Integrative taxonomy suggests hidden diversity within the fish genus Cyttopsis (Zeiformes, Parazenidae)
Rafael Bañón, Francisco Baldó, Juan Carlos Arronte, Alejandro de Carlos, José‐ Daniel Barreiro‐Vázquez, Ángel Sebastián Comesaña, David Barros‐García

TL;DR
This study reveals hidden diversity within the fish species Cyttopsis rosea using both molecular and morphological analyses.
Contribution
The study identifies cryptic diversity in Cyttopsis rosea through integrative taxonomy, suggesting multiple species instead of a single one.
Findings
Molecular analysis identified three molecular operational taxonomic units within Cyttopsis rosea.
Morphological differences in caudal peduncle length, vertebrae count, and lateral line scales suggest distinct species.
An updated taxonomic key for the Parazenidae family is provided based on new data and literature review.
Abstract
Nine specimens of Cyttopsis rosea (Zeiformes: Parazenidae) were collected during scientific surveys at three different locations in the northeast Atlantic. All nine specimens were included in the molecular analysis, adding new cytochrome c oxidase subunit I sequences to public databases. Of these, six specimens were retained for detailed morphological examination. Morphological measurements and counts agree with previous descriptions and confirm the identification of C. rosea. However, molecular species delimitation analyses suggested cryptic diversity, identifying three molecular operational taxonomic units, contrary to the current status of a single species with a worldwide distribution. The integration of classical and molecular taxonomy proved essential for accurately delimiting and characterizing this species, enhancing our understanding of its intraspecific morphological and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
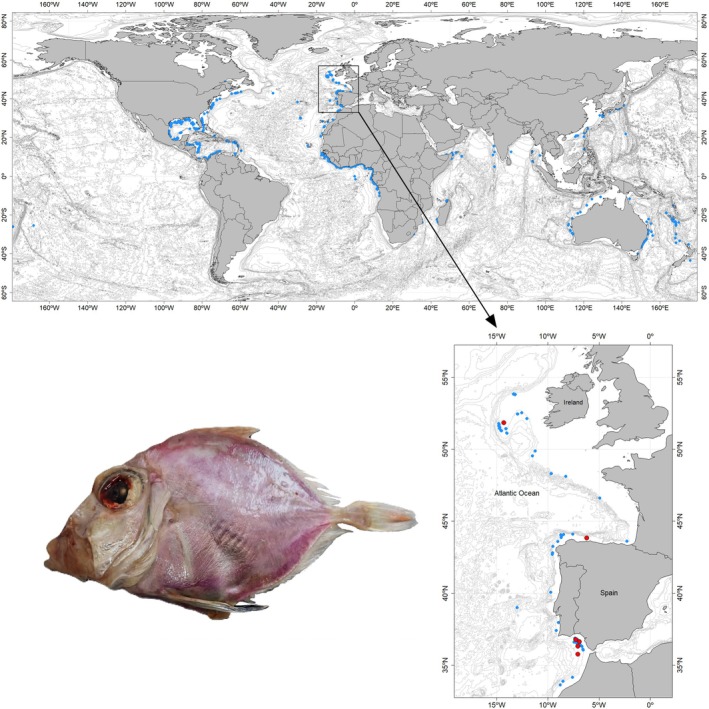 FIGURE 1
FIGURE 1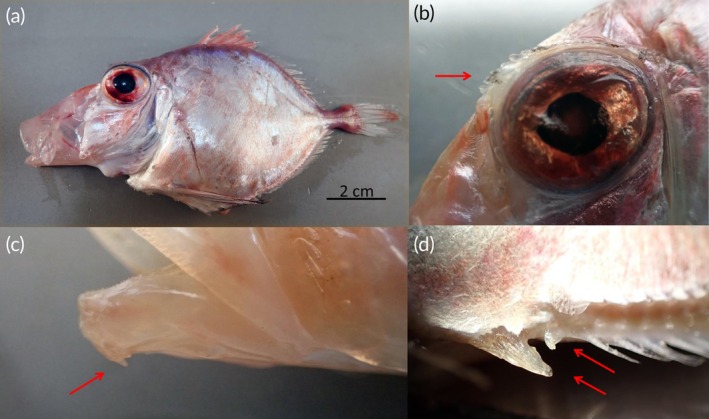 FIGURE 2
