Gestational and lactational dietary supplementation with live yeast partially attenuates inflammatory responses to lipopolysaccharide challenge in newly weaned piglets
Yuechi Fu, Abiola S Lawal, Timothy A Johnson, Theresa M Casey, Jun Xie, Olayiwola Adeola, Kolapo M Ajuwon

TL;DR
Adding live yeast to the diets of pregnant and nursing sows may help reduce inflammation in piglets after weaning when they are exposed to bacterial toxins.
Contribution
This study shows maternal live yeast supplementation can partially reduce inflammation in piglets after weaning.
Findings
YLPS piglets had lower rectal temperatures and reduced TNF-α levels in the ileal mucosa compared to CLPS piglets.
Piglets from LY-supplemented sows showed higher mRNA levels of several cytokines and E-cadherin protein in the ileal and jejunal mucosa.
YLPS piglets had reduced NF-κB and TLR4 mRNA in the liver and lower NF-κB and MyD88 gene expression in the mesenteric lymph node.
Abstract
Weaning is an abrupt event in the life of piglets that adversely affects metabolic homeostasis, leading to poor nutrient absorption, increased susceptibility to enteric pathogens, and reduced growth performance. Few studies have examined the effects of maternal dietary live yeast (LY) supplementation on the responses of piglets subjected to an immunological challenge immediately after weaning. The aim of this study was to evaluate the effects of gestational and lactational dietary LY supplementation on inflammatory and antioxidant markers in newly weaned piglets challenged with lipopolysaccharide (LPS). On day 77 of gestation, 40 sows were randomly assigned to two dietary treatments: without (CON) or with LY supplementation at 0.05% of the diet during gestation and 0.1% during lactation. Within 24 h postweaning, 16 piglets with similar weights were selected from each maternal group and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
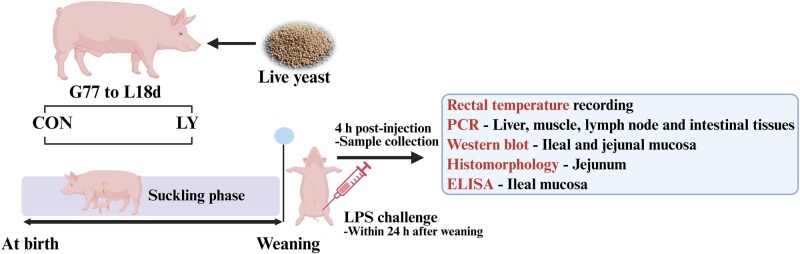 Figure 1
Figure 1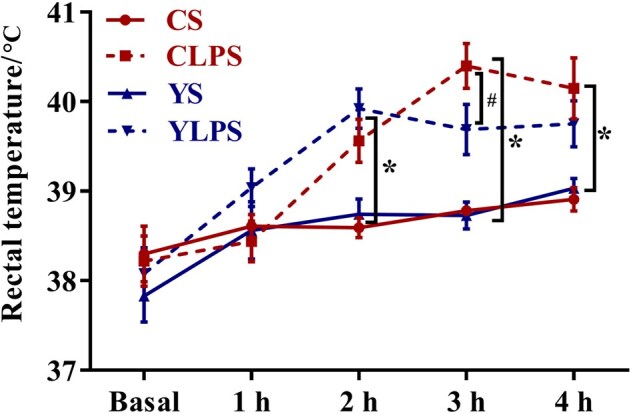 Figure 2
Figure 2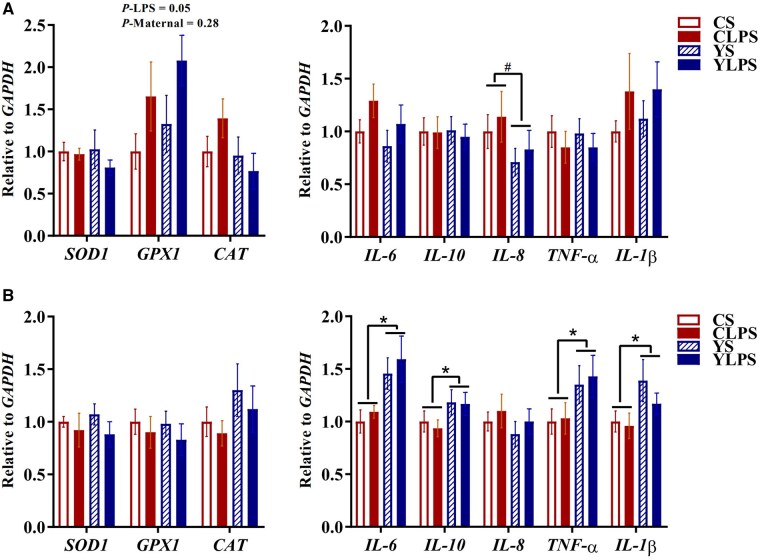 Figure 3
Figure 3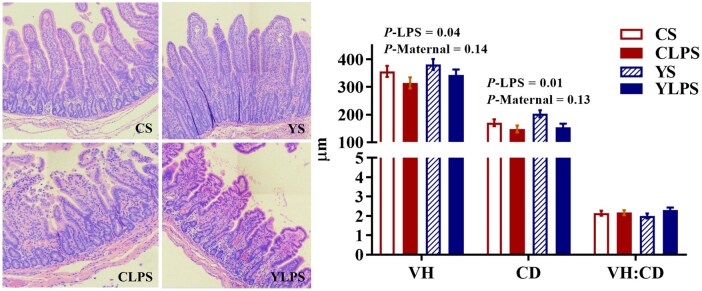 Figure 4
Figure 4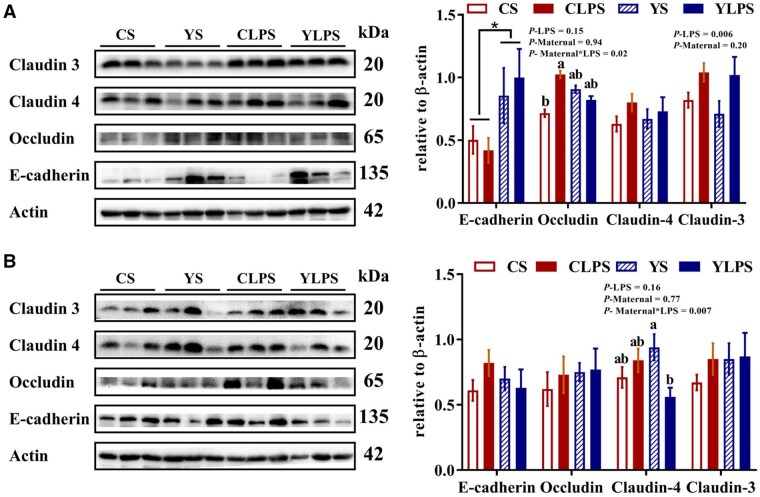 Figure 5
Figure 5| Ingredient, % | Gestation | Lactation |
|---|---|---|
|
| 76.95 | 57.62 |
|
| 17.50 | 35.00 |
|
| 1.20 | 3.00 |
|
| 1.69 | 1.63 |
|
| 1.30 | 1.50 |
|
| 0.15 | 0.15 |
|
| 0.10 | 0.10 |
|
| 0.01 | 0.01 |
|
| 0.10 | 0.10 |
|
| 0.50 | 0.50 |
|
| 0.21 | 0.10 |
|
| 0.25 | 0.25 |
|
| 0.04 | 0.04 |
|
| 100.00 | 100.00 |
|
| ||
|
| 3286.3 | 3352.8 |
|
| 146.9 | 214.5 |
|
| 9.0 | 9.6 |
|
| 5.8 | 7.2 |
|
| 3.7 | 4.5 |
|
| 6.0 | 10.3 |
| Item (pg/mg) | CON | LY |
|
| ||||
|---|---|---|---|---|---|---|---|---|
| Saline | LPS | Saline | LPS | Maternal diet | Challenge | Maternal diet × Challenge | ||
