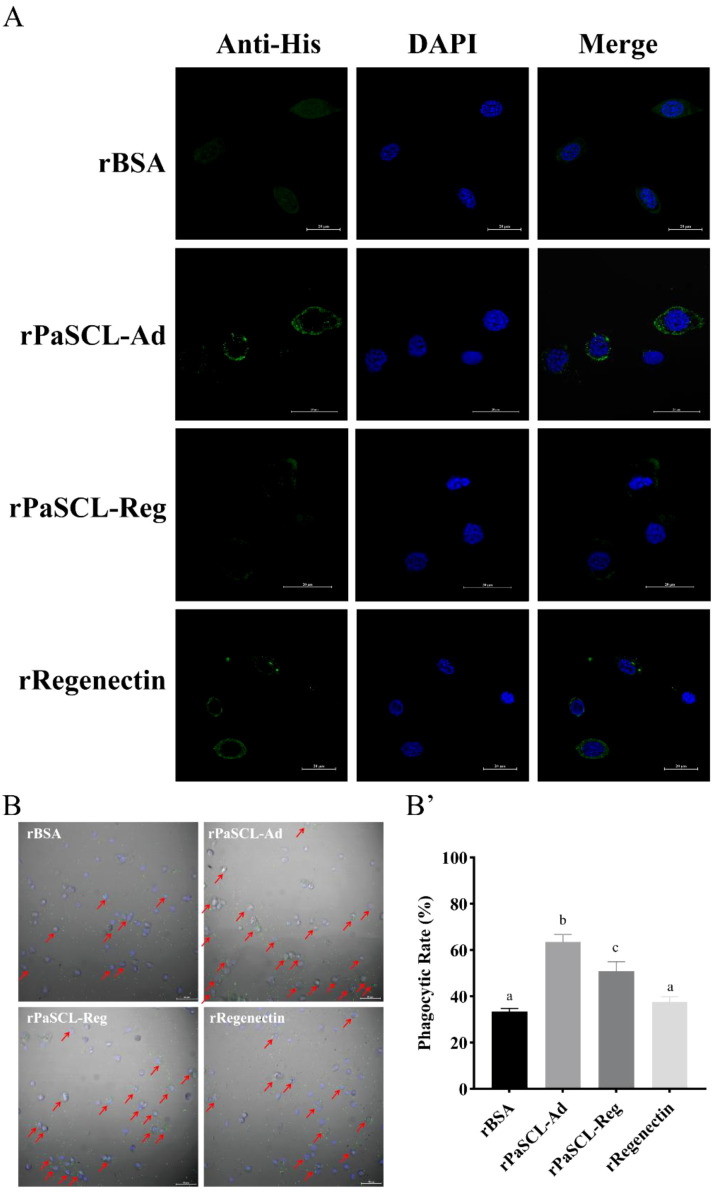 Figure 7
Figure 7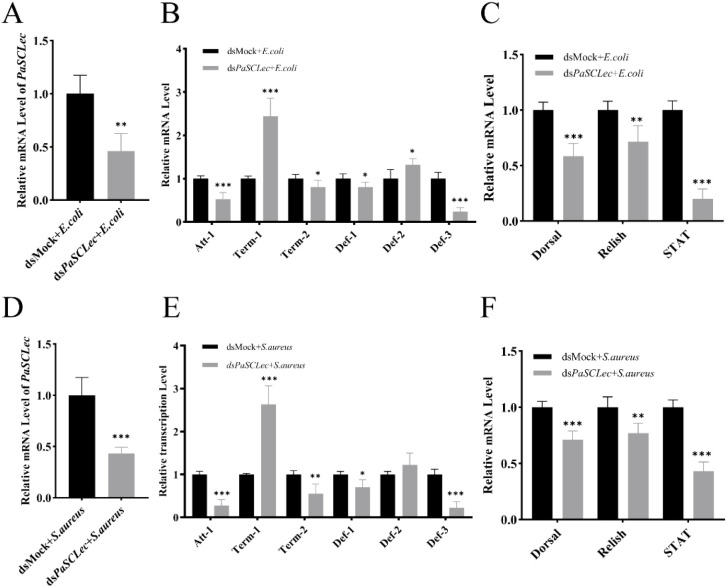 Figure 8
Figure 8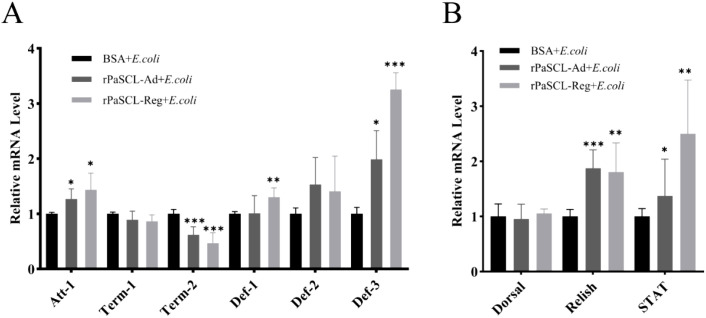 Figure 9
Figure 9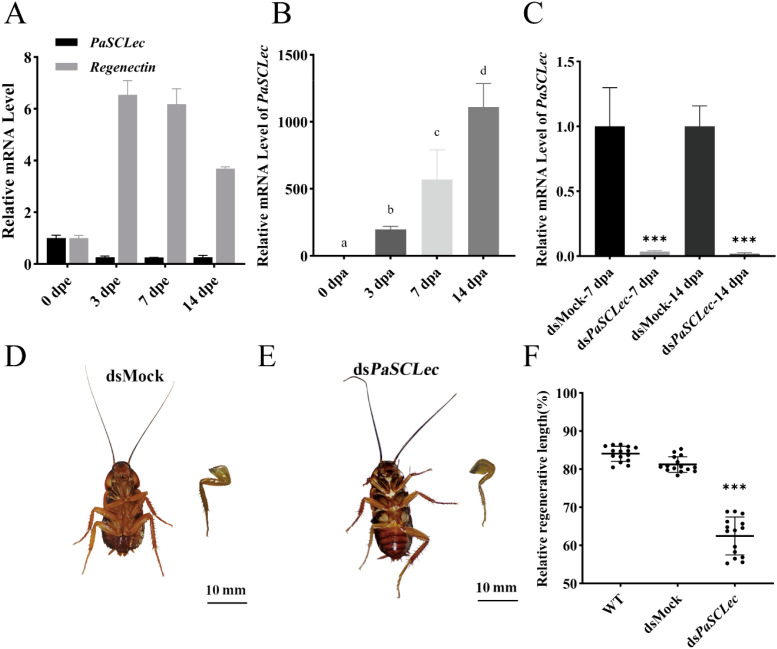 Figure 10
Figure 10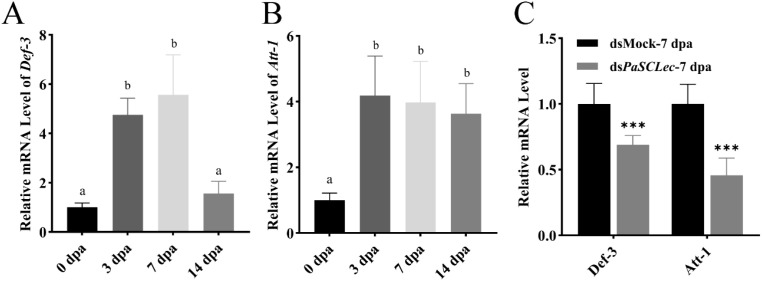 Figure 11
Figure 11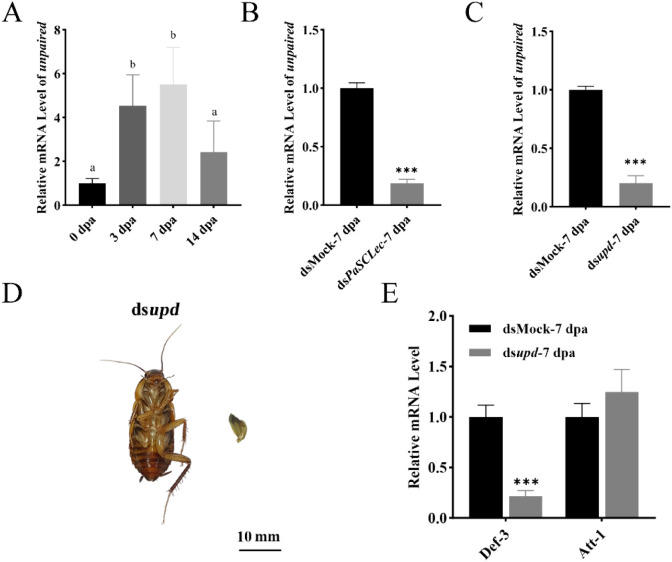 Figure 12
Figure 12- —National Natural Science Foundation of China10.13039/501100001809
- —Major Science and Technology Project of Hainan Province10.13039/501100013072
- —Natural Science Foundation of Hainan Province10.13039/501100004761
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Glycosylation and Glycoproteins Research · Immunotherapy and Immune Responses
Introduction
1
C-type lectins (CTLs), a major class of pattern recognition receptors (PRRs), play essential roles in innate immunity (1, 2) and occur either as transmembrane receptors or as soluble secreted proteins (3). Previous studies have demonstrated that CTLs participate in both immune responses and developmental processes in invertebrates. (4, 5). In Tribolium castaneum, multiple CTLs have been identified and shown to contribute to immune responses and development (6–9). In Drosophila melanogaster, DL2 and DL3 can agglutinate E. coli and promote encapsulation and melanization in vitro (10, 11). In Rhynchophorus ferrugineus, RfCTL27 recognizes Gram-negative bacteria and induces antimicrobial peptide expression to eliminate pathogens (12). In Ostrinia furnacalis, IML-10 promotes hemocyte aggregation by binding directly to hemocyte surfaces, thereby facilitating encapsulation (13). In addition, CLEC3A activates the PI3K–AKT pathway to accelerate cell proliferation. (14). In vitro analyses have also shown that Collectin-11, a soluble C-type lectin, activates EGFR signaling and directly promotes murine melanoma cell proliferation (15). Collectively, these findings indicate that CTLs are critical regulators of both immunity and development.
Insects are invertebrates of substantial economic, ecological, pathological, and medicinal importance (16–18). Periplaneta americana is widely used as a raw material in traditional Chinese medicine (19). Extracts of P. americana exhibit strong regenerative properties (20–22), which may be associated with its remarkable leg regeneration capabilities (23). Previous studies have shown that CTLs from P. americana act as opsonins that mediate immune responses (24–26). Regenectin localizes near newly formed epidermal cells, is secreted into the regenerating leg saccule, and assembles around myoblasts to facilitate the in site formation of leg muscle fibers (27). A potential link exists between immune responses and leg regeneration, and C-type lectins may play a central role in this process. Transcriptome analyses previously identified two PaSCLec isoforms, PaSCL-Reg and PaSCL-Ad, both of which are highly expressed during leg regeneration in P. americana (28). However, the biological function of PaSCLec remain unclear.
