Space Use and Movements During Egg Laying Associated With Nest Fate and Female Survival in Eastern Wild Turkeys
Paige E. Goodman, Nicholas W. Bakner, Nickolas A. Gulotta, Erin E. Ulrey, Bret A. Collier, Michael J. Chamberlain

TL;DR
Female wild turkeys that move more during egg laying have lower nest success but higher survival, while sharing space with other females improves nest success.
Contribution
This study links female movement and space use during egg laying to nest success and survival in wild turkeys using GPS data and Bayesian analysis.
Findings
Increased daily movement during laying decreases nest success by 1.73% per ~700 m traveled.
Greater overlap with conspecifics increases nest success by 4.76% per additional overlapping female.
Movement weakly correlates with increased female survival during incubation.
Abstract
Reproduction is a fundamental aspect of a species' life history that is energetically costly, yet critical for population sustainability and genetic diversity. Wild turkeys exhibit high rates of nest loss and female mortality during reproduction, prompting females to make decisions related to spatial and movement decisions during nesting. Using GPS data from eastern wild turkeys ( Meleagris gallopavo silvestris ), we assessed female movements and space use during laying and evaluated potential impacts of those metrics on nest success and female survival during incubation. We used a Bayesian logistic regression to estimate nest success and female survival based on space use, daily movements, and range overlap with conspecifics during the laying period. We found that with each increase of ~700 m in average daily distance traveled during laying, there was a 1.73% decrease in the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
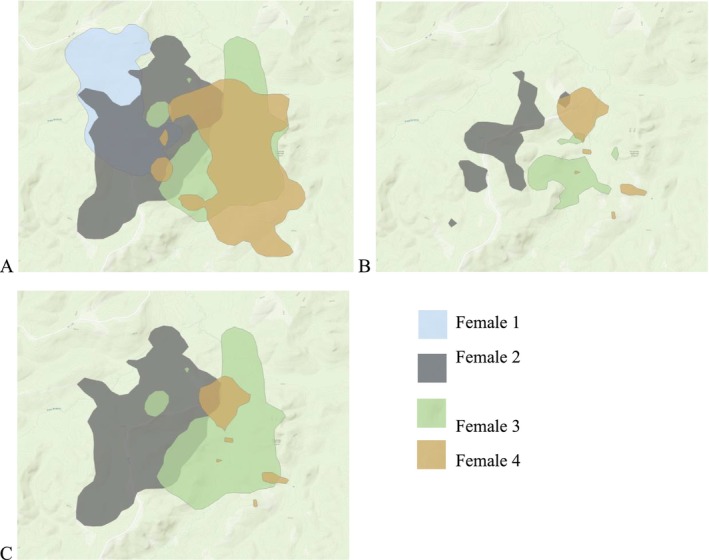 FIGURE 1
FIGURE 1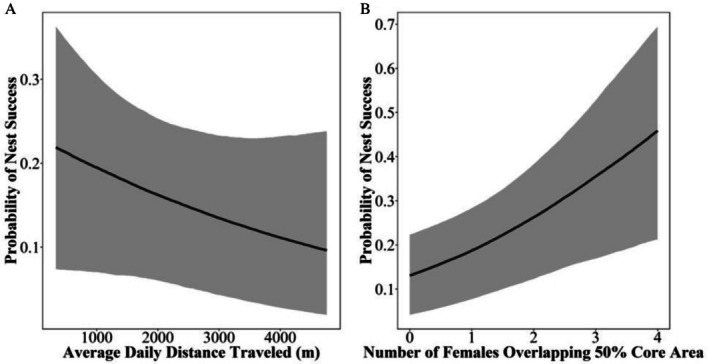 FIGURE 2
FIGURE 2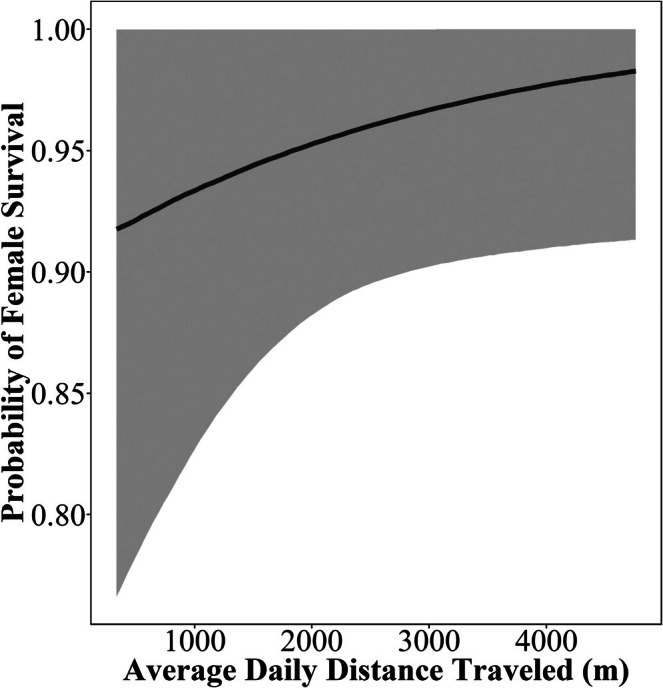 FIGURE 3
FIGURE 3| Covariate | Mean | SD |
|---|---|---|
| Average daily distance traveled (m) | 2151.14 | 699.93 |
| 50% Core area (ha) | 16.39 | 10.09 |
| 95% Home range (ha) | 89.67 | 59.32 |
| Mean number of females that overlapped core areas | 0.47 | 0.78 |
| Mean number of females that overlapped home ranges | 0.75 | 1.06 |
| Mean number of females with home range that overlapped core area | 0.60 | 0.91 |
| Covariate | Lower 95% CI | Estimate | Upper 95% CI | pd |
|---|---|---|---|---|
| Intercept | −2.81 | −1.69 | −1.11 | 100 |
| Average daily distance traveled | −0.43 | −0.15 | 0.09 | 89.47 |
| Number of females 50% overlapping | 0.11 | 0.34 | 0.65 | 99.84 |
| Nesting Attempt | ||||
| Nest Attempt 2 | −0.30 | 0.36 | 1.40 | 83.40 |
| Nest Attempt 3 | −1.11 | 0.33 | 2.18 | 65.67 |
| Covariate | Lower 95% CI | Estimate | Upper 95% CI | pd |
|---|---|---|---|---|
| Intercept | 2.06 | 3.06 | 9.82 | 100.00 |
| Average daily distance traveled | −0.13 | 0.25 | 0.70 | 92.39 |
| Number of females 50% overlapping | −0.16 | 0.17 | 0.75 | 84.33 |
| Nesting Attempt | ||||
| Nest Attempt 2 | −1.68 | 0.38 | 1.24 | 73.90 |
| Nest Attempt 3 | −4.23 | −0.37 | 1.22 | 66.84 |
- —Georgia Department Of Natural Resources10.13039/100016261