|
| 13.99 | 16.26 | 19.31 | 12.96 | 2.03 | 0.62 | 0.33 | 0.05 |
|
| 108.14 | 120.39 | 106.40 | 90.43 | 7.07 | 0.03 | 0.78 | 0.05 |
| Maternal diet | CON | LY |
|
| ||||
|---|---|---|---|---|---|---|---|---|
| Saline | LPS | Saline | LPS | Maternal diet | Challenge | Maternal diet × Challenge | ||
|
| ||||||||
|
| 1.00 | 0.92 | 1.37 | 1.02 | 0.30 | 0.41 | 0.45 | 0.63 |
|
| 1.00 | 1.45 | 1.08 | 1.49 | 0.31 | 0.84 | 0.16 | 0.94 |
|
| 1.00 | 0.61 | 1.30 | 0.67 | 0.12 | 0.17 | 0.0007 | 0.34 |
|
| 1.00 | 2.85 | 0.87 | 2.72 | 0.24 | 0.55 | <0.0001 | 0.98 |
|
| 1.00 | 1.70 | 0.91 | 1.55 | 0.16 | 0.45 | 0.0005 | 0.84 |
|
| 1.00 | 1.57 | 1.19 | 1.72 | 0.17 | 0.31 | 0.003 | 0.92 |
|
| 1.00 | 2.91 | 1.09 | 1.57 | 0.33 | 0.07 | 0.002 | 0.04 |
|
| 1.00 | 2.14 | 1.06 | 3.32 | 0.47 | 0.84 | <0.0001 | 0.63 |
|
| 1.00 | 3.83 | 1.03 | 3.31 | 0.41 | 0.54 | <0.0001 | 0.49 |
|
| 1.00 | 5.39 | 1.02 | 3.27 | 0.42 | 0.02 | <0.0001 | 0.02 |
|
| ||||||||
|
| 1.00 | 0.53 | 0.78 | 0.52 | 0.12 | 0.31 | 0.004 | 0.36 |
|
| 1.00 | 0.95 | 0.64 | 0.78 | 0.16 | 0.13 | 0.78 | 0.56 |
|
| 1.00 | 0.74 | 0.65 | 0.52 | 0.13 | 0.04 | 0.15 | 0.59 |
|
| 1.00 | 1.22 | 1.07 | 1.39 | 0.20 | 0.53 | 0.16 | 0.81 |
|
| 1.00 | 2.01 | 1.03 | 1.15 | 0.17 | 0.03 | 0.004 | 0.02 |
|
| 1.00 | 1.64 | 1.30 | 1.16 | 0.23 | 0.70 | 0.27 | 0.10 |
|
| 1.00 | 1.83 | 1.13 | 1.41 | 0.23 | 0.49 | 0.01 | 0.20 |
|
| 1.00 | 0.76 | 0.54 | 0.19 | 0.19 | 0.19 | 0.85 | 0.30 |
|
| 1.00 | 1.79 | 0.78 | 0.92 | 0.21 | 0.02 | 0.04 | 0.13 |
|
| 1.00 | 0.96 | 0.64 | 0.65 | 0.15 | 0.02 | 0.93 | 0.84 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Behavior and Welfare Studies · Animal Nutrition and Physiology · Gut microbiota and health
Introduction
Weaning is a routine procedure in pig husbandry, although it is associated with nutritional, immunological, and physiological disturbances in piglets (Pluske et al., 2018). These perturbations result from multiple events that occur during weaning, including transportation, handling, sudden dietary transition, and environmental change. Consequently, weaning stress-induced diarrhea is associated with gut dysbiosis, growth retardation, and increased mortality (Tang et al., 2022). As this period is associated with poor pig performance, several management, dietary, and feeding strategies have been employed to mitigate the adverse effects of weaning stress. Some of these strategies target the gestating and lactating sows, with the expectation that the benefits are transferable to piglets after weaning. A few of these approaches include provision of adequate housing during gestation, adjusting sow feeding or supplementing sow diets with probiotics and prebiotics during pregnancy and lactation (Blavi et al., 2021).
