Frost fighters: The discovery of a tripartite molecular module that confers cold stress tolerance in apple
Margot Raffeiner

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
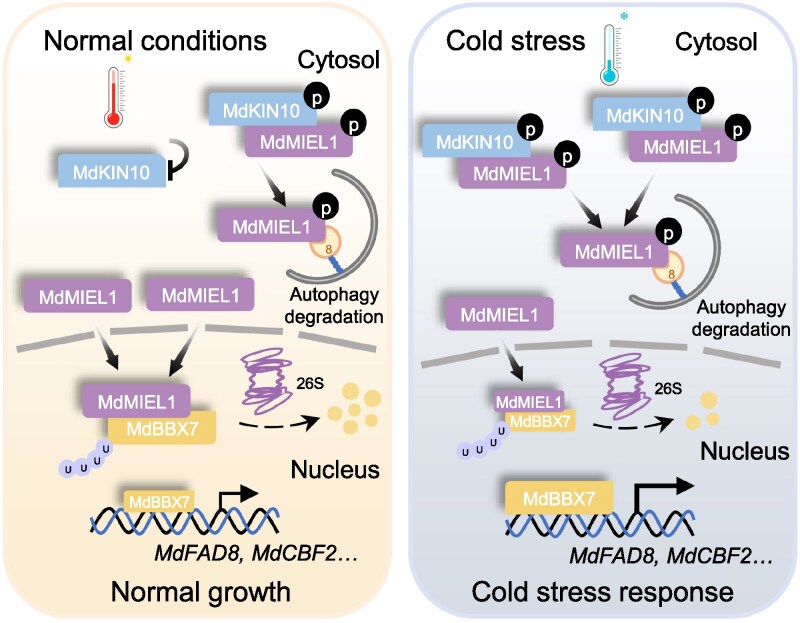 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Ion Channels and Receptors · Light effects on plants
When exposed to freezing temperatures (below 0 °C) or chilling conditions (0 to 15 °C), it gets a little chaotic inside the cell. For example, proteins that usually keep cell membranes flexible and functional begin to malfunction, leading to injury and, in crop species such as apple (Malus × domestica), severe yield losses (Unterberger et al. 2018). To restore balance, plants rely on protein degradation pathways that help remove damaged or misfolded proteins. Two major systems take the lead: proteasomal degradation, where a large protease complex directly cleaves proteins, and autophagy, in which cargos are engulfed by vesicles called autophagosomes and delivered to the vacuole for enzymatic breakdown (Raffeiner et al. 2023).
Because cold stress is a serious problem in apple cultivation, considerable research has been directed toward uncovering the molecular mechanisms of cold tolerance. Recently, a key discovery was the identification of a negative regulator called MYB30-Interacting E3 Ligase 1 (MdMIEL1), a RING-type E3 ligase whose stability must be carefully controlled to allow normal development and proper stress responses (An et al. 2020).
In new work, Fang Zhi and colleagues (Zhi et al. 2025) dissected how MdMIEL1 is regulated under cold stress. First, they discovered that MdMIEL1 interacts with MdKIN10, the catalytic α-subunit of the plant master regulator kinase Sucrose nonfermenting 1 (SNF1)-related protein kinase 1 (SnRK1) (Shen et al. 2009). Interestingly, this study further revealed that MdKIN10 activity itself is increased specifically under cold conditions, highlighting its role in freezing tolerance. Not only does MdKIN10 interact with MdMIEL1, but it also phosphorylates it, marking it for degradation (Fig. 1).
To shed more light on the role of this interaction during cold acclimatization, the researchers generated transgenic apple calli that overexpressed either the wild-type (WT) MdMIEL1 or a nonphosphorylatable mutant (MdMIEL1^S198A^) alone or alongside MdKIN10. Under cold treatment, MdMIEL1 protein abundance decreased, but this effect was amplified by coexpression with MdKIN10. At the same time, increased phosphorylation of WT MdMIEL1 in MdKIN10 overexpressing calli was observed. In contrast, MdMIEL1^S198A^ protein remained stable, showing that phosphorylation by MdKIN10 is essential for MdMIEL1 turnover.
