Developmental single-cell atlas of coronary vessel growth and cardiomyocyte interaction in zebrafish
Muhammad Abdul Rouf, Gülsüm Kayman Kürekçi, Shaoqiu Zhang, Stéphanie Larrivée Vanier, Sarah M. Kamel, Ann Nee Lee, Ruey-Bing Yang, Shih-Lei Lai, Rubén Marín-Juez

TL;DR
This study maps coronary vessel development in zebrafish, showing how blood vessels and heart muscle cells interact during growth.
Contribution
The first comprehensive single-cell atlas of coronary vessel development in zebrafish is created, revealing cellular and molecular milestones.
Findings
Coronary vessels act as scaffolds to guide cardiomyocyte growth during development.
Vegfa signaling is critical for coronary vessel–cardiomyocyte interactions.
A single-cell RNA sequencing atlas identifies new coronary markers and dynamic cellular interactions.
Abstract
Cardiac morphogenesis requires the intricate coordination of different cell types and molecular cues. Coronary vessel formation is essential for heart development, yet how coronary vessels grow and contribute to ventricular wall formation remains poorly understood. Here, we combine high-resolution imaging and new genetic tools to systematically analyze coronary vasculature development in zebrafish at each millimeter increment from 7 to 30 mm of body length and identify cellular and molecular milestones defining four distinct coronary network developmental stages. Our data show how coronary vessels expand, pattern, specify and act as vascular scaffolds to guide cardiomyocyte growth throughout development. Manipulating coronary network growth by Vegfa signaling perturbation through gain- and loss-of-function approaches disrupts coronary vessel–cardiomyocyte interactions and cardiomyocyte…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
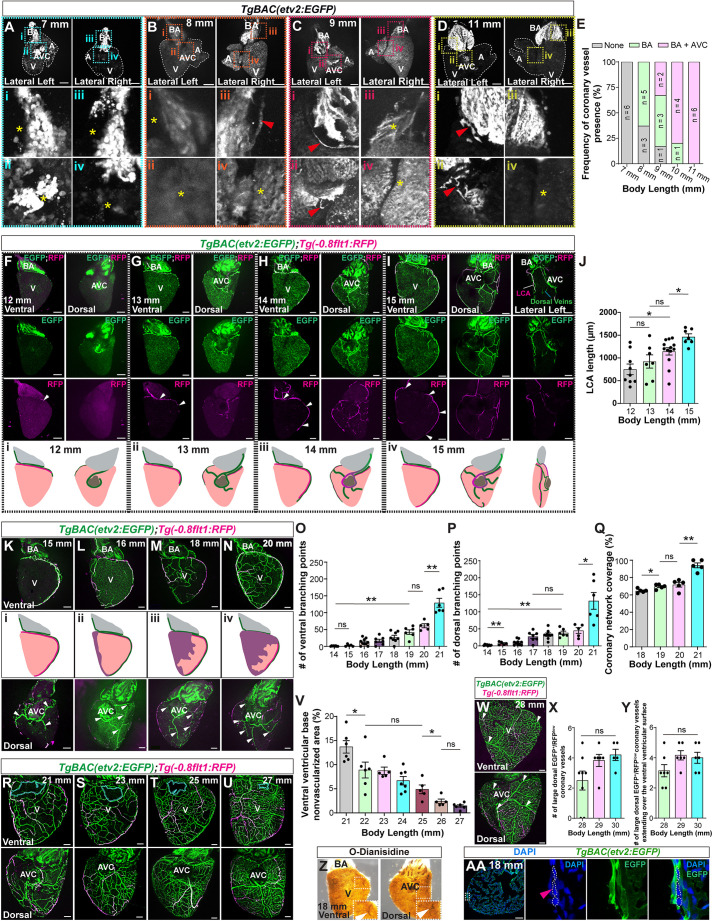 Fig. 1
Fig. 1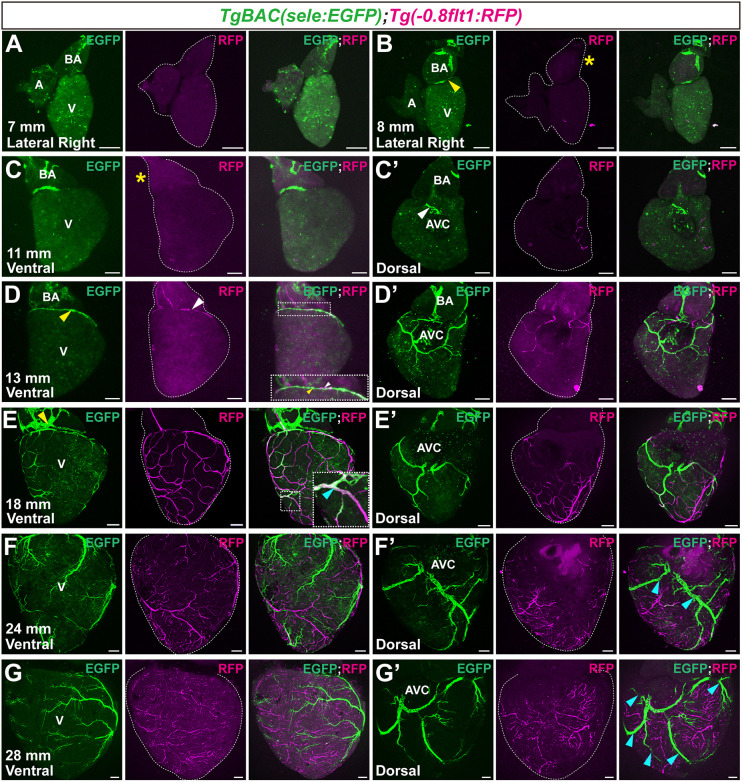 Fig. 2
Fig. 2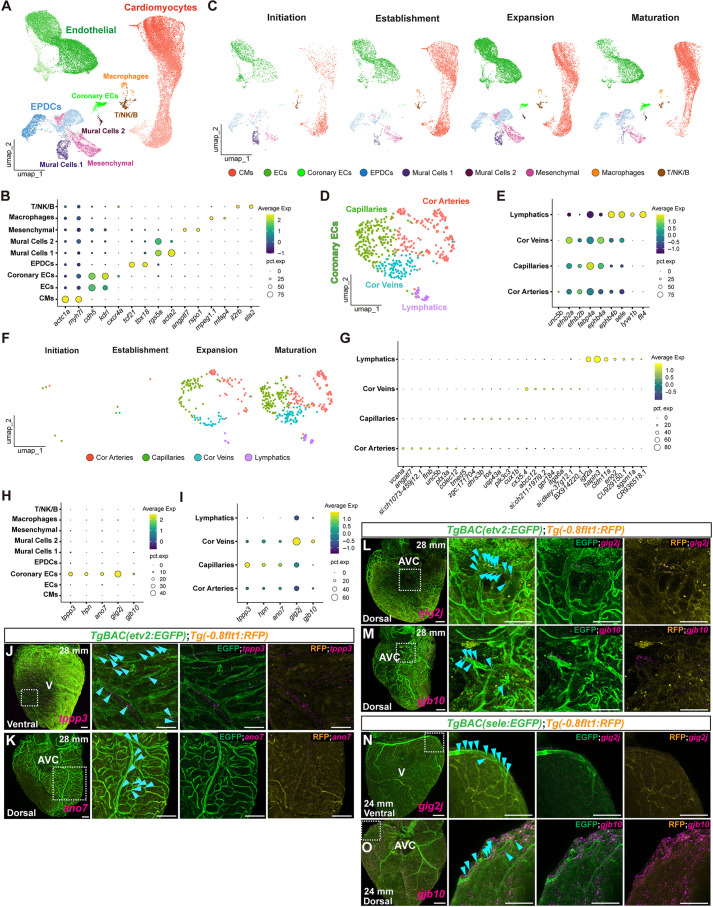 Fig. 3
Fig. 3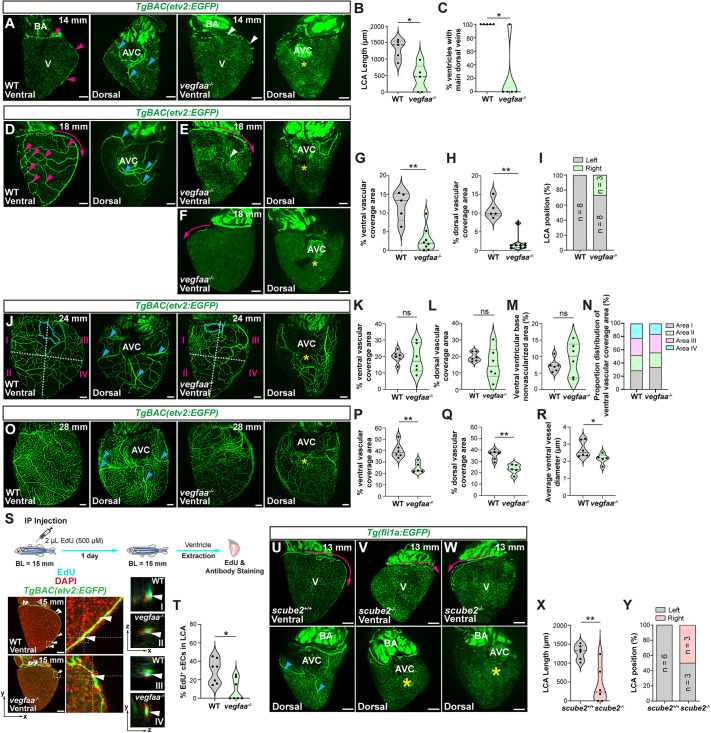 Fig. 4
Fig. 4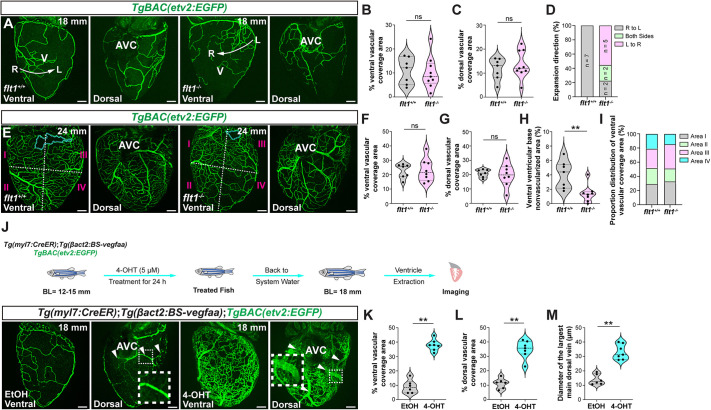 Fig. 5
Fig. 5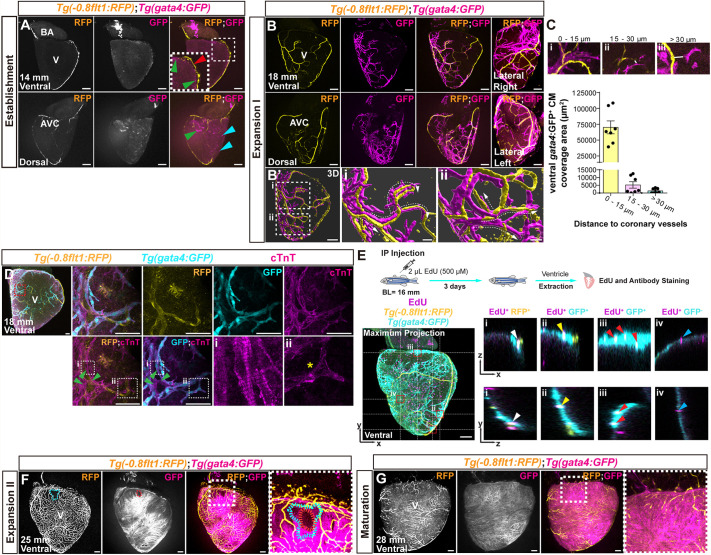 Fig. 6
Fig. 6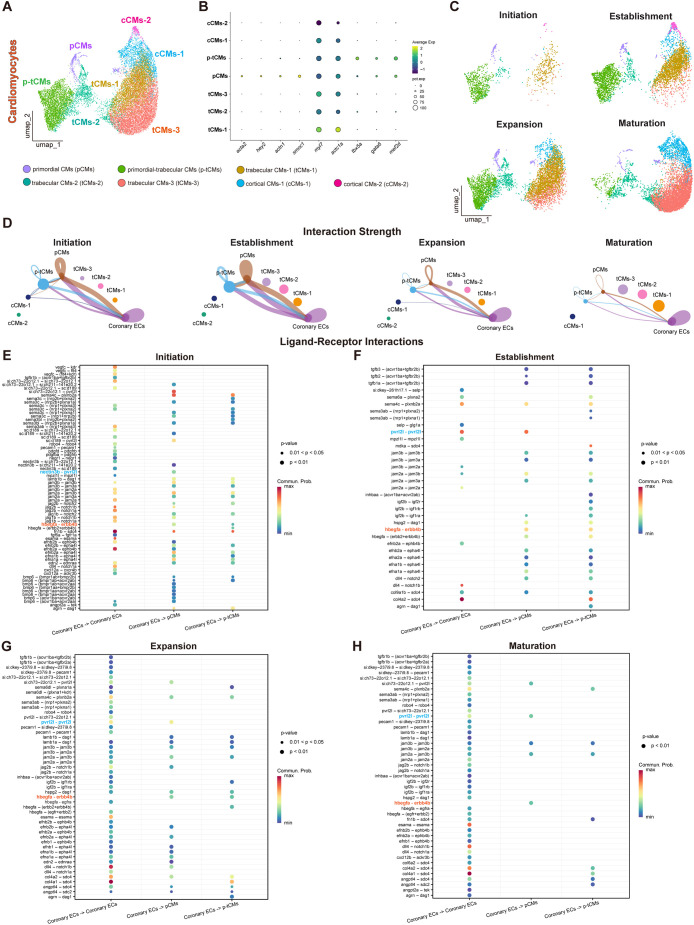 Fig. 7
Fig. 7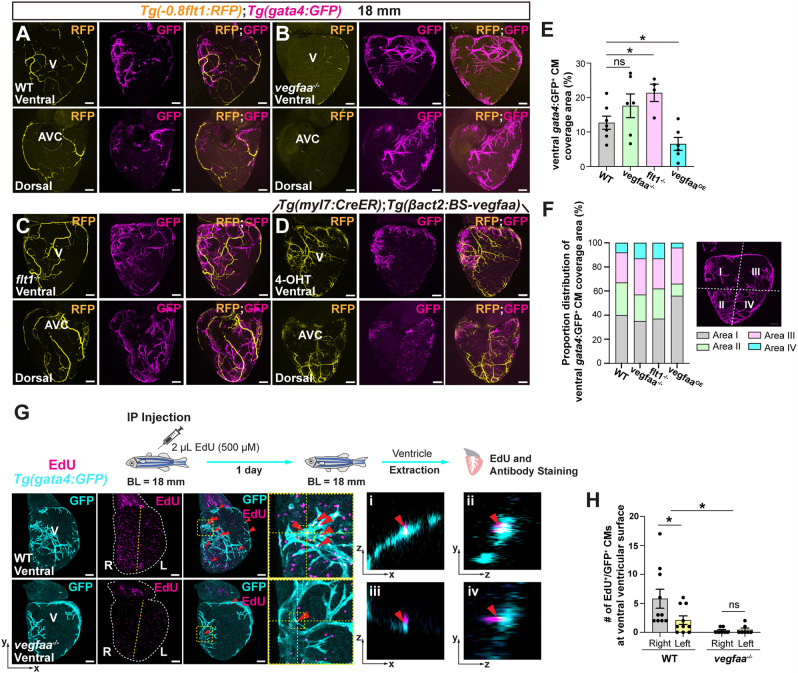 Fig. 8
Fig. 8- —Fonds de recherche du Québechttp://dx.doi.org/10.13039/501100020951
- —Natural Sciences and Engineering Research Council of Canadahttp://dx.doi.org/10.13039/501100000038
- —Canadian Institutes of Health Researchhttp://dx.doi.org/10.13039/501100000024
- —Natural Sciences and Engineering Research Council of Canada and the Canadian Institutes of Health Research
- —Université de Montréalhttp://dx.doi.org/10.13039/501100005242
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCongenital heart defects research · Zebrafish Biomedical Research Applications · Developmental Biology and Gene Regulation
INTRODUCTION
The formation of a functional coronary vascular network is essential to support cardiac muscle growth. As cardiac morphogenesis progresses, coronary circulation arises to meet the increasing metabolic demands of the thickening ventricular wall (D'Amato et al., 2022; Red-Horse et al., 2010; Tian et al., 2015; Tomanek, 2005). In mammals, coronary vessels develop during embryogenesis primarily from epicardium-derived cells (EPDCs), the endocardium and sinus venosus-derived endothelial cells (ECs) that invade the subepicardial and myocardial spaces, ultimately assembling into an intricate vascular network (D'Amato et al., 2022; Nakajima and Imanaka-Yoshida, 2013; Red-Horse et al., 2010; Riley, 2012; Wu et al., 2012). This process involves tightly coordinated interactions among coronary ECs, cardiomyocytes (CMs) and epicardial cells (Günthel et al., 2018; Gupta and Poss, 2012; Harrison et al., 2015; Marín-Juez et al., 2019; Pires-Gomes and Pérez-Pomares, 2013; Red-Horse et al., 2010; Wu et al., 2012).