Yeast and yeast-based byproducts, including yeast culture, live yeast (LY), and yeast cell wall components such as β-glucans and mannan oligosaccharides, have been utilized as dietary supplements to modulate metabolic processes and enhance immune function in food animal production across various growth stages (Burdick Sanchez et al., 2021; Patterson et al., 2022). Commercially available LY products for animal feeding are typically manufactured in coated or encapsulated granular forms to protect yeast stability and viability during diverse storage and environmental conditions (Arslan et al., 2015). Live yeast exerts probiotic effects through viable yeast cells and prebiotic functions through polysaccharide-rich cell wall fractions and fermentation-derived metabolites, which support gastrointestinal health and host physiological function through multiple mechanisms (Fan et al., 2025). These include modulation of the immune system by promoting immune cell proliferation or activating macrophages and neutrophils to release inflammatory cytokines (Kogan and Kocher, 2007; Monroy-Salazar et al., 2012), enhancement of the host antioxidant defense system by upregulating the synthesis of antioxidant enzymes such as superoxide dismutase and catalase (Long et al., 2021), or supporting intestinal barrier function by stimulating mucin secretion and increasing expression of key junctional proteins, including proteins located in tight junctions and adherens junctions (Fan et al., 2024). Moreover, when fermented, LY cell wall components such as β-glucan and mannan oligosaccharides produce short-chain fatty acids and contribute to gastrointestinal acidification, thereby inhibiting pathogenic bacteria, reshaping gut microbiota composition, and increasing microbial diversity (Kiros et al., 2019). Consequently, research evidence indicates that supplementing LY, either directly in nursery pig diets or indirectly through the sow, enhances piglet growth performance, nutrient digestibility (Lu et al., 2019), and intestinal development (Fu et al., 2024), while also reducing the instances of antimicrobial resistance to antibiotics (Chance et al., 2022).
Exposure to bacterial pathogens represents a major challenge for weanling pigs. This period represents a time during which piglets are vulnerable to intestinal and systemic inflammation in response to bacterial infection from organisms such as Escherichia coli. Lipopolysaccharide (LPS) obtained from E. coli is often used to model this inflammatory response (Yao et al., 2017; Guevara et al., 2024). Although previous studies have reported the effects of maternal or postweaning LY supplementation on intestinal health and immune function in pigs several weeks after weaning (Trckova et al., 2014; Law et al., 2024), few have investigated the effects of maternal LY supplementation on tissue-specific inflammatory responses in piglets facing an inflammatory stimulus immediately after weaning. Therefore, the objective of the present study was to determine the impacts of supplementing LY to sow diets on inflammatory and antioxidant markers in selected tissues (intestinal mucosa, liver, muscle, and mesenteric lymph node) of piglets challenged with LPS immediately after weaning.
Materials and Methods
All protocols used in this study were approved by the Purdue University Institutional Animal Care and Use Committee (No. 1111000145).
Animals and experimental design
The management and dietary treatment of sows during late gestation and lactation were described in our previous study (Fu et al., 2024). Briefly, on day 77 of gestation, a total of forty sows (Landrace × either Duroc or Yorkshire) were randomly assigned to two dietary groups based on their expected farrowing date and parity (*n *= 20): basal diet only (CON) or the basal diet supplemented with live yeast (LY, Saccharomyces cerevisiae, Vistacell, AB Vista, Marlborough, Wiltshire, UK). The LY product was mixed directly into the feed at inclusion rates of 0.05% during gestation and 0.10% during lactation. According to manufacturer specifications and previous studies using this product (Lu et al., 2019), LY contains approximately 2 × 10^10^ CFU/g viable yeast cells; however, viable counts were not quantified in the current study. The basal diet (Table 1) was formulated to meet or exceed the nutrient requirements for primiparous sows according to the National Research Council (NRC, 2012).
E. coli lipopolysaccharide challenge
Within 24 h postweaning, thirty-two healthy male piglets from sixteen sows per treatment (one piglet per sow, average weight of the CON group: 6.11 ± 0.20 kg, average weight of the LY group: 6.23 ± 0.23 kg) were selected on the day of weaning (average weaning age: 19.2 days), and the weaned piglets were individually housed in a separate cage (0.9 × 0.4 × 0.6 m) prior to the immunological challenge. Lipopolysaccharide (LPS; E. coli serotype O55: B5; Sigma Chemical Inc., St Louis, MO, USA) was reconstituted in sterile saline (9 g/L NaCl) to prepare a solution at a concentration of 2.5 mg/L. Piglets in each treatment were intraperitoneally injected with 25 μg LPS per kg BW to induce an inflammatory response (Weber and Kerr, 2008; Gu et al., 2017), while the other sixteen were intraperitoneally injected with an equivalent amount of sterile saline. This resulted in four treatment groups (*n *= 8 per group): 1) control + saline (CS), 2) live yeast + saline (YS), 3) control + LPS (CLPS), and 4) live yeast + LPS (YLPS). The piglets were euthanized 4 h after lipopolysaccharide challenge (Gu et al., 2017). A schematic view of the experimental design is provided in Figure 1.
Schematic view of the experimental design (Created in https://BioRender.com). Abbreviations: CON, control; G, gestation; L, lactation; LY, live yeast; LPS, lipopolysaccharide.