FIGURE 2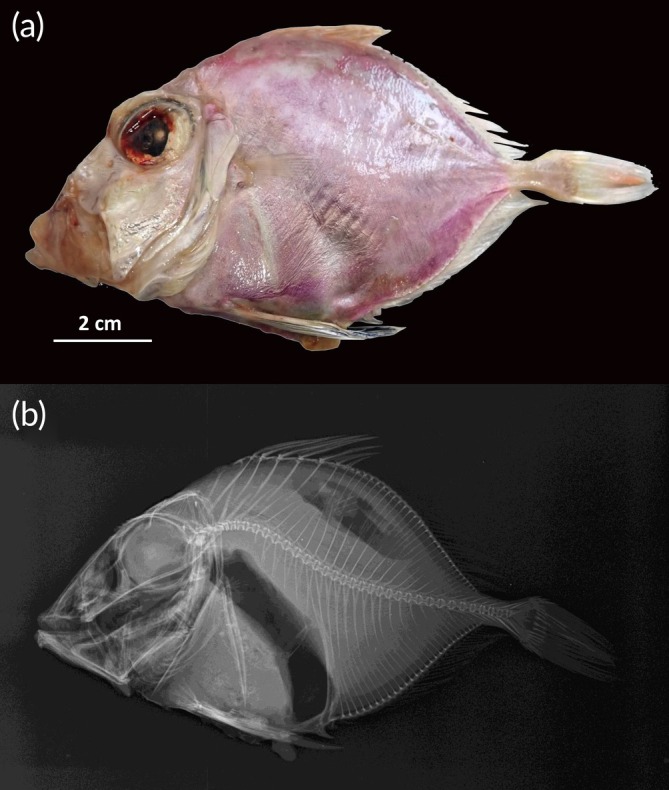 FIGURE 3
FIGURE 3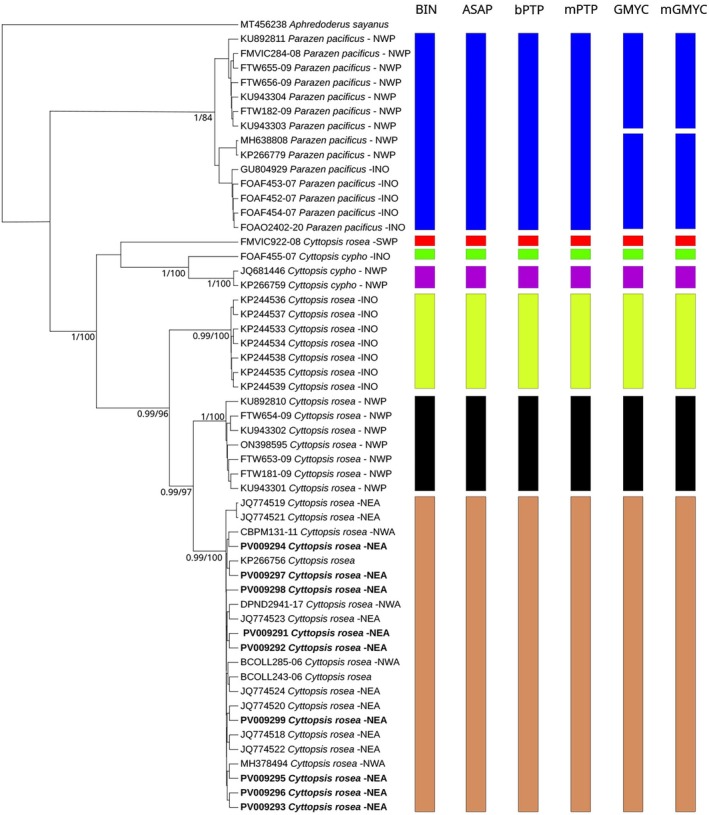 FIGURE 4
FIGURE 4| Character | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Total length (mm) | 137 | 136 | 77 | 135 | 91 | 150 |
| Standard length (mm) | 114 | 113 | 63 | 112 | 75 | 124 |
| As % | ||||||
| Head length | 38.2 | 42.5 | 44.4 | 39.3 | 42.4 | 41.5 |
| Snout length | 16.3 | 19.5 | 18.6 | 17.2 | 19.3 | 19.8 |
| Postorbital length | 6.6 | 7.5 | 8.3 | 6.5 | 7.6 | 7.8 |
| Eye diameter | 15.4 | 15.9 | 15.9 | 15.2 | 14.1 | 14 |
| Interorbital space | 8.3 | 8.6 | 8.4 | 8.6 | 8 | 9.1 |
| Predorsal length | 52.3 | 55.0 | 55.2 | 52.3 | 56.7 | 54 |
| Dorsal base fin length | 52.2 | 52.7 | 47.3 | 52.3 | 49.7 | 53.2 |
| Preanal length | 58.8 | 61.2 | 54.0 | 59.8 | 57.9 | 60.1 |
| Anal base fin length | 42.6 | 38.5 | 34.1 | 40.8 | 38.4 | 40.7 |
| Prepectoral length | 38.0 | 39.6 | 40.5 | 42.3 | 41.1 | 37.5 |
| Preventral length | 40.4 | 39.2 | 36.8 | 35.5 | 41.3 | 38.5 |
| Pelvic‐anal distance (LPA2) | 14.0 | 17.7 | 16.7 | 16.3 | 19.2 | 18.9 |
| Pectoral‐anal distance (LPA1) | 31.1 | 30.1 | 30.6 | 35.0 | 33.3 | 28.9 |
| Pectoral‐pelvic (LPP) | 20.2 | 20.1 | 19.0 | 19.6 | 19.1 | 18.5 |
| Dorsal‐pectoral (LDP1) | 26.3 | 26.1 | 23.8 | 24.4 | 26 | 26.6 |
| Dorsal‐pelvic (LDP2) | 49.4 | 50.6 | 46.3 | 51.2 | 50.7 | 45.8 |
| Pectoral fin length | 13.4 | 13.5 | 13.0 | 14.1 | 14.4 | 13.9 |
| Pelvic fin length | 34.5 | 37.2 | 46.2 | 34.1 | 40.9 | 31.2 |
| Body depth | 56.1 | 56.2 | 53.8 | 58.7 | 56.3 | 54.7 |
| Upper jaw length | 18.4 | 17.7 | 19.0 | 18.8 | 20 | 18.5 |
| Lower jaw length | 24.6 | 22.6 | 23.0 | 22.7 | 22.9 | 23.1 |
| Caudal peduncle minimum height | 5.7 | 5.3 | 6.0 | 5.5 | 6 | 5.2 |
| Caudal peduncle length | 6.6 | 7.7 | 9.0 | 7.6 | 7.7 | 8.3 |
| Length 3° dorsal spine (longest) | 17.5 | 18.9 | 14.4 | 15.2 | 16 | 16.1 |
| Length 1° anal spine | 3.9 | 4.6 | 2.7 | 3.3 | 3.7 | 3.5 |
