Macrophage-derived chemokines in T cell regulation: implications for cancer immunotherapy
Kunpeng Zhang, Jingjing Liu, Qi Liu, Ningning Zhu, Baodong Ye

TL;DR
This review explores how macrophage-produced chemokines influence T cell behavior in cancer, offering insights into improving immunotherapy.
Contribution
The paper systematically reviews the dual roles of macrophage-derived chemokines in T cell regulation and cancer immunotherapy.
Findings
Macrophage-derived chemokines modulate T cell homeostasis, differentiation, and metabolic reprogramming.
TAMs secrete CCL22 and CCL5, which promote immunosuppression and CD8+ T cell exhaustion.
M1-like macrophages produce CXCL9 and CXCL10, enhancing anti-tumor immunity through CD8+ T cell activation.
Abstract
Macrophages are pivotal regulators of immunity, with intercellular communication being a central mechanism of their function. Among these communications, chemokines act as critical messengers in macrophage-T cell crosstalk. This review systematically elucidates the notable roles of macrophage-derived chemokines in modulating T cell homeostasis, particularly concentrating on their influence on both CD4+ and CD8+ T cell differentiation, proliferation, exhaustion, secretory activity, metabolic reprogramming (involving glycolysis and OXPHOS), chemotaxis, and memory formation. In the tumor microenvironment (TME), the dualistic nature of chemokines was highlighted: tumor-associated macrophages (TAMs) could secrete immunosuppressive factors, such as CCL22 and CCL5, recruiting inhibitory cells and inducing CD8+ T cell exhaustion. In contrast, M1-like macrophages could produce CXCL9 and CXCL10,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
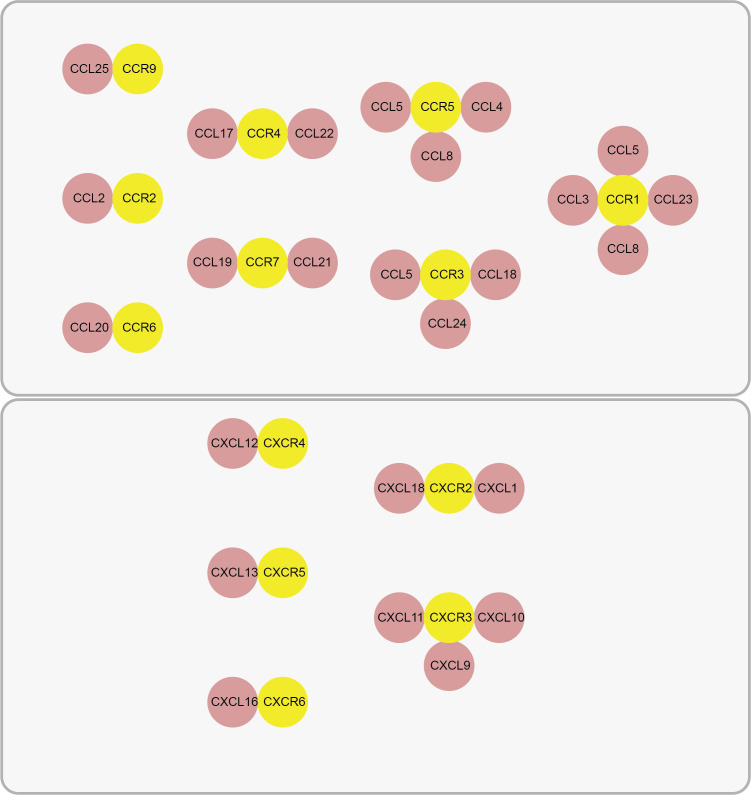 Figure 1
Figure 1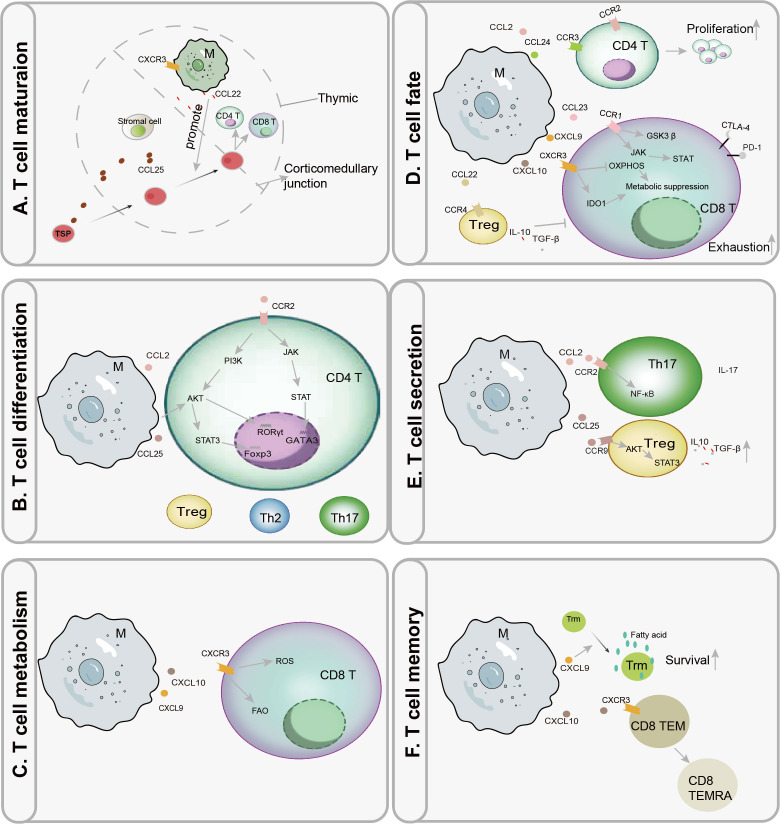 Figure 2
Figure 2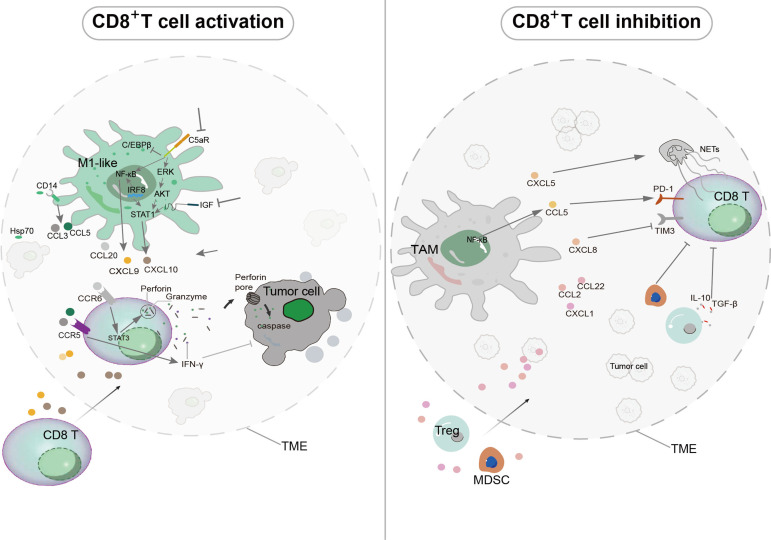 Figure 3
Figure 3- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune cells in cancer · Chemokine receptors and signaling · Single-cell and spatial transcriptomics
Introduction
1
Chemokines represent a family of signaling proteins that are indispensable for coordinating immune cell migration, activation, and intercellular communication. These functions are critical for maintaining physiological immunity and are also intimately involved in the pathogenesis of various diseases (1). As prolific sources of these signals, macrophages emerge as pivotal regulators of T cell biology. Their secretion of a diverse array of chemokines allows them to modulate multiple facets of T cell activity, including development, differentiation, metabolic reprogramming, migration, and effector functions (2–6). This regulatory role is particularly consequential within the tumor microenvironment (TME), where the dysregulated expression of chemokines by tumor-associated macrophages (TAMs) is a well-established driver of immune evasion and tumor progression (5).
