Stage-dependent trade-offs in thermal performance: fluctuating temperatures reverse larval and adult fitness in Anopheles gambiae and An. coluzzii
Mauro Pazmiño-Betancourth, Alena Miller, Maria Katsoni, Ivan Casas Gomez-Uribarri, Fredros O Okumu, Simon A Babayan, Francesco Baldini

TL;DR
Malaria mosquitoes Anopheles gambiae and An. coluzzii show opposite survival and performance patterns in larvae and adults when exposed to fluctuating versus constant temperatures.
Contribution
Reveals species-specific and stage-dependent thermal performance trade-offs in malaria mosquitoes under fluctuating temperatures.
Findings
An. gambiae larvae survive better under fluctuating temperatures, while adults perform better under constant temperatures.
An. coluzzii larvae survive better under constant temperatures, but adults thrive under fluctuating temperatures.
Development time is slightly longer under fluctuating temperatures for both species.
Abstract
Temperature is a key environmental factor influencing the development, survival, and transmission potential of malaria vectors. While most laboratory studies use constant temperature (CT) regimes, mosquitoes in natural habitats experience fluctuating temperatures (FTs), which may affect their life-history traits. We investigated the effects of CT (27 °C) and FT (27 ± 3 °C) on larval and adult traits of 2 major malaria vectors, Anopheles gambiae and An. coluzzii, under laboratory conditions. We measured larval survival, development time, adult body size, and adult survival, using survival and mixed-effects models. Species-specific and stage-specific responses to temperature regimes were observed. An. gambiae larvae exhibited higher survival under FT, while An. coluzzii larvae survived better under CTs. However, this pattern reversed in adulthood: An. coluzzii adults showed increased…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
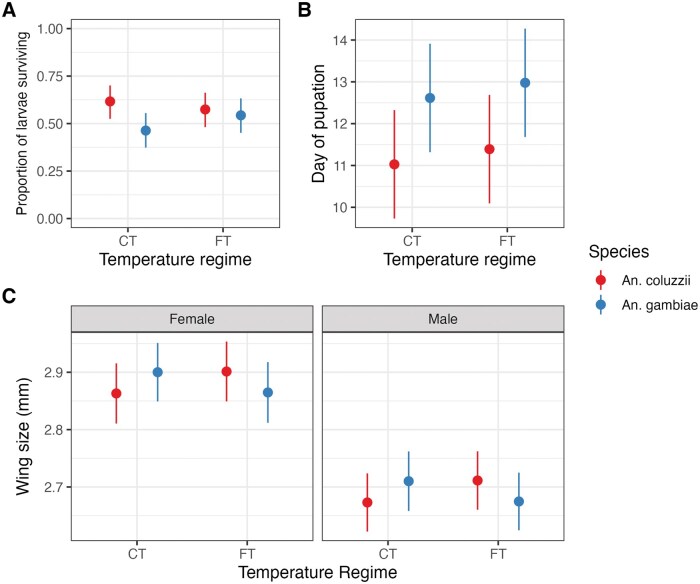 Fig. 1
Fig. 1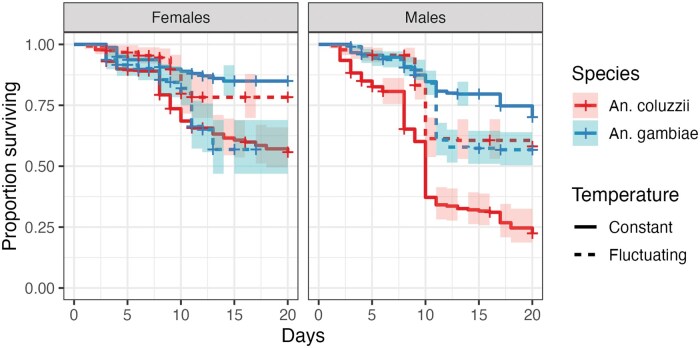 Fig. 2
Fig. 2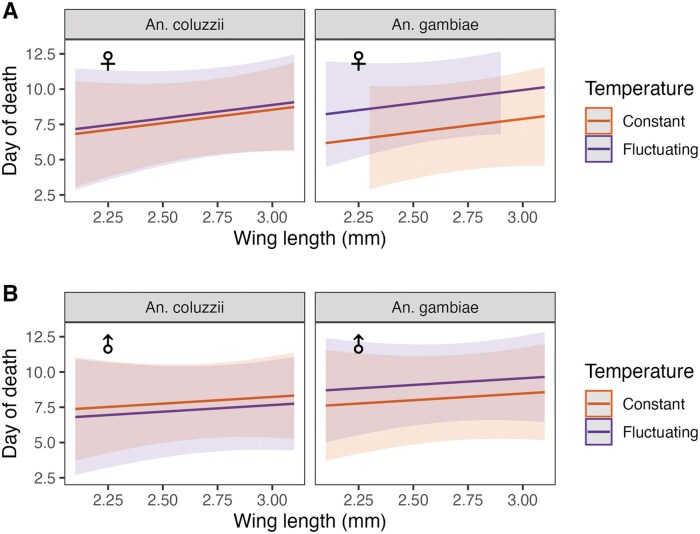 Fig. 3
Fig. 3| Type of model | Response variable | Explanatory variables | χ2 |
|
|---|---|---|---|---|
|
| Larval survival | Temperature × Species | 16.24 | <0.001 |
|
| Development time | Temperature × Species | 3.15 | 0.07 |
| Temperature | 20.85 | <0.001 | ||
| Species | 361.48 | <0.001 | ||
|
| Wing length (main effect) | Sex | 124.51 | <0.001 |