Here, we investigated whether the secreted C-type lectin PaSCLec contributes to immune regulation and leg regeneration. We found that bacterial challenge significantly increased the mRNA levels of total PaSCLec (including PaSCL-Reg and PaSCL-Ad) in hemocytes. In addition, both PaSCL-Reg and PaSCL-Ad exhibited bacterial-binding activity, agglutinating capacity, and antibacterial properties. Furthermore, PaSCLec knockdown impaired leg regeneration, likely by modulating the expression of unpaired. These findings enhance our understanding of the roles of secreted C-type lectins in immune responses and leg regeneration, and further expand the functional diversity attributed to CTLs.
Materials and methods
2
Insects
2.1
All Cockroaches were provided by American cockroach breeding farm in Chizhou city, Anhui province, China. The colony was maintained at 27°C with a relative humidity of 70-80% in breathable plastic box, and the animals received lab mice diet and water. To obtain pools of synchronized animals, the oothecae were hatched uniformly and molt numbers were recorded. Newly 6^th^ instar nymphs were selected from the colony and placed in separate plastic boxes, supplied with mice diet and water. The experimental cockroaches were anesthetized with ice for sample collection.
Bioinformatics analysis of PaSCLec
2.2
The full-length transcript sequence of PaSCLec was obtained from cockroach leg by transcriptomic sequencing. Transcript sequence was used for primer design to obtain full-length cDNAs by Rapid Amplification of cDNA Ends (RACE) using the HiScript-TS 5’/3’ RACE Kit (Vazyme, China). A cDNA copy of PaSCLec was obtained using the following primers: 3′RACE (5′- ATGGGCGCTGGAAGCTGTATACCGG-3′) and 5′RACE (5′ -AGGGGTTCATGCGGCTTATGA-3′). The sequence of PaSCLec was subjected to nanopore sequencing by Beijing Tsingke Biotech Co., Ltd. The full-length cDNA sequence of PaSCL-Reg and PaSCL-Ad were amplified by RT-PCR using corresponding primers and re-sequenced for confirmation (Supplementary Table S1). Homology analysis of PaSCL-Reg and PaSCL-AD amino acid sequence were performed using BLAST (http://www.ncbi.nlm.nih.gov/). Multiple sequences alignment analysis was performed using the DNAMAN. The characterizations of protein were predicted on the ExPASY server (http://www.expasy.org/). SignalP 5.0 server was used to predict the signal peptide (https://services.healthtech.dtu.dk/service.php?SignalP-5.0). Domain architecture prediction of the proteins was performed using SMART (http://smart.embl-heidelberg.de/). MEGA 5 was used for phylogenetic analysis. The modeling structures were generated by predicted by using Alphafold3 server (https://alphafoldserver.com/).