Consequently, this review aimed to systematically elucidate the mechanisms through which macrophage-derived chemokines govern T cell homeostasis and immune function. This review advances beyond previous reviews by systematically addressing the multifaceted roles of macrophage-derived chemokines in modulating T cell homeostasis, particularly concentrating on their influence on metabolic reprogramming, including glycolysis and oxidative phosphorylation (OXPHOS). In contrast to earlier reviews that primarily concentrated on immune activation or suppression, this review explored the dualistic nature of chemokines in the TME, emphasizing their context-dependent roles in promoting both anti-tumor immunity and immune evasion. Additionally, it provided a comprehensive analysis of specific chemokine signaling axes, such as CCL2/CCR2 and CXCL10/CXCR3, and their therapeutic potential in cancer immunotherapy. By integrating these novel insights, this review provided a more complete understanding of the complex interaction between macrophages and T cells, highlighting emerging strategies for potentiating T cell-mediated anti-tumor responses through targeted chemokine modulation.
Macrophage origin and plasticity
2
Macrophages originate from embryonic precursors or are derived from bone marrow monocytes (7, 8). Their defining characteristic is remarkable functional plasticity, enabling them to adapt to diverse microenvironments and meet varying immunological demands. Historically, macrophages were broadly categorized into two primary phenotypes: the classically activated phenotype (often termed M1-like), which is pro-inflammatory, and the alternatively activated phenotype (often termed M2-like), associated with anti-inflammatory and tissue-repair properties (9). This classification was originally based on the helper T cell (Th)1/Th2 paradigm and reflected the dual roles of macrophages in both the initiation and resolution of inflammation. However, it is currently understood that macrophage phenotypes are not fixed while rather exist along a dynamic spectrum, continuously regulated by local microenvironmental cues.
Although initial models simplified macrophage activation into two major types, including pro-inflammatory M1-like and anti-inflammatory M2-like, it is widely recognized that macrophages exhibit a remarkable functional diversity. Their phenotypic state can be shaped by a range of stimuli from the surrounding microenvironment. For instance, interferon-γ (IFN-γ) alone or in combination with lipopolysaccharide promotes a pro-inflammatory, M1-like macrophage phenotype, while interleukin (IL)-4 and IL-13 drive polarization toward an anti-inflammatory, M2-like profile (10, 11). In the TME, for instance, TAMs mainly exhibit functional properties resembling the M2-like phenotype and may contribute to immune evasion and disease progression (5).