| Temperature × Species | 5.85 | <0.01 |
| Type of model | Response variable | Explanatory variables | χ2 |
|
|---|---|---|---|---|
|
| Survival | Temperature × Species | 78.32 | <0.001 |
| Sex | 29.74 | <0.001 | ||
|
| Survival | Temperature × Species | 23.82 | <0.001 |
| Wing length | 36.19 | <0.001 |
- —Bill and Melinda Gates Foundation10.13039/100000865
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Physiological and biochemical adaptations · Invertebrate Immune Response Mechanisms
Introduction
Climate change—including increased global temperatures (Climate Action Tracker 2023) and the rising frequency of extreme weather events (Stocker 2014, Fischer and Knutti 2015)—is predicted to impact the ecology of mosquitoes and their ability to transmit malaria and other vector-borne diseases (VBDs) (Thomson and Stanberry 2022). Indeed, the life-history traits of mosquitoes, including the major malaria vectors Anopheles gambiae, An. coluzzii, An. arabiensis, and An. funestus, are heavily influenced by environmental factors such as temperature, humidity, and rainfall (Afrane et al. 2012, Parham et al. 2012, Charlwood 2019). As ectotherms, mosquitoes are highly sensitive to temperature, which influences both their aquatic and terrestrial life stages (Caminade et al. 2019, Villena et al. 2022), as well as the development of the malaria parasite within the vector (Paaijmans et al. 2013a, Suh et al. 2024). As global warming is altering not only average daily temperatures but also the amplitude and frequency of temperature fluctuations (Colinet et al. 2015), understanding how fluctuating temperatures (FTs) affect mosquito ecology and malaria transmission dynamics under future climate change scenarios is vital [28]. This is also critical for informing climate-resilient vector control interventions (Ferguson and Govella 2023) and to optimize mosquito rearing protocols for sterile insect techniques (SIT), Wolbachia-based approaches, and gene drive technologies (Wang et al. 2025). Indeed, while most laboratory studies rear mosquitoes under constant temperature (CT) conditions (Christiansen-Jucht et al. 2014, Agyekum et al. 2021, Barr et al. 2023, 2024, Martin and Hillyer 2024), data on the effects of FT on mosquito ecology remain limited (Lyons et al. 2013, Paaijmans et al. 2013b, Davies et al. 2016, Hidalgo et al. 2016, 2018, Mamai et al. 2016).