Sample collection
Rectal temperature was measured using a digital thermometer before the LPS injection for baseline measurement and then hourly thereafter. At 4 h post-injection, piglets were euthanized for collecting mesenteric lymph node, liver, muscle, jejunal mucosa, and ileal mucosa. Mucosal samples were collected from gut sections corresponding to the jejunum and ileum by scraping the lining of the gastrointestinal tract using a glass slide. Mucosal scrapings were transferred into microfuge tubes, one tube contained TRIzol reagent (Invitrogen, Carlsbad, CA, USA) to preserve for RNA extraction, and the other tube stored samples for protein extraction, both samples were snap-frozen in liquid nitrogen and stored at −80°C until analysis. Additionally, a 5 cm segment of the mid jejunum was flushed with ice-cold phosphate-buffered saline and placed in 10% neutral-buffered formalin for histological analysis.
Jejunal histomorphology
Formalin-fixed jejunal samples were processed at the Purdue Histology and Phenotyping Laboratory (Purdue University, West Lafayette, IN, USA) for histology analysis. Six tissue sections containing intact lamina propria were selected from each piglet for measurement. Villus height (VH) and crypt depth (CD) were measured in micrometers using ImageJ software (NIH, Bethesda, MD, USA). For each sample, VH and CD values were averaged, and the villus height-to-crypt depth (VH:CD) ratio was calculated.
RT-qPCR
Gene expression in tissue samples was measured using a reverse transcription quantitative polymerase chain reaction (RT-qPCR), as previously described (Fu et al., 2024). Briefly, total RNA was extracted from frozen tissue samples including liver, muscle, lymph node, as well as the mucosa of jejunum and ileum using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The concentration of RNA was quantified using a Nanodrop 1000 instrument (Thermo Scientific, Waltham, MA, USA), and the RNA integrity was verified using 1% agarose gel electrophoresis. One μg of RNA was used for reverse transcription using M-MLV reverse transcriptase (Promega, Madison, WI, USA). Afterward, transcript amplification was performed using the CFX-96 real-time PCR detection system (Bio-Rad, Hercules, CA, USA) with the SYBR green RT-PCR mix (Bimake, Houston, TX, USA). The PCR protocol was as follows: 1 cycle of 95°C for 3 min and 40 cycles of 95°C for 10 s, followed by 30 s at an annealing temperature for each primer and 65°C for 30 s. Inflammatory and toll-like receptor 4 pathway-related markers, including interleukin (IL)-6, IL-1β, IL-8, IL-10, tumor necrosis factor (TNF)-α, toll-like receptor 4 (TLR4), inhibitor of kappa B alpha (IKBA), myeloid differentiation factor 88 (MYD88), nuclear factor kappa B (NF-κB), and antioxidant markers, superoxide dismutase (SOD)1, catalase (CAT), and glutathione peroxidase (GPX)1 were detected by RT-qPCR using their respective primers. Target mRNA levels were normalized to Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and analyzed using the 2^−ΔΔCt^ method, with results expressed as fold changes relative to the CS group (Livak and Schmittgen, 2001). The primer sequences are provided in Supplementary Table S1.
ELISA
Total protein levels in the ileal mucosal homogenates were determined using a Pierce BCA protein assay (Thermo Scientific, Waltham, MA, USA). Concentrations of TNF-α and IL-10 were quantified using respective commercial ELISA kits TNF-α and IL-10 (Life Diagnostics, Inc., West Chester, PA, USA) following the manufacturer’s protocols. Data were expressed as pg/mg of protein.
Western blotting
The methods for analyzing tight junction proteins by Western blot were described in Fu et al. (2024). Briefly, ileal and jejunal mucosa samples were homogenized and lysed in 1 × radio-immunoprecipitation assay buffer supplemented with 1% phosphatase and protease inhibitor cocktail (Thermofisher, Waltham, MA, USA) using TissueLyser (Qiagen, Valencia, CA, USA) at 30 Hz twice for 2 min. Protein concentrations were determined by a bicinchoninic acid assay kit (Sigma-Aldrich, St Louis, MO, USA). Proteins were resolved on a 10% SDS polyacrylamide gel and transferred onto nitrocellulose membranes (Bio-Rad, Hercules, CA, USA) using a semi-dry transfer system (Bio-Rad, Hercules, CA, USA). After transfer, membranes were cut into sections according to molecular weight from the marker lane and blocked with 5% bovine serum albumin dissolved in Tris-buffered saline with 0.1% Tween at room temperature for 2 h, and then incubated with corresponding primary antibodies at a dilution of 1:1,000 overnight at 4°C. The following primary antibodies were used: anti-claudin-4 (Catalog No. 329400; Life Technologies, Carlsbad, CA, USA), anti-claudin-3, occludin (Catalog No. ab15102 and ab31721, respectively; Abcam, Cambridge, MA, USA), anti-β-Actin (13E5) rabbit mAb, and E-cadherin (#4970 and #3195; Cell Signaling Technology, MA, USA). Membranes were washed at least four times with TBST and then incubated with corresponding secondary antibodies (anti-rabbit or anti-mouse IgG-horseradish peroxidase; Cell Signaling Technology, Danvers, MA, USA) at a dilution of 1:25,000 at room temperature for 1 h. Protein bands were visualized on a FluorChem imager (Proteinsimple, San Jose, CA, USA) using immobilon chemiluminescent HRP substrate (Millipore, Billerica, MA, USA). Raw densitometric readings were obtained and quantified using ImageJ (NIH, Bethesda, MD, USA) and averaged for eight samples from each treatment. The relative abundance of each target protein was expressed as the ratio of target protein to β-actin.