Functionally, macrophages with an M1-like orientation are adept at phagocytosing pathogens, clearing damaged cells, and presenting antigens, which are crucial for host defense (12). In contrast, those with an M2-like orientation are specialized in tissue remodeling and repair, typically displaying reduced antigen-presentation capacity (13). Beyond these primary functions, macrophages are key modulators of adaptive immune responses through the secretion of chemokines, playing a pivotal role in regulating T cell functions and overall immune responses (14).
Chemokine structure and function
3
Chemokines are a family of low-molecular-weight (8–10 kDa) signaling proteins characterized by their short half-lives (15). Their classification into four distinct subfamilies (CC, CXC, XC, and CX3C) is defined by the arrangement of conserved N-terminal cysteine residues. A key structural determinant is the presence or absence of an intervening amino acid between the first two cysteines; this motif critically influences the protein’s tertiary structure, stability, and receptor binding affinity (15). The primary biological function of chemokines is executed through their binding to specific G-protein-coupled receptors (GPCRs), as shown in Figure 1, which illustrates the receptor-ligand interaction and the resulting signaling cascade. This ligand-receptor interaction initiates intracellular signaling cascades that govern essential leukocyte behaviors, including directed migration, survival, and functional polarization (16–18). Consequently, chemokines are fundamental orchestrators of immune cell trafficking, activation, and communication, thereby coordinating the responses of lymphocytes (B and T cells) and innate immune cells like natural killer (NK) cells (19–21). This precise coordination is indispensable for launching effective antimicrobial defenses and for maintaining immune homeostasis (22). However, pathogenic dysregulation of chemokine signaling disrupts this equilibrium and contributes to a spectrum of diseases, ranging from chronic inflammatory and autoimmune disorders to cancer (18). For instance, within the TME, specific chemokines facilitate the recruitment of immunosuppressive cell populations—including regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs)—which in turn promote immune evasion, tumor progression, and metastasis (23). The central role of chemokines in both health and disease underscores their significant potential as therapeutic targets for immunomodulation.
Chemokines and their receptors. CCR9 Ligand (CCL25); CCR2 Ligand (CCL2); CCR6 Ligand (CCL20); CCR4 Ligand (CCL17, CCL22); CCR7 Ligand (CCL19, CCL21); CCR5 Ligand (CCL4, CCL5, CCL8); CCR3 Ligand (CCL5, CCL18, CCL24); CCR1 Ligand (CCL3, CCL5, CCL8, CCL23). CXCR4 Ligand (CXCL12); CXCR5 Ligand (CXCL13); CXCR6 Ligand (CXCL16); CXCR2 Ligand (CXCL1, CXCL18); CXCR3 Ligand (CXCL9, CXCL10, CXCL11).
Macrophage-derived chemokines
4
Macrophages are integral architects of T cell homeostasis, primarily through their regulated secretion of chemokines, as depicted in Figure 2. This secretory activity critically shapes multiple facets of T cell biology, from development to effector function. Accordingly, this section will dissect the specific mechanisms by which macrophage-derived chemokines modulate pivotal T cell activities. We will highlight their indispensable, yet context-dependent, roles in maintaining immune equilibrium and in driving the pathogenesis of various diseases.
Macrophage-derived chemokines orchestrate T cell homeostasis, differentiation, and function. (A) Stromal cell-derived CCL25 facilitates the entry of thymus-seeding progenitors (TSPs) into the thymus. Subsequently, macrophage (M)-derived CCL22 guides their migration to the corticomedullary junction, the site for CD4+ and CD8+ T cell lineage commitment. CX3CR1+ macrophages contribute to central tolerance by supporting the negative selection of self-reactive T cells. (B) Macrophage-derived CCL2 binding to CCR2 activates the JAK-STAT/GATA3 pathway to promote Th2 polarization. Concurrently, CCL2/CCR2 signaling via the PI3K-AKT pathway upregulates RORγt to drive Th17 differentiation. In contrast, CCL25 signaling through AKT/STAT3/Foxp3+ facilitates regulatory T cell (Treg) development. (C) The CXCL9/10-CXCR3 axis enhances reactive oxygen species (ROS) levels and promotes fatty acid oxidation (FAO) in CD8+ T cells, optimizing them for effector functions. (D) Macrophage-derived CCL2/CCR2 and CCL24/CCR3 signaling promotes CD4+ T cell expansion. Conversely, CCL23/CCR1 engagement upregulates immune checkpoint molecules (CTLA-4, PD-1) on CD8+ T cells via GSK3β or JAK-STAT pathways, promoting exhaustion. Additionally, CXCL9/10 signaling via CXCR3 induces a metabolically suppressive state by downregulating oxidative phosphorylation (OXPHOS) and enhancing indoleamine 2,3-dioxygenase 1 (IDO1)-mediated tryptophan degradation. CCL22/CCR4 signaling stimulates Tregs to produce immunosuppressive cytokines (IL-10, TGF-β), indirectly inhibiting CD8+ T cell proliferation. (E) Macrophages activate NF-κB via CCL2/CCR2 to stimulate IL-17 production in Th17 cells. Through CCL25/CCR9-mediated AKT/STAT3 activation, they promote the secretion of IL-10 and TGF-β by Tregs. (F) Macrophage-secreted CXCL9 recruits tissue-resident memory T cells (Trm) to survival niches rich in fatty acids. Furthermore, CXCL10/CXCR3 signaling drives the differentiation of CD8+ effector memory T cells (TEM) into terminally differentiated effector cells (TEMRA).