Preparation of rPaSCL-Reg and rPaSCL-Ad
2.3
The codon-optimized sequences of rPaSCL-Reg and rPaSCL-Ad were synthesized de novo by Beijing Tsingke Biotech Co., Ltd and subsequently cloned into the plasmid (pET28a) before transformation into BL21 (DE3) cells to generate recombinant plasmids. Positive clones underwent PCR screening with primers T7-F and T7-R, followed by sequence confirmation. A single colony of transformed BL21 (DE3) cells was induced to express the recombinant protein with 0.1 mM IPTG at 28°C for 6 h. The resulting rPaSCL-Reg and rPaSCL-Ad were purified using a Ni column (Qiagen, Germany) and underwent desalination via Amicon^®^ Ultra (Merck, Germany), followed by analysis using 10% SDS-PAGE and Coomassie brilliant blue R-250 staining. Recombinant protein concentration was determined using BCA protein Assay Kit (Solarbio, China).
Western blot analysis
2.4
Following SDS-PAGE separation, the purified proteins were transferred to a nitrocellulose membrane (Pall, America). The membranes underwent blocking for 2 h with 5% non-fat milk in 1×TBST (10 mM Tris·HCl, 150 mM NaCl, 0.1% Tween, pH 7.2-7.6 in 25°C). After washing, the membrane was incubated with 1/5000 primary antibody (Rabbit anti-His-tag antibody, Abclonal, China) in TBST containing 5% non-fat milk at 4°C for 12 h. The washed membrane was then incubated with a secondary antibody (HRP-conjugated Goat Anti-Rabbit IgG, BBI, China) at 37°C for 2 h. The Western blot analysis was performed on UVP ChemStudio815 using an enhanced Chemiluminescence Substrate Kit (Yuanye, China).
Recombinant protein binding assay
2.5
Bacterial cultures were grown overnight and harvested by centrifugation at 8000 r for 10 min. The bacteria underwent three washing cycles with TBS (10 mM Tris·HCl, 150 mM NaCl, pH 7.2-7.6 in 25°C) and were resuspended in TBS to achieve an OD600 of 1.0. The bacterial suspension (100 μL) in TBS was incubated with purified rPaSCL-Reg and rPaSCL-Ad (50 μg) for 60 min at room temperature under mild rotation. Following incubation, the bacteria were washed four times with TBS. The final bacterial suspension underwent 10% SDS-PAGE and Western blot analysis. Rabbit anti-His-tag antibody (Abclonal, China, 1:5000) served as the primary antibody, while HRP-conjugated Goat Anti-Rabbit IgG (BBI, China, 1:10, 000) functioned as the secondary antibody, respectively.
An Elisa was employed to assess the sugar binding specificity of rPaSCL-Reg and rPaSCL-Ad. LPS from E. coli and PGN from S. aureus were selected for the assay. 96-well ELISA Plates were coated with 10 μg of polysaccharide and incubated at 4°C overnight. After five TBS washes, the microplates underwent blocking with BSA (1 mg/mL, 200 μL) at 37°C for 2 h, followed by TBS washing. Purified rPaSCL-Reg and rPaSCL-Ad (final concentration 0–80 μg/mL in TBS with 0.1 mg/mL BSA) were added to each coated well and incubated at 37°C for 2 h. The plate underwent five TBS washes. Subsequently, Rabbit anti-His-tag antibody (1:2000) was added (100 μL per well) and incubated at 37˚C for 2 h; 100 μL of HRP-conjugated Goat Anti-Rabbit IgG (1:5000) was introduced to each well at 37°C for 1 h. Following five TBS washes, color development occurred with TMB (BBI, China) at room temperature for 30 min. The OD value was measured at 450 nm. Each binding assay was performed in triplicate.
Agglutination test
2.6
Gram-positive bacteria (Staphylococcus aureus and Bacillus subtilis) and Gram-negative bacteria (Escherichia coli and Salmonella Typhimurium) were selected to evaluate the bacterial agglutination properties of rPaSCL-Reg and rPaSCL-Ad. A volume of 25 μL rPaSCL-Reg and rPaSCL-Ad (100 μg/mL) was incubated with an equal volume of bacterial suspension (1×10^8^ CFU/mL) at room temperature (~25°C) for 1 h, both with and without 10 mmol/L CaCl_2_. rBSA in TBS and TBS+Ca^2+^ (10 mM Tris·HCl, 150 mM NaCl, 10 mM CaCl_2_, pH 7.2-7.6 in 25°C) served as negative controls. Following incubation, agglutination was observed and documented using a fluorescence microscope (Nikon, Japan).
Antibacterial activity assays
2.7
A total of 100 μg of each protein was incubated with a bacterial suspension (1 × 10^5^ CFU/mL) in fresh LB (Tryptone 10 g/L, Yeast Extract 5g/L, NaCl 10g/L) in a 96-well culture plate. CaCl_2_ was added to a final concentration of 10 mM, with each well containing a total volume of 200 μL. The plate was incubated at 37 °C for 24 h. Bacterial growth was assessed by measuring the absorbance at 600 nm using a microplate reader. Each assay was performed in triplicate wells per protein and repeated in three independent experiments. rBSA served as a negative control. Data are presented as mean ± standard deviation (SD), and statistical significance was evaluated using student t-test.
Immunocytochemical analysis
2.8
Hemolymph extracted from P. americana was fixed using 200 μL of anticoagulant mixture (62 mM NaCl, 100 mM glucose, 10 mM EDTA, 30 mM Sodium citrate, 26 mM citric acid) and 4% paraformaldehyde, followed by centrifugation at 600 g for 10 min at 4°C. The isolated hemocytes were placed on poly-L-lysine-Prep slides (BBI, China) to facilitate cell adhesion during microscopic analysis, washed with TBS, and blocked with 5% BSA at 37°C for 30 min. Following TBS washing, the hemocytes were incubated with 10 μg rPaSCL-Reg or rPaSCL-Ad at 4°C for 12 h. The samples were then washed with TBS and incubated with 5% anti-His-tag antibody (1:1000 in 5% BSA) at room temperature for 2 h, followed by TBS washing and incubation with Alexa Fluor 488-conjugated Goat anti-rabbit IgG (Abclonal, China, 1:1000 in 5% BSA) for 1 h at 37°C in darkness. After six washes, the hemocytes were treated with 2-(4-Amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI, Beyotime, China) for 10 min at room temperature and washed six times. Fluorescence was examined using a laser scanning confocal microscope (LSCM) (Nikon, Japan). Nis-ElemeViewer software was utilized to assess the binding capability of rPaSCL-Reg and rPaSCL-Ad to hemolymph membranes.
Phagocytosis assay
2.9
To assess the phagocytic activity of hemocytes, Hemolymph was collected with anticoagulant and centrifuged at 600 g for 10 min at 4 °C to isolate hemocytes. Hemocytes were gently washed twice with TBS and resuspended. Escherichia coli was cultured overnight, harvested by centrifugation at 5000 rpm for 10 min, washed three times with TBS, and resuspended. Bacteria were labeled with 0.1 mg/mL FITC which dissolved in DMSO for 2 h in the dark, washed three times with TBS to remove excess dye, and finally resuspended in TBS at an OD600 = 1. For the phagocytosis assay, 100 µL of hemocyte suspension (1×10^6^ cells/mL) was added to each well of a confocal culture dish containing culture medium. The treatment group received 10 µg of rPaSCL-Reg and rPaSCL-Ad, whereas the control group received rBSA, with three replicates per group. Cells were incubated at 28 °C for 1 h. Subsequently, FITC-labeled bacteria were added at a 1:10 ratio of hemocytes to bacteria. Hemocytes were fixed with 4% paraformaldehyde for 30 min, washed three times with PBS, and 10 µL of the cell-bacteria mixture was placed onto a microscope slide for 10 min sedimentation in a humid chamber. Phagocytosis was observed under a LSCM (Nikon, Japan), and images were captured. For each treatment, at least 150 hemocytes from randomly selected fields were counted to calculate the phagocytic rate.