Taking advantage of its strengths as a model organism and the rich repertoire of genetic tools available, recent studies have begun to investigate the mechanisms of coronary vessel formation in zebrafish (Bakis et al., 2023; Chiba et al., 2025; Duca et al., 2024; Gancz et al., 2019; Harrison et al., 2015; Karra et al., 2018; Lowe et al., 2019; Marín-Juez et al., 2019). At approximately 45 days post-fertilization (dpf), ECs from the atrioventricular canal (AVC) sprout and expand over the dorsal ventricular surface to form the coronary plexus (Chiba et al., 2025; Harrison et al., 2015). At the same stage, a subset of trabecular CMs breach the primordial CM layer to form the cortical myocardium (Gupta and Poss, 2012; Gupta et al., 2013). Cortical CMs follow the track of the developing coronary vasculature to populate the ventricular surface (Marín-Juez et al., 2019). A similar association between coronary ECs and CMs occurs during mouse heart development (DeBenedittis et al., 2022). Moreover, disruption of coronary development results in defective CM growth, mediated by impaired angiocrine signaling (Chiang et al., 2023; DeBenedittis et al., 2022; Rhee et al., 2018, 2021). These findings underscore the role of coronary vessels as active regulators of myocardial wall formation. However, the precise developmental dynamics of coronary vessel formation and interaction with CMs remain largely unknown.
Here, we combined high-resolution imaging, novel and established tissue-specific transgenic lines, functional manipulation of Vegfa signaling, and single-cell transcriptomics to construct a roadmap of coronary development. Spanning early emergence to maturation, we identify developmental milestones, define four distinct stages termed initiation, establishment, expansion and maturation, and show how the coronary network progressively populates the ventricular surface. These analyses reveal spatially coordinated proliferation between developing coronary ECs and adjacent CMs, supporting a vessel-guided model of myocardial expansion. Additionally, single-cell RNA sequencing (scRNA-seq) of 37,554 ventricular cells across developmental stages shows dynamic gene expression programs and cell type-specific interactions. Together, this integrative approach delineates the temporal and cellular architecture of coronary vascular development and identifies its instructive role in ventricular morphogenesis.
RESULTS
Coronary vasculature emergence and growth dynamics
To track coronary development, we used body length from the snout to the base of the tail as a proxy for developmental stage, as it more accurately reflects developmental progression than chronological age in zebrafish (Singleman and Holtzman, 2014). We used the TgBAC(etv2:EGFP) transgenic line to label coronary ECs and track coronary vessel emergence. When the body length was <8 mm, we observed no vessel-like structures on the heart (Fig. 1A,E). At this stage, etv2:EGFP expression was strong in the AVC, reported to be one of the origins of the coronary vasculature in zebrafish (Harrison et al., 2015) (Fig. 1A). As etv2 is a transcription factor expressed in endothelial progenitors (Sumanas and Lin, 2006; van Bueren and Black, 2012), it is possible that this upregulation reflects the activation of vasculogenesis. At 8 mm of body length, the first coronary vessel was observed on the bulbus arteriosus (BA) (Fig. 1B,E), consistent with the extension of the hypobranchial artery onto the BA (Mizukami et al., 2023). At 9 mm, a second vessel appeared from the AVC (Fig. 1C,E), consistent with previous findings (Harrison et al., 2015). By 11 mm, both BA and AVC vessels continued to grow around the BA-ventricular base junction and the AVC, respectively (Fig. 1D,E). Therefore, between 8 and 11 mm of body length the first two coronary vessels emerge and expand over the BA and AVC regions. We refer to this period of coronary development as the initiation stage (Fig. S1A).