Macrophage-derived chemokines orchestrate thymocyte migration and selection
4.1
The thymus-seeding progenitors (TSPs) to the thymus are guided by stromal cell-derived CCL25, a crucial first step in T cell ontogeny that enables their differentiation into early T cell precursors (24). Following this entry, the migratory journey continues, as macrophage-derived CCL22 directs immature double-positive (CD4^+^ CD8^+^) T cells to the corticomedullary junction for subsequent pre-maturation events (2, 25). As illustrated in Figure 2, CX3CR1^+^ macrophages contribute to central tolerance by facilitating the negative selection of self-reactive thymocytes through antigen presentation (26). Beyond selection, these macrophages also promote T cell proliferation via the phagocytic clearance of apoptotic cells and bolster thymic epithelial cell (TEC)-T cell crosstalk through the secretion of CXCL16 and CCL2 (27, 28). The final stage of thymic development involves the egress of mature, naïve T cells; this process is triggered by CCL19 and CCL21, which activate Rac/RhoA signaling pathways to mediate their departure into the peripheral lymphatic system, thereby ensuring systemic distribution and establishing a pool of cells primed for immune surveillance (29–31).
Chemokine cues from macrophages direct CD4+ T cell differentiation
4.2
Following activation, naïve CD4^+^ and CD8^+^ T cells undergo differentiation into specialized effector subsets to orchestrate targeted immune responses. This critical process is potently modulated by chemokines secreted by macrophages (1, 32), as illustrated in Figure 2. As shown in Figure 2, the differentiation of CD4^+^ T cells into subsets like Treg, Th2, or Th17 is influenced by chemokines secreted by macrophages. For example, CXCL10 produced by M1-like macrophages promotes Th1 differentiation, while CCL17 from M2-like macrophages drives Th2 polarization.
As shown in Figure 2, macrophage-derived chemokines, like CCL2, CCL25, play a critical role in guiding the differentiation of CD4^+^ T cell subsets, including Treg, Th2, and Th17 cells. Signaling through the CCL2/CCR2 axis, for instance, can promote Th1 differentiation by upregulating the expression of key cytokines IFN-γ and IL-12 (33, 34). Furthermore, CCL2/CCR2 engagement activates downstream JAK/STAT signaling cascades, which regulate the master transcription factors T-bet and GATA3 to drive Th2 cell polarization (35, 36). Other macrophage-derived chemokines, including CXCL9, CXCL10, and CCL20, are also instrumental in stimulating effector T cell differentiation (37–39). The source of these signals is critical; notably, CXCL10 produced by pro-inflammatory M1 macrophages robustly drives Th1 polarization, thereby enhancing cell-mediated immunity (40). Conversely, M2 macrophages—often characterized by upregulated CD36 or histamine receptors—are a significant source of CCL17, a chemokine that promotes Th2 polarization and reinforces anti-inflammatory responses (41, 42).
The differentiation of Th17 cells is similarly regulated by chemokine cues. CCL2 facilitates Th17 commitment by modulating the local concentrations of TGF-β and IL-6, which in turn activate the PI3K/AKT/RORγt pathway (43, 44). The regulatory role of macrophage chemokines is complex and often indirect. Intriguingly, reduced secretion of CCL2, CCL3, CCL4, CCL5, and CXCL10 by M1 macrophages has been correlated with an expanded peripheral Treg population, underscoring the nuanced, often inhibitory role of these signals in regulatory T cell biology (45). In contrast, direct promotion of Treg development occurs via macrophage-derived CCL25, which activates AKT/STAT3/Foxp3 signaling to foster Treg differentiation while concurrently suppressing Fas/FasL-mediated apoptosis in contexts such as endometrial metaplasia (46). Additionally, CXCL1 supports Treg development through activation of the NF-κB/Foxp3 pathway (47).
Beyond these direct mechanisms, macrophage-derived chemokines can shape T cell fate indirectly by remodeling the local microenvironment. A key example is provided by islet macrophages, which express CXCL16 to scavenge oxidized low-density lipoproteins; this metabolic clearance activity indirectly creates a microenvironment conducive to CD8^+^ T cell differentiation (48).