Scanning electron microscopy
2.10
Morphological changes in recombinant protein-bound bacteria were examined using field-emission scanning electron microscopy (Verios G4 UC, Thermo Fisher Scientific, America). Bacterial suspensions (1×10^8^ CFU/mL) were incubated with rPaSCL-Reg and rPaSCL-Ad at 2 mg/mL in TBS and TBS+Ca^2+^ (TBS, 10 mM CaCl_2_) at 37°C for 2 h, using rBSA protein (2 mg/mL) as the negative control. After incubation, cells underwent fixation with 2.5% (v/v) glutaraldehyde in 0.1 M Phosphate buffer at 4°C for 12 h, followed by gradual dehydration using increasing concentrations of ethanol (30%, 50%, 70%, 80%, 90%, and 100%) for 20 min at each step. The samples were subsequently dried, gold coated, and analyzed using field-emission scanning electron microscopy.
Real-time quantitative PCR
2.11
For gene expression analyses, 10-fold diluted cDNA served as templates for qRT-PCR. Reactions were performed in triplicate using ChamQ Universal SYBR qPCR Master Mix (Vazyme, Q711). The relative expression levels were calculated using the 2^-ΔΔCt^ method and there were three biological replicates and three technical replicates for each sample. Actin was chosen as a reference gene for qRT-PCR analysis (20). The primers used for qRT-PCR and the amplification efficiency (29) of each primer are shown in Supplementary Table S1.
Double-stranded RNA treatment
2.12
Double-stranded RNA (dsRNA) was synthesized using the T7 RiboMAX Express RNA interference (RNAi) System (Promega, P1700). Following purification, dsRNA was prepared at a concentration of 2 μg/μL, and 2μg was microinjected into the coxa of each nymph. To sustain RNAi efficiency throughout the regeneration process, injections were repeated every 7 days (15 nymphs per replicate, three biological replicates). The dsRNA-synthesizing primer sequences are detailed in Supplementary Table S1. A dsMock targeting a clone vector sequence was used as the negative control. For the analysis of differentially expressed genes following RNAi of target genes, dsRNA was performed together with amputation, and the coxa and trochanter at 7 days post-amputation (dpa) were harvested.
Results
3
Cloning and sequence analysis of PaSCLec
3.1
Through transcriptome analysis of leg regeneration in Periplaneta americana, a unique secreted C-type lectin (PaSCLec) with high expression was identified. Notably, an alternate donor site event in the transcript of PaSCLec generating a frameshift mutation revealed two distinct isoforms (designated PaSCL-Ad and PaSCL-Reg) (Supplementary Figure S1). Using 5′/3′ RACE and Nanopore sequencing, we found that transcripts corresponding to PaSCL-Ad constituted approximately 70–80% of the total isoform population (Primer 3′RACE: 79.06%; Primer 5′RACE: 75.46%). Therefore, the transcript of PaSCL-Ad was considered the main transcript. Sequence analysis showed the ORF of PaSCL-Ad (accession no. XP_069685827.1) encodes 214 amino acids with a molecular weight (MW) of 23.9 kDa and an isoelectric point of 4.97, while the ORF of PaSCL-Reg (accession no. XP_069685825.1) encodes 377 amino acids with a molecular weight (MW) of 41.3 kDa and an isoelectric point of 4.65. Both PaSCL-Ad and PaSCL-Reg proteins contain a 22-amino acid signal peptide and a 173-amino acid C-type lectin-like domain (Figure 1A). The amino acid sequence analysis indicates PaSCL-Reg contains 163 additional amino acids compared to PaSCL-Ad at its C-terminal (Figure 1B). A comparison of protein structure reveled that the overall architecture of PaSCL-Ad is relatively compact, with a well-defined core domain (Figure 1B1). In contrast, although PaSCL-Reg also possesses a recognizable core structure, it appears more loosely organized, suggesting that its overall structural stability may be lower (Figure 1B2). Phylogenetic analysis of PaSCL-Ad, PaSCL-Reg and other CTLs from invertebrate and vertebrate species revealed that, unlike Regenectin, PaSCL-Ad and PaSCL-Reg cluster with CTLs from Gryllus bimaculatus, which also possesses leg regeneration capability (Figure 1C).
Sequence analysis of PaSCL-Reg and PaSCL-Ad (A) Red square and pink hexagon represent signal peptide and CTLD domain. (B) Multiple alignments of the PaSCL-Reg with PaSCL-Ad. The structure of the PaSCL-Reg (B1) and PaSCL-Ad (B2) were analyzed using the online Alphafold3 server. (C) Phylogenetic analysis of PaSCL-Reg and PaSCL-Ad with other CTLs from various species. All amino acid sequences were collected from NCBI, and the tree was constructed with MEGA X software.
Expression profile of total PaSCLec
3.2
Tissue expression profiling revealed that total PaSCLec exhibited highest expression in the hemolymph, followed by fat body, coxa and gut (Figure 2A). To investigate the potential role of PaSCLec in innate immunity, sixth instar P. americana were subjected to bacterial injection to induce a systemic immune challenge. Compared with the control group, the mRNA levels of PaSCLec in hemolymph were significantly upregulated at 24 h post-injection with either E. coli (Figure 2B) or S. aureus (Figure 2C). These results suggest PaSCLec is involved in the immunoregulatory network of P. americana.
*Expression profile of total PaSCLec. (A) Relative mRNA levels of PaSCLec in different tissues. Relative mRNA levels of PaSCLec in hemolymph after E. coli(B) and S. aureus(C) treatment. The results are the mean and standard errors of three biological replicates. Different letters on the error bar indicate statistically significant differences at p < 0.05 level (ANOVA in association with Tukey’s HSD test). Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001). ANOVA, one-way analysis of variance; HSD, honestly significant difference; IB: injection buffer (control); mRNA, messenger ribonucleic acid.
Expression and purification of rPaSCL-Ad and rPaSCL-Reg
3.3
The alternative splicing variants rPaSCL-Ad and rPaSCL-Reg were expressed using a prokaryotic expression system to assess their biological activities and purified using the Ni-NTA Agarose (QIAGEN, Germany). The purification of rPaSCL-Ad and rPaSCL-Reg was verified by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (Figure 3A). To assess the immune functions of rPaSCL-Ad and rPaSCL-Reg, another P. americana C-type lectin, Regenectin, which is expressed during leg regeneration but has no reported immune activity, was also purified as a comparative control (Figure 3B).The purified rPaSCL-Ad, rPaSCL-Reg and Regenectin were analyzed using Western blot (Figure 3C, D, E), revealing apparent molecular weights of approximately ~26 kDa and ~40 kDa for rPaSCL-Ad and rPaSCL-Reg respectively, which correspond precisely to their predicted sizes.