In ectotherms, the association between temperature and life-history traits such as development, survival, and reproduction—commonly described through thermal performance curves (TPCs)—typically follows a unimodal function: performance increases from zero at a critical minimum (CT_min_), peaks at an optimum (T_opt_), and declines to zero at a critical maximum (CT_max_) temperature. While T_opt_ represents the temperature at which metabolic performance is maximized, CT_min_ and CT_max_ mark physiological thresholds beyond which cellular functions are severely impaired due to protein denaturation, disrupted membrane fluidity, mitochondrial dysfunction, and oxidative stress (Duffy et al. 2015, Bodlah et al. 2023). In mosquitoes, TPCs have been consistently quantified for traits including larval and adult survival, development rate, biting frequency, and reproductive output, highlighting performance constraints at both thermal extremes (Mordecai et al. 2013, Johnson et al. 2015, Villena et al. 2022).
Typically, TPCs exhibit an accelerating temperature–performance relationship below T_opt_ and a decelerating relationship above T_opt_, as shown in early work by Kaufmann (Kaufmann 1932) and known as Jensen’s inequality (Jensen 1906). This framework helps explain discrepancies between insect performance under CTs and FTs with the same mean. Specifically, when FT occurs in the accelerating portion of the curve (below T_opt_), performance often exceeds that observed under CT; conversely, when FT occurs above T_opt_, performance tends to be reduced compared to CT (Colinet et al. 2015). The magnitude and direction of these effects depend on 2 key factors: the thermal sensitivity of the trait in question (i.e., how steep or curved the TPC is), and the amplitude of temperature fluctuations. Traits with greater thermal sensitivity (steeper TPCs) and environments with larger thermal amplitudes exhibit stronger deviations between FT and CT outcomes (Colinet et al. 2015). Global warming is expected to increase the occurrence of these deviations. Since thermal sensitivity likely varies among mosquito species, current models of mosquito development may underestimate the significance of FT.
Mosquitoes undergo larval development in aquatic habitats that often experience marked diurnal temperature fluctuations. In sub-Saharan Africa, the primary malaria vectors exhibit species-specific habitat preferences. For instance, Anopheles funestus tends to occupy larger, deeper, and more stable water bodies (Kahamba et al. 2024), whereas members of the An. gambiae sensu lato complex typically breed in shallow, sunlit, and often ephemeral pools (Onen et al. 2021). Within this complex, An. gambiae and An. coluzzii differ in their geographic distributions, ecological preferences, and roles in malaria transmission (Wiebe et al. 2017, Mbama Ntabi et al. 2024). An. coluzzii predominates in savannah regions, particularly in southern West Africa, where it breeds in more permanent and human-modified habitats such as rice fields (Cassone et al. 2014). In contrast, An. gambiae is more common in forested areas, primarily in northern and central regions, and prefers temporary, rain-fed water bodies such as puddle margins (Gimonneau et al. 2010, Mattah et al. 2017, Koukpo et al. 2019). Although these ecological differences influence the seasonal dynamics of malaria transmission—An. coluzzii being more prevalent during the dry season and An. gambiae during the rainy season (Akogbéto et al. 2018)—the extent to which their distinct ecologies affect thermal sensitivity remains unknown.
The adaptation of An. gambiae and An. coluzzii to different habitats suggests that their thermal sensitivity and experienced thermal fluctuations may be different. However, while some data are available for An. gambiae and other Anopheles species, information on An. coluzzii is scarce. Most studies in laboratory conditions have relied on CT, with few using FT (Lyons et al. 2013, Paaijmans et al. 2013b, Davies et al. 2016, Hidalgo et al. 2016, 2018, Mamai et al. 2016, Shocket et al. 2025). Studies on life-history traits of An. gambiae using CT have shown that larval development is faster at high temperatures (Bayoh and Lindsay 2003), while both body size and survival decrease (Agyekum et al. 2021, 2022, Barr et al. 2023). Similar trends have been found in An. coluzzii with added interaction of survival with humidity (Faiman et al. 2017). The studies using FT have shown that An. coluzzii can adjust its cuticular hydrocarbons, increase its metabolic rate, and decrease its protein content when reared in low-temperature, low-humidity conditions, while An. gambiae does not (Hidalgo et al. 2016, 2018). However, life history and survival studies with extended follow-up times in both species are lacking. In other Anopheles species, it has been shown that FTs that include CT_min_ and CT_max_ values (18 to 35 °C) increase the survival of An. arabiensis larvae when reared in a mix with An. quadriannulatus (Davies et al. 2016). FT has a minimal effect on the survival of adult An. arabiensis when fluctuations are large (±20 °C) with a maximum temperature of 35 °C, compare to An. funestus whose survival greatly decreases even with small fluctuations (±10 °C) with a maximum temperature of 30 °C (Lyons et al. 2013). Overall, the literature collectively indicates that FTs alter mosquito life-history responses relative to constant conditions. Since mosquitoes are naturally exposed to FT conditions, there is a need to expand our understanding of these effects on vectors relevant for malaria transmission (Pascual et al. 2009, Paaijmans et al. 2010, 2013b, Shocket et al. 2025).