Statistical analysis
Data was analyzed by two-way analysis of variance using the PROC MIXED procedure of SAS (SAS Inst. Inc., Cary, NC, USA) for a split-plot design, with sow diet as the whole-plot and immunological challenge as the sub-plot. The Shapiro-Wilk test was used to analyze the normality of the data. Significance was set at *P *≤ 0.05, and a tendency was defined at 0.05 < *P *≤ 0.10. Results were illustrated using GraphPad Prism 7.03 (GraphPad Software Inc., San Diego, CA, USA).
Results
Effects of maternal dietary live yeast supplementation on rectal temperature
At 2 h post-injection, LPS challenge led to increased rectal temperature compared with saline group, and this effect was still observable up to 4 h post-injection (*P *< 0.05, Figure 2). At 3 h post-injection, piglets in the YLPS group tended to have a lower rectal temperature compared to those in the CLPS group (*P *= 0.09).
*Effects of maternal dietary live yeast supplementation on rectal temperature of newly weaned piglets in response to a lipopolysaccharide challenge (n = 8). Abbreviations: CS, control + saline; YS, yeast + saline; CLPS, control + lipopolysaccharide; YLPS, yeast + lipopolysaccharide. Indicates significances (P ≤ 0.05); #Represents trends (0.05 < P ≤ 0.10).
Effects of maternal dietary live yeast supplementation on expression of inflammatory- and antioxidant-related markers in the intestinal mucosa
In the jejunal mucosa, LPS challenge increased mRNA abundance of GPX1 compared with the saline group (*P *= 0.05, Figure 3A), whereas mRNA abundance of SOD1 and CAT was unaffected (*P *> 0.05). In addition, piglets from LY-supplemented sows tended to have lower mRNA levels of IL-8 (*P *= 0.09), while neither maternal diet nor LPS challenge influenced mRNA abundance of inflammatory cytokines, including IL-6, IL-10, TNF-α, and IL-1β (*P *> 0.05).
*Effects of maternal dietary live yeast supplementation on mRNA abundance of inflammatory- and antioxidant-related markers in the (A) jejunal and (B) ileal mucosa of newly weaned piglets in response to a lipopolysaccharide challenge (n = 8). Indicates significances (P ≤ 0.05); #Represents trends (0.05 < P ≤ 0.10). Abbreviations: CAT, catalase; CS, control + saline; CLPS, control + lipopolysaccharide; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; GPX1, Glutathione peroxidase 1; IL, interleukin; SOD, superoxide dismutase; TNF, tumor necrosis factor; YS, yeast + saline; YLPS, yeast + lipopolysaccharide.
In the ileal mucosa, piglets from LY-supplemented sows had higher mRNA abundance of IL-6, IL-10, TNF-α, and IL-1β (*P *< 0.05, Figure 3B), whereas no effect of maternal diet was observed for mRNA abundance of IL-8 or for LPS-induced effects in these genes (*P *> 0.05). Expression of antioxidant-related genes, including SOD1, GPX1, and CAT in the ileal mucosa did not differ in response to maternal diet or LPS challenge (*P *> 0.05). Additionally, TNF-α concentrations in the ileal mucosa were higher in piglets from CON sows than those from LY-supplemented sows (*P *< 0.05; Table 2), whereas no effect of LPS challenge was detected (*P *> 0.05). An interaction between maternal diet and LPS challenge was observed for both TNF-α and IL-10 concentrations (*P *= 0.05).
Effects of maternal dietary live yeast supplementation on the expression of inflammatory- and antioxidant-related markers in the liver and muscle
In the liver, mRNA abundance of CAT was downregulated by LPS (*P *< 0.05, Table 3), while no maternal dietary effect was observed (*P *> 0.05). Neither maternal diet nor LPS challenge influenced mRNA abundance of SOD1 and GPX1 (*P *> 0.05). The mRNA abundance of both pro- and anti-inflammatory cytokines, including TNF-α, IL-6, and IL-10, was upregulated in the LPS-challenged groups (*P *< 0.05), but maternal diet had no effects on these genes (*P *> 0.05). LPS challenge also elevated the mRNA abundance of TLR4, MYD88, IKBA, and NF-κB (*P *< 0.05), while effects of maternal diet were detected for NF-κB (*P *< 0.05) and a tendency was observed for TLR4 (*P *= 0.07), with piglets from LY-supplemented sows exhibiting lower mRNA abundance under LPS challenge compared with those from CON sows. In addition, interactions were observed for mRNA abundance of TLR4 and NF-κB (*P *< 0.05).
In the muscle, a trend toward an interaction between maternal diet and LPS challenge was observed for mRNA abundance of GPX1 (*P *= 0.10; Supplementary Table S2), along with a tendency for higher mRNA abundance of IL-6 in LPS-challenged piglets (*P *= 0.09). However, neither maternal diet nor LPS challenge affected mRNA abundance of SOD1, CAT, IL-10, or TNF-α (*P *> 0.05).
Effects of maternal dietary live yeast supplementation on expression of inflammatory- and antioxidant-related markers in the lymph node
In the lymph node, LPS challenge reduced mRNA abundance of SOD1 compared with the saline group (*P *< 0.05, Table 3), while no effect of maternal diet was observed (*P *> 0.05). Neither maternal diet nor LPS challenge affected GPX1 expression (*P *> 0.05). Piglets from LY-supplemented sows had lower gene expression of CAT than those from CON sows (*P *< 0.05), while no effect of LPS challenge was detected (*P *> 0.05). In response to LPS challenge, piglets from CON sows showed higher mRNA abundance of IL-10 than those from LY-supplemented sows (*P *< 0.05), with an interaction between maternal diet and LPS challenge observed for IL-10 (*P *< 0.05). Neither maternal diet nor LPS challenge affected mRNA abundance of IL-6 (*P *> 0.05), whereas a trend toward an interaction between maternal diet and LPS challenge was observed for mRNA abundance of TNF-α (*P *= 0.10). In addition, LPS challenge upregulated mRNA abundance of TLR4 and MYD88 compared with the saline group (*P *< 0.05), while no effects of maternal diet were detected (*P *> 0.05). Furthermore, piglets from LY-supplemented sows had lower mRNA abundance of NF-κB and MYD88 than those from CON sows (*P *< 0.05); however, neither maternal diet nor LPS challenge affected mRNA abundance of IKBA (*P *> 0.05).