The changes in protein abundance of MdMIEL1 that depend on its phosphorylation status under cold stress were also reflected phenotypically. Plants overexpressing MdMIEL1 were more sensitive to cold, showing lower survival rates as well as higher electrolyte leakage, reflecting greater cellular damage. However, plants expressing a constitutively phosphorylated version (MdMIEL1^S198D^) showed better growth under cold stress compared with those expressing the nonphosphorylated version, further underscoring the relevance of MdMIEL1's phosphorylation for cold stress adaptation. The importance of KIN10 was also confirmed using transgenic plants: those overexpressing MdKIN10 had higher survival rates after freezing, whereas loss of MdKIN10 impaired tolerance.
But how is MdMIEL1 degraded? Testing different chemical inhibitors that block either autophagy or proteasomal degradation, the authors demonstrated that under cold stress, MdMIEL1 is cleared primarily via the autophagy pathway. Supporting this, MdMIEL1 was shown to interact directly with the Autophagy-related protein 8i (ATG8i), a key autophagy protein responsible for delivering substrates to autophagosomes. This interaction depended on MdMIEL1's phosphorylation status, again linking KIN10 activity to MdMIEL1 degradation. As expected, overall autophagic activity was higher during cold stress, helping the plant to eliminate this negative regulator when temperatures drop.
Ultimately, the study further found the protein MdBBX7 to be degraded via the proteasome after being targeted and ubiquitinated by MdMIEL1. MdBBX7 enhances cold tolerance by increasing cold response gene expression and promoting fatty acid desaturation to increase membrane fluidity. Together, the authors uncovered a regulatory network where MdKIN10 phosphorylates MdMIEL1 that is removed from the cell via autophagic degradation, thereby allowing positive regulators of cold stress response such as MdBBX7 to be activated.
In summary, this work identified a critical molecular module that helps apple trees to boost their tolerance to freezing temperatures, knowledge that might be applied in the future to bring more resilient apple varieties to market.
Recent related articles in The Plant Cell:
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1An JP et al An apple MYB transcription factor regulates cold tolerance and anthocyanin accumulation and undergoes MIEL 1-mediated degradation. Plant Biotechnol J. 2020:18:337–353. 10.1111/pbi.13201.31250952 PMC 6953192 · doi ↗ · pubmed ↗
- 2Raffeiner M, Zhu S, González-Fuente M, Üstün S. Interplay between autophagy and proteasome during protein turnover. Trends Plant Sci. 2023:28:698–714. 10.1016/j.tplants.2023.01.013.36801193 · doi ↗ · pubmed ↗
- 3Shen W, Reyes MI, Hanley-Bowdoin L. Arabidopsis protein kinases GRIK 1 and GRIK 2 specifically activate Sn RK 1 by phosphorylating its activation loop. Plant Physiol. 2009:150:996–1005. 10.1104/pp.108.132787.19339507 PMC 2689985 · doi ↗ · pubmed ↗
- 4Unterberger C et al Spring frost risk for regional apple production under a warmer climate. P Lo S One. 2018:13:e 0200201. 10.1371/journal.pone.0200201.30044808 PMC 6059414 · doi ↗ · pubmed ↗
- 5Wang L et al Reduced proteasome degradation of Hsf B 2a at higher temperatures is responsible for the inhibition of anthocyanin synthesis in pear. Plant Cell. 2025:37:102. 10.1093/PLCELL/KOAF 102.40324390 · doi ↗ · pubmed ↗
- 6Wang X et al PUB 25 and PUB 26 dynamically modulate ICE 1 stability via differential ubiquitination during cold stress in Arabidopsis. Plant Cell. 2023:35:3585–3603. 10.1093/PLCELL/KOAD 159.37279565 PMC 10473228 · doi ↗ · pubmed ↗
- 7Zhang C et al Brassinosteroid-signaling kinase 4 activates mitogen-activated protein kinase 4 to enhance cold stress tolerance in maize. Plant Cell. 2025:37:koaf 234. 10.1093/PLCELL/KOAF 234.41026892 · doi ↗ · pubmed ↗
- 8Zhi F et al Md KIN 10-mediated phosphorylation of E 3 ubiquitin ligase Md MIEL 1 leads to its autophagic degradation under cold stress in apple. Plant Cell. 2025:koaf 284. 10.1093/plcell/koaf 284.41417625 · doi ↗ · pubmed ↗