In this study, we compared the effects of CT and FT temperatures, both with a mean of 27 °C (the standard laboratory temperature for mosquito rearing), on key larval and adult life-history traits in An. gambiae and An. coluzzii in controlled laboratory settings. Our findings reveal contrasting species-specific responses across life stages. An. gambiae exhibited improved larval performance under FT relative to CT, whereas An. coluzzii performed better under CT. In adulthood, however, the pattern reversed: An. coluzzii showed increased survival and body size under FT, while An. gambiae fared better under CT. We hypothesized that An. gambiae, adapted to ephemeral, thermally variable habitats, would perform better under FTs, while An. coluzzii, associated with more stable aquatic environments, would benefit from constant conditions, though these patterns may vary across life stages.
Materials and Methods
Temperature Design
We assessed the effect of 2 temperature regimes on life-history traits of An. gambiae and An. coluzzii using environmental incubators (CLF Plant Climatics). The 2 temperature regimes were 27 °C (CT) and 27 ± 3 °C (FT), with daily temperature range (DTR) fluctuating between 24 °C and 30 °C, peaking at 12:00 pm, based on typical diurnal patterns in sub-Saharan Africa (Charlwood 2017, Suh et al. 2020), Supplementary Fig. S1). The shape of temperature fluctuations was programmed manually to be approximately sinusoidal. The rate of change between the different programmed points was linear. The 2 regimes had the same mean temperature; thus, any changes in life-history traits between the two would depend on the presence/absence of the fluctuation. We programmed a photoperiod of 12:12 h (L:D) cycle, with alternations occurring at 8:00 AM and 8:00 PM, and relavtive humidity (RH) was programmed at 75 ± 10% (as per the chamber’s specifications) across all experiments. This was constantly monitored using independent data loggers, which confirmed that the value always stayed within range.
Larval Rearing, Temperature Regimes, and Feeding
Two strains were used in the study: Anopheles gambiae Kisumu strain originating from Kisumu, Kenya, and Anopheles coluzzii, Ngousso strain from Yaoundé, Cameroon (Harris et al. 2010). For each replicate and species, 10 trays with 200 larvae and 500 ml of deionized water were placed into each environmental chamber. Larvae were fed with Tetramin Tropical Flakes and Tetra Pond Pellets (Tetra Ltd., United Kingdom) at approximately 0.92 mg/larva/day and reared into adults. For body size assessment, pupae were collected and placed in emergence cages. To control for the effects of larval development rate on adult size, each emergence cage contained the same number of pupae from larvae with fast, normal, and slow development rates. This was achieved by combining pupae collected during early (8 to 10 d), peak (11 to 14 d), and late (15 to 16 d) pupation times in each cage (n = 10 pupae per time point). For adult survival analysis, a total of 400 pupae were put into each cage and reared under the same conditions as they were reared as larvae. The day the pupae emerged into adults was assigned as day 0. Adults were fed with 5% glucose solution ad libitum. Females were blood-fed human blood through a membrane feeder 3 to 4 d after emergence and every 3 to 4 d subsequently. Three independent experimental blocks were conducted over consecutive weeks.