- —South Carolina Department of Natural Resources10.13039/100010242
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAvian ecology and behavior · Animal Behavior and Reproduction · Bird parasitology and diseases
Introduction
1
Reproduction is a fundamental aspect of a species' life history that is energetically costly, yet critical for population sustainability and genetic diversity (Avise 1996). Resource allocation theory governs how females allocate resources between producing offspring and individual growth and maintenance. Resource allocation posits that resources allocated to one aspect of an organism's life, such as survival, cannot simultaneously be allocated to another, such as reproduction. Consequently, females must balance energetic demands of producing offspring with resource acquisition and evading predation. Among avian taxa, the mortality rates of females during the nesting period play a significant role in the choice of reproductive tactics (Fontaine and Martin 2006), leading to a gradient of reproductive tactics where individuals prioritize productivity and survival differently (Jones 1989). The initiation of egg laying and incubation represents critical phases for avian species, demanding spatial confinement and considerable energy expenditure (Deeming and Reynolds 2015). For uniparental incubators, females must balance remaining at the nest site and making recess movements away from the nest location to secure resources (Williams 1996).
During incubation, uniparental ground nesters exhibit behaviors similar to those of central place foragers (Orians and Pearson 1979). When individuals are tethered to a centralized location like a nest, there may be space use restrictions and availability of resources may be limited (Kacelnik 1984; Van Gils and Tijsen 2007; Lalla et al. 2022). In species with relatively long incubation periods, individuals may be confined for extended temporal periods, requiring them to prioritize resource acquisition and minimize predation risk. To minimize predation risk during incubation, ground nesters often choose nest locations in dense vegetation or use behavioral tactics to avoid or hide from predators (Wiebe and Martin 1997; Smith et al. 2012). Individuals may familiarize themselves with certain areas to identify the less risky choices to lay their nest, and balance finding places with foraging resources while avoiding drawing attention from predators (Olsson et al. 2008). Likewise, individuals may also be drawn to similar areas on the landscape due to shared preferences for particular vegetation types. However, individuals must be aware of proximity to conspecifics, as spatial association can facilitate the acquisition or exchange of information about predators, resources, or social cues (Samplonius et al. 2017; Shen et al. 2017), which ultimately may prompt them to adopt various space use and movement tactics during incubation (Doligez et al. 2002; Hatchwell 2009). However, being near conspecifics may expose the individual to increased predation risk and conspecific parasitism, which has been documented in multiple species of ground nesting birds (Yom‐Tov 2001; Krakauer 2008; Deeming and Reynolds 2015; Sullivan et al. 2022).