Macrophage-derived chemokines regulate T cell immunometabolism
4.3
Cellular metabolism provides the fundamental biochemical foundation for all T cell functions, fueling processes from activation and proliferation to the execution of effector duties (49–54). The burgeoning field of immunometabolism has more recently identified chemokine signaling as a pivotal regulator of these metabolic programs (55, 56). A key mechanism involves the CXCL9/10–CXCR3 axis, which augments the cytotoxic potential of CD8^+^ T cells by elevating reactive oxygen species (ROS) and enhancing fatty acid oxidation (FAO) (57, 58). Paradoxically, this same signaling pathway can simultaneously suppress broader lipid metabolic pathways, illustrating the complex and nuanced nature of chemokine-mediated metabolic control. Beyond direct effects on T cells, macrophages can impose metabolic constraints through environmental remodeling. For instance, macrophage-derived CCL8 fosters a hypoxic niche by promoting aerobic glycolysis. This metabolic reprogramming of the TME functionally excludes cytotoxic T lymphocytes (CTLs) and suppresses their anti-tumor activity (5). Despite these advances, the mechanisms through which macrophage-secreted chemokines influence the metabolism of CD4^+^ T helper, Treg, and other lymphoid subsets represent a significant and promising frontier for future research.
Dual roles of macrophage-derived chemokines in T cell proliferation and exhaustion
4.4
The functional efficacy of T cells is intrinsically linked to their local abundance. In concert with survival cytokines, chemokines are critical mediators of T cell expansion and proliferation (59). For instance, macrophage-derived CCL24 acting through CCR3, or CCL2 signaling via CCR4, can activate the proliferation of naïve CD4^+^ T cells (60).
However, the role of macrophage-derived chemokines is not universally proliferative. In some contexts, they can suppress T cell numbers. For example, CCL18 secreted by M2 macrophages has been shown to indirectly reduce CD4^+^ T cell abundance in HIV-1 patients (61). A primary mechanism for this reduction is the induction of T cell exhaustion. As illustrated in Figure 2, chemokines, such as CXCL9 and CXCL10, play a crucial role in mediating T cell exhaustion, particularly in the TME, by upregulating exhaustion markers, including PD-1 on CD8^+^ T cells (62). This process is mediated through several distinct pathways. Firstly, certain chemokines directly upregulate exhaustion-associated proteins on T cells (63, 64). Macrophage-derived CCL23, CXCL9/10/11, and CCL5 upregulate CTLA-4 or PD-1 on CD8^+^ T cells via phosphorylation of glycogen synthase kinase 3β (GSK3β) or activation of the JAK/STAT pathway (56, 65, 66). Chemokines can alter the metabolic microenvironment to induce exhaustion (67). Overexpression of CXCL9 and CXCL10 by macrophages leads to the downregulation of OXPHOS and enhances indoleamine 2,3-dioxygenase 1 (IDO1)-mediated tryptophan degradation, thereby depleting nutrients essential for T cell proliferation (56). Furthermore, indirect mechanisms also play a key role. Stimulation through CCL22/CCR4 promotes the production of immunosuppressive factors such as IL-10 and TGF-β by Tregs; CCL8 establishes a hypoxic niche that indirectly suppresses CD8^+^ T cell expansion (5, 68).
Regulation of T cell cytokine secretion by macrophage-derived chemokine signaling
4.5
The secretion of effector molecules is a fundamental mechanism by which T cells execute their immune functions. A growing body of evidence indicates that macrophage-derived chemokines are potent regulators of this secretory activity. For example, signaling through CCL22 or the CXCL10/CXCR3 axis enhances IFN-γ production in T cells, a response that can exacerbate disease pathogenesis (40, 69). The underlying mechanisms involve the activation of distinct intracellular signaling pathways. In Th17 cells, CCL2 promotes IL-17 production by activating NF-κB. This IL-17 then creates a positive feedback loop by stimulating macrophages to secrete CCL20 through the NF-κB/MAPK/PI3K cascade, thereby facilitating the recruitment of more CCR6^+^ Th17 cells to the site (43, 44, 70). Conversely, a contrasting immunomodulatory effect is seen with CCL25, which activates the AKT/STAT3 pathway in Tregs, inducing the overexpression of anti-inflammatory cytokines like IL-10 and TGF-β (46). As depicted in Figure 2, the regulatory effects of chemokines can be influenced by macrophage polarization. M1-like macrophages typically enhance immune responses, while M2-like macrophages suppress them, contributing to immune evasion in the TME, suggesting a context-dependent mechanism of chemokine-mediated immunomodulation. Collectively, these findings demonstrate that macrophage-derived chemokines can polarize T cell responses in either a pro-inflammatory or anti-inflammatory direction. This dichotomous effect is likely determined by the phenotypic state of the macrophage source, highlighting a sophisticated, context-dependent mechanism of immunomodulation.
Macrophage-derived chemokines recruiting suppressive and effector T cells
4.6
T cell migration is a highly orchestrated process directed by chemokine gradients. These gradients activate GPCR signaling pathways, triggering cytoskeletal reorganization that enables targeted movement (71–75). This mechanism is fundamental not only for routine immunological surveillance but also serves as a key indicator of immune cell infiltration in pathological states (76). Macrophages regulate T cell recruitment by secreting a diverse array of chemokines that specifically attract distinct T cell subsets. For instance, CCL2 and CCL22 selectively recruit Tregs, while CXCL9 and CXCL10 are pivotal for attracting effector CD8^+^ T cells into tumors, thereby enhancing anti-tumor immunity.