*Characterization of coronary development. (A-D) Whole-mount TgBAC(etv2:EGFP) hearts from 7-mm (A), 8-mm (B), 9-mm (C) and 11-mm (D) -long zebrafish. High-magnification images of BA (i and iii) and AVC (ii and iv) regions are shown. Yellow asterisks indicate the absence of coronary vessels. Red arrowheads point to coronary vessels at the BA and AVC regions. (E) Frequency of coronary vessel presence in the BA and AVC regions in 7- to 11-mm-long zebrafish. (F-I) Whole-mount TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) ventricles from 12-mm (F), 13-mm (G), 14-mm (H) and 15-mm (I) -long zebrafish. Schematic representations of coronary vessel development at each of the stages are shown below (i-iv). White arrowheads point to the LCA expanding across the left ventricular curvature from base to apex. (J) LCA length in 12- to 15-mm-long zebrafish. (K-N) Whole-mount TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) ventricles from 15-mm (K), 16-mm (L), 18-mm (M) and 20-mm (N) -long zebrafish. Schematic representations of coronary network expansion (purple shadowed areas) are shown (i-iv). White arrowheads point to putative main dorsal veins. (O,P) Number of coronary branching points at the ventral (O) and dorsal (P) ventricular surfaces in 14- to 21-mm-long zebrafish. (Q) Percentage of coronary network coverage area at the ventral ventricular surface in 18- to 21-mm-long zebrafish. (R-U) Whole-mount TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) ventricles from 21-mm (R), 23-mm (S), 25-mm (T) and 27-mm (U) -long zebrafish. Cyan dotted lines outline the nonvascularized area at the ventral ventricular base. (V) Nonvascularized area percentage at the ventral ventricular base in 21- to 27-mm-long zebrafish. (W) Whole-mount TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) ventricle from a 28-mm-long zebrafish. White arrowheads point to EGFP+/RFPlow vessels. (X,Y) Number of large EGFP+/RFPlow coronary vessels on the dorsal ventricular surface (X) and extending over the ventral ventricular surface (Y). (Z) Erythrocytes in a whole-mount ventricle from an 18-mm-long zebrafish stained with O-dianisidine. Insets show magnifications of the boxed areas. White arrowheads point to vessel-like structures. (AA) TgBAC(etv2:EGFP) ventricular section from an 18-mm-long zebrafish stained for EGFP (coronary ECs, green) and DNA (DAPI, blue). Boxed area in left-hand image is shown at higher magnification on the right. Magenta arrowhead points to DAPI+ cells in the lumen of an etv2:EGFP+ coronary vessel. Dashed lines outline DAPI+ cells in the lumen of an etv2:EGFP+ coronary vessel. Data in graphs expressed as mean±s.e.m. ns, no significant difference. *P<0.05, *P<0.01 (two-tailed, Mann–Whitney U test). A, atrium; AVC, atrioventricular canal; BA, bulbus arteriosus; V, ventricle. Scale bars: 100 μm.
To assess vessel identity, we analyzed ventricles from TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) fish. The combination of these transgenes identifies coronary ECs (EGFP^+^/RFP^+^) and coronary arteries (EGFP^+^/RFP^high^) (Harrison et al., 2015; Marín-Juez et al., 2016, 2019). Between 12 and 15 mm of body length, both the BA and the AVC vasculature continued to expand. At 12 mm, the BA sprout reached the ventricular base and upregulates −0.8flt1:RFP, indicating arterial identity (Fig. 1F). This vessel continued to expand over the left ventricular curvature to reach the apex (Fig. 1G-J). Based on its localization along the left ventricular curvature and arterial identity, we termed this vessel as the left coronary artery (LCA). The AVC-derived vasculature expanded over the dorsal ventricular surface and formed additional branches that extended dorsally and ventrally along the left curvature toward the apex, with some dorsal vessels becoming EGFP^+^/RFP^high^ (Fig. 1F-J). We also observed a distinct dorsal EGFP^+^/RFP^low^ vessel, potentially of AVC origin, which was located dorsal to and close to the LCA (Fig. 1I). This vessel and the LCA extended together along the left ventricular curvature toward the apex (Fig. 1I). Therefore, between 12 and 15 mm of body length, the dorsal vascular plexus is established, and the LCA grows to reach the apex. We refer to this period as the establishment stage (Fig. S1A).
As development progressed from 15 to 20 mm of body length, coronary vessels continued to grow and branch across both the dorsal and ventral ventricular surfaces (Fig. 1K-P, Fig. S2A-D). The coronary plexus continued to populate the dorsal surface and extended ventrally, consistent with observations in other organisms (Kattan et al., 1999; Red-Horse et al., 2010). From 16 mm onward, the coronary network expanded across the right ventricular curvature and sprouted toward the LCA, establishing a right-to-left expansion pattern over the ventral surface (Fig. 1L-N,Q), similar to observations in mice (Chen et al., 2014). When zebrafish reached 20 mm of body length, the expanding plexus contacted the LCA (Fig. 1N). To measure this ventral expansion, we quantified the number of branching points and the extent of coronary network coverage (Fig. 1O-Q). Our data show that both parameters increased significantly between 15 and 19 mm and remained unchanged between 19 and 20 mm. At 21 mm, after the expanding front contacted the LCA, both branching points and coverage increased significantly again compared with previous stages. These data show that the coronary plexus undergoes consistent growth from 15 to 20 mm of body length and then enters a second expansion phase at 21 mm.
From 21 to 27 mm, the expanding ventral coronary network became increasingly dense and complex (Fig. 1R-U, Figs S2E-H and S3A-D). In contrast to previous stages, an increasing number of EGFP^+^/RFP^high^ vessels were observed, suggesting progressive arterialization (Fig. S2E-H). At this stage, part of the ventral ventricular base surface remained consistently nonvascularized until fish reached 27 mm of body length (Fig. 1R-V). We define the developmental window spanning from 15 to 27 mm of body length as the expansion stage (Fig. S1A) and subdivide it into ‘expansion I’ (15-20 mm, first wave of coronary branching and coverage increase) and ‘expansion II’ (21-27 mm, second expansion wave and presence of the ventral ventricular base nonvascularized area).
From 28 to 30 mm, the ventral ventricular base nonvascularized area disappeared as the coronary network fully covered the ventricular surface (Fig. S2I-K) and no further increase in network coverage was observed (Fig. S3E-H). The ventricle presents a well-defined dorsal network of two to five large EGFP^+^/RFP^low^ vessels, potentially main coronary veins, that in most instances expanded to reach the ventral surface (Fig. 1W-Y). Coronary arteries were well established with branches clearly visible on both ventricular surfaces (Fig. S2I-K). We define this stage as the maturation stage (Fig. S1A).
To determine the onset of coronary circulation, we performed O-dianisidine staining at different stages. Vessel-like structures were not clearly detectable until animals reached 18 mm of body length (Fig. 1Z, Fig. S4A,B). To further support this observation, we analyzed TgBAC(etv2:EGFP) ventricular sections and found that coronary vessels at this stage were indeed lumenized and contained DAPI^+^ cells (Fig. 1AA). Next, we performed 5-ethynyl-2-deoxyuridine (EdU) incorporation assays on TgBAC(etv2:EGFP) ventricles before (16 mm) and after (19 mm) the onset of coronary perfusion. We found that the number of EdU^+^ coronary ECs was increased at 19 mm compared with 16 mm (Fig. S4D-F), indicating that the onset of blood flow coincides with an increase in coronary EC proliferation.
TgBAC(sele:EGFP) labels coronary veins
Several transgenic lines have been developed to study arterio-venous development in zebrafish. However, while arterial reporters remain active in the adult heart, venous marker expression becomes restricted to cardiac lymphatics, preventing visualization of coronary veins (Harrison et al., 2015; Marín-Juez et al., 2016, 2019). To circumvent this limitation, we generated a TgBAC(sele:EGFP) zebrafish line. sele encodes E-selectin, a cell adhesion molecule expressed on the EC surface (Collins et al., 1991). In zebrafish embryos, sele is highly expressed in the posterior cardinal vein (PCV) (Sun et al., 2015).
To determine whether this transgene is specifically expressed in venous ECs, we examined its expression throughout development in combination with Tg(−0.8flt1:RFP) and Tg(dll4:TagRFP), both well-established arterial markers (Bussmann et al., 2010, 2011; Chong et al., 2011; Marín-Juez et al., 2016). We found strong sele:EGFP expression in venous vessels in adult fin rays (Fig. S5A) as well as in the PCV and venous intersegmental vessels (vISVs) at 5 dpf (Fig. S5B,C). We also analyzed TgBAC(sele:EGFP) in combination with Tg(lyve1b:dsRed) to visualize lymphatic vessels and found minimal EGFP expression in the thoracic duct (TD) at 5 dpf (Fig. S5D). Next, we analyzed coronary vein and artery development from initiation to maturation stages in TgBAC(sele:EGFP); Tg(−0.8flt1:RFP) ventricles. No vessel-like structures were found at 7 mm (Fig. 2A). At 8-11 mm (initiation stage), the first BA and AVC sele:EGFP^+^ vessels were observed (Fig. 2B-C′), suggesting venous identity of these emerging vessels. At 13 mm of body length (establishment stage), two adjacent vessels, one EGFP^+^/RFP^−^ and another one EGFP^−^/RFP^high^ (the developing LCA), developed across the ventricular base and expanded toward the left ventricular curvature (Fig. 2D,D′). At this stage, the dorsal coronary plexus was composed of EGFP^+^/RFP^−^ and EGFP^−^/RFP^high^ vessels. At 18 mm (expansion I stage), the venous plexus further developed dorsally and expanded ventrally following the previously observed right-to-left pattern (Fig. 2E). At this stage, some vessels near the expanding vascular front co-expressed both sele:EGFP and −0.8flt1:RFP, suggesting venous-to-capillary or arterial conversion (Hen et al., 2015; Red-Horse and Siekmann, 2019; Red-Horse et al., 2010; Xu et al., 2014) (Fig. 2E). At 24-28 mm (expansion II to maturation stages), the main coronary veins and arteries were distinguishable by marker expression, with sele:EGFP expression being restricted to coronary veins (Fig. 2F-G′). At this stage, the coronary veins primarily populated the dorsal ventricular half, with most extending ventrally, consistent with our data (Fig. 1W, Fig. S2I-K) and previous reports (Harrison et al., 2015; Marín-Juez et al., 2016).