Adult Survival and Body Size
Adult survival was monitored for 20 d. Dead mosquitoes were collected, sexed, and counted daily. Wing lengths (distance from the alula notch to the tip) of dead males and females were measured using DinoCapture 2.0, under a Leica MZ95 microscope, and used as a proxy for body size (Maïga et al. 2012). Observers were not blinded.
Statistical Analysis
Analyses were carried out using R version 4.4.0 (R Core Team 2021), as follows: (1) For larval development time to pupation (L1 to pupation), a linear mixed model (LMM) was run using the “lmer” function from the “lme4” (Bates et al. 2015) package in R. Day of pupation was used as the response variable, temperature regime, species and their interaction were the explanatory variables with replicate as random effect. (2) For larval survival (measured as the proportion of L1 larvae that developed into pupae), a generalized linear mixed model (GLMM) with binomial distribution was run using the “glmer” function. Larval survival was the response variable, temperature regime, species, and their interaction were the explanatory variables, and replicate was the random effect. (3) To examine the effect of temperature on body size, an LMM was run using the “lmer” function. Wing length was used as the response variable. Temperature regime, species, and their interaction were used as explanatory variables; replicate was treated as a random effect.
Survival data were analyzed using Cox’s proportional hazards and parametric models using the package “survival” (Therneau and Grambsch 2000) and “flexsurvreg” (Jackson 2016). Proportional hazard assumptions were tested using the function cox.zph from the survival package. For parametric survival, the Akaike information criterion (AIC) was used to choose the best-fit distribution (Supplementary Table S1). A random intercept for replicate was included using a frailty term in the Cox model. Explanatory variables were species (2 levels), sex, and temperature regime (CT or FT), and the interaction between temperature and species. A separate Cox’s proportional hazards model was developed for females only, incorporating wing length as an additional explanatory variable. The significance of each variable was calculated through stepwise model simplification based on likelihood ratio tests using the “lmerTest” package for all models. Model assumptions were tested using the “DHARMa” package (Hartig 2024). Residuals showed no evidence of overdispersion or spatial/temporal autocorrelation.
Results
Larval Survival and Development Time
An. coluzzii larvae survival was higher at CT than at FT, whereas An. gambiae survival was higher under FT (interaction between species and temperature χ^2^ = 16.24, P < 0.001; Fig. 1A, Table 1).
Influence of temperature and species on larval survival, developmental time, and body size. (A) Predicted average number of larvae that survived stratified by species and different temperature conditions. Points represent the mean, and the lines represent the 95% confidence intervals. (B) Day of pupation by species and temperature regime. (C) Predicted wing sizes of An. gambiae (blue) and An. coluzzii (red) are divided into females and males under constant and fluctuating temperature conditions.
Conversely, while there was no interaction between temperature regime and species on developmental time, both variables were associated with the time that L1 larvae took to reach the pupal stage (Species: χ^2^ = 361.48, P < 0.001, temperature regime: χ^2^ = 20.85, P < 0.001) (Table 1), but the interaction was not significant. Unexpectedly, the developmental time in *An. coluzzii—*predicted to be a slower pace species—was, in fact, slightly shorter under both temperature regimes (Fig. 1B). Anopheles gambiae larvae took 1.58 d longer to pupate than An. coluzzii (95% CI: 1.43 to 1.74, P < 0.001). Overall pupation for both species was slightly faster at CT, as larvae under FT required an extra 0.36 d (95% CI: 0.21 to 0.52) to pupate.
Body Size
To understand if the temperature regime influenced pupal growth, the wing length of adults emerging from pupae collected from different days was measured as a proxy of their body size. There was a strong association between wing length and sex (χ^2^ = 124.51, P < 0.001), and a significant interaction between temperature and species (χ^2^ = 5.84, P < 0.01; Table 1). Specifically, An. coluzzii were larger when reared under FT than CT, whereas An. gambiae showed the opposite pattern (Fig. 1C).