Female eastern wild turkeys ( Meleagris gallopavo silvestris , hereafter wild turkey) are uniparental ground‐nesters that have long incubation periods (e.g., 25–30 days) during which they must identify suitable foraging opportunities and concealment spots to evade predators effectively (Healy 1992; Conley et al. 2015). Ground‐nesting coupled with extended incubation periods, along with a reliance on crypsis for survival and exposure to multiple predator guilds, results in increased vulnerability to nest loss by females (Blomberg et al. 2015). Throughout incubation, female wild turkeys exhibit central place foraging behavior, making periodic foraging excursions from the nest site (Conley et al. 2015; Bakner et al. 2019, 2024; Lohr et al. 2020). To survive the incubation period and reproduce successfully, female wild turkeys must identify resource‐rich areas that offer both foraging opportunities and predator concealment (Green 1982; Martin 1998).
A suite of studies has examined how vegetation structure at and around nest sites affects nest success; however, the extant literature remains inconclusive, with reported results being contradictory (Byrne and Chamberlain 2013; Yeldell et al. 2017; Bakner et al. 2019; Lohr et al. 2020; Keever et al. 2022). Similarly, early studies inferred that female wild turkeys prospected for nest locations (Badyaev 1995; Chamberlain and Leopold 2000), but contemporary studies have shown that such prospecting does not occur (Conley et al. 2015; Collier et al. 2019). However, although prenesting and laying ranges may overlap little, female wild turkeys display increased daily movements and reduced space use during the laying phase (Schofield 2019). Such movement patterns are consistent with prospecting behavior, whereby individuals explore local conditions to identify suitable nest sites and profitable foraging patches prior to incubation (Pärt and Doligez 2003). Under movement‐based decision frameworks, behavioral decisions are expected to reflect information gained through prior space use rather than assumptions about site familiarity per se (Matthiopoulos et al. 2005). Specifically, during laying but before entering the incubation phase, female wild turkeys engage in prospecting behavior, exploring their surroundings to assess resource distribution (Bakner et al. 2024), which may facilitate efficient travel to and from resources while minimizing mortality risks (Piper 2011; Wakefield et al. 2015).
The nesting ecology of female wild turkeys has been extensively studied. Contemporary literature has noted that turkey populations exhibit high rates of nest loss (Byrne and Chamberlain 2013; Crawford et al. 2021), prompting females to adopt spatial and movement decisions to combat these issues (Lohr et al. 2020). However, despite extensive research on nest locations and vegetation at nest sites (Porter 1992; Fuller et al. 2013; Streich et al. 2015; Little et al. 2016), the behaviors of female wild turkeys during the laying period remain relatively unstudied (Bakner et al. 2024). For instance, it is plausible that movements exhibited by female wild turkeys during the laying period could impact nest success and female survival, both of which can influence population trajectories (Vangilder and Kurzejeski 1995; Lehman et al. 2022). Bakner et al. (2024) demonstrated that the number of times females revisited areas within their incubation ranges did not influence nest fate, but rather, the number of patches visited during incubation recesses did. However, it is unclear if female movements during laying influence metrics of nest success and female survival during incubation.
Our objectives were to describe space use of female eastern wild turkeys during the egg laying period, including home range and core area sizes, average daily distance traveled, and overlap in space use with other females, and to relate these metrics to nest success and female survival during incubation. Contemporary studies continue to demonstrate that movements of wild turkeys can have important fitness consequences related to both individual survival and reproductive success (Lohr et al. 2020; Gulotta et al. 2024, 2025a, 2025b). Therefore, we hypothesized that metrics of space use and movements by females during the laying period would influence nest success and female survival rates during incubation. Specifically, we predicted that females who traveled greater daily distances and had larger home ranges and core areas during laying would experience greater individual survival and rates of nest success, as increased movement may enhance their knowledge of local resources and escape routes, thereby reducing their risk of predation. Secondly, we hypothesized that maintaining core areas that overlapped with other females would affect nest success and female survival rates during incubation. We predicted that females who maintained overlapping core areas during the laying period would have decreased individual survival and nest success rates because of increased attraction of predators to the area.