TgBAC(sele:EGFP) labels coronary veins during development. (A) Whole-mount TgBAC(sele:EGFP); Tg(−0.8flt1:RFP) heart from a 7-mm-long zebrafish. (B-C′) Whole-mount TgBAC(sele:EGFP); Tg(−0.8flt1:RFP) hearts from 8-mm (B) and 11-mm (C,C′) -long zebrafish. Yellow arrowhead points to a sele:EGFP+ vessel in the BA-ventricular base junction. Yellow asterisks indicate absence of −0.8flt1:RFP expression. White arrowhead points to a sele:EGFP+ vessel around the AVC. (D-E′) Whole-mount TgBAC(sele:EGFP); Tg(−0.8flt1:RFP) ventricles from 13-mm (D,D′) and 18-mm (E,E′) -long zebrafish. (D) Yellow arrowhead points to an EGFP+/RFP− (vein) vessel. White arrowhead points to an EGFP−/RFPhigh (LCA, artery) vessel. (E) Yellow arrowhead points to BA EGFP+ lymphatics. Cyan arrowhead points to an EGFP+/RFP+ vessel. (F-G′) Whole-mount TgBAC(sele:EGFP); Tg(−0.8flt1:RFP) ventricles from 24-mm (F,F′) and 28-mm (G,G′) -long zebrafish. Cyan arrowheads point to main dorsal coronary veins. Insets show magnifications of the boxed areas. A, atrium; AVC, atrioventricular canal; BA, bulbus arteriosus; V, ventricle. Scale bars: 100 μm.
Although no vessel-like structures were observed on the ventricle at 7 mm (Fig. 2A), we observed sele:EGFP^+^/−0.8flt1:RFP^−^ cells on the BA. Previous studies have reported a population of BA lymphatic ECs at early developmental stages, before lymphatics populate the ventricle (Gancz et al., 2019; Harrison et al., 2019). Moreover, sele:EGFP was also lowly expressed in some TD lyve1b:dsRed^+^ lymphatics (Fig. S5D). We analyzed TgBAC(sele:EGFP); Tg(lyve1b:dsRed) ventricles and found that these cells co-express both reporters, indicating lymphatic identity (Fig. S5E,F). These data show that two distinct vascular populations co-exist on the BA at the initiation stage. Our analyses suggest that this population of BA lymphatics continue to expand at later stages (Fig. S5E-H), potentially reaching the ventricular surface during late expansion and maturation stages (Fig. S5I,J). Ventricular lymphatics only retained lyve1b:dsRed expression and closely followed coronary veins (Fig. S5J,K).
Collectively, these results show that TgBAC(sele:EGFP) is expressed by coronary venous ECs in zebrafish. This new marker enables dissection of venous identity, emergence and spatial distribution during coronary network development.
Single-cell profiling identifies dynamic cellular transitions and markers defining distinct coronary subtypes
To better understand the cellular composition and interactions during heart development, we performed scRNA-seq on zebrafish ventricles at the initiation (10 mm), establishment (14 mm), expansion (18 mm) and maturation (28 mm) stages (Fig. S6A).
We identified and annotated all major cardiac cell types present at each developmental stage based on the expression of established marker genes (Fig. 3A,B, Fig. S6D). Our analysis revealed stage-specific changes in cellular composition across the four stages (Fig. 3C, Fig. S6B-F). Notably, coronary EC populations were lower at the initiation and establishment stages but showed a significant increase during the expansion and maturation stages. Similarly, macrophage and CM populations also increased progressively (Fig. 3C, Fig. S6E,F). We also observed changes in other cell populations, including EPDCs and mesenchymal cells (Fig. 3C, Fig. S6E,F). Interestingly, we identified two distinct populations of mural cells designated as ‘mural cells 1’ and ‘mural cells 2’. The mural cells 1 number fluctuated throughout development, whereas mural cells 2 displayed a pattern similar to that of coronary ECs (Fig. 3C, Fig. S6E,F). To further resolve heterogeneity among cell populations potentially involved in coronary development, we performed secondary subclustering analyses of ECs, EPDCs and mural cells 2. EC subclustering identified five distinct subpopulations increasing from initiation to expansion stages, possibly reflecting progressive coronary vessel growth and remodeling (Fig. S7A,B). We also subclustered EPDCs and mural cells 2 and identified six EPDC subtypes and two mural cell 2 subtypes (Fig. S7C-F).
Single-cell transcriptome atlas during coronary development. (A-C) Combined uniform manifold approximation and projection (UMAP) plot (A) and split UMAP plots (C) visualizing nine cell types identified across cardiac development and dot plot showing average marker gene expression and abundance corresponding to each cell type (B). (D-F) Combined UMAP plot (D) and split UMAP plots (F) of coronary ECs visualizing four subtypes identified across cardiac development and dot plot showing average expression and abundance of selected marker genes corresponding to each subtype (E). (G) Dot plot showing average expression and abundance of top seven marker genes in coronary ECs. (H,I) Dot plots showing average expression and abundance of selected highly expressed marker genes of coronary ECs in all cell types (H) and in coronary EC subtypes (I). (J-M) In situ HCR for tppp3 (J), ano7 (K), gig2j (L), gjb10 (M) coupled with immunostaining for EGFP (green) and RFP (yellow) on whole-mount TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) ventricles. Cyan arrowheads point to EGFP+/RFPlow or EGFP+/RFP− showing HCR signal. AVC, atrioventricular canal; V, ventricle. (N,O) In situ HCR for gig2j (N), gjb10 (O) coupled with immunostaining for EGFP (green) and RFP (yellow) on whole-mount TgBAC(sele:EGFP); Tg(−0.8flt1:RFP) ventricles. Cyan arrowheads point to EGFP+/RFPlow or EGFP+/RFP− showing HCR signal. Dotted boxes indicate the regions shown at higher magnification to the right. T/NK/B, T cells/natural killer cells/B cells. Scale bars: 100 μm.
Marker gene expression analyses of the coronary EC subclustering identified arterial (unc5b and efnb2) (Adams et al., 1999; Gale et al., 2001; Navankasattusas et al., 2008), capillary (fabp4a) (van der Ark-Vonk et al., 2024), venous (ephb4 and sele) (Adams et al., 1999; Sun et al., 2015; Wang et al., 1998) and lymphatic (lyve1b and flt4) (Jackson, 2004; Jerafi-Vider et al., 2021) ECs (Fig. 3D,E). These cell types increased from the initiation to the maturation stages (Fig. 3F) and displayed distinguishable gene expression signatures (Fig. 3G).
In addition, these analyses identified tppp3, hpn, ano7, gig2j and gjb10 as new markers showing high and restricted expression in coronary ECs (Fig. 3H). Specifically, tppp3, hpn and ano7 were enriched in capillaries, while gig2j and gjb10 were mainly expressed in coronary veins (Fig. 3I). Using in situ hybridization chain reaction (HCR) coupled with immunostaining on whole-mount ventricles, we observed that tppp3 and ano7 were expressed mainly in capillary-like vessels and some large veins (etv2:EGFP*^+^/−0.8flt1*:RFP^−^) (Fig. 3J,K). gig2j and gjb10 were expressed in main dorsal veins (Fig. 3L-O). hpn levels were not clearly detectable by HCR.
Manipulation of Vegfa signaling impairs coronary network formation
Vegfa is a key regulator of angiogenesis and its depletion leads to severe vascular and developmental defects as well as embryonic lethality across vertebrate species (Carmeliet et al., 1999; Hudlicka et al., 1992; Rossi et al., 2016; Taimeh et al., 2013), including zebrafish (Rossi et al., 2016). We have previously shown that vegfaa mutants can be rescued to adulthood, providing a unique model in which to study the role of Vegfaa in coronary development (Marín-Juez et al., 2016). At 14 mm (establishment stage), rescued vegfaa^−/−^ exhibited defective LCA and main dorsal vein growth compared to wild-type (WT) fish (Fig. 4A-C). At 18 mm (expansion I), dorsal and ventral vessel expansion was severely reduced in rescued vegfaa^−/−^ (Fig. 4D-H). In addition, we noticed that LCA localization was altered in several vegfaa^−/−^ ventricles, with the LCA growing along the right ventricular curvature (Fig. 4F,I). At 24 mm (expansion II), the vegfaa^−/−^ coronary network expanded and reached coverage levels similar to those in WTs (Fig. 4J-M). While at this stage the overall ventricular coverage was not significantly different, rescued vegfaa^−/−^ ventricles exhibited a greater degree of variability (Fig. 4J,N). To further analyze these differences, we evaluated the coronary network distribution in WT and mutant ventricles. To assess their spatial distribution, we divided the ventral ventricular surface into four quadrants and measured vascular coverage in each of these areas. While WT fish displayed relatively uniform coronary network distribution among the four quadrants, rescued vegfaa^−/−^ clustered coronaries in area I and displayed sparse coverage in area IV (Fig. 4J,N). At 28 mm (maturation), rescued vegfaa^−/−^ exhibited incomplete and reduced coronary coverage, characterized by thinner vessels and a disorganized vascular network compared to WT siblings (Fig. 4O-R), consistent with previous findings (Marín-Juez et al., 2016). Vegfa regulates endothelial development and specification (Carmeliet et al., 1996; Jin et al., 2017). To analyze possible arterial phenotypes, we analyzed vegfaa^−/−^ Tg(−0.8flt1:RFP) ventricles. vegfaa^−/−^ ventricles from establishment to maturation showed reduced and sparse RFP expression, suggesting either defects in arterialization or dysregulation of transgene expression (Fig. S8A-D).