Effects of maternal dietary live yeast supplementation on jejunal histomorphology and tight junction protein expression in the intestinal mucosa
Jejunal villus height and crypt depth were reduced in LPS-challenged piglets compared with the saline group (P < 0.05, Figure 4), whereas no effect of maternal diet was observed (*P *> 0.05). Neither maternal diet nor LPS challenge affected the villus-to-crypt ratio (*P *> 0.05). Piglets from LY-supplemented sows exhibited higher E-cadherin protein abundance in the jejunal mucosa compared with those from CON sows (*P *< 0.05; Figure 5A and Supplementary Figure S1—see online supplementary material for a colour version of this figure), while no maternal dietary effects were detected for protein abundance of occludin, claudin-3, and claudin-4 (*P *> 0.05). An interaction between maternal diet and LPS challenge was observed for occludin abundance (*P *< 0.05), and LPS challenge increased claudin-3 protein abundance in the jejunal mucosa (*P *< 0.05). In the ileal mucosa, an interaction between maternal diet and LPS challenge was observed for claudin-4 protein abundance (*P *< 0.05, Figure 5B and Supplementary Figure S1—see online supplementary material for a colour version of this figure), whereas neither maternal diet nor LPS challenge affected the protein abundance of E-cadherin, occludin, claudin-3, and claudin-4 (*P *> 0.05).
Effects of maternal dietary live yeast supplementation on jejunal histomorphology of newly weaned piglets in response to a lipopolysaccharide challenge (n = 8). Left panel: representative image from 1 piglet per treatment; Right panel: quantification of 8 samples per treatment. Abbreviations: CD, crypt depth; CS, control + saline; YS, yeast + saline; CLPS, control + lipopolysaccharide; YLPS, yeast + lipopolysaccharide; VH, villus height; VH: CD, villus height-to-crypt depth ratio. Magnification: × 10.
*Effects of maternal dietary live yeast supplementation on tight junction protein expression in the (A) jejunal and (B) ileal mucosa of newly weaned piglets in response to a lipopolysaccharide challenge (n = 8). a-bLeast squares means without a common superscript differ at P < 0.05. Indicates effect of maternal diet (P < 0.05). Left panel: representative western blot; Right panel: quantification of 8 samples per treatment. Abbreviations: CS, control + saline; YS, yeast + saline; CLPS, control + lipopolysaccharide; YLPS, yeast + lipopolysaccharide.
Discussion
Maternal transmission of immunomodulatory components occurs both prenatally and postnatally. During gestation, inflammatory mediators, microbial metabolites, and other bioactive compounds can cross the placenta, whereas postnatally, cytokines, immunoglobulins, and growth factors are delivered through colostrum and milk, thereby shaping neonatal immune function and early growth (Gormley et al., 2024). Therefore, dietary intervention during pregnancy and lactation represents a strategic approach for modulating intestinal development, immune resilience, and antioxidant defenses in the offspring. In the present study, we investigated the effects of dietary live yeast (LY) supplementation to sows during late pregnancy and lactation on inflammatory and antioxidant markers in various tissues of newly weaned piglets subjected to a LPS challenge.
The LPS challenge led to elevated rectal temperature at 2 h post-injection and lasted until 4 h, confirming the establishment of an acute inflammatory response. This is in agreement with previous studies that reported LPS-induced hyperthermia in piglets (Carroll et al., 2004). At 3 h post-injection, piglets from LY-supplemented sows (YLPS) exhibited a trend toward lower rectal temperatures compared with piglets from CON sows (CLPS), suggesting that maternal dietary LY supplementation might attenuate the LPS-induced febrile response. Intestinal barrier dysfunction contributes to inflammation and oxidative stress by allowing the translocation of luminal components, such as LPS, bacteria, and antigens, into systemic circulation (Ghosh et al., 2020). This translocation activates immune response, leading to release of pro-inflammatory cytokines both locally and systemically (Kim et al., 2012). Although the LPS challenge had limited effects on local inflammatory responses in the jejunal and ileal mucosa, including the mRNA abundance of IL-6, IL-10, TNF-α, and IL-1β, piglets from LY-supplemented sows exhibited higher ileal mucosal mRNA levels of both pro- and anti-inflammatory cytokines (IL-6, TNF-α, IL-1β, and IL-10). The increased expression of these cytokines may be attributed to the immune-priming properties of β-glucans (a key component of the LY cell wall), which may have altered the immune properties of colostrum or milk during the suckling phase (Broadway et al., 2015). Moreover, a tendency for lower mRNA level of IL-8 was observed in the jejunal mucosa of piglets from LY-supplemented sows. IL-8 is a chemokine primarily involved in the recruitment, activation, and migration of neutrophils to sites of mucosal injury or infection (Matsushima et al., 2022). The reduced IL-8 expression in piglets from LY-supplemented sows suggests a reduced pro-inflammatory response in the gut mucosa of the pigs (Kaiserman et al., 2022). In agreement with this, these piglets also exhibited lower TNF-α concentrations in the ileal mucosa and a higher E-cadherin protein expression in the jejunal mucosa compared with piglets from CON sows. E-cadherin is a marker of cell-cell adhesion, and higher mucosal E-cadherin abundance may indicate a stronger mucosal barrier capable of limiting neutrophil infiltration during inflammatory response (Lialios and Alimperti, 2025). Therefore, maternal immunomodulatory benefits conferred by LY supplementation, possibly through alterations in the immune properties of colostrum and milk, may help suppress excessive mucosal inflammation in piglets (Lewicka et al., 2019). Although maternal systemic and metabolic biomarkers were not measured in the present study, previous studies have reported that maternal dietary supplementation with LY increases plasma immunoglobulin G and decreases liver-associated enzyme activities such as aspartate aminotransferase and gamma-glutamyl transpeptidase in sows (Peng et al., 2020), which may reflect an improved immunometabolic status in LY-supplemented sows. These maternal alterations, together with LY-induced changes in the immunological properties of colostrum and milk, may modify the immune environment transmitted to offspring both in utero and postnatally, through colostrum and milk, thereby contributing to the attenuated mucosal cytokine responses observed in YLPS piglets.