The expression and purification of rPaSCL-Ad, rPaSCL-Reg and rRegenectin. rPaSCL-Reg, rPaSCL-Ad and rRegenectin were detected by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS–PAGE) (A, B) and Western blot (C-E). Lane M: molecular marker; Lane 1: crude supernatant protein extracts of bacteria with pET-28a; Lane 2, lane 4 and lane 6, crude supernatant protein extracts of bacteria with pET-28a-PaSCL-Ad, pET-28a-PaSCL-Reg and pET-28a-PaSCL-Regenectin proteins; Lane 3, lane 5 and lane 7, purified pET-28a-PaSCL-Ad, pET-28a-PaSCL-Reg and pET-28a-PaSCL-Regenectin proteins; Lane 8, lane9 and lane 10, western blot based on the purified pET-28a-PaSCL-Ad, pET-28a-PaSCL-Reg and pET-28a-PaSCL-Regenectin proteins.
Bacterial binding and agglutinating activity of rPaSCL-Ad and rPaSCL-Reg
3.4
C-type lectin exhibits pathogen recognition functions through binding to the Pathogen-Associated Molecular Patterns (PAMPs). Direct binding analysis using enzyme-linked immunosorbent assay (ELISA) demonstrated that rBSA had no binding ability (Figure 4A) and rPaSCL-Ad and rPaSCL-Reg bound to lipopolysaccharide (LPS) from E. coli and peptidoglycan (PGN) from S. aureus, with binding ability showing dose dependence (Figures 4B, C). Additionally, rRegenectin displayed lower binding capacity compared to rPaSCL-Ad and rPaSCL-Reg at equivalent concentrations (Figure 4D). Furthermore, rPaSCL-Ad demonstrated binding capability to Mannan and β-Glucan at low doses (PAMPs predominantly found in fungal cell walls). Western blotting analysis confirmed the binding affinity of rPaSCL-Ad and rPaSCL-Reg to bacteria. The results indicated that rPaSCL-Ad exhibited superior binding capacity for microorganisms including GP bacteria S. aureus, Bacillus subtilis, and GN bacteria Salmonella Typhimurium and E. coli (Figure 4E).
Binding affinity of rPaSCL-Ad, rPaSCL-Reg and rRegenectin. Binding of rBSA (A), rPaSCL-Ad (B), rPaSCL-Reg (C) and rRegenectin (D) to polysaccharide, analyzed by ELISA. ELISA, enzyme-linked immunosorbent assay; LPS, lipopolysaccharide; PGN, peptidoglycan. Binding of rPaSCL-Ad, rPaSCL-Reg and rRegenectin to bacteria (E. coli, S. aureus, B. subtilis and S. typhimurium) was detected by Western blot (E). Line 1: E. coli; Line 2: S. aureus; Line 3: S. typhimurium; Line 4: B. subtilis..
The agglutination activity of all recombinant proteins was evaluated using FITC method across various bacteria. All recombinant proteins demonstrated agglutination activity toward bacteria in the presence of Ca^2+^ (Figure 5), except for the control rBSA (Figure 5A). rPaSCL-Ad demonstrated the strongest agglutination activity (Figure 5B). Both rPaSCL-Reg and rRegenectin exhibited comparatively weaker agglutination, with rRegenectin showing almost no detectable activity toward E. coli (Figures 5C, D); however, no agglutinating activity was observed toward any bacteria without Ca^2+^ (Supplementary Figure S2), indicating Ca^2+^-dependent agglutinating activity.
Agglutination activities of rPaSCL-Ad, rPaSCL-Reg and rRegenectin with 10mM Ca2+ (recombinant proteins+bacteria). E. coli(A), S. aureus(B), S. typhimurium(C)B. subtilis(D). The corresponding control groups are shown in Supplementary Figure S2.
rPaSCL-Ad exhibits bacterial antibacterial activity and destroy the membrane integrity
3.5
Following the observation that rPaSCL-Reg and rPaSCL-Ad bind to bacterial surfaces, their potential bacteriostatic or bactericidal activity was examined. Bacterial cultures exposed to intact rPaSCL-Reg, rPaSCL-Ad and rRegenectin at concentrations of 100 μg/ml showed significant variations in bacterial growth compared to control cultures with rBSA control group. The results revealed that only rPaSCL-Ad demonstrated direct bacteriostatic activity against all bacteria (Figure 6A), while rPaSCL-Reg and rRegenectin specifically inhibited B. subtilis growth (Figure 6B and C). Notably, scanning electron microscopy (SEM) analysis revealed that both GN E. coli and GP S. aureus bacteria exhibited membrane pore formation following rPaSCL-Ad treatment (Figure 6B). In contrast, rPaSCL-Reg did not compromise cell membrane integrity of GN or GP bacteria (Supplementary Figure S3).
*Direct bacteriostatic and bacteriocidal activities in vitro of rPaSCL-Ad, rPaSCL-Reg and rRegenectin. (A) The rPaSCL-Ad, rPaSCL-Reg and rRegenectin were incubated with E. coli, S. aureus, S. typhimurium and B. subtilis for 6 h, 12 h and 18 h Bacterial growth was assessed by measuring absorbance at 600 nm. rBSA protein was used as a negative control; (B) Scanning electron microscopy (SEM) was used to examine the observation on the interaction between rPaSCL-Ad and bacteria. Representative SEM images of E. coli (a), S. aureus (b), S. typhimurium (c), B. subtilis (d) are shown. Arrows indicate membrane pore formation. Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001).
rPaSCL-Ad and rPaSCL-Reg binds to bacterial surface components and to the Periplaneta americana hemocyte surface
3.6
To determine whether rPaSCL-Ad, rPaSCL-Reg and rRegenectin are involved in hemocyte-mediated phagocytosis, their ability to bind to the hemocyte surface of Periplaneta americana was examined by immunocytochemical analysis. The results showed that all three recombinant proteins bound to the hemocyte surface (Figure 7A).
*rPaSCL-Ad and rPaSCL-Reg bind to the hemocyte surface and enhance the phagocytic rate. (A) Immunocytochemical analysis was performed to examine the binding of rPaSCL-Ad and rPaSCL-Reg to the hemocyte surface. (B) Immunocytochemical assays were conducted to assess rPaSCL-Ad and rPaSCL-Reg enhanced phagocytosis in vitro. B’ The phagocytic rate was calculated after 2h, and statistical significance was analyzed using one-way ANOVA: *p < 0.05, **p < 0.01, **p < 0.001. Arrows indicate sites of hemocyte phagocytic activity.
Subsequently, collected hemocytes were placed in cell culture plates and pre-incubated separately with each recombinant proteins, using rBSA as the control group. FITC-labeled E. coli was then added for co-incubation. After fixation, hemocytes phagocytosis of E. coli was visualized by microscopy and quantitatively analyzed. The results demonstrated that rPaSCL-Ad and rPaSCL-Reg significantly enhanced hemocyte phagocytosis of E. coli compared with the control, whereas rRegenectin had no detectable effect (Figure 7B and B’). These findings suggest that rPaSCL-Ad and rPaSCL-Reg promote hemocyte phagocytic activity by facilitating interactions between bacterial and hemocytes.