*Loss of Vegfa signaling impairs coronary network formation. (A) Whole-mount TgBAC(etv2:EGFP) ventricles from 14-mm-long WT and rescued vegfaa−/−. Magenta arrowheads point to the LCA, blue arrowheads point to main dorsal coronary veins, white arrowheads point to defective LCA, and yellow asterisk indicates absence of main dorsal coronary veins. (B,C) LCA length (B) and percentage of ventricles with main dorsal coronary veins (C) in WT and rescued vegfaa−/− 14-mm-long zebrafish. (D-F) Whole-mount TgBAC(etv2:EGFP) ventricles from 18-mm-long WT (D) and vegfaa−/− (E,F). Magenta arrows indicate the direction and position of LCA growth. Magenta arrowheads point to coronary plexus branching points. Blue arrowheads point to main dorsal coronary veins. White arrowhead points to defective ventral vasculature. Yellow asterisks indicate absence of main dorsal coronary veins. (G-I) Percentage of the ventral (G) and dorsal (H) vessel coverage, and LCA position (I). (J) Whole-mount TgBAC(etv2:EGFP) ventricles from 24-mm-long WT and rescued vegfaa−/−. Cyan dotted lines outline the nonvascularized area at the ventral ventricular base. Blue arrowheads point to main dorsal coronary veins. Yellow asterisk indicates absence of large dorsal coronary veins. White dashed lines divide the ventricular surface into four areas. (K-N) Percentage of the ventral (K) and dorsal (L) vessel coverage, ventral ventricular base nonvascularized area (M) and proportion distribution of ventral vascular coverage (N) in 24-mm-long WT and rescued vegfaa−/−. (O) Whole-mount TgBAC(etv2:EGFP) ventricles from 28-mm-long WT and rescued vegfaa−/−. Blue arrowheads point to main dorsal coronary veins. Yellow asterisk indicates absence of large dorsal coronary veins. (P-R) Percentage of the ventral (P) and dorsal (Q) vessel coverage, and average ventral vessel diameter (R). (S) Whole-mount TgBAC(etv2:EGFP) ventricles of WT and rescued vegfaa−/− ventricles from 15-mm-long zebrafish stained for EGFP (coronary ECs, green), EdU (proliferating cells, cyan) and DNA (DAPI, red). White arrowheads point to EdU+/EGFP+ cells in the LCA. xz axis orthogonal views of WT (I) and rescued vegfaa−/− (II) ventricles and yz axis orthogonal views of WT (III) and rescued vegfaa−/− (IV) ventricles are shown. Schematic summarizes the experimental protocol. BL, body length; IP, intraperitoneal. (T) Percentage of EdU+/EGFP+ cells in the LCA in 15-mm-long WT and rescued vegfaa−/−. (U-W) Whole-mount Tg(fli1a:EGFP) ventricles from scube2+/+ (U) and scube2−/− (V,W) 13-mm-long zebrafish. Magenta arrows indicate the direction and position of LCA growth. Blue arrowheads point to main dorsal coronary veins, and yellow asterisks indicate absence of main dorsal coronary veins. (X,Y) LCA length (X) and percentage position (Y) in 13-mm-long scube2+/+ and scube2−/−. Data in graphs are expressed as mean±s.e.m. ns, no significant difference. *P<0.05, *P<0.01 (two-tailed, unpaired Student's t-test). AVC, atrioventricular canal; BA, bulbus arteriosus; V, ventricle. Scale bars: 100 μm.
Since we observed a severe reduction in LCA growth in vegfaa^−/−^ ventricles, we hypothesized that defective proliferation might be the underlying cause for this defect. To test this possibility, we performed EdU incorporation assays coupled with immunostaining in whole-mount TgBAC(etv2:EGFP) ventricles from 15 mm fish. We found a significant reduction in the percentage of EdU^+^ coronary ECs within the LCA of vegfaa^−/−^ compared to WT (Fig. 4S,T). Moreover, our data indicate LCA mislocalization in some vegfaa^−/−^ ventricles (Fig. 4F,I). This phenotype could be due to alterations in developmental patterning caused by defective Vegfa signaling. To test this possibility, we analyzed LCA development in scube2^−/−^ fish. Scube2 modulates Vegfa signaling to induce angiogenesis (Ali, 2020; Lin et al., 2017) and functions upstream of Sonic Hedgehog (Shh) signaling during developmental patterning (Kawakami et al., 2005; Woods and Talbot, 2005). At 13 mm, scube2^−/−^ exhibited defective LCA and dorsal coronary vein growth, recapitulating vegfaa^−/−^ phenotypes (Fig. 4U-X). Furthermore, 50% of scube2^−/−^ display mislocalized LCA positioning, like vegfaa^−/−^ ventricles (Fig. 4W,Y). Together, these data indicate that Vegfa signaling regulates LCA growth and localization.
As retinoic acid (RA) signaling regulates endothelial proliferation via Vegf and Shh pathways (Bohnsack et al., 2004; Lai et al., 2003), we next examined the expression of RA signaling-related genes in our scRNA-seq datasets. We found that several RA pathway components, including aldh1a2, raraa, rarab, rxraa and rxrab were highly expressed in ECs, coronary ECs, EPDCs, mesenchymal cells, mural cells and immune cell populations mainly during expansion stage. The expression of genes encoding RA-degrading enzymes (cyp26a1 and cyp26c1) was not detectable (Fig. S9).
Recent studies have shown that VEGF can either promote or inhibit angiogenesis in a dose-dependent manner (Pontes-Quero et al., 2019). Similarly, stimulation of Vegfa signaling through flt1 deletion enhances heart regeneration while strong Vegfaa overexpression impairs it (Karra et al., 2018; Wang et al., 2024). To assess how different levels of Vegfa signaling induction regulate coronary development, we analyzed flt1 mutants and the Tg(myl7:CreER); Tg(βactin2:loxP-mTagBFP-STOP-loxP-vegfaa) overexpression line [hereafter referred to as Tg(myl7:CreER); Tg(βact2:BS-vegfaa)] at the different stages.
First, we analyzed flt1^−/−^ at the expansion I stage (18 mm). At this stage, there was no difference in coronary coverage between flt1^−/−^ and WT siblings (Fig. 5A-C). However, the directional expansion of the coronary network was altered. In flt1^−/−^ fish, the coronary network predominantly expanded left to right, contrasting with the right-to-left pattern observed in WTs (Fig. 5A,D). At the expansion II stage (24 mm), vessel coverage remained similar in flt1^−/−^ and WT siblings (Fig. 5E-G), yet the spatial distribution of the coronary network in flt1^−/−^ fish was irregular. Specifically, areas I and III of the ventral ventricular surface appeared hypervascularized in flt1^−/−^ (Fig. 5E,I), resulting in a significant reduction of the ventral ventricular base nonvascularized area (Fig. 5E,H). At the maturation stage (28 mm), no differences in total coronary coverage were observed between flt1^−/−^ and WT siblings (Fig. S10A-D). Tg(−0.8flt1:RFP) expression was strongly upregulated in flt1^−/−^ coronary ECs (Fig. S8E-G).
*Coronary network development in conditions of increased Vegfaa signaling. (A) Whole-mount TgBAC(etv2:EGFP) ventricles from 18-mm-long flt1+/+ and flt1−/−. White arrows from R (right) and L (left) represent the direction of coronary network expansion. (B-D) Percentage of the ventral (B) and dorsal (C) vessel coverage, and direction of coronary network expansion (D) in 18-mm-long flt1+/+ and flt1−/−. (E) Whole-mount TgBAC(etv2:EGFP) ventricles from 24-mm-long flt1+/+ and flt1−/−. Cyan dotted lines outline the nonvascularized area at the ventral ventricular base. White dashed lines divide the ventricular surface into four areas. (F-I) Percentage of the ventral (F) and dorsal (G) vessel coverage, ventral ventricular base nonvascularized area (H) and proportion distribution of ventral vascular coverage (I) in 24-mm-long flt1+/+ and flt1−/−. (J) Whole-mount Tg(myl7:CreER); Tg(βact2:BS-vegfaa); TgBAC(etv2:EGFP) ventricles from 18-mm-long control (EtOH, ethanol) and tamoxifen (4-OHT) treated (vegfaaOE) fish. Dotted boxes indicate the regions shown at higher magnification in insets. White arrowheads point to main dorsal coronary veins. Schematic summarizes the experimental protocol. BL, body length. (K-M) Percentage of the ventral (K) and dorsal (L) vessel coverage, and diameter of the largest main dorsal coronary vein (M) in 18-mm-long control and 4-OHT-treated zebrafish. Data in graphs are expressed as mean±s.e.m. ns, no significant difference. *P<0.01 (two-tailed, unpaired Student's t-test). AVC, atrioventricular canal; V, ventricle. Scale bars: 100 μm.
Next, to further test the effect of strong Vegfa signaling stimulation, we overexpressed vegfaa using the Tg(myl7:CreER); Tg(βact2:BS-vegfaa) line crossed with the TgBAC(etv2:EGFP); Tg(−0.8flt1:RFP) background. Following tamoxifen administration, treated ventricles displayed pronounced hypervascularization and distorted vascular architecture compared to control ventricles (Fig. 5J-L). Moreover, vegfaa overexpression led to a threefold dilation of the main dorsal coronary veins (Fig. 5J,M). As no distinctive LCA could be observed after vegfaa overexpression, we analyzed Tg(−0.8flt1:RFP) ventricles. As expected, based on observations in flt1^−/−^ ventricles, RFP expression was strongly upregulated in all coronary vessels, preventing us from assessing arterial identity (Fig. S8H). Interestingly, −0.8flt1:RFP was ectopically expressed in endocardial cells in recombined Tg(myl7:CreER); Tg(βact2:BS-vegfaa) ventricles, likely reflecting the strong induction of Vegfaa in these fish (Fig. S8H). These data further support that the changes in −0.8flt1:RFP expression observed upon Vegfa manipulation result, at least in part, from altered vegfaa expression levels, rather than reflecting changes in specification.
Lastly, we examined Vegf receptor expression across distinct coronary EC subtypes in our scRNA-seq datasets. flt1 and kdrl were expressed in all coronary ECs with lower expression in lymphatic ECs, kdr was expressed in all clusters and flt4 expression was largely restricted to lymphatic ECs (Fig. S11).
Collectively, our results show that Vegfa signaling does not simply control coronary vessel growth, but also dictates specific aspects of network patterning, including LCA positioning, directional expansion across the ventricle, and regional distribution of vessel density.