| Distance between pelvic fin bases | 10.2 | 11.3 | 12.9 | 12.5 | 11.5 | 10.3 |
| Meristic | ||||||
| Dorsal fin rays | VII + 27 | VIII+27 | VII + 29 | VII + 28 | VII + 29 | VII + 29 |
| Anal fin rays | I + 30 | II + 30 | I + 30 | II + 29 | I + 30 | II + 29 |
| Pectoral fin rays | 14 | 14 | 14 | 14 | 14 | 14 |
| Ventral fin rays | 9 | 9 | 9 | 9 | 9 | 9 |
| Developed gillraker | 1 + 10 | 2 + 10 | 1 + 9 | 2 + 10 | 2 + 9 | 2 + 8 |
| Branchiostegal rays | 7 | 7 | 7 | 7 | 7 | 7 |
| Vertebrae | 32 | 32 | 32 | 32 | 32 | 32 |
| Scales in lateral line | 66 | 72 | 69 | 70 | 69 | 70 |
| Soft dorsal‐fin base scutes | 27 | 25 | 28 | 26 | 27 | 28 |
| Soft anal‐fin base scutes | 28 | 30 | 27 | 27 | 28 | 27 |
| Caudal fin rays | 4 + 13 + 4 | 4 + 13 + 4 | 3 + 13 + 4 | 3 + 13 + 3 | 3 + 13 + 4 | 3 + 13 + 3 |
| Character | Atlantic | Pacific |
|---|---|---|
| Total length (mm) | ||
| Standard length (mm) | 63–124 | 56.5–186.6 |
| As % | ||
| Head length | 37–44.4 | 38.8–47.6 |
| Snout length | 16.3–25.6 | 17.8–23.3 |
| Postorbital length | 6.5–8.3 | – |
| Eye diameter | 13–15.9 | 14.7–19.1 |
| Interorbital space | 6.6–9.1 | 5.8–8.0 |
| Predorsal length | 52.3–59.5 | 50.4–59.8 |
| Dorsal base fin length | 47.3–53.2 | 48.2–55.4 |
| Preanal length | 50.8–67.8 | 50.3–61.5 |
| Anal base fin length | 34.1–42.6 | 36.5–42 |
| Prepectoral length | 37.5–42.3 | – |
| Preventral length | 35.5–41.3 | – |
| Pelvic‐anal distance (LPA2) | 14–19.2 | 11–23.7 |
| Pectoral‐anal distance (LPA1) | 28.9–35 | 24.6–33.5 |
| Pectoral‐pelvic distance (LPP) | 18.5–20.2 | 19.3–26.4 |
| Dorsal‐pectoral distance (LDP1) | 23.8–26.6 | 23.4–29.2 |
| Dorsal‐pelvic distance (LDP2) | 45.8–51.2 | 44.7–58 |
| Pectoral fin length | 13–14.4 | 12.4–17.6 |
| Pelvic fin length | 31.2–46.2 | 28.4–48.8 |
| Body depth | 47.4–60.7 | 47.7–60.8 |
| Upper jaw length | 17.7–20.5 | 16.4–22.7 |
| Lower jaw length | 22.6–24.6 | 22.1–26.9 |
| Caudal peduncle minimum height | 5.2–6 | 4.3–5.9 |