Methods
2
Study Area
2.1
We conducted research across study sites in Georgia, South Carolina, and Louisiana, USA. In Georgia, we conducted research on 3 wildlife management areas (WMAs), including Silver Lake, Cedar Creek, and B.F. Grant WMAs. The Silver Lake WMA, located in southwest Georgia, was owned and managed by the Georgia Department of Natural Resources—Wildlife Resources Division (GADNR). Silver Lake WMA was dominated by mature pine (Pinus spp.) forests and forested wetlands. Overstory species were predominately longleaf pine ( P. palustris ), loblolly pine ( P. taeda ), slash pine ( P. echinata ), oaks (Quercus spp.), and sweetgum ( Liquidambar styraciflua ). Prescribed fire was applied on an approximately 2‐ to 3‐year return interval. Cedar Creek WMA was owned by the US Forest Service (USFS) and managed in partnership with the Georgia Department of Natural Resources‐Wildlife Resources Division (GADNR). Cedar Creek was composed primarily of loblolly pine uplands, mixed hardwood‐pine forests, and hardwood lowlands containing mostly oaks, sweetgum, and hickory (Carya spp.). Prescribed fire was applied to pine‐dominated stands on an approximately 3–5‐year return interval. B.F. Grant was owned by the Warnell School of Forestry and Natural Resources at the University of Georgia and managed jointly by GADNR and the Warnell School. B. F. Grant was dominated by loblolly pine stands, agricultural lands, mixed hardwood‐pine forest, and hardwood lowlands of similar species composition as Cedar Creek.
Our two study sites in South Carolina included the Webb WMA Complex and the Savannah River Site (SRS). We conducted research on the Webb WMA Complex from January 2014–March 2018. The Webb WMA Complex was a combination of three adjacent WMAs (Webb, Palachacola, and Hamilton Ridge). These WMAs covered a 10,483‐ha area and were owned and managed by the South Carolina Department of Natural Resources (SCDNR) and consisted primarily of longleaf, loblolly, and slash pine forests with hardwood stands along riparian corridors, and expanses of bottomland hardwood wetlands. Management activities included prescribed fire, timber management, fallow field management, and maintaining agricultural field plots focused on enhancing habitat for wildlife species. During 2021–2023, we conducted research on SRS, which was a 78,000‐ha tract owned by the United States Department of Energy and jointly managed by the USFS. Approximately 94% of SRS was dominated by loblolly, longleaf, and bottomland hardwood forests. The remaining forested areas included forested swamps, riparian areas, and mixed hardwood stands. Approximately 30% of SRS was managed for red cockaded woodpeckers ( Picoides borealis ), with prescribed fire applied on a 3–5 year return interval. For detailed descriptions of site conditions on SRS and Webb WMA see Wightman et al. (2019).
We also conducted research on two sites in Louisiana. The western Louisiana (WLA) site included Kisatchie National Forest (KNF) and Fort Johnson Wildlife Management Area (FJWMA). The KNF was owned and managed by the United States Forest Service (USFS), whereas FJWMA was jointly owned by the USFS and the United States Army. Both sites were composed of forest dominated by pines, hardwood bottoms, and forested wetlands, and included forest openings and roads throughout the sites. Our study site in southeastern Louisiana (SELA) included a broad collection of private properties and Sandy Hollow WMA owned by the Louisiana Department of Wildlife and Fisheries. This study site consisted of rolling hills with longleaf pine plantations, hardwood riparian zones, and agricultural areas used primarily for grazing and row crops.
Data Collection
2.2
We captured female wild turkeys using rocket nets baited with cracked corn during January–March from 2014 to 2023. We classified captured individuals as juvenile or adult based on barring on the ninth and tenth primary feathers (Dickson 1992). We marked each individual with an aluminum rivet tarsal band (National Band and Tag company, Newport, Kentucky, USA) and a backpack style GPS/UHF transmitter (e‐obs GmbH Grünwald, Germany) or GPS/VHF transmitter (Lotek UK Ltd., Wareham, Dorset, UK). All individuals were released at the capture location immediately following processing. The capture, handling, and marking procedures were approved by the Institutional Animal Care and Use Committee at the University of Georgia (Protocol Numbers A2014 06‐008‐R2, A2014 06‐008‐R2, A2019 01‐025‐R2, A2020 06‐018‐R1, and A2021 11‐024‐Y1‐A0) and Louisiana State University (Protocol Numbers A2014‐013, A2015‐07, and A2018‐13).
