First record of winter pregnant males of two pipefish species in a Mediterranean coastal lagoon
Adrián Guerrero‐Gómez, Antonio Zamora‐López, Antonio Andrés Herrero‐Reyes, Francisco J. Oliva‐Paterna, Jorge Madrid‐Ruiz, Víctor M. Álvarez‐Navarro, Rocío Peñalver, Mar Torralva

TL;DR
For the first time, male pipefish were found to be pregnant in winter in a Mediterranean lagoon, possibly due to recent warming trends.
Contribution
First recorded winter breeding event for two pipefish species in response to thermal anomalies.
Findings
Pregnant males of Syngnathus abaster and Syngnathus typhle were observed in January 2025 in the Mar Menor lagoon.
The size structure of S. abaster in winter 2025 differed from colder winters, suggesting phenological shifts.
This indicates potential impacts of climate change on the reproductive dynamics of syngnathids.
Abstract
Syngnathus abaster Risso, 1827 and Syngnathus typhle L., 1758 are key components of fish assemblages in European transitional waters, with well‐documented reproductive cycles typically occurring from spring to autumn. However, recent warming trends in the western Mediterranean has raised questions about potential shifts in their breeding phenology. In January 2025, three pregnant S. abaster males and one S. typhle male were detected in the Mar Menor Coastal lagoon (western Mediterranean), representing the first recorded winter breeding event for both species. The size structure of S. abaster in winter 2025 also contrasted with colder winters, further supporting this interpretation. These findings may reflect a response to recent thermal anomalies and highlight the phenological plasticity of these species. To our knowledge, this is the first indication of potential phenological shifts in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1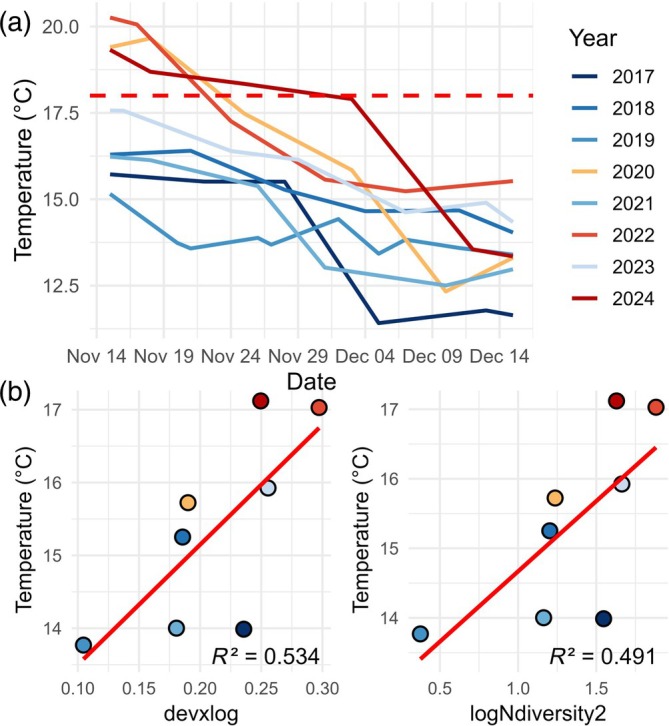 Figure 2
Figure 2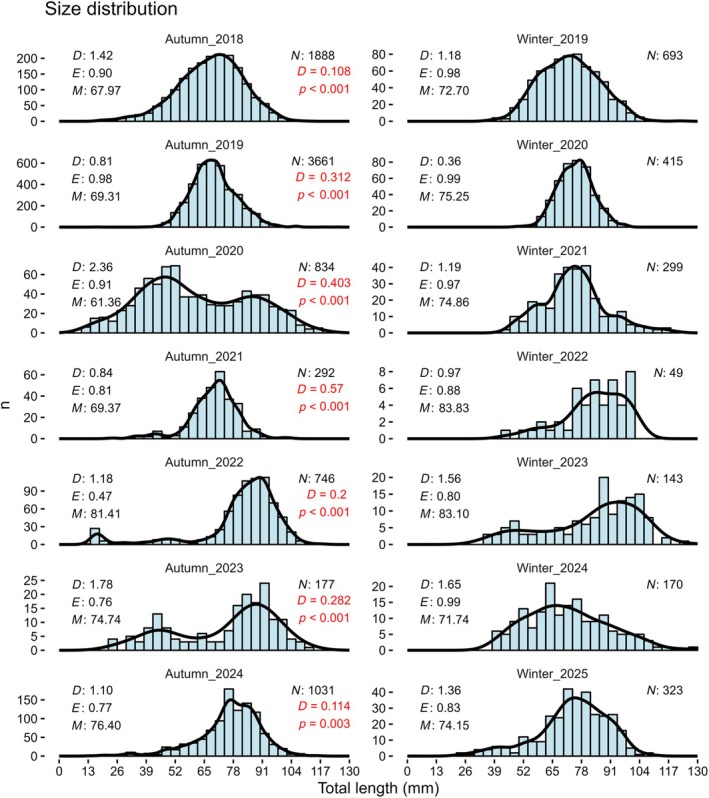 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquatic life and conservation · Marine Ecology and Invasive Species · Fish Biology and Ecology Studies
Syngnathids are a charismatic flagship group with high relevance in the fish assemblages of European coastal lagoons (Pérez‐Ruzafa et al., 2007; Scapin et al., 2018; Vincent et al., 2011). Among them, Syngnathus abaster Risso, 1827 and Syngnathus typhle L., 1758 are the most representative species, exhibiting well‐documented reproductive strategies (Monteiro & Vieira, 2017; Rodríguez et al., 2021; Vincent et al., 1995). Males of both species carry fertilized eggs in fully enclosed brood pouches on their abdominal side, receiving multiple partial clutches from different females (Vincent et al., 1995). Mating success is size‐dependent, with larger males of S. typhle and S. abaster producing more offspring and also initiating breeding earlier in the season (Cunha et al., 2015; Flanagan et al., 2017). These species typically reproduce from spring to autumn as water temperatures rise, with no reports of winter reproduction (Flanagan et al., 2017; Franzoi et al., 1993; Monteiro & Vieira, 2017; Rodríguez et al., 2021; Tsikliras et al., 2010). However, recent episodic increases in water temperature in the western Mediterranean have raised questions about potential changes in syngnathid reproductive cycles (Atalah et al., 