The liver serves as a central organ for metabolism and detoxification, responsible for processing toxins, purifying the blood, and eliminating waste products generated by other tissues (Zhou et al., 2022). Under normal living conditions, liver-resident macrophages in piglets are capable of clearing low levels of circulating LPS from the bloodstream (Guo et al., 2025). However, accumulation of LPS beyond the capacity of the liver causes hepatocyte damage and activates hepatic macrophages, triggering the release of inflammatory cytokines and the production of reactive oxygen species (Kaur et al., 2006). In the current study, LPS reduced mRNA abundance of antioxidant enzyme catalase (CAT) and upregulated inflammatory cytokines including IL-6, IL-10, and TNF-α in the liver, which is consistent with previous studies showing that LPS reduces antioxidant capacity and promotes the release of inflammatory cytokines (Wen et al., 2023; Yu et al., 2023). Moreover, LPS activates toll-like receptor 4 (TLR4) signaling in both hepatocytes and Kupffer cells, initiating a cascade of intracellular events central to innate immune activation (Nakamoto and Kanai, 2014). Upon LPS binding, TLR4 recruits the adaptor protein myeloid differentiation factor 88 (MYD88), which activates downstream kinases that phosphorylate the inhibitor of NF-κB, IKBA. Phosphorylated IKBA is subsequently ubiquitinated and degraded, thereby allowing NF-κB to translocate from the cytoplasm to the nucleus and induce transcription of pro-inflammatory cytokines such as TNF-α and IL-6 (Pålsson-McDermott and O’Neill, 2004). In the present study, LPS challenge activated markers of the TLR4-MYD88-NF-κB signaling pathway in piglets, as evidenced by increased the hepatic mRNA abundance of TLR4, MYD88, NF-κB, IL-6, and TNF-α. We also observed an increase in IKBA mRNA abundance, which likely reflects the NF-κB-mediated negative feedback commonly observed during inflammatory activation (Hoffmann et al., 2002). Notably, YLPS piglets exhibited lower hepatic NF-κB and TLR4 mRNA abundance compared with CLPS piglets, suggesting a potential attenuation of innate immune activation. This modulation may be partially attributable to yeast-derived nucleotides. For instance, Li et al. (2022a) reported that maternal nucleotide supplementation inhibited TLR4 and NF-κB expression in LPS-treated piglets. However, the specific anti-inflammatory components of LY remain to be identified.
Lymph nodes are essential peripheral immune organs in mammals, characterized by a highly organized structure that facilitates migration and activation of immune cells, and functions as immunological checkpoints in host defense (Zhang et al., 2023). Despite their importance, limited studies have examined the immune response in lymph nodes, particularly in livestock species such as pigs. In the present study, LPS reduced the mRNA abundance of SOD1 in the lymph node, a key antioxidant enzyme that mitigates oxidative damage by catalyzing the dismutation of superoxide radicals (Eleutherio et al., 2021). This reduction suggests that LPS-induced inflammation may be accompanied by compromised antioxidant defense mechanisms in both hepatic and lymphatic tissues. In addition, the LPS challenge increased the mRNA abundance of markers in the TLR4-mediated inflammatory pathway, including TLR4 and MYD88 in challenged piglets. It also elevated IL-10 expression, reflecting a compensatory anti-inflammatory response (Alexander et al., 2021). However, maternal dietary LY supplementation reduced mRNA abundance of MYD88 and NF-κB in the lymph nodes of piglets. Although NF-κB activation is primarily regulated post-translationally (Hayden and Ghosh, 2012), reductions in NF-κB and MYD88 transcription may reflect a lower signaling capacity of the pathway to initiate inflammatory responses. Furthermore, the observed interaction for IL-10 between maternal diet and LPS challenge suggests that maternal LY modulates the piglet’s regulatory response to LPS. Rather than indicating immunosuppression, this reduced activation may reflect a more balanced inflammatory response, whereby maternal LY supplementation enhances tolerance and prevents excessive inflammatory response to LPS (Liu et al., 2023). Our recent findings indicate that gestational and lactational supplementation of LY to sows increases the abundance of Ig-like domain-containing protein and complement proteins (Complement C8 alpha chain and C1q domain-containing proteins) in the colostrum and IgG heavy chain in milk (Fu et al., 2025). Similar findings also suggest that maternal dietary LY supplementation improves IgG concentrations in the colostrum and subsequently increases plasma IgG levels in piglets (Jang et al., 2013). Thus, these enhancements in colostrum and milk immunological quality may modulate the development and responsiveness of the offspring’s immune system (Reyes-Camacho et al., 2020).