Within the TME, this recruitment is often co-opted to promote immunosuppression. For example, TAM-derived CCL2 enhances the infiltration of Tregs, thereby reinforcing an immunosuppressive state (77). Similarly, macrophage-secreted CCL22 potently enhances Treg chemotaxis—a process modulated by type I interferon signaling or the nuclear translocation of p65/NF-κB (78–80). The expression level of CCL22 by M2 macrophages can predict CD4^+^ T cell abundance in the TME, and intriguingly, its expression is positively regulated by Treg-derived CXCL18, suggesting a cross-regulatory loop (81–83). Furthermore, CCL22 also facilitates the infiltration of CCR4^+^ Th2 cells in breast cancer, promoting a pro-tumorigenic immune contexture (84).
In contrast to these immunosuppressive roles, other macrophage-derived chemokines are critical for anti-tumor immunity. The CXCL9 and CXCL10 chemokines act synergistically to modulate CD8^+^ T cell chemotaxis and are essential for effective immune surveillance (85). Specifically, CXCL9 and CXCL10 derived from M1 macrophages play a critical role in promoting the migration of CD8^+^ T cells into tumors, a process indispensable for the successful killing of tumor cells (86–88). CXCL9 further cooperates with CCL5 to enhance the infiltration of CD8^+^ T cells into solid tumors (37).
CCL5 itself enhances the accumulation of CD8^+^ T cells via binding to its receptor, CCR5, thereby amplifying anti-tumor immune responses (89). These roles for CCL5 have been characterized not only in cancer but also in inflammatory diseases such as atherosclerosis and arthritis, and are frequently associated with the activation of inflammatory signaling pathways like NF-κB (90–93). Macrophage plasticity, exemplified by the ability of macrophages to switch between pro-inflammatory and anti-inflammatory states, introduces a dynamic regulatory mechanism that shapes the immune microenvironment. This plasticity enables macrophages to adapt their chemokine secretion patterns in response to different stimuli, influencing T cell differentiation, polarization, and functional outcomes in both tumorigenic and inflammatory contexts. Further studies indicated that repolarization from an M2-like to an M1-like phenotype could lead to a marked upregulation of CCL5 expression, resulting in the increased CD8^+^ T cell tissue infiltration (94).
Macrophage-derived chemokines orchestrate memory T cell differentiation and survival
4.7
The formation of memory T cells is essential for long-term adaptive immunity, enabling a rapid and potent response upon pathogen re-exposure. This process is critically dependent on chemokine-mediated recruitment (95). During the inflammatory phase, macrophages secrete chemokines that direct the migration of naïve T cells to secondary lymphoid organs, which are key sites for the initial differentiation of memory precursors (95). Additionally, monocyte-derived macrophages produce chemokines such as CCL5, CXCL9, and CXCL10, which are vital for the positioning, survival, and differentiation of tissue-resident memory T cell (Trm) precursors (96, 97).
M1 macrophages contribute to this process by producing CXCL9, which recruits CD8^+^ Trm cells to inflammatory sites enriched with fatty acids that support T cell survival (98). A fascinating positive feedback loop exists within the TME: CD8^+^ Trm cells promote CXCL9 transcription in macrophages via the IFN-γ/p-STAT1 axis, which in turn reinforces Trm retention and survival (99). This mechanism ensures swift responses to recurrent infections and enhances durable immune surveillance.
The therapeutic relevance of these pathways is increasingly apparent. Collectively, this chemokine-driven regulation of memory T cell recruitment, differentiation, and survival underscores the central role of macrophages in bridging innate and adaptive immunity, ensuring a potent and rapid response to recurrent challenges.
Cancer
5
Within the TME, the anti-tumor efficacy of CD8^+^ T lymphocytes is frequently compromised by both a decline in their absolute numbers and a state of functional impairment. This dysfunction inhibits their primary killing mechanisms, which include the perforin-granzyme/caspase cascade and IFN-γ-mediated pathways (100, 101). This immunosuppressed state is further exacerbated by Tregs, which secrete inhibitory molecules like IL-10 and TGF-β and upregulate checkpoint ligands that suppress the perforin-granzyme axis (100, 102, 103).
The polarization state of tumor-infiltrating macrophages is a critical determinant of this balance. In recent years, the divergent roles of pro-inflammatory M1 macrophages and immunosuppressive TAMs have garnered significant attention. Polarization towards the TAM phenotype, driven by tumor-derived signals and metabolic abnormalities, directly and indirectly exacerbates immunosuppression. In stark contrast, M1 macrophages typically counteract these effects and promote immune activation (104, 105). This section will focus on the mechanisms by which these two macrophage subsets differentially modulate the tumor-killing capacity of CD8^+^ T cells through their secretion of chemokines.