PaSCLec regulates the expression of antimicrobial peptides
3.7
To investigate whether PaSCLec indirectly participates in immune responses through AMP expression regulation, dsPaSCLec was synthesized in vitro and injected into 6th P. americana. Following stimulation with E. coli and S. aureus, decreased relative mRNA levels of PaSCLec were observed (Figures 8A, D). Six AMPs were evaluated: Attacin-1 (Att-1), Termicin-1 (Term-1), Termicin-2 (Term-2), Defensin-1 (Def-1), Defensin-2 (Def-2) and Defensin-3 (Def-3). Bacterial challenges significantly downregulated the expression of antimicrobial peptides Att-1, Term-2, Def-1 and Def-3 (Figures 8B, E). Conversely, Term-1 and Def-2 expression were equally upregulated in the dsPaSCLec transfected cockroaches. To examine the potential role of PaSCLec in AMP regulation in vivo, its involvement in Toll, IMD, and JAK/STAT pathways was investigated via transcription factors Dorsal, Relish, and STAT. The results indicated significant reduction in Dorsal, Relish and STAT expression (Figures 8C, F).
*Expression of transcription factors and antimicrobial peptides (AMPs) following bacterial challenge after PaSCLec knockdown. Relative mRNA levels of PaSCLec following injection with E. coli(A) or S. aureus(D). The mRNA expression levels of AMPs (B, E) and transcription factors (C, F) were measured after injection of E. coli and S. aureus at 24 h, respectively. Each group consisted of 15 larvae injected with dsRNA, and three biological replicates were performed. Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001).
To validate these findings, dsPaSCLec transfected cockroaches were initially injected with rPaSCL-Ad and rPaSCL-Reg protein respectively, followed by E. coli challenge as previously described. The results demonstrated significantly increased Relish and STAT expression in response to E. coli challenge in both rPaSCL-Ad-rescued and rPaSCL-Reg-rescued cockroaches (Figure 9A). Additionally, the expression of Att-1 and Def-3, along with transcription factors Relish and STAT, showed significant increases compared to control cockroaches (Figure 9B). These findings indicate that PaSCLec specifically upregulates the expression of Att-1 and Def-3, suggesting that PaSCLec regulates AMP expression through multiple pathways.
*Expression of transcription factors and AMPs in rPaSCL-Ad-rescued and rPaSCL-Reg rescued cockroaches following E. coli challenge. The mRNA levels of AMPs (A) and transcription factors (B) were analyzed by qRT-PCR in rPaSCL-Ad- and rPaSCL-Reg-injected cockroaches. rBSA was used as the control. Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001).
PaSCLec is involved in reg regeneration of P. americana
3.8
To investigate whether PaSCLec has an effect on leg regeneration, we first evaluated the expression of PaSCLec at 0, 3, 7 and 14 days post-ecdysis (dpe) or dpa. The results showed that the expression of PaSCLec was significantly upregulated during leg regeneration (Figures 10A, B). Then, RNAi of PaSCLec was performed and the interference efficiency of PaSCLec was measured on 7 dpa and 14 dpa, and the mRNA level of PaSCLec was significantly reduced compared with control (Figure 10C). Injection of dsPaSCLec caused the regenerated legs to exhibit morphological abnormalities, while no change was observed when dsMock was injected (Figures 10D, E). The regenerative length (Regenerated leg/Contralateral leg, %) was measured after RNAi treatments of genes (Figure 10F). In the wild-type (WT) group (N = 15), the mean relative regenerative length was 84.046% ± 1.98% (mean ± standard deviation, SD). The dsMock group, which served as a non-targeting control (N = 15), exhibited a similar relative regenerative length of 81.214% ± 2.06%. In contrast, the dsPaSCLec group, in which PaSCLec was knocked down (N = 15), showed a significantly reduced relative regenerative length of 62.473% ± 4.98%. Statistical analysis revealed that both WT and dsMock groups had markedly higher relative regenerative lengths compared to the dsPaSCLec group.
*The C-type lectin PaSCLec regulates leg morphogenesis. (A) Relative mRNA level of PaSCLec and Regenectin during normal development. (B) Relative mRNA level of PaSCLec during leg regeneration. (C) Silencing efficiency of dsPaSCLec RNAi at 7 and 14 dpa. (D) Phenotype of regenerated legs in the dsMock-injected group. (E) Phenotype of regenerated legs in the dsPaSCLec-injected group. (F) Relative regenerative length of regenerated legs in Wide Type (WT), dsMock- and dsPaSCLec-treated groups. Each group including 15 larvae was injected into dsRNA. Different letters on the error bar indicate statistically significant differences at p < 0.05 level (ANOVA in association with Tukey’s HSD test). Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001).
PaSCLec regulates the expression of AMPs during leg regeneration
3.9
To investigate PaSCLec’s immune function during leg regeneration, we measured mRNA levels of Def-3 and Att-1 regulated by PaSCLec in leg regeneration. Results indicated sustained expression of Def-3 and Att-1 during early regeneration phases (Figures 11A, B), which were downregulated after injection of dsPaSCLec (Figure 11C).
*Effects of PaSCLec on Def-3 and Att-1 during leg regeneration. (A) Relative mRNA level of Def-3 during leg regeneration. (B) Relative mRNA level of Att-1 during leg regeneration. (C) Relative mRNA level of Def-3 and Att-1 determined by qRT-PCR following dsPaSCLec treatment. Each group consisted of 15 larvae injected with dsRNA. Different letters above the error bars indicate statistically significant differences at p < 0.05 level (ANOVA in association with Tukey’s HSD test). Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001).
PaSCLec and JAK/STAT signaling may synergistically regulate immunity and leg regeneration in Periplaneta americana
3.10
To explore the latent interaction of JAK/STAT signaling pathways with PaSCLec during leg regeneration, RNAi was performed to knockdown the expression of PaSCLec in P. americana. The expression of unpaired was upregulated at 3 dpa and 7 dpa and reduced significant at 7 dpa when the PaSCLec was silenced (Figures 12A, B). Our results showed that PaSCLec knockdown leads to reduced unpaired mRNA levels during leg regeneration, suggesting that PaSCLec may modulate the transcription of JAK/STAT associated cytokines. To confirm the role of the JAK/STAT pathway and specifically the functions of the ligand unpaired, we performed to knockdown the expression of unpaired. Silencing efficiency of RNAi was detected by on 7 dpa (Figure 12C). Knockdown of unpaired in the JAK/STAT signaling disrupted the entire regeneration process (Figure 12D) and the mRNA level of Def-3 was significantly impaired (Figure 12E). In contrast, the mRNA level of Att-1 was upregulated (Figure 12E). Because unpaired is a canonical activator of the JAK/STAT pathway, this correlation implies a possible functional relationship between PaSCLec and JAK/STAT-mediated processes in immunity and regeneration. Unpaired is a canonical activator of the JAK/STAT pathway, this correlation implies a possible functional relationship between PaSCLec and JAK/STAT mediated processes in immunity and regeneration. Nevertheless, the involvement of JAK/STAT signaling remains speculative, as the current study does not provide direct mechanistic evidence. We did not detect physical interactions, receptor activation, or downstream readouts such as STAT phosphorylation or nuclear translocation. Thus, additional experimental work will be required to define whether and how PaSCLec interacts with the JAK/STAT pathway in future.