We programmed Lotek transmitters to record 1 GPS location nightly to provide a roost location and hourly GPS locations from 0500 to 2000 (Standard Time and according to the appropriate time zones) for the duration of the study (Cohen et al. 2018). We programmed e‐obs transmitters to record hourly GPS locations from 0400 to 2000 for the duration of the study and used the 0400 point as the roost location. Units collected data until the battery died or the individual died, and the transmitter was retrieved. To monitor the status and locations of all birds, we used radiotelemetry via a UHF handheld command unit or VHF receiver. We used a BaseStation (e‐obs GmbH Grünwald, Germany) or a PinPoint VHF Commander (Lotek UK Ltd., Wareham, Dorset, UK) to remotely download data from transmitters at least once a week from January to July. During the nesting period, we increased the monitoring and data downloads of females to ≥ 2 times per week. We viewed GPS locations and determined incubation initiation by noting when female location became centralized around a single point (Conley et al. 2015; Yeldell et al. 2017; Wood et al. 2019). We did not disturb the nests while females were incubating. Instead, we continued to monitor them daily from a distance > 20 m via VHF/UHF. Upon nest termination, we located the nest site to determine if hatching had occurred and considered nests successful if ≥ 1 egg hatched (Conley et al. 2016). We continued to monitor females for additional nest attempts until reproductive activity ceased.
Analysis
2.3
We performed data processing and analysis in program R (v.4.3.1; R Development Core Team 2023). We processed and cleaned the raw GPS data by removing locations that had dilution of precision values (DOP) > 7. To determine dates of the onset of the egg laying and incubation period, we mapped our spatial–temporal data using FireTail (Schäuffelhut Berger GmbH, Unterhaching, Germany) and ArcPro (Environment Systems Research Institute, Redlands, California, USA). We determined locations for each nesting attempt by when an individual's locations became concentrated around a single point for several days (Guthrie et al. 2011; Conley et al. 2015; Yeldell et al. 2017; Wood et al. 2019). We determined the initiation of incubation as the first time an individual remained on the nest overnight (Bakner et al. 2019; Lohr et al. 2020) and then evaluated hourly locations for the previous 20 days to determine when a female initially visited the nest site. After identifying an incubating bird, we used a 27.5 m buffer (accounting for conservative GPS error; Bakner et al. 2019) to identify the date of first visit. We considered this first visit to the nest as the date of nest initiation and the beginning of the laying period. We recognize that this methodology could be biased by a day if the female visited the nest prior, but we based this approach on previously published works (Conley et al. 2016; Collier et al. 2019).
Covariates
2.4
For each female, we estimated average daily distance traveled for the laying period for each nesting attempt by summing the total distance traveled daily within the laying period (number of days laying) divided by length (in days) of the laying period. We calculated 95% home ranges and 50% core areas during the laying period by fitting dynamic Brownian bridge movement models (dBBMMs) to the time‐specific location data (Cohen et al. 2018) using package move (Kranstauber et al. 2018) in program R (v.4.3.1; R Core Team 2023). We used an error estimate of 20 m, a moving window size of 7 locations, and a margin setting of 3 locations (Byrne et al. 2014; Cohen et al. 2018). We kept window and margin size constant to account for changes in GPS sampling frequency because we failed to see any measurable effects of altering these values when we began our analysis (Cohen et al. 2018).
We quantified range overlap by categorizing whether simultaneously monitored females had overlapping ranges (intersected = 1, no intersection = 0). Specifically, we calculated these metrics across three different combinations: 95% home range that overlapped with another 95% home range, 50% core area that overlapped with another 50% core area, and 50% core area that overlapped with a 95% home range (Figure 1). Additionally, we calculated how many individual females overlapped with one another during the laying period using these same three combinations (Figure 1).
95% and 50% utilization ranges during the laying period for four female eastern wild turkeys to depict how covariates used to evaluate overlap in space use among females were determined. (A) depicts overlapping 95% utilization (home ranges), (B) depicts overlapping 50% utilization (core areas), and (C) depicts 95% home ranges that overlap to 50% core area.
In preliminary analyses, we assessed the correlations among all covariates using Pearson's correlation coefficient. We found that average daily distance traveled, home range size, and core area size were highly correlated (r = 0.79–0.86), suggesting significant multicollinearity among these spatial metrics. Similarly, substantial correlations were observed among the overlap variables (r = 0.67–0.92). To mitigate effects of multicollinearity and simplify the model, we selected average daily distance traveled to represent the spatial movement metrics and the number of females overlapping core areas to represent the overlap measures in our final analyses. This selection was based on their relevance to our research objectives and their comprehensive representation of the respective variable groups.