TAM-derived chemokines facilitate CD8+ T cell dysfunction
5.1
TAMs employ a repertoire of chemokines to enact immunosuppressive programs that ultimately inhibit CD8^+^ T cell function (Figure 3). A primary strategy involves the indirect suppression of CD8^+^ T cells via the recruitment of other inhibitory cells. Specifically, TAM-secreted CCL2, CCL22, and CXCL1 facilitate the infiltration of Tregs and MDSCs (23, 82, 106–108). The critical role of CCL22 is underscored by studies showing that anti-CCL22 antibodies significantly reduce intratumoral Treg accumulation (109). Beyond recruitment, TAMs can drive CTL exhaustion through neutrophil engagement. For instance, via NF-κB-mediated upregulation of CXCL5, TAMs stimulate neutrophil extracellular trap (NET) formation, which ultimately promotes a state of CD8^+^ T cell exhaustion (105).
Tumor-associated macrophages (TAMs) and M1 macrophages regulate CD8+ T cell-mediated tumor immunity through chemokine signaling. (CD8+ T cell activation). Pharmacological inhibition of C5aR enhances phosphorylation of ERK, AKT, and NF-κB, while suppressing C/EBPβ phosphorylation. Blockade of insulin-like growth factor (IGF) signaling upregulates STAT1, and overexpression of IRF8 enhances both NF-κB and STAT1 signaling. These interventions promote the secretion of CXCL9 and CXCL10 by M1 macrophages, which facilitate CD8+ T cell recruitment into the tumor microenvironment (TME). M1 macrophage-derived CCL20 binds to CCR6 and activates STAT3, stimulating CD8+ T cells to produce perforin. Granzyme then enters tumor cells through perforin pores, initiating caspase-mediated apoptosis. Tumor-derived Hsp70 (heat shock protein 70) triggers CCL3 and CCL5 secretion from M1-like macrophages via CD14 activation, and the released CCL3 and CCL5 stimulate IFN-γ production in CD8+ T cells through CCR5 signaling. (CD8+ T cell inhibition) TAM-derived CCL2, CCL22, and CXCL1 recruit myeloid-derived suppressor cells (MDSCs) or regulatory T cells (Tregs) — which secrete IL-10 and TGF-β — into the TME (dotted circle), thereby inhibiting CD8+ T cell function. CXCL5 stimulates neutrophil extracellular trap (NET) formation, leading to CD8+ T cell exhaustion. CCL5 upregulates the exhaustion marker PD-1 on CD8+ T cells, while CXCL8 downregulates the survival marker TIM-3.
Moreover, TAM-derived chemokines can directly impair CD8^+^ T cell function and promote tumor progression. In glioblastoma, a synergistic effect between TAM-secreted CCL8 and an IL-1β-induced hypoxic microenvironment collectively suppresses CD8^+^ T cell activity (5). In clear cell renal cell carcinoma, TAM-derived CCL5 directly upregulates the expression of the exhaustion marker PD-1 on CD8^+^ T cells (110). Furthermore, in advanced colorectal cancer, TAMs suppress TIM-3—a critical molecule for CD8^+^ T cell proliferation and effector function—through the IL-8 (CXCL8)/CXCR2 axis (111). These findings collectively underscore the pivotal role of TAM-derived chemokines in facilitating tumor immune escape.
M1 macrophage-derived chemokines enhance the tumor-killing capacity of CD8+ T cells
5.2
In direct opposition to TAMs, M1 macrophages secrete chemokines that are critical for initiating and sustaining effective anti-tumor T cell responses, making them attractive therapeutic targets (Figure 3). A key function of M1 macrophages is the production of CXCL9, CXCL10, and CXCL11, which facilitate the recruitment and intratumoral positioning of CXCR3-expressing CD8^+^ T cells. This process is fundamental for establishing a favorable microenvironment for immune checkpoint inhibitor therapies, such as anti-PD-(L)1 treatment (112, 113). In the context of atezolizumab plus bevacizumab combination therapy for advanced hepatocellular carcinoma, macrophage-derived CXCL10 promotes the preferential differentiation of CXCR3^+^ CD8^+^ effector memory T cells into highly cytotoxic PD1^−^ CD45RA^+^ terminal effector cells (TEMRA) (88).
The signaling pathways that drive the production of these chemokines are diverse and can be therapeutically modulated. For example, in ovarian cancer, pharmacological inhibition of the immunosuppressive C5a receptor enhances phosphorylation of ERK, AKT, and NF-κB while suppressing C/EBPβ phosphorylation, resulting in increased CXCL9 production (114). Similarly, blockade of insulin-like growth factor (IGF) signaling upregulates STAT1 transcription and inhibits STAT3 phosphorylation in M1 macrophages, thereby promoting the expression of CXCL9 and CXCL10 (115). In colorectal cancer liver metastases, microwave ablation combined with αPD-L1 therapy upregulates the transcription factor IRF8, which enhances both NF-κB and JAK-STAT1 signaling to stimulate CXCL9 production in M1 macrophages (116).
Beyond recruitment, M1 macrophage-derived chemokines can directly augment the cytotoxic arsenal of CD8^+^ T cells. Macrophage-secreted CCL20 binds to CCR6 (CD196) on CD8^+^ T cells, promoting STAT3-dependent transcription of perforin and enhancing its expression (117). Tumor-derived Hsp70 (heat shock protein 70) triggers CCL3 and CCL5 secretion from M1-like macrophages via CD14 activation, and the released CCL3 and CCL5 stimulate IFN-γ production in CD8^+^ T cells through CCR5 signaling (118).