2024). Although there is currently no direct evidence linking temperature shifts to modifications in the breeding phenology of these species, changes in the reproductive cycles of other syngnathid species in offshore waters have been previously reported (Kirby et al., 2006; Monteiro et al., 2017), raising the question of whether warming trends may influence their reproductive cycles. Here, we present data on pregnant males of S. abaster and S. typhle collected during an 8‐year monitoring programme in the Mar Menor coastal lagoon. In the case of the more abundant species, S. abaster, we also examined winter size structure in relation to temperature patterns, aiming to explore whether recent thermal anomalies may be associated with changes in reproductive timing. These findings contribute to our understanding of syngnathid responses to environmental variability and provide relevant insights for the management of transitional ecosystems.
Between January 2018 and January 2025, we conducted seasonal fish sampling in the shallow areas of the Mar Menor using a beach seine net (depth <1.5 m, 24 sampling campaigns; see Zamora‐López et al., 2023). In addition, between October 2023 and January 2025, we have carried out seasonal fish sampling in the deeper areas (between 1.5 and 6.5 m) using an epibenthic trawl net (n = 6) known as a ‘gánguil’. After collection, fish specimens were temporarily placed in plastic containers and photographed on a millimetre‐scaled plastic tray, allowing for total body length measurements using ImageJ software (Schneider et al., 2012). To investigate phenological changes, we analysed the size distributions of the most abundant species, S. abaster, in shallow areas, which offered the greatest number of temporal replicates. S. typhle was excluded from size analyses due to very low winter catch rates (ranging between 0 and 6 individuals). Size distribution comparisons were initially conducted visually using histograms of relative frequency patterns between autumn and the subsequent winter. Differences in distributions were statistically tested using Kolmogorov–Smirnov tests for each autumn–winter pair. We then performed a permutational analysis of variance (PERMANOVA; Anderson, 2017) on winter size data, nesting years into two groups based on whether average water temperatures during the period 15 November to 15 December exceeded the species' thermal threshold for mating (18°C; Fedonenko et al., 2016; Silva, Monteiro, Vieira, & Almada, 2006). This period was selected because it represents the last part of the year in which temperatures may still exceed the mating threshold, and it precedes the winter sampling dates, falling within the reported duration of embryo development in the brood pouch for this species (1 month; Silva, Monteiro, Almada, & Vieira, 2006). Additionally, we examined mean size, size evenness, size diversity, log‐transformed size diversity and log‐transformed size dispersion, following the metrics described by Quintana et al. (2016). These variables were correlated with mean temperature during the same pre‐winter period using Spearman's rank correlation. All statistical analyses were conducted using R software (Version 4.3.0) (R Core Team, 2025), and temperature data for the Mar Menor were obtained from CARM (2025).