The LPS challenge induced a decrease in villus height and crypt depth in the jejunum of piglets, indicating the potential intestinal injury following endotoxin exposure. This is consistent with previous findings that LPS induces morphological alterations in the digestive tract and increased mucosal permeability (Li et al., 2022a). The intestinal barrier is primarily composed of epithelial cells that are tightly interconnected by adherence and junction complexes, including E-cadherin, claudins, and occludin, to form a selectively permeable barrier that maintains intestinal integrity (Zhang et al., 2015). Reduced expression of these proteins may indicate a disruption of the intestinal barrier, contributing to increased intestinal permeability and facilitating the translocation of luminal antigens and bacteria into systemic circulation (Liu et al., 2023). In the current study, the effect of maternal LY supplementation on tight junction proteins was dependent on LPS challenge, as reflected in the different abundances of jejunal occludin and ileal claudin-4. In addition, piglets from LY-supplemented sows had a higher protein abundance of E-cadherin in the jejunal mucosa compared with piglets from CON sows, regardless of LPS challenge. This maternal effect suggests that LY supplementation may enhance epithelial cell adhesion and support mucosal integrity under both normal and challenged conditions. This pattern is consistent with our previous findings that gestational and lactation LY supplementation increases the abundance of junctional proteins such as E-cadherin, occludin, and claudin-3 in the small intestines of suckling and newly weaned piglets (Fu et al., 2024). Maternal nutrition modulates the maternal entero-mammary route and colostrum or milk composition, which may affect epithelial development and tight junction expression of offspring (Li et al., 2022b). In addition, our data indicate that maternal LY supplementation enriched bacterial taxa associated with SCFA production in piglets during suckling and early postweaning period (Y. Fu, unpublished data). The SCFAs, especially butyrate, strengthen the intestinal barrier by activating AMP-activated protein kinase, which promotes the assembly and reorganization of tight junction proteins (Peng et al., 2009). Future studies are needed to identify the specific yeast-derived components and metabolites that mediate these maternal programming effects on neonatal intestinal development and immune maturation, whether through modulation of placental nutrient and immune signaling or through direct transfer to the offspring via colostrum and milk.
Overall, supplementing sow diets with LY partially inhibited the inflammatory response to LPS challenge in the liver, lymph node, and intestinal mucosa of newly weaned piglets, supporting the possibility that LY exerts immunomodulatory effects through maternal-neonatal transfer. However, there is a nuanced effect of LY in the modulation of the inflammatory response to LPS that is tissue specific, illustrating the complexity of whole-body immune regulation.
Conclusion
In conclusion, although the LPS challenge induced an inflammatory response characterized by elevated rectal temperature, intestinal barrier disruption, oxidative stress, and tissue-specific inflammatory response, gestational and lactational dietary supplementation with LY partially alleviated these adverse effects. Key points of LY effects included suppression of TLR4-mediated pro-inflammatory signaling pathways in the liver and lymph nodes, reducing local inflammation, and improving adherens junction protein expression in the intestinal mucosa. These findings suggest that maternal dietary live yeast supplementation could enhance the immune resilience of piglets, offering a promising nutritional strategy to mitigate weaning stress and inflammation during the early postweaning period.
Supplementary Material
skaf435_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alexander A. F. , Kelsey I., Forbes H., Miller-Jensen K. 2021. Single-cell secretion analysis reveals a dual role for IL-10 in restraining and resolving the TLR 4-induced inflammatory response. Cell Rep. 36(12):109728. 10.1016/j.celrep.2021.10972834551303 PMC 8995750 · doi ↗ · pubmed ↗
- 2Arslan S. , Erbas M., Tontul I., Topuz A. 2015. Microencapsulation of probiotic Saccharomyces cerevisiae var. boulardii with different wall materials by spray drying. LWT–Food Sci. Technol. 63(1):685–690. 10.1016/j.lwt.2015.03.034 · doi ↗
- 3Blavi L. , Solà-Oriol D., Llonch P., López-VergéS., Martín-Orúe S. M., Pérez J. F. 2021. Management and feeding strategies in early life to increase piglet performance and welfare around weaning: a review. Animals. 11(2):302. 10.3390/ani 1102030233503942 PMC 7911825 · doi ↗ · pubmed ↗
- 4Broadway P. , Carroll J., Sanchez N. 2015. Live yeast and yeast cell wall supplements enhance immune function and performance in food-producing livestock: a review. Microorganisms. 3(3):417–427. 10.3390/microorganisms 303041727682097 PMC 5023245 · doi ↗ · pubmed ↗
- 5Burdick Sanchez N. C. , Broadway P. R., Carroll J. A. 2021. Influence of yeast products on modulating metabolism and immunity in cattle and swine. Animals. 11(2):371. 10.3390/ani 1102037133540746 PMC 7913008 · doi ↗ · pubmed ↗
- 6Carroll J. A. , Fangman T. J., Hambach A. K., Wiedmeyer C. E. 2004. The acute phase response in pigs experimentally infected with Escherichia coli and treated with systemic bactericidal antibiotics. Livest. Prod. Sci. 85(1):35–44. 10.1016/S 0301-6226(03)00115-5 · doi ↗
- 7Chance J. A. , De Rouchey J. M., Amachawadi R. G., Ishengoma V., Nagaraja T. G., Goodband R. D., Woodworth J. C., Tokach M. D., Kang Q., Loughmiller J. A. et al. 2022. Influence of yeast-based pre- and probiotics in lactation and nursery diets on nursery pig performance and antimicrobial resistance of fecal Escherichia coli. J. Anim. Sci. 100(6):skac 166. 10.1093/jas/skac 16635524733 PMC 9167580 · doi ↗ · pubmed ↗
- 8Eleutherio E. C. A. , Silva Magalhães R. S., de Araújo Brasil A., Monteiro Neto J. R., de Holanda Paranhos L. 2021. SOD 1, more than just an antioxidant. Arch. Biochem. Biophys. 697:108701. 10.1016/j.abb.2020.10870133259795 · doi ↗ · pubmed ↗