Spatiotemporal dynamics of coronary vessel–CM interactions during heart development
Previous studies have shown that coronary vessels act as scaffolds for growing CMs during development and regeneration in zebrafish (Marín-Juez et al., 2019). To profile this interaction, we used double-transgenic Tg(−0.8flt1:RFP); Tg(gata4:GFP) fish to label coronary ECs and growing cortical CMs, respectively (Gupta and Poss, 2012; Kikuchi et al., 2010; Marín-Juez et al., 2019). At 14 mm, we observed that gata4:GFP^+^ CMs emerged along the ventricular base, the left ventricular curvature and near the AVC region (Fig. 6A), consistent with previous reports (Gupta et al., 2013). Exclusively at this stage, we observed gata4:GFP^+^ cells with epicardial morphology sparsely distributed over the ventricle (Fig. 6A). We performed Caveolin-1 (Cav-1) staining, an epicardial marker, on Tg(gata4:GFP) ventricular sections and found that these cells were Cav-1^+^, indicating epicardial identity (Fig. S12A). At 18 mm, gata4:GFP^+^ cortical CMs populated the ventral surface, paralleling the coronary network expansion and showing an intimate association with the developing vasculature (Fig. 6B,B′). To assess the spatial relationship between coronary vessels and CM growth, we quantified ventral CM coverage relative to coronary vessel proximity, grouping distances into 0-15 μm, 15-30 μm and >30 μm. The majority of growing CMs were located within 0-15 μm of developing coronaries (Fig. 6C), suggesting close spatial coordination.
Coronary vessel–CM interactions during development. (A) Whole-mount Tg(−0.8flt1:RFP); Tg(gata4:GFP) ventricle from a 14-mm-long zebrafish. Red arrowhead points to the LCA. Green arrowheads point to GFP+ cortical CMs. Inset shows magnified view of the boxed region. Cyan arrowheads point to morphologically distinct GFP+ cells. (B,B′) Whole-mount a Tg(−0.8flt1:RFP); Tg(gata4:GFP) ventricle from an 18-mm-long zebrafish showing expanding GFP+ cortical CMs in close proximity to developing coronary vessels (B) and a 3D reconstruction of the ventral ventricular surface (B′). High-magnification images of the ventral 3D reconstruction are shown (i,ii). White dashed arrows indicate CM growth expansion toward the vascular front. (C) Ventral ventricular cortical coverage area of gata4:GFP+ CMs located at 0-15, 15-30 and >30 µm from the nearest RFP+ coronary vessel. Example images are shown for each of the categories with the relevant distance marked by white bars (i-iii). (D) Whole-mount Tg(−0.8flt1:RFP); Tg(gata4:GFP) ventricle from an 18-mm-long zebrafish stained for RFP (coronary ECs, yellow), GFP (growing cortical CMs, cyan) and cardiac troponin T (cTnT, sarcomeres, magenta). Green arrowheads point to gata4:GFPlow CMs displaying well-organized sarcomeres. Yellow asterisk marks gata4:GFP+ CMs at the expanding vascular front lacking defined sarcomeres. (E) Whole-mount Tg(−0.8flt1:RFP); Tg(gata4:GFP) zebrafish ventricle stained for RFP (coronary ECs, yellow), GFP (growing cortical CMs, cyan) and EdU (proliferating cells, magenta). xz and yz orthogonal views show proliferating coronary ECs (i; EdU+/RFP+, white arrowheads), proliferating CMs in close proximity to coronaries (ii; EdU+/GFP+, yellow arrowheads), proliferating CMs near an expanding vascular front (iii; EdU+/GFP+, red arrowheads) and GFP− proliferating cells in the trailing region behind the expanding vascular front (iv; EdU+/GFP−, blue arrowheads). Schematic summarizes the experimental protocol. BL, body length. (F,G) Whole-mount Tg(−0.8flt1:RFP); Tg(gata4:GFP) ventricles from 25-mm (F) and 28-mm (G) -long zebrafish. Dotted lines outline the nonvascularized area (cyan) and lack of cortical CM (gata4:GFP+) coverage (red) at ventral ventricular base. Dashed boxes indicate regions shown at higher magnification to the right. AVC, atrioventricular canal; BA, bulbus arteriosus; V, ventricle. Scale bars: 100 μm.
Next, we performed whole-mount cardiac troponin T (cTnT) staining in Tg(−0.8flt1:RFP); Tg(gata4:GFP) ventricles to analyze sarcomere arrangement. CMs trailing behind the expanding front displayed defined sarcomeres arranged perpendicularly to near coronaries, indicating CM orientation and structural maturation. In contrast, CMs at the expanding front exhibited poorly defined sarcomeres (Fig. 6D). To test whether the spatial proximity between CMs and coronary ECs reflects coordinated growth dynamics, we performed EdU incorporation assays combined with immunostaining. We observed EdU^+^ cells in both expanding coronary vessels and adjacent CMs, indicating synchronized proliferation (Fig. 6. Ei). EdU^+^ CMs were consistently localized near developing coronary vessels (Fig. 6. Eii). Moreover, EdU^+^ CMs were enriched at the expanding front (Fig. 6. Eiii), whereas most EdU^+^ cells behind were gata4:GFP^−^ (Fig. 6. Eiv). These findings suggest that coronary vessels and neighboring CMs undergo coordinated proliferation specifically at the myo-vascular expanding front.
At the expansion II stage (25 mm), gata4:GFP^+^ CMs continued to expand over the ventricular surface (Fig. 6F). Notably, the ventral ventricular base, the last area to be vascularized (Fig. 1R-U), was also the last area to be populated by gata4:GFP^+^ CMs, further indicating coronary-CM coordination (Fig. 6F). At the maturation stage (28 mm), the entire ventricular surface was populated by both cortical CMs and coronary vessels (Fig. 6G). Overall, these data show that the developing coronary network functions as a vascular scaffold for cortical CMs, with the spatial expansion of these CMs mirroring that of developing coronaries (Fig. S13A).
To better understand this interaction, we first performed secondary CM subclustering of our scRNA-seq datasets and identified seven CM subtypes. Primordial CMs (pCMs) were the least abundant and characterized by high expression of acta2, hey2, actn1 and smoc1 (Carey et al., 2024; Tsedeke et al., 2021) (Fig. 7A,B). Trabecular CMs (tCMs) with three subclusters (tCMs-1, tCMs-2 and tCMs-3) were the most abundant subtype and displayed strong expression of the canonical myocardial markers myl7 and actc1a, which are associated with contractile differentiated CMs (Carey et al., 2024) (Fig. 7A,B). Cortical CMs (cCMs) with two subclusters (cCMs-1 and cCMs-2) expressed lower levels of myl7 and actc1a, and lacked expression of tbx5a, as previously described (Carey et al., 2024; Sánchez-Iranzo et al., 2018) (Fig. 7A,B). We also identified a CM population that co-expressed markers of both primordial (actn1, smoc1) and trabecular (tbx5a, gata6, mef2d) CMs. Given this hybrid expression profile, we designated this cluster as primordial-trabecular CMs (p-tCMs), suggesting a transitional or intermediate CM state (Fig. 7A,B). Across developmental stages, we observed dynamic shifts in CM subtype abundance. Both pCMs and p-tCMs increased from initiation to expansion stages then declined at maturation. tCMs followed a similar trend. In contrast, cCMs expanded progressively from initiation to maturation (Fig. 7C), consistent with cortical wall formation (Gupta and Poss, 2012).
Coronary vessel–CM molecular interactions during cardiac development. (A-C) Combined UMAP plot (A) and split UMAP plots (C) of CMs visualizing seven subtypes identified across cardiac development, and dot plot showing average expression and abundance of selected marker genes corresponding to each subtype (B). (D) Circle plots showing differential cell–cell communication networks between coronary ECs and CM subtypes across cardiac development. Each node in the circle plot represents a cell cluster, with the node size being proportional to cell number. Edges connect nodes, indicating communication between cell clusters. Edge width represents communication strength. (E-H) Potential ligand–receptor interactions within coronary ECs, and between coronary ECs with primordial CMs (pCMs) and primordial-trabecular CMs (p-tCMs) across developmental stages of initiation (E), establishment (F), expansion (G) and maturation (H). nectin3b-pvrl2l/pvrl2l-pvrl2l and hbegfa-erbb4b interactions are highlighted in blue and orange text, respectively.
Next, we performed cell–cell communication analyses between coronary ECs and the seven CM clusters (Figs S6D and S14A,B). Coronary ECs appeared to mainly interact with pCMs and p-tCMs. The interaction strength between these cell types increased gradually from initiation to establishment then decreased at later stages (Fig. 7D). We also found interactions among pCMs, p-tCMs and cCMs-1 throughout development (Fig. 7D). Other cardiac components, including EPDCs, also appeared to interact with coronary ECs and CMs. Likewise, macrophage interactions with coronary ECs and CMs were observed throughout development (Fig. S14C).
As we found that coronary ECs mainly interact with pCMs and p-tCMs, we performed ligand–receptor analysis between these cell types. Within coronary ECs, we observed higher number of ligand–receptor pairs at expansion and maturation stages (Fig. 7G,H). Coronary EC-pCM ligand–receptor pairs decreased gradually from initiation to maturation stages, while coronary EC-p-tCM ligand–receptor pairs increased from initiation to establishment stages (Fig. 7E-H). Notably, the hbegfa–erbb4b interaction was predicted to be strong between coronary ECs, and pCMs and p-tCMs, from initiation to establishment stages (Fig. 7E-H). EC-secreted HB-EGF promotes CM proliferation, survival and maturation (Iwamoto and Mekada, 2006; Iwamoto et al., 2003). In addition, the nectin3b–pvrl2l heterophilic interaction was predicted exclusively during initiation stage, between coronary ECs and pCMs (Fig. 7E), whereas the pvrl2l–pvrl2l homophilic interaction observed in coronary ECs and coronary ECs-pCMs peaked at the establishment stage (Fig. 7F-H). Pvrl2l (nectin-2-like) mediates Ca^2+^-independent cell–cell adhesion, suggesting a role in supporting physical integration and alignment of coronary ECs and CMs during vessel expansion (Kinugasa et al., 2012; Takai et al., 2003). To assess whether subtype-specific interactions correlate with proliferation, we examined cell cycle-related gene expression. Proliferation markers including mki67, pcna, aurka and mcm5 were enriched from initiation to expansion stages, particularly in pCMs and p-tCMs (Fig. S15).
Altogether, these data identify complex signaling interactions between coronary ECs and CMs during cardiac development.