Here, we report the detection of three pregnant S. abaster males and one S. typhle male in the waters of the Mar Menor coastal lagoon (western Mediterranean Sea) between 14 and 21 January 2025 (Figure 1). An individual of S. abaster (85 mm total length) was detected in shallow areas, while the other two (60 mm and 106 mm) and the S. typhle individual (223 mm) were found in deeper areas. These species breed from spring to autumn, covering at most February to October for S. abaster and April to October for S. typhle (Monteiro & Vieira, 2017; Takahashi et al., 2003; Tsikliras et al., 2010). In both species, males brood their embryos in a brood pouch for approximately 1 month, although low temperatures may extend this period (Ahnesjö, 2008; Silva, Monteiro, Almada, & Vieira, 2006). Therefore, the pregnant individuals observed in January likely mated during November–December, when water temperatures remained within the suitable range for reproduction. This pattern was especially marked in 2024, when suitable conditions persisted until early December (Figure 2), suggesting that anomalously warm autumns may lead to an extension of the species' known breeding season.
The size distribution of S. abaster in winter 2025 more closely resembled that of the preceding autumn, as indicated by the highest p value and a low D statistic in the Kolmogorov–Smirnov tests performed between seasonal pairs (Figure 3). Additionally, 2025 exhibited a unique transition in size structure between autumn and winter, characterized by a decrease in mean size and an increase in diversity and evenness, driven by the emergence of new juveniles. Other years with warm late autumns, such as 2022–2023, partially mirrored this pattern but did not show a reduction in mean size. This shift aligns with the unusually warm conditions recorded in late autumn 2024, which provided approximately 10 more days of suitable temperatures for S. abaster mating compared to previous warm years (2020–2021 and 2022–2023) (Figure 2). Furthermore, this pattern was supported by the PERMANOVA test, which revealed that size structure was significantly influenced by warmer years—those exceeding 18 °C after mid‐November—highlighting the potential role of elevated water temperatures during this period in triggering phenological changes in syngnathid species (Table 1). Moreover, average water temperature during these dates showed significant positive correlations with diversity and size deviation descriptors of the size structure, but only after applying a logarithmic transformation (Table 2 and Figure 2). This suggests that higher temperatures promote greater variability among smaller (juvenile) size classes. The log transformation dampens the influence of the largest individuals, highlighting changes within the juvenile spectrum and further supporting an extended breeding period for the species under such conditions.
On the other hand, between late 2019 and 2021, the Mar Menor coastal lagoon experienced successive critical eutrophic episodes, resulting in severe and widespread mass mortality events affecting aquatic fauna (Fernández‐Alías et al., 2022). These disturbances led to a marked decline in the abundance of S. abaster during the subsequent years (2021–2024; Guerrero‐Gómez et al., 2024; Zamora‐López et al., 2023), as evidenced by notably reduced detection rates and a capture frequency approximately 50% lower than in 2025 (Figure 3). The absence of brooding males during the winter of 2020–2021 and 2022–2023, despite thermal conditions relatively comparable to those in 2024–2025, may reflect diminished population densities post‐mortality, potentially limiting mating opportunities and reducing the probability of encounter. Moreover, these eutrophic stress events may have compromised the somatic condition of surviving individuals, thus could have diminished their reproductive readiness or capacity (Almeida et al., 2023).