Nest Fate and Female Survival Models
2.5
We constructed two separate Bayesian logistic regression models to test our hypothesis regarding the effect of female movements and space use during the laying period on nest fate and female survival during incubation. For both models we included fixed effects of average daily distance traveled, total number of individual females who overlapped core areas during the laying period, nesting attempt, and a random effect of individual wild turkey. We treated the probability of nest fate and survival (success or failure; alive or dead) as a Bernoulli distribution. To aid model fit and allow for direct comparison of effect sizes of each predictor variable, we standardized all fixed effects included in the models using the scale function in R (v.4.3.1; R Core Team 2023). We fitted models using package brms in program R (Bürkner 2017). We computed 4 MCMC chains for 8000 iterations, discarding the first 1000 iterations as a burn‐in (Gelman and Rubin 1992). We calculated 95% credible intervals (hereafter CrI) that provided a metric of uncertainty and present the median of the posterior distribution. We then computed the probability of direction which describes the proportion of the posterior distribution that is in the direction of the estimated effect (Makowski, Ben‐Shachar, Chen, et al. 2019; Makowski, Ben‐Shachar, and Lüdecke 2019). We interpret effects where the CrI does not overlap zero or with a probability of direction ≥ 89% as showing support for an effect (Makowski, Ben‐Shachar, Chen, et al. 2019, Makowski, Ben‐Shachar, and Lüdecke 2019). Additionally, for statistically significant effects with effect estimates near zero, we calculated the region of practical equivalence (ROPE), which estimates the proportion of the posterior distribution that lies within a range considered too small to have biological relevance (Makowski, Ben‐Shachar, Chen, et al. 2019, Makowski, Ben‐Shachar, and Lüdecke 2019). For both models, we inspected trace and density plots, and all estimated parameters had R‐hat values < 1.1, indicating that all chains converged (Gelman 2004).
Results
3
We used 872 nesting attempts (initial attempts = 626, renesting attempts = 246) by 643 females (567 adults, 76 juveniles) for our nest fate and female survival analysis. Of the 872 nests, 210 (24%) hatched and 662 (76%) failed. Of the 643 females that initiated nests, 82 (13%) were depredated during incubation. Home range size during the laying period ranged from 3.02 ha to 584.11 ha with a mean of 16.39 ha, and core area size ranged from 0.62 ha to 73.31 ha with a mean of 16.39 ha (Table 1). Average daily distance traveled during the laying period ranged from 334.06 m to 4761.46 m with a mean of 2151.14 m (Table 1).
For nest fate, we found support for an increase in average daily distance traveled negatively influencing nest fate (β = −0.15, 95% CrI = −0.43–0.09, PD = 89.47%; Table 2, Figure 2). Although most of the posterior distribution for daily distance traveled during laying fell within the ROPE (60.09%), the effect cannot be conclusively considered biologically negligible. Specifically, with each increase of approximately 700 m (one standard deviation of variable, Table 1) in average daily distance traveled during laying, there was a 1.73% decrease in the probability of nest success, a minimal magnitude of effect that may not reflect a biologically meaningful relationship. We also found support that the total number of females that overlapped core areas (β = 0.34, 95% CrI = 0.11–0.65, PD = 99.84%; Table 2, Figure 2) positively influenced nest fate. An increase of 1 overlapping female (one standard deviation) was associated with a 4.76% increase in the probability of nest success. Lastly, we found no evidence that nest fate varied across nesting attempts (Table 2).
TABLE 2: Posterior means, 95% credible intervals, and probability of direction (pd) statistic for covariates used to model nest fate for female eastern wild turkeys across the southeastern United States during 2014–2023. Estimates > 0 positively influenced daily nest survival, whereas estimates < 0 negatively influenced survival.
Probability of nest success for female wild turkeys as a function of (A) average daily distance traveled (m) and (B) the number of females who had overlapping 50% core areas with a female during the laying period. Error bars and gray areas represent the 95% credible intervals.
For female survival during incubation, we found weak support that an increase in average daily distance traveled during laying positively influenced female survival (β = 0.25, 95% CrI = −0.13–0.70, PD = 92.39%, ROPE = 33.30%; Table 3, Figure 3). Specifically, with each increase of approximately 700 m (one standard deviation of variable, Table 1) traveled daily there was a 0.93% increase in the probability of female survival during incubation, which is likely not a biologically meaningful effect. We observed no effects of the number of females that overlapped 50% core areas during the laying period on female survival while incubating (Table 3). Likewise, we found no differences in female survival across nesting attempts (Table 3).
TABLE 3: Posterior means, 95% credible intervals, and probability of direction (pd) statistic for covariates measured during the laying period and used to model survival of female wild turkeys during incubation across the southeastern United States during 2014–2023. Estimates > 0 positively influenced survival, whereas estimates < 0 negatively influenced survival.