*PaSCLec and unpaired regulate the Def-3 expression and leg regeneration through a feedback loop. (A) Relative mRNA level of unpaired during leg regeneration. (B) Relative mRNA level of unpaired following dsPaSCLec treatment. (C) RNAi silencing efficiency of dsupd at 7 dpa. (D) RNAi treatment significantly disrupted the entire leg regeneration process. (E) Relative mRNA level of PaSCLec, Def-3 and Att-1 following dsupd treatment. Each group consisted of 15 larvae injected with dsRNA. Different letters above the error bars indicate statistically significant differences at p < 0.05 level (ANOVA in association with Tukey’s HSD test). Asterisk indicates significant differences compared with values of the control (student’s t test, *p < 0.05, **p < 0.01, **p < 0.001).
Discussion
4
Secretory CTLs in the insect immune system are soluble PRRs that mediate innate immune defense by specifically recognizing evolutionarily conserved glycan motifs on pathogen surfaces (30, 31). In this study, we identified a novel secretory C-type lectin gene, PaSCLec, which has undergone evolutionary divergence through an alternative donor-site splicing event. This splicing event introduces a frameshift mutation that produces two distinct isoforms, PaSCL-Reg and PaSCL-Ad. Both PaSCL-Ad and PaSCL-Reg retain identical signal peptides and CLECT domains in their tertiary structures (Figure 1). However, the two isoforms exhibit functional divergence, including distinct ligand-binding specificities and differential regulatory effects on immune cell signaling pathways. A similar phenomenon has been reported in Procambarus clarkii, where the spliced isoform PcGal4-L-CRD exhibits stronger binding activity than PcGal4-L (32). These findings suggest that the strong upregulation of PaSCLec during P. americana leg regeneration contributes to tissue-specific immunoregulation and may be involved in pathogen recognition.
Tissue expression profiling revealed that PaSCLec is highly expressed in the hemolymph (Figure 2A), consistent with observations in other insect CTLs (33, 34). Previous studies indicate that secretory C-type lectins in the hemolymph modulate innate immunity through pathogen recognition (35) and antimicrobial peptide (AMP) induction (36), supporting the hypothesis that PaSCLec contributes to immune regulation. To test this hypothesis, Gram-negative E. coli and Gram-positive S. aureus were injected into P. americana. Following bacterial injection, PaSCLec expression in the hemolymph was first downregulated and subsequently upregulated (Figures 2B, C). A similar expression pattern has been reported in Tribolium castaneum and Octodonta nipae (7, 37). This response suggests that bacteria may transiently suppress CTL expression in the hemolymph as a strategy to evade host immune defenses (38). The decrease in PaSCLec mRNA levels after bacterial infection supports this possibility, although the underlying mechanism remains to be elucidated. Recognition and binding of bacterial carbohydrate structures, including LPS and PGN (39), likely underlie its agglutination activity (40). Accumulating evidence shows that CTLs display dose-dependent binding affinity toward both LPS and PGN (37, 41–43). The mutant rPaSCL-Ad exhibited significantly higher binding affinity for PAMPs than rPaSCL-Reg and rRegenectin (Figures 4A–D). These results indicate that although both PaSCLec and Regenectin are upregulated during limb regeneration, only the mutant rPaSCL-Ad shows strong in vitro binding to PAMPs, supporting its involvement in immune defense. Bacterial binding assays further confirmed the superior binding affinity of rPaSCL-Ad (Figure 4E), reinforcing this conclusion. Subsequent assays demonstrated that rPaSCL-Ad, rPaSCL-Reg, and rRegenectin agglutinate both GP and GN bacteria in a Ca²^+^-dependent manner, with rPaSCL-Ad displaying the strongest activity (Figure 5). This outcome is consistent with previous reports (44, 45).
Because both rPaSCL-Ad and rPaSCL-Reg bind and agglutinate bacteria, we next examined their bacteriostatic and bactericidal activities. Previous studies reported that rMaCTL inhibits the growth of multiple bacterial species in vitro (46). A similar inhibitory effect was observed for rPaSCL-Ad (Figure 6A). Similar effects were observed in the mutant rPaSCL-Ad (Figure 6A). SEM analysis showed that rPaSCL-Ad disrupted the cellular structures of E. coli and S. aureus (Figure 6D), potentially due to its strong binding to LPS and PGN. The truncated rPaSCL-Ad displayed marked antibacterial activity, whereas rPaSCL-Reg lacked detectable activity. Structural analysis suggests that the functional divergence arises from an exposed membrane-active segment in PaSCL-Ad that mediates membrane perforation. In contrast, the extended C-terminal region of PaSCL-Reg likely shields or neutralizes this segment. However, S. typhimurium and B. subtilis remained intact cellular structures. Both proteins may induce partial bacterial inactivation through agglutination-mediated mechanisms. Similar bactericidal activity exists in other CTLs. For example, the crab lectin EsIgLectin kills V. parahaemolyticus and S. aureus (47). The newly identified lectin Perlucin exhibits bactericidal activity against V. parahaemolyticus (48). The Pacific Abalone CTL shows broad-spectrum activity against both GP and GN bacteria (49).
The recombinant CTL enhances hemocyte phagocytic capacity by binding to hemocyte surface receptors or PAMPs. Previous research demonstrated that rTcCTL2 binds to microbes and promotes hemocyte-mediated phagocytosis in vitro (50). MjCC-CL binds to E. coli via recognition of cell-surface polysaccharides and also interacts with self-ligands on shrimp hemocyte surfaces (51). Similarly, rPaSCL-Ad and rPaSCL-Reg demonstrated binding to the hemocyte surface (Figure 7A), and in vitro experiments showed rPaSCL-Ad more effectively enhanced hemocyte phagocytic activity (Figure 7B and B’), suggesting their potential role as opsonins.
As PRRs, C-type lectins primarily recognize invading pathogens and trigger downstream immune responses. For example, CTL16 activates immune signaling pathways that promote AMP production (52). Knockdown of TcCTL15 has been shown to reduce the mRNA levels of Dif, Rel, and STAT (6). A novel chimeric CTL identified in Marsupenaeus japonicus induces the expression of specific AMPs through the JAK/STAT pathway (51). Following dsPaSCLec transfection in P. americana, subsequent challenges with E. coli and S. aureus resulted in the downregulation of four AMPs (Att-1, Term-2, Def-1 and Def-3) and transcription factors Dorsal, Relish, and STAT. These results indicate that PaSCLec plays a regulatory role in the immune response (Figure 8).