| Caudal peduncle length | 6.6–9 | 10.8–13.7 |
| L 3° dorsal spine (longest) distance between pelvic fin bases | 14.4–18.9 | 12–17.4 |
| Meristic | ||
| Dorsal fin rays | VII‐IX + 27–29 | VII + 27–30 |
| Anal fin rays | I‐II + 29–30 | I + 28–31 |
| Pectoral fin rays | 13–14 | 14–15 |
| Ventral fin rays | 9 | 9 |
| Developed gillraker | 1–2 + 8–10 | 1–2 + 8–11 |
| Branchiostegal rays | 7 | – |
| Vertebrae | 32 | 30–31 |
| Scales in lateral line | 66–72 | 73–82 |
| Soft dorsal‐fin base scutes | 25–28 | 27–29 |
| Soft anal‐fin base scutes | 27–30 | 27–30 |
| Caudal fin rays | 3–4 + 13 + 3–4 | – |
- —European Maritime, Fisheries and Aquaculture Fund10.13039/100032853
- —Fundação para a Ciência e a Tecnologia10.13039/501100001871
- —Xunta de Galicia10.13039/501100010801
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIdentification and Quantification in Food · Fish biology, ecology, and behavior · Fish Biology and Ecology Studies
INTRODUCTION
1
Zeiformes is a primarily benthopelagic order of acanthomorph fishes that live and feed near the seafloor at depths of 50–1000 m. They are distributed globally in tropical and temperate regions, with some species having near global ranges, while others are regional endemics (Peters et al., 2024). The family Parazenidae comprises three genera and four species of zeiform fishes distributed worldwide in tropical and temperate waters on the continental shelf and slope of the Atlantic, Pacific and Indian Oceans. The family was reviewed by Heemstra (1980) and later by Tyler et al. (2003) and Grande et al. (2018) based on molecular and morphological characters, confirming the monophyly of the family. The parazenid genus Cyttopsis Gill, 1862 contains two valid species, the Rosy dory Cyttopsis rosea (Lowe, 1843), found in the Atlantic and Indo‐West Pacific, and the little dory Cyttopsis cypho (Fowler, 1934), which is restricted to the eastern Indian and western Pacific Oceans (Mizumachi et al., 2022).