Probability of survival for female wild turkeys during incubation as a function of the average daily distance traveled (m) during the laying period. Gray areas represent the 95% credible intervals.
Discussion
4
We used hourly spatial locations to describe space use and movements of female eastern wild turkeys during the egg laying period and subsequently related those spatial behaviors to individual survival and nest success during the incubation period. We found that daily distances traveled by females during laying only weakly influenced both nest fate and female survival during incubation. We also found that overlapping core areas with conspecifics during egg laying positively influenced nest fate. Our findings contribute to a growing body of literature suggesting that movements and space use of female wild turkeys during portions of their reproductive period may influence metrics of reproductive success (Fontaine and Martin 2006), and specific to wild turkeys, such tactics are occurring during both egg laying and incubation (Bakner et al. 2019; Lohr et al. 2020).
The egg laying period is significant as it represents a temporal period when female movements are restricted, therefore limiting their space use and access to available resources (Kacelnik 1984; Van Gils and Tijsen 2007). We found that on average female wild turkeys traveled 2151 m per day during the egg laying period and maintained home ranges and core area sizes of 90 ha and 16 ha, respectively. We note that only a single previous study evaluated movements of female turkeys during laying, with that work reporting larger range sizes which included the eastern and Rio Grande wild turkey subspecies (Schofield 2019). However, Schofield (2019) used a 99% utilization distribution to calculate home range sizes, whereas we used a 95% utilization distribution to calculate home range size as is consistent with other studies using dBBMM (Byrne et al. 2014; Cohen et al. 2018). Previous studies on spring home range sizes in other Galliformes have shown that species inhabiting open landscapes, such as sharp‐tailed grouse ( Tympanuchus phasianellus ), sage grouse ( Centrocercus urophasianus ), and black grouse ( Lyrurus tetrix ), often have home ranges > 150 ha (Ballard and Robel 1974; Rippin and Boag 1974; Wallestad and Pyrah 1974; Svedarsky 1979), whereas those in denser, forested habitats, like ruffed grouse ( Bonasa umbellus ), spruce grouse ( Canachites canadensis ), western capercaillie ( Tetrao urogallus ), and blue grouse ( Dendragapus obscurus ), maintain home ranges < 50 ha (Herzog and Boag 1978; Maxson 1978; Hannon et al. 1982; Wegge 1985; Wegge and Rolstad 1986). Therefore, the home ranges we observed for female wild turkeys during egg laying are comparable to what has been reported in other species using a similar mating system during the breeding and reproductive periods.
Movements during reproductive periods can influence individual metrics of fitness and reproductive success (Lyon and Anderson 2003; Beck et al. 2006). Females must balance the energetic demands of producing offspring, resource acquisition, and evading predators, which leads to spatial and movement decisions (Jones 1989; Boggs 1992; Fontaine and Martin 2006; Deeming and Reynolds 2015). Our findings showed weak support for a negative association between an increase in average daily distance traveled by females during egg laying and nest fate, as well as a weak positive influence on female survival. Although these effect sizes were small, the observed patterns were similar to findings by Lohr et al. (2020), who demonstrated females who took longer incubation recesses during the incubation period had lower nest success but greater individual survival. Increased daily movement may reflect exploratory behaviors related to resource assessment or predator avoidance; however, any such effects appear limited in magnitude and should be interpreted cautiously (Williams 1996; Gehr 2020; Bakner et al. 2024). Alternatively, some nesting sites could be in areas that are more resource‐rich than others, requiring females that use resource‐rich nesting areas to travel lesser distances than their counterparts. As such, we cannot rule out that resources were heterogeneously distributed across the landscape, and resource distribution could be driving the observed pattern of greater nest loss with increasing distances traveled.
The presence of conspecifics can influence female behaviors and reproductive success in a variety of avian species (Ramsey et al. 1999; Kivelä et al. 2014). Previous research demonstrates that female birds may use conspecifics as cues to select nesting sites, and that social information from conspecifics positively influences reproductive success (Forsman et al. 2007; Hoi and Griggio 2012). We found that increases in the number of overlapping core areas with conspecific females positively influenced nest success in wild turkeys, which suggests that females perceive reproductive advantages to sharing space with conspecifics during the laying period. We speculate that these findings may stem from females benefiting from information acquisition relative to predation risk and resource distribution, which has been demonstrated in various bird species (Thornton and Raihani 2008; Farine et al. 2012; Firth and Sheldon 2016; Samplonius et al. 2017). However, increased overlap may also arise from nonsocial processes, such as multiple females independently selecting similar nesting cover or landscape features that confer higher nest success (Doligez et al. 2002; Danchin et al. 2004), or from predator swamping effects that reduce per‐capita predation risk in areas of higher nest density (Lima 2009). Because we did not document direct interactions among females and could not account for unmarked conspecifics, we caution that the spatial overlap we observed should not be interpreted as evidence of coordinated or cooperative behavior. Future research explicitly quantifying social interactions, habitat features, and predator responses will be necessary to disentangle these mechanisms.