Impaired coronary vessel formation via Vegfaa manipulation disrupts CM expansion
Next, we used our genetic models for Vegfa signaling manipulation to test how alterations in coronary development affect CM growth. We focused on the expansion stage, characterized by strong coronary-cortical CM interplay and active growth. At this stage, vegfaa^−/−^ fish did not follow the typical right-to-left CM expansion, displaying instead a more symmetric growth pattern (Fig. 8A,B). CM expansion was also altered in both flt1^−/−^ and Tg(myl7:CreER); Tg(βact2:BS-vegfaa) tamoxifen-treated (vegfaa^OE^) ventricles (Fig. 8C,D). While ventral CM coverage was unchanged between WT and vegfaa^−/−^ fish, it appeared to be increased in flt1^−/−^ ventricles (Fig. 8A-C,E), consistent with previous studies showing enhanced CM regeneration in these mutants (Wang et al., 2024). Interestingly, inducing higher vegfaa levels using the Tg(myl7:CreER); Tg(βact2:BS-vegfaa) line led to a significant reduction in total CM coverage (Fig. 8D,E). Next, we analyzed differences in CM distribution. In WT fish, CM coverage was predominant in areas I and II, mirroring the coronary expansion pattern (Fig. 8A,F). In contrast, vegfaa^−/−^ and flt1^−/−^ fish displayed reduced CM coverage in these areas with increased coverage of areas III and IV. vegfaa overexpression using the Tg(myl7:CreER); Tg(βact2:BS-vegfaa) line shifted CM expansion distribution toward areas I and III, indicating alterations in regional expansion (Fig. 8D,F). It is possible that vegfaa overexpression in Tg(myl7:CreER); Tg(βact2:BS-vegfaa) fish impairs the mitogenic potential of Vegfaa, leading to mislocalized and restricted CM expansion as reported during regeneration (Karra et al., 2018). These data suggest that precise coronary developmental patterning, orchestrated by Vegfa signaling, is essential for regulating CM growth and expansion to populate the ventricular surface.
*Manipulation of Vegfa signaling disrupts coronary–CM interactions. (A-D) Whole-mount Tg(−0.8flt1:RFP); Tg(gata4:GFP) ventricles from 18-mm-long WT (A), rescued vegfaa−/− (B), flt1−/− (C) and Tg(myl7:creER); Tg(βact2:BS-vegfaa) treated with 4-OHT (vegfaaOE) (D). (E,F) Percentage (E) and proportion distribution (F) of ventral gata4:GFP+ CM coverage area in 18-mm-long WT, rescued vegfaa−/−, flt1−/− and Tg(myl7:creER); Tg(βact2:BS-vegfaa) treated with 4-OHT (vegfaaOE). (G) Whole-mount Tg(gata4:GFP) ventricles from 18-mm-long WT and vegfaa−/− stained for GFP (growing cortical CMs, cyan) and EdU (proliferating cells, magenta). Yellow dashed lines divide the ventral ventricular surface into right (R) and left (L) sides. Red arrowheads point to EdU+/GFP+ cells. Dashed boxes indicate regions shown at higher magnification to the right. Orthogonal views of WT in the xz axis (i) and yz axis (ii) and vegfaa−/− in the xz axis (iii) and yz axis (iv) are shown. (H) Number of EdU+/GFP+ CMs on the right and left sides of the ventral ventricular surface in WT and rescued vegfaa−/− 18-mm-long zebrafish. Data in graphs are expressed as mean±s.e.m. ns, no significant difference. P<0.05 (two-tailed, Mann–Whitney U test). Schematic summarizes the experimental protocol. BL, body length; IP, intraperitoneal; AVC, atrioventricular canal; V, ventricle. Scale bars: 100 μm.
To further determine whether CM proliferation was affected under conditions of impaired coronary development, we performed EdU incorporation assays coupled with immunostaining in Tg(gata4:GFP) vegfaa^−/−^ mutants. In WT fish, EdU^+^ CMs were significantly enriched on the right side of the ventral ventricular surface. This spatial asymmetry was lost in vegfaa^−/−^ fish accompanied by a significant reduction in the total number of EdU*^+^* CMs (Fig. 8G,H).
Overall, these findings indicate that coronary vessel development is coupled with CM expansion, and that disruption of coronary growth through impaired Vegfa signaling compromises cortical myocardial expansion.
DISCUSSION
We assemble a developmental roadmap for coronary vasculature formation and show how coronary vessels play an instructive role in shaping the ventricular wall through cellular and molecular interactions. Our findings highlight both conserved and species-specific aspects of coronary vasculature development, provide new molecular tools for investigating coronary vessel identity and reveal how coronary vessel–CM interactions are modulated by angiogenic signaling levels.
This study reveals a dual-origin model of coronary vessel formation in zebrafish, identifying the emergence of coronary sprouts from both the BA and the AVC. Our data indicate that the earliest vessel arises from the BA, followed by a second vessel sprouting from the AVC. Previous studies have shown that the coronary vasculature initiates from the AVC endothelium in zebrafish (Harrison et al., 2015), while in giant danio (Devario malabaricus), the coronary vasculature has a similar dual origin (Shifatu et al., 2018). Also, more recently Mizukami et al. showed in zebrafish that the hypobranchial artery expands caudally to reach the BA at 36 dpf (Mizukami et al., 2023), further supporting our observations. Interestingly, this pattern parallels the coronary vessel emergence in the human embryonic heart, where the early coronary plexus forms around the peritruncal region of the outflow tract (Bogers et al., 1989; Silva-Junior et al., 2009; Tomanek, 2005).
Based on anatomical landmarks and quantitative mapping, we have defined four stages of coronary network development: initiation, establishment, expansion and maturation. The coronary network expands in a dorsal-to-ventral and right-to-left direction, suggesting that either internal cardiac architecture or local vascular cues guide this process. These patterns of coronary vessel expansion are consistent with those observed in mammals (Chen et al., 2014; Red-Horse et al., 2010).
Despite growing interest in coronary vessel biology, the venous portion of the coronary network has remained uncharacterized owing to the lack of specific markers (Gancz et al., 2019; Harrison et al., 2015, 2019; Marín-Juez et al., 2016). Here, we developed a TgBAC(sele:EGFP) line that is specifically expressed in coronary venous ECs. Using this tool, we monitored coronary vein development and observed that venous vessels emerge early and expand progressively following the dorsal-to-ventral and right-to-left expansion pattern typical of coronary growth. Notably, sele:EGFP expression was undetectable in −0.8flt1:RFP^high^ arterial vessels, allowing for the first time venous and arterial coronary vessels to be distinguished in zebrafish.
Further scRNA-seq analyses identified arterial, venous, capillary and lymphatic ECs. The relative abundance of these endothelial populations changed across developmental stages, with all subtypes showing progressive expansion from the initiation to the maturation stages, paralleling our in vivo data. In addition to known markers, our dataset identified new subtype-specific marker genes. Specifically, tppp3, hpn and ano7 were enriched in capillaries and gig2j and gjb10 in coronary veins.
Our data highlight the central role of Vegfa signaling in orchestrating multiple aspects of coronary development. Loss of vegfaa impairs LCA formation, EC proliferation, and spatial patterning, resulting in hypovascularization and disorganized vessel architecture. Conversely, increased Vegfa signaling via flt1 deletion or vegfaa overexpression leads to regional hypervascularization, increased vessel density, and disrupted spatial organization. These phenotypes align with previous studies showing that Vegfa regulates angiogenesis in a dose-dependent manner, establishing vascular hierarchy and morphogenetic fidelity (Karra et al., 2018; Wang et al., 2024).
To further investigate how alterations in coronary network formation affect CM development, we analyzed CM growth in these Vegfa signaling gain- and loss-of function models. In vegfaa mutants, directional CM expansion was lost, and overall proliferation reduced. In flt1 mutants, we observed enhanced CM coverage, consistent with observations in cardiac regeneration (Wang et al., 2024). Interestingly, stronger vegfaa overexpression reduced CM coverage, suggesting that excessive Vegfaa decouples endothelial-CM developmental programs, as previously proposed during heart regeneration (Wang et al., 2024). It is worth noting that the coronary vasculature in these animals was strongly perturbed, raising the possibility that instructive angiocrine signals might also be impaired in these vessels. These data underscore the need for precisely tuned angiogenic signaling to maintain the coordinated growth of endothelial and myocardial tissues.
Finally, our scRNA-seq analyses identified stage-specific shifts in CM populations, including primordial, trabecular and cortical subtypes, recapitulating previous lineage-tracing analyses reporting late emergence of trabecular-derived cortical CMs (Gupta and Poss, 2012). We also observed a marked increase in cortical CMs throughout development, mirroring that of the coronary plexus, further supporting the coupling between cortical CM growth and coronary network expansion. Interestingly, the pCM and p-tCM subtypes showed the strongest interactions with coronary ECs, suggesting that developing coronaries might signal to pCMs and p-tCMs to support cortical CM expansion. In addition, ligand–receptor interaction analyses revealed signaling axes, such as hbegfa–erbb4b (paracrine cues) and pvrl2l–pvrl2l (cell–cell adhesion), known to regulate myocardial proliferation and endothelial behavior (Iwamoto and Mekada, 2006; Iwamoto et al., 2003; Kinugasa et al., 2012; Takai et al., 2003). These data support a model in which coronary ECs act as both instructive and structural components, actively shaping the cardiac microenvironment through temporally and spatially regulated signaling programs. The extent to which the interaction between coronaries and CMs is direct or mediated through intermediate factors, or involves both mechanisms, warrants further investigation.
MATERIALS AND METHODS
Zebrafish lines and manipulations
All experimental procedures involving zebrafish (Danio rerio) were conducted in strict adherence to institutional and national animal welfare regulations. In this study, zebrafish were raised in system water with 20 fish per 3-l tank. We used the previously published lines TgBAC(etv2:EGFP)^ci1^ (Proulx et al., 2010), Tg(−0.8flt1:RFP)^hu5333^ (Bussmann et al., 2010), Tg(dll4:TagRFP)^sfc7^ (Marín-Juez et al., 2016), Tg(lyve1b:dsRed)^nz101^ (Okuda et al., 2012), Tg(fli1a:EGFP)^y1^ (Lawson and Weinstein, 2002), Tg(myl7:CreER)^pd10^ (Kikuchi et al., 2010), Tg(βactin2:loxP-mTagBFP-STOP-loxP-vegfaa)^pd262^ (Karra et al., 2018), Tg(gata4:GFP)^ae1^ (Heicklen-Klein and Evans, 2004), vegfaa^bns1^ (Rossi et al., 2016), flt1^bns29^ (Matsuoka et al., 2016) and scube2^as404^ (Tsao et al., 2022). To induce recombination in juvenile fish, Tg(myl7:CreER); Tg(βact2:BS-vegfaa) fish were bathed in 5 µM tamoxifen (4-OHT; Sigma-Aldrich, H7904) for 24 h in dark conditions as described previously (Karra et al., 2018) and put them back in system water after treatment.