Adult Survival
The effect of temperature on adult survival differed significantly between the 2 species (χ^2^ = 78.32, P < 0.001). In contrast to the effect of temperature regime on the 2 species during larval stages, during adulthood, An. coluzzii live longer at FTs compared to constant ones, while the opposite was observed for An. gambiae. Sex also influenced survival (χ^2^ = 29.74, P < 0.001), as males generally survived less than females (Fig. 2). To test the effect of body size on survival, a model including only females was created. When wing length was included, the significant effect of the interaction between temperature and species was still observed (χ^2^ = 23.82, P < 0.001). As expected, wing length was positively associated with survival (χ^2^ = 36.19, P < 0.001; Table 2) as larger individuals tended to live longer (Fig. 3). This finding suggests that the effects of temperature regimes on the survival of the 2 species were independent of their body size.
Kaplan–Meier survival plot of adult females and males, An. gambiae (blue line) and An. coluzzii (red line) reared under constant (solid line) and fluctuating temperature regimes (dashed line). Mortality of mosquitoes was recorded up to day 20. The shaded areas depict the 95% confidence intervals.
Relationship between longevity and wing length (mm) of An. gambiae and An. coluzzii at constant (orange line) and fluctuating (purple line) temperatures for females (A) and males (B). The relationship is visualized by the line of best fit (regression line), and the shaded areas represent the 95% confidence intervals.
Discussion
This study compared the effect of CT and FT on life-history traits of 2 Anopheles species, An. gambiae and An. coluzzii. We found that An. gambiae had higher larval survival and longer pupation time under FT, but larger adult body size and better adult survival under CT. In contrast, An. coluzzii showed higher larval survival under CT, but increased pupation time, larger adult size, and better adult survival under FT, showing species-specific thermal sensitivities across life stages.
These differences could be explained by the innate hard-wired preferences of different types of breeding sites for each species. Indeed, An. gambiae exploits temporary breeding sites (Hidalgo et al. 2018, Ebhodaghe et al. 2024), typically small and shallow, where temperature fluctuates more. On the contrary, An. coluzzii can be found in a wider variety of breeding sites with a preference to more permanent breeding sites, which are large and deep, where temperature variation is reduced (Gimonneau et al. 2012, Hidalgo et al. 2018). In contrast, An. arabiensis larvae show increased survival under FT (Davies et al. 2016), highlighting species-specific responses (Davies et al. 2016). However, similar to An. gambiae, FT increases larval survival in An. stephensi (Paaijmans et al. 2010). Both species pupated more slowly at FT, with an extra 0.36 d required to pupate. Similar studies on An. arabiensis and An. quadriannulatus, showed faster developmental times at CTs compared to fluctuating (Davies et al. 2016). Overall, An. coluzzii pupated faster across both temperature regimes compared to An. gambiae. This increase in fitness of An. coluzzii has been reported before in laboratory colonies, and it might be caused by colony-specific genetic drift (Gimonneau et al. 2014).
We found that the effect of FT on adult body size differed between species. An. gambiae adults reared under CT emerged larger than those reared under FT, whereas An. coluzzii exhibited the opposite pattern, with larger adults emerging under FT. These differences may reflect species-specific responses in larval survival. In An. gambiae, higher larval mortality under CT could have led to reduced competition for resources among the surviving individuals, enabling greater growth. Conversely, in An. coluzzii, improved larval survival under CT may have resulted in increased intraspecific competition, resulting in smaller emerging adults (Jannat and Roitberg 2013). Furthermore, differences in larval survival between the 2 species may have led to variations in bacterial proliferation and the accumulation of toxic waste products, which could have subsequently influenced larval development. The differences in wing lengths are the opposite of what was observed for larval survival, showing that when larval survival is higher for 1 species in 1 temperature regime, their body size is reduced, suggesting that larval density and competition—higher if more larvae survive—impacted on adult body size. While larval density was not directly manipulated, differences in survival may have led to varying levels of intraspecific competition, potentially influencing adult size.