To validate these findings, dsRNA-transfected P. americana were first injected with rPaSCL-Ad or rPaSCL-Reg proteins and then challenged with E. coli as described above. Although both rPaSCL-Ad and rPaSCL-Reg regulate AMP expression, their functions likely differ because rPaSCL-Ad contains a truncated structure. Specifically, rPaSCL-Ad appears to function primarily as an opsonin, whereas rPaSCL-Reg plays a major role in immune responses to microbial challenge (Figure 9). Recent studies have shown that dimerization of CLEC2D alters its ligand-binding properties and thereby modulates immune responses (53).A single carbohydrate-binding site per CML1 protomer was located at the dimer interface, where both protomers contribute to ligand engagement. This interaction results in a hexavalent lectin (54). These findings suggest that PaSCLec exerts antimicrobial activity by activating classical immune pathways, thereby selectively inducing AMP expression in response to E. coli and S. aureus challenge.
Cockroaches demonstrate superior regeneration capabilities within the class Insecta (23). qRT-PCR results revealed that PaSCLec was highly expressed during leg regeneration (Figures 10A, B), implying that PaSCLec possibly plays an important role in regulating leg regeneration of P. americana. Previous studies have shown that REG3A, a secreted C-type lectin of the regenerating (REG) family, promotes the growth of PDAC cell lines by binding directly to the extracellular domain of EGFR (55). In addition, CLEC3A activates the PI3K–AKT signaling pathway to enhance cell proliferation (14). RNAi experiments demonstrated that knockdown of PaSCLec caused distinct morphological abnormalities in regenerated legs (Figures 10B–F). These findings further indicate that PaSCLec plays an essential role in leg regeneration in P. americana.
Previous studies have shown that AMP expression is markedly upregulated at wound edges after skin injury, indicating that AMPs may act as wound-healing stimulators (56, 57). In addition, AMPs modulate host immune responses, thereby enhancing pathogen clearance and promoting tissue repair (58). In our study, the mRNA levels of Att-1 and Def-3 were upregulated during leg regeneration but were significant reduced in the dsPaSCLec group (Figure 11). These findings indicate that PaSCLec regulates the expression of Att-1 and Def-3 during bacterial immune responses and leg regeneration. However, the specific functions of these genes require further validation through in vitro and in vivo experiments as well as bioinformatic analyses.
We preliminarily investigated the mechanism by which PaSCLec influences leg regeneration. Previous studies showed that JAK/STAT signaling contributes to leg regeneration in Gryllus bimaculatus (59). Our results further demonstrate that PaSCLec may modulates immune responses through the JAK/STAT signaling pathway. The JAK/STAT pathway is an evolutionarily conserved signaling cascade that plays critical roles in development, physiological homeostasis, and regenerative responses following infection or tissue injury (60). Therefore, we examined the expression of unpaired, a ligand of the JAK/STAT pathway after knockdown of PaSCLec. The mRNA levels of unpaired were significantly reduced at 3 dpa and 7 dpa (Figures 12A, B), suggesting that PaSCLec may mediate the expression of unpaired.
Knockdown of unpaired significantly impaired the regeneration process (Figures 12C, D), consistent with previous research (20). Additionally, unpaired knockdown led to a significant decrease in the expression of Def-3(Figure 12E), indicating that Def-3 is regulated by the JAK/STAT pathway. BmCTL5 has been proposed as an important PRR regulating the JAK/STAT pathway (61). In contrast, Att-1 is likely regulated by other pathways, such as the Toll or Imd pathway (62), as its expression increased significantly after unpaired knockdown. Although knockdown of PaSCLec and unpaired caused morphological defects in regenerated legs, this study primarily relied on external phenotypic measurements to assess regenerative outcomes. In addition, JAK/STAT signaling pathway activity and the protein-level of PaSCLec were not determined. Consequently, our findings provide only a preliminary insight into the role of PaSCLec in regulating immunity and regeneration. Future studies incorporating histological examinations and functional assays will be necessary to comprehensively elucidate the mechanisms by which PaSCLec regulates regenerative processes.
In conclusion, we identified a novel secreted C-type lectin, PaSCLec, from P. americana. PaSCLec exhibited two major functions in P. americana. First, its two isoforms (PaSCL-Reg and PaSCL-Ad) participate in immune responses by mediating bacterial agglutination, exerting direct bacteriostatic activity, and regulating AMP expression to resist pathogenic invasion. Second, PaSCLec is highly expressed during leg regeneration. Knockdown of PaSCLec impairs this process and significantly reduces the mRNA levels of unpaired, suggesting that PaSCLec may regulate leg regeneration by modulating the JAK/STAT signaling pathway. Thus, additional experimental work will be required to define whether and how PaSCLec interacts with the JAK/STAT pathway in future. These finding expand our understanding of secreted C-type lectins and provide a theoretical foundation for elucidating their roles in regenerative processes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dodd RB Drickamer K . Lectin-like proteins in model organisms: implications for evolution of carbohydrate-binding activity. Glycobiology. (2001) 11:71R–9R. doi: 10.1093/glycob/11.5.71R, PMID: 11425795 · doi ↗ · pubmed ↗
- 2Kim T Ri S Ju K Shi W Zhou W Yu Y . A C-type lectin with a single carbohydrate-recognition domain (CRD) containing unique QPN/WDD motifs from Tegillarca granosa is involved in the innate immune defense. Fish Shellfish Immunol. (2023) 142:109093. doi: 10.1016/j.fsi.2023.109093, PMID: 37722437 · doi ↗ · pubmed ↗
- 3Cambi A Koopman M Figdor CG . How C-type lectins detect pathogens: C-type lectins and pathogens. Cell Microbiol. (2005) 7:481–8. doi: 10.1111/j.1462-5822.2005.00506.x, PMID: 15760448 · doi ↗ · pubmed ↗
- 4Li M Li C Ma C Li H Zuo H Weng S . Identification of a C-type lectin with antiviral and antibacterial activity from pacific white shrimp Litopenaeus vannamei. Dev Comp Immunol. (2014) 46:231–40. doi: 10.1016/j.dci.2014.04.014, PMID: 24792214 · doi ↗ · pubmed ↗
- 5Huang X Huang Y Shi Y-R Ren Q Wang W . Function of a novel C-type lectin with two CRD domains from Macrobrachium rosenbergii in innate immunity. Dev Comp Immunol. (2015) 49:121–6. doi: 10.1016/j.dci.2014.11.015, PMID: 25475962 · doi ↗ · pubmed ↗
- 6Wang S Ai H Zhang Y Bi J Gao H Chen P . Functional analysis of a multiple-domain CTL 15 in the innate immunity, eclosion, and reproduction of tribolium castaneum. Cells. (2023) 12:608. doi: 10.3390/cells 12040608, PMID: 36831275 PMC 9954269 · doi ↗ · pubmed ↗
- 7Zhang Y Ai H Wang Y Zhang P Du L Wang J . A pattern recognition receptor C-type lectin Tc CTL 14 contributes to immune response and development in the red flour beetle, Tribolium castaneum. Insect Sci. (2023) 30:1363–77. doi: 10.1111/1744-7917.13161, PMID: 36518010 · doi ↗ · pubmed ↗
- 8Chen P Ai H Liu Z Li C Li B . The dual functions of a newly identified C-type lectin (Tc CTL 17) in the immunity and development of Tribolium castaneum. Bull Entomol Res. (2025) 115:251–64. doi: 10.1017/S 0007485324000920, PMID: 40099412 · doi ↗ · pubmed ↗