This anti-tumor activity is reinforced by powerful cytokine feedback loops. Elevated levels of IFN-γ within the TME, often secreted by activated T cells, stimulate M1 macrophages to further increase their secretion of CXCL9 and CXCL10. This creates a positive feedback loop that amplifies T cell recruitment and enhances tumor clearance (119). The expression profile of these macrophage-derived chemokines has even been proposed as an independent prognostic biomarker in cancer, offering valuable insights for designing personalized treatment strategies (110, 120). Yi et al. (121) summarized the current understanding and therapeutic implications of targeting cytokine and chemokine signaling pathways in cancer. By exploring the roles of these molecules in tumor biology and the immune response, they highlighted the development of novel therapeutic agents aimed at modulating these pathways to combat cancer.
Conclusions
6
This review has elucidated the complex mechanisms through which macrophage-derived chemokines govern T cell biology, emphasizing their context-dependent roles in the TME. It highlighted how chemokines from immunosuppressive TAMs, such as CCL2 and CCL22, could mediate immune evasion by recruiting regulatory cell populations and promoting T cell exhaustion. In contrast, chemokines, such as CXCL9 and CXCL10 secreted by pro-inflammatory macrophages are pivotal in driving anti-tumor immunity by enhancing the infiltration and activation of cytotoxic CD8^+^ T cells. Consequently, targeting these chemokine-mediated regulatory mechanisms represents a notable direction for the development of novel cancer immunotherapies.
Although the traditional M1/M2 macrophage paradigm has provided useful insights, recent advances in single-cell transcriptomics have revealed a more complex and diverse spectrum of macrophage phenotypes. Previous research has demonstrated that macrophage populations exhibit significant heterogeneity, and their functional roles in immunity and tumor progression cannot be fully captured by the M1/M2 model. Therefore, understanding this macrophage diversity is crucial for refining therapeutic strategies.
Moreover, indirect mechanisms, including metabolic reprogramming pathways, such as IDO1-mediated tryptophan degradation, remain underexplored, while are critical in modulating T cell responses in the TME. To address these challenges, future research must utilize cutting-edge tools, including single-cell transcriptomics, spatial transcriptomics, and multiplexed imaging. These technologies, which have already provided valuable insights into macrophage diversity, are essential for deconvoluting specific macrophage subsets, mapping their interactions with T cells, and profiling their chemokine expression patterns in spatial contexts. Targeting these chemokine-mediated regulatory mechanisms represents a remarkably promising direction for the development of novel cancer immunotherapies. This comprehensive approach may pave the way for more precise and targeted immunotherapies that exploit chemokine networks, overcoming current limitations and employing the full potential of the immune system to combat cancer.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mantovani A Sica A Sozzani S Allavena P Vecchi A Locati M . The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. (2004) 12:677–86. doi: 10.1016/j.it.2004.09.015, PMID: 15530839 · doi ↗ · pubmed ↗
- 2Mc Cabe A Mac Namara KC . Macrophages: Key regulators of steady-state and demand-adapted hematopoiesis. Exp Hematol. (2016) 4:213–22. doi: 10.1016/j.exphem.2016.01.003, PMID: 26806720 PMC 4852701 · doi ↗ · pubmed ↗
- 3Norment AM Bevan MJ . Role of chemokines in thymocyte development. Semin Immunol. (2000) 5:445–55. doi: 10.1006/smim.2000.0261, PMID: 11085177 · doi ↗ · pubmed ↗
- 4Van Raemdonck K Umar S Palasiewicz K Volkov S Volin MV Arami S . CCL 21/CCR 7 signaling in macrophages promotes joint inflammation and Th 17-mediated osteoclast formation in rheumatoid arthritis. Cell Mol Life Sci. (2020) 7:1387–99. doi: 10.1007/s 00018-019-03235-w, PMID: 31342120 PMC 10040247 · doi ↗ · pubmed ↗
- 5Sattiraju A Kang S Giotti B Chen Z Marallano VJ Brusco C . Hypoxic niches attract and sequester tumor-associated macrophages and cytotoxic T cells and reprogram them for immunosuppression. Immunity. (2023) 8:1825–43.e 6. doi: 10.1016/j.immuni.2023.06.017, PMID: 37451265 PMC 10527169 · doi ↗ · pubmed ↗
- 6Liu N Wang X Steer CJ Song G . Micro RNA-206 promotes the recruitment of CD 8(+) T cells by driving M 1 polarisation of Kupffer cells. Gut. (2022) 8:1642–55. doi: 10.1136/gutjnl-2021-324170, PMID: 34706869 PMC 9279850 · doi ↗ · pubmed ↗
- 7Epelman S Lavine KJ Randolph GJ . Origin and functions of tissue macrophages. Immunity. (2014) 1:21–35. doi: 10.1016/j.immuni.2014.06.013, PMID: 25035951 PMC 4470379 · doi ↗ · pubmed ↗
- 8Locati M Curtale G Mantovani A . Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol. (2020) 15):123–47. doi: 10.1146/annurev-pathmechdis-012418-012718, PMID: 31530089 PMC 7176483 · doi ↗ · pubmed ↗