We reached two primary conclusions from our work, including that increased movements by females only weakly influence nest success and female survival rates during incubation, and that overlapping space use with conspecifics can positively influence reproduction in wild turkeys. Positive fitness consequences via interactions with conspecifics have been observed in other avian species (Ramsey et al. 1999; Kivelä et al. 2014; Joly et al. 2024) likely reflecting a combination of social and nonsocial processes operating across heterogeneous landscapes, rather than a single mechanism driving movement and spatial overlap (Doligez et al. 2002; Danchin et al. 2004; Lima 2009). Furthermore, various factors can influence conspecific interactions in wild turkeys, including social dominance and relatedness. Previous research noted that female turkeys join social groups prior to breeding composed mostly of unrelated individuals (Watkins et al. 2025), and contemporary research suggests that these social groups tend to nest in the same area (Ulrey et al. 2023). Future research should explicitly identify the social, behavioral, and habitat‐driven mechanisms that link movement behavior and spatial overlap to variation in vital rates, particularly nest success and female survival, during the reproductive period.
Author Contributions
Paige E. Goodman: conceptualization (equal), data curation (lead), formal analysis (equal), investigation (supporting), methodology (supporting), writing – original draft (lead). Nicholas W. Bakner: conceptualization (supporting), formal analysis (supporting), methodology (supporting), writing – review and editing (supporting). Nickolas A. Gulotta: formal analysis (supporting), methodology (supporting), writing – review and editing (supporting). Erin E. Ulrey: formal analysis (supporting), methodology (supporting), writing – review and editing (supporting). Bret A. Collier: formal analysis (supporting), methodology (supporting), writing – review and editing (supporting). Michael J. Chamberlain: conceptualization (supporting), funding acquisition (lead), investigation (supporting), project administration (lead), supervision (lead), writing – review and editing (supporting).
Funding
This work was supported by the Georgia Department Of Natural Resources and South Carolina Department of Natural Resources.
Ethics Statement
This research was conducted with approval from the Institutional Animal Care and Use Committee at the University of Georgia (Protocol #A2014 06‐008‐R2, A2014 06‐008‐R2, A2019 01–025‐R2, A2020 06‐018‐R1, and A2021 11‐024‐Y1‐A0) and Louisiana State University (Protocol #A2014‐013, A2015‐07, and A2018‐13).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Avise, J. C. 1996. “Three Fundamental Contributions of Molecular Genetics to Avian Ecology and Evolution.” Ibis 138: 16–25.
- 2Badyaev, A. V. 1995. “Nesting Habitat and Nesting Success of Eastern Wild Turkeys in the Arkansas Ozark Highlands.” Condor 97: 221–232.
- 3Bakner, N. W. , L. R. Schofield , C. Cedotal , M. J. Chamberlain , and B. A. Collier . 2019. “Incubation Recess Behaviors Influence Nest Survival of Wild Turkeys.” Ecology and Evolution 9: 14053–14065.31938503 10.1002/ece 3.5843 PMC 6953688 · doi ↗ · pubmed ↗
- 4Bakner, N. W. , E. E. Ulrey , B. A. Collier , and M. J. Chamberlain . 2024. “Prospecting During Egg Laying Informs Incubation Recess Movements of Eastern Wild Turkeys.” Movement Ecology 12: 4.38229127 10.1186/s 40462-024-00451-3PMC 10792941 · doi ↗ · pubmed ↗
- 5Ballard, W. P. , and R. J. Robel . 1974. “Reproductive Importance of Dominant Male Greater Prairie Chickens.” Auk 91: 75–86.
- 6Beck, J. L. , K. P. Reese , J. W. Connelly , and M. B. Lucia . 2006. “Movements and Survival of Juvenile Greater Sage Grouse in Southeastern Idaho.” Wildlife Society Bulletin 34: 1070–1078.
- 7Blomberg, E. J. , D. Gibson , and J. S. Sedinger . 2015. “Biases in Nest Survival Associated With Choice of Exposure Period: A Case Study in North American Upland Game Birds.” Condor 117: 577–588.
- 8Boggs, C. L. 1992. “Resource Allocation: Exploring Connections Between Foraging and Life History.” Functional Ecology 6: 508–518.