Cyttopsis rosea is a demersal species found between 150 and 962 m depth (Bañón et al., 1997; Maurin & Quéro, 1981). It was originally described by Lowe (1843) as Zeus roseus on the basis of two specimens from the Atlantic Madeira Islands. It is currently distributed worldwide, occurring in the western Atlantic from Venezuela to Canada, in the eastern Atlantic from the British Isles to Gabon, in the western Indian Ocean from Somalia to South Africa (KwaZulu‐Natal), the Maldives and India, and in the western Pacific off Japan, eastern and western Australia, New Caledonia and New Zealand (Heemstra, 2022; Munroe et al., 2015). Cyttopsis itea Jordan & Fowler, 1902 and Paracyttopsis scutatus Gilchrist & von Bonde, 1924 are considered synonyms of C. rosea (Heemstra, 1980).
In the eastern Atlantic, C. rosea has been found to be expanding its range northwards into European Atlantic waters as a result of warming waters associated with climate change and the tropicalization phenomenon. It was reported in Portugal in 1963 and has expanded northwards to 43°N in 1968, 44°N in 1969, 49°N in 1987 and 52°N in 1988 (Quéro et al., 2023). According to the Red List of the International Union for Conservation of Nature (IUCN), C. rosea is considered globally to be of Least Concern (Munroe et al., 2015).
Molecular taxonomy, including the analysis of DNA sequence data, provides a strong assessment of genetic relationships and has revealed inconsistencies in taxa defined by traditional phenotypic methods. The analysis of DNA barcode sequences using various clustering techniques offers an efficient method for recognizing putative species (operational taxonomic units) (Kekkonen & Hebert, 2014). Integrated morphological and molecular analyses allow more accurate identification of fish species, facilitating the description of new species and the discovery of synonymous or cryptic species (Bañón et al., 2024). Molecular tools have been successfully used to highlight some of these findings in zeiform fishes (Costa et al., 2012; Kai & Tashiro, 2019; Ward et al., 2008).
The aim of this research was to add new morphological and molecular data from C. rosea caught in northeast Atlantic waters to the global dataset, and to review the available taxonomic and biogeographical knowledge to document the possible occurrence of cryptic diversity.
MATERIALS AND METHODS
2
Ethics
2.1
The species described in this paper are not listed as threatened on either the IUCN Red List or any national, regional or local schedules of protected fauna. All required permits for collection were obtained from the Instituto Español de Oceanografía (IEO), Consejo Superior de Investigaciones Científicas (CSIC).