Generation of the TgBAC(sele:EGFP) zebrafish line
To generate TgBAC(sele:EGFP) fish, the BAC clone DKEY-51E6 containing sele was modified by Red/ET recombineering technology (Gene Bridges) as previously described (Bussmann and Schulte-Merker, 2011; Suster et al., 2011).
Histological analyses
O-dianisidine staining was performed as described (Detrich et al., 1995). Ventricular section and whole-mount immunostaining were performed as previously described (Bakis et al., 2023; Marín-Juez et al., 2016). Primary antibodies used were: anti-GFP (Aves Labs, CGFP-1020; 1:500), anti-tRFP (Evrogen/Sapphire North America, AB233; 1:500), anti-Caveolin-1 (BD Transductions Laboratories, 610407; 1:100), anti-cTnT (Developmental Studies Hybridoma Bank, ct3; 1:100). Secondary antibodies used were Alexa Fluor 488 Goat Anti-Chicken IgG (H+L) (Thermo Fisher Scientific, A-11039; 1:750), Alexa Fluor 568 Goat Anti-Mouse IgG (H+L) (Thermo Fisher Scientific, A-11004; 1:750), Alexa Fluor 568 Goat Anti-Rabbit IgG (H+L) (Thermo Fisher Scientific, A-11036; 1:750) and Alexa Fluor 647 Goat Anti-Rabbit IgG (H+L) (Thermo Fisher Scientific, A-21244; 1:750). DAPI (4′,6-diamidino-2-phenylindole) was used as a nuclear counterstain. For HCR RNA-FISH (Molecular Instruments), the manufacturer's protocol for whole-mount zebrafish embryos and larvae was followed with modifications during sample preparation and fixation as previously described (Bakis et al., 2023). EdU staining was performed by using the Click-iT™ EdU Cell Proliferation Kit for Imaging with Alexa Fluor™ 555 dye (Invitrogen, C10338) following the manufacturer's instructions. EdU (2 µl, 500 µM) was injected intraperitoneally 1 day or 3 days before the extraction of the heart, depending on the experiment.
Imaging and quantification
Anesthetized fish were dissected, and ventricles were extracted and immediately put in freshly prepared 1× PBS (Sigma-Aldrich, P4417) for at least 5 min and rinsed three times followed by 4% paraformaldehyde treatment for 5 min. Ventricles were then mounted on a glass-bottom dish (ibidi, 80206) using 1.5% low melting agarose (Sigma-Aldrich, A4018). The ventricles were submerged in agarose without touching the bottom surface of the dish, and were later flipped to image both sides of ventricles. Imaging was performed on a Leica TCS SP8 laser scanning confocal microscope. O-dianisidine staining was imaged using a Nikon SMZ18 stereomicroscope. Quantifications were performed using ImageJ/Fiji from maximum projected confocal images. 3D reconstruction was performed using Imaris 10.2 (Oxford Instruments). Vascular coverage area quantifications were carried out using Tubeness (ImageJ/Fiji).
Single-cell dissociation
One pool of ventricles per developmental stage – initiation (10 mm) 25 ventricles, establishment (14 mm) 18 ventricles, expansion (18 mm) 12 ventricles and maturation (28 mm) 5 ventricles – was prepared using WT zebrafish. Euthanized fish were dissected, and ventricles were extracted in calcium-, magnesium-free HBSS (Sigma-Aldrich, H9394) supplemented with 20 U/ml heparin (Sigma-Aldrich, H3393). Ventricles were washed in fresh HBSS and transferred into 1 ml of 1 mg/ml of collagenase (Gibco, 17101015) dissolved in HBSS for approximately 30 min digestion on a thermal mixer at 32°C and 700 rpm. Ventricles were mechanically disrupted by pipetting every 10 min. Samples were centrifuged at 2000 rpm (∼376 g) for 5 min at room temperature (RT) and supernatants discarded. Cells were digested with 1 ml 1× TrypLE Express Enzyme (Gibco, 12605010) for 15 min on a thermal mixer at 32°C and 700 rpm and pipetted every 5 min. Enzymatic digestion was stopped by adding 1 ml of 20% fetal bovine serum (Thermo Fisher Scientific, A3160701) in HBSS to extracts. Tissue extracts were strained through a 70 μm cell strainer and centrifuged at 2000 rpm (∼376 g) for 5 min at RT. The supernatants were discarded, and pellets were washed with 1 ml of 20% fetal bovine serum in HBSS. After centrifugation at 2000 rpm (∼376 g) for 5 min at RT, cells were resuspended in 80 μl of 20% fetal bovine serum in HBSS and strained through a 40 μm cell strainer. Cells were counted and viability (>85%) was assessed using 0.4% Trypan Blue (Sigma-Aldrich, T8154). Cells were kept on ice and immediately processed for scRNA-seq.
scRNA-seq and analysis
For scRNA-seq, 14,000 cells were used for each sample and the Chromium Next GEM Single Cell 3′ Reagent Kit v3.1 (10x Genomics, PN-1000269) was used following the manufacturer's protocol. Each sample was indexed individually using the Dual Index Kit TT Set A (10x Genomics, PN-1000215). After quality control, libraries were run on a NovaSeq 6000 system (Illumina) at a depth of approximately 300 M reads/sample. FASTQ files were processed individually in 10x Genomics Cloud Analysis using the Cell Ranger Count v6.1.2 pipeline and reads were aligned to GRCz11 v4.3.2 reference genome (Lawson et al., 2020). Filtered gene expression matrices were imported and further processed in Seurat v4 (Hao et al., 2021). Samples were integrated to create a single aggregated Seurat object, and low-quality cells containing <200 unique features were removed. Batch effect correction was performed using the ‘data integration’ and ‘ScTransform’ functions in Seurat. To increase the retrieval of CMs characterized with high mitochondrial RNA content, an initial filtering was applied to remove cells with >40% mitochondrial RNA (Carey et al., 2024). After initial clustering and annotation of CMs, non-CM cells with >12% mitochondrial RNA were excluded from further analysis. The remaining cells were then re-clustered and annotated based on the expression of marker genes. Following normalization, data visualization was performed in Seurat, incorporating previously published R code with modifications (Carey et al., 2024). Quality control metrics are listed in Table S1. The number of cells retained at each filtering step is listed in Table S2. All marker genes in each cluster are listed in Table S3. The potential ligand–receptor interactions in each developmental stage are listed in Table S4 (initiation), Table S5 (establishment), Table S6 (expansion) and Table S7 (maturation).
Statistical analysis
Data were statistically analyzed and graphics were created in GraphPad Prism v.10. When two groups were compared, comparative statistics were performed using two-tailed, unpaired Student's t-test or Mann–Whitney U-test for non-parametric tests. Data were considered significant at P<0.05. The number of samples are mentioned in the graphs and/or represented by dots in the graphs. The P-values are indicated in the figure legends. Data in graphs are expressed as mean±s.e.m.
Supplementary Material
10.1242/develop.205065_sup1Supplementary information
Table S1. Summary of scRNA-seq QC Metrics
Table S2. Number of cells retained
Table S3. All markers at each cluster
Table S4. Initiation all interactions
Table S5. Establishment all interactions
Table S6. Expansion all interactions
Table S7. Maturation all interactions
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adams, R. H., Wilkinson, G. A., Weiss, C., Diella, F., Gale, N. W., Deutsch, U., Risau, W. and Klein, R. (1999). Roles of ephrin B ligands and Eph B receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis. Genes Dev. 13, 295-306. 10.1101/gad.13.3.2959990854 PMC 316426 · doi ↗ · pubmed ↗
- 2Ali, H. (2020). SCUBE 2, vascular endothelium, and vascular complications: a systematic review. Biomed. Pharmacother. 127, 110129. 10.1016/j.biopha.2020.11012932278240 · doi ↗ · pubmed ↗
- 3Bakis, I., Sun, Y., Abd Elmagid, L., Feng, X., Garibyan, M., Yip, J. K., Yu, F. Z., Chowdhary, S., Fernandez, G. E., Cao, J. et al. (2023). Methods for dynamic and whole volume imaging of the zebrafish heart. Dev. Biol. 504, 75-85. 10.1016/j.ydbio.2023.09.00237708968 PMC 10841891 · doi ↗ · pubmed ↗
- 4Bogers, A. J. J. C., Gittenberger-De Groot, A. C., Poelmann, R. E., Péault, B. M. and Huysmans, H. A. (1989). Development of the origin of the coronary arteries, a matter of ingrowth or outgrowth? Anat. Embryol. 180, 437-441. 10.1007/BF 003051182619086 · doi ↗ · pubmed ↗
- 5Bohnsack, B. L., Lai, L., Dolle, P. and Hirschi, K. K. (2004). Signaling hierarchy downstream of retinoic acid that independently regulates vascular remodeling and endothelial cell proliferation. Genes Dev. 18, 1345-1358. 10.1101/gad.118490415175265 PMC 420359 · doi ↗ · pubmed ↗
- 6Bussmann, J. and Schulte-Merker, S. (2011). Rapid BAC selection for tol 2-mediated transgenesis in zebrafish. Development 138, 4327-4332. 10.1242/dev.06808021865323 · doi ↗ · pubmed ↗
- 7Bussmann, J., Bos, F. L., Urasaki, A., Kawakami, K., Duckers, H. J. and Schulte-Merker, S. (2010). Arteries provide essential guidance cues for lymphatic endothelial cells in the zebrafish trunk. Development 137, 2653-2657. 10.1242/dev.04820720610484 · doi ↗ · pubmed ↗
- 8Bussmann, J., Wolfe, S. A. and Siekmann, A. F. (2011). Arterial-venous network formation during brain vascularization involves hemodynamic regulation of chemokine signaling. Development 138, 1717-1726. 10.1242/dev.05988121429983 PMC 3074448 · doi ↗ · pubmed ↗