The conditions experienced during the larval stage might have influenced also adult-stage outcomes. While An. gambiae adults showed higher survival under CT, An. coluzzii adults survived better under FT—a reversal of the larval survival patterns observed. This suggests that reduced larval competition, resulting from higher mortality (i.e., An. gambiae at CT and An. coluzzii at FT), may have conferred survival advantages to those individuals that reached adulthood. Importantly, our models included body size as an explanatory variable, indicating that these species-specific differences in adult survival are not simply mediated by body size, but may instead reflect selection processes acting during the larval stage. For example, the suboptimal temperature conditions that led to increased larval mortality may have acted as a selective filter, allowing only the most robust individuals to reach adulthood. Consequently, selective mortality during the larval stage may result in a cohort of more robust adults, potentially confounding direct temperature effects on adult survival—where the relationship between temperature and adult traits is conditioned on having survived the larval stage, rather than being a direct effect of the temperature regime itself. Moreover, the different survival to temperatures might not be fully mediated by their larval development, but instead by a different physiological response during the adult stage. The observed species differences may also be influenced by species-specific thermal performance. Since our FT conditions reached up to 30 °C—likely exceeding Topt for both species—the FT treatment could have differently reduced their performance during periods when temperatures surpassed their optimal range, depending on the shape of their TPCs.
Conclusion
This study highlights how even small daily temperature fluctuations can influence the life-history traits of Anopheles mosquitoes, with distinct responses observed between species under fluctuating versus CT conditions. These findings suggest that laboratory studies using CTs may misrepresent field performance, particularly for species like An. gambiae that exploit thermally variable habitats. This shows the importance of including daily variations when studying the impact of temperature and other environmental conditions on mosquito ecology. Future works should include larger fluctuations in temperature as well as humidity to gain a deeper understanding of the effects of ecological factors on life-history traits.
Supplementary Material
tjaf189_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Afrane YA , Githeko AK, Yan G. 2012. The ecology of Anopheles mosquitoes under climate change: case studies from the effects of deforestation in East African highlands. Ann. N Y Acad. Sci. 1249:204–210. 10.1111/j.1749-6632.2011.06432.x 22320421 PMC 3767301 · doi ↗ · pubmed ↗
- 2Agyekum TP , Arko-Mensah J, Botwe PK, et al. 2022. Effects of elevated temperatures on the growth and development of adult Anopheles gambiae (s.l.) (Diptera: Culicidae) Mosquitoes. J. Med. Entomol. 59:1413–1420. 10.1093/jme/tjac 04635452118 PMC 9278826 · doi ↗ · pubmed ↗
- 3Agyekum TP , Botwe PK, Arko-Mensah J, et al. 2021. A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate. IJERPH. 18147255 10.3390/ijerph 1814725534299706 PMC 8306597 · doi ↗ · pubmed ↗
- 4Akogbéto MC , Salako AS, Dagnon F, et al. 2018. Blood feeding behaviour comparison and contribution of Anopheles coluzzii and Anopheles gambiae, two sibling species living in sympatry, to malaria transmission in Alibori and Donga region, northern Benin, West Africa. Malar. J. 17:307. 10.1186/s 12936-018-2452-930134912 PMC 6106899 · doi ↗ · pubmed ↗
- 5Barr JS , Estevez-Lao TY, Khalif M, et al. 2023. Temperature and age, individually and interactively, shape the size, weight, and body composition of adult female mosquitoes. J. Insect Physiol. 148:104525. 10.1016/j.jinsphys.2023.10452537236342 · doi ↗ · pubmed ↗
- 6Barr JS , Martin LE, Tate AT, et al. 2024. Warmer environmental temperature accelerates aging in mosquitoes, decreasing longevity and worsening infection outcomes. Immun. Ageing. 21:61. 10.1186/s 12979-024-00465-w 39261928 PMC 11389126 · doi ↗ · pubmed ↗
- 7Bates D , Mächler M, Bolker B, et al. 2015. Fitting linear mixed-effects models using lme 4. J. Stat. Soft. 67:1–48. 10.18637/jss.v 067.i 01 · doi ↗
- 8Bayoh MN , Lindsay SW. 2003. Effect of temperature on the development of the aquatic stages of Anopheles gambiae sensu stricto (Diptera: Culicidae). Bull. Entomol. Res. 93:375–381. 10.1079/BER 200325914641976 · doi ↗ · pubmed ↗