The influence of temperature on syngnathid reproductive phenology is well documented, with numerous studies linking monthly increased temperatures to breeding activity (Monteiro et al., 2005; Silva et al., 2007; Silva, Monteiro, Vieira, & Almada, 2006; Taylan et al., 2018). There is also evidence suggesting that global warming is advancing and extending the breeding season of some offshore syngnathids, potentially altering sexual selection dynamics (Kirby et al., 2006; Monteiro et al., 2017). In addition, climate projections indicate a potential decline in Mediterranean syngnathid populations under continued warming scenarios (Monteiro et al., 2023). In line with this, laboratory experiments have also confirmed the potential thermal stress induced by climate change on syngnathid species (Costa et al., 2023). However, to our knowledge, this is the first documented evidence of phenological shifts in syngnathids from transitional waters associated with an unusual increase in temperature. These situations could become more frequent under ongoing global warming (Atalah et al., 2024). Confined transitional systems like the Mar Menor are particularly vulnerable to climate change (De Pascalis et al., 2012), making our observations highly relevant for understanding the potential response of this charismatic group in European transitional ecosystems. Further research should assess the hatching success and survival of juveniles from winter matings to better infer the long‐term implications of climate‐induced shifts in reproductive timing.
AUTHOR CONTRIBUTIONS
Adrián Guerrero‐Gómez: Writing – original draft, methodology, formal analysis, data curation, conceptualization. Antonio Zamora‐López: Writing – review and editing, methodology, investigation. Antonio Andrés Herrero‐Reyes: Writing – review and editing, methodology, investigation. Francisco José Oliva‐Paterna: Writing – review and editing, validation, supervision, project administration, investigation, funding acquisition, conceptualization. Jorge Madrid‐Ruiz: Methodology, investigation. Víctor Manuel Álvarez‐Navarro: Methodology, investigation. Rocío Peñalver: Methodology, investigation. Mar Torralva: Writing – review and editing, validation, supervision, project administration, methodology, investigation, funding acquisition, conceptualization.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahnesjö, I. (2008). Behavioural temperature preference in a brooding male pipefish Syngnathus typhle . Journal of Fish Biology, 73, 1039–1045.
- 2Almeida, D. , Cruz, A. , Llinares, C. , Torralva, M. , Lantero, E. , Fletcher, D. H. , & Oliva‐Paterna, F. J. (2023). Fish morphological and parasitological traits as ecological indicators of habitat quality in a Mediterranean coastal lagoon. Aquatic Conservation: Marine and Freshwater Ecosystems, 1229, 11–16.
- 3Anderson, M. J. (2017). Permutational multivariate analysis of variance (PERMANOVA). In N. Balakrishnan , T. Colton , B. Everitt , W. Piegorsch , F. Ruggeri , & J. L. Teugels (Eds.), Wiley Stats Ref: Statistics reference online (pp. 1–15). John Wiley & Sons, Inc.
- 4Atalah, J. , Ibañez, S. , Aixalà, L. , Barber, X. , & Sánchez‐Jerez, P. (2024). Marine heatwaves in the western Mediterranean: Considerations for coastal aquaculture adaptation. Aquaculture, 588, 740917.
- 5CARM . (2025). Canal Mar Menor. https://www.canalmarmenor.es/ (accessed Mar 12, 2025).
- 6Costa, A. B. , Correia, M. , Silva, G. , Lopes, A. F. , & Faria, A. M. (2023). Performance of the long‐snouted seahorse, Hippocampus guttulatus, under warming conditions. Frontiers in Marine Science, 10, 1–11.
- 7Cunha, M. , Berglund, A. , & Monteiro, N. M. (2015). The intrinsically dynamic nature of mating patterns and sexual selection. Environmental Biology of Fishes, 98, 1047–1058.
- 8De Pascalis, F. , Pérez‐Ruzafa, A. , Gilabert, J. , Marcos, C. , & Umgiesser, G. (2012). Climate change response of the mar Menor coastal lagoon (Spain) using a hydrodynamic finite element model. Estuarine, Coastal and Shelf Science, 114, 118–129.