Specimen collection and processing
2.2
Nine specimens of C. roseus were collected through trawling during several research surveys conducted in various areas of the North Atlantic Ocean (Figure 1): (i) Porcupine 2022, conducted off the west coast of Ireland; (ii) Demersales 2022, in Galician waters and the Cantabrian Sea, and (iii) ARSA 2022 and ARSA 2024, conducted in the Gulf of Cadiz (southern Spain). Specimens were preliminarily identified on board, and entire specimens or tissues were preserved frozen (−28°C). Following tissue extraction for molecular analysis, six voucher specimens were preserved frozen and deposited in the fish collection at the Museo Luis Iglesias de Ciencias Naturais in Santiago de Compostela (MHN USC), under reference numbers MHN USC 25228‐1 to 25228‐6. Additional tissue samples from three specimens collected in the Gulf of Cadiz were used for DNA species delimitation analyses and are deposited at the Universidade de Vigo.
Sampling location of Cyttopsis rosea specimens (red dots). The global distribution of the species is also shown (GBIF.org, 2025; OBIS, 2025), including historical records from the Spanish research bottom trawl surveys in the Porcupine Bank, the north coast of Spain and the Gulf of Cadiz (light blue dots).
Morphological analyses
2.3
Morphological analysis was carried out according to Mizumachi et al. (2022). Measurements were taken to the nearest 0.1 mm, using a digital calliper, except for total length (L T) and standard length (L S), which were measured to the lower millimetre using an ichthyometer. A comprehensive review of morphological characters reported in the ichthyological literature was compiled to search for geographic differences in the species. Vertebrate counts were determined from radiographs taken using mammography computed radiography equipment (Fujifilm Capsula XL II) with exposure factors of 44 kVp × 35 mAs.
DNA extraction, PCR amplification and sequencing
2.4
DNA extraction was performed from muscle tissue samples preserved in 70% ethanol at −20°C using the E.Z.N.A. Tissue DNA Kit (Omega Bio‐Tek) according to the manufacturer's instructions. The DNA barcode region of the mitochondrial gene encoding the enzyme cytochrome c oxidase I was amplified using the polymerase chain reaction. For this purpose, the Horse‐Power Taq DNA Polymerase Master Mix (Canvax) was employed in combination with the C_FishF1t1‐C_FishR1t1 universal primer cocktail for fish DNA barcoding (Ivanova et al., 2007). The temperature regime was as follows: 98°C for 5 min, 35 cycles of 94°C for 35 s, 52°C for 35 s and 72°C for 50 s, and 72°C for 7 min. Sanger cycling sequencing reactions were performed with BigDye Terminator v3.1, and the resulting products resolved in a SeqStudio Genetic Analyser at the Centro de Apoyo Científico y Tecnológico a la Investigación facilities (University of Vigo). Trace files were interpreted and DNA alignment was conducted using MEGA version 11 (Tamura et al., 2021).
DNA sequences, specimen photographs and associated metadata have been deposited in the Barcode of Life Database (BOLD Systems; www.boldsystems.org) as part of the projects titled ‘Porcupine Bank Fish’ (code PORF, process ID: PORF357‐25) and ‘Unusual Atlantic Fishes’ (code UNAFI, process ID UNAFI 014‐25 to UNAFI 021‐25). Additionally, the barcode sequences have been submitted to GenBank (https://www.ncbi.nlm.nih.gov/genbank/) under accession numbers PV009291‐PV009299.
Phylogenetic analyses
2.5
Publicly available cytochrome c oxidase subunit I (COI) sequences previously assigned to the family Parazenidae were retrieved from BOLD‐Systems and GenBank repositories (February 2025). After validation, the final dataset comprised 54 COI sequences belonging to family Parazenidae plus one outgroup species (Piraten perch Aphredoderus sayanus [Gilliams, 1824]). To apply delineation methods based on phylogenetic tree topologies, trees were constructed using both Bayesian inference for generalized mixed yule coalescent (GMYC) and maximum likelihood (ML) for Bayesian Poisson tree processes (bPTPs). To this end, an optimal partition strategy and a substitution model were determined using PartitionFinder v1.1.1 (Lanfear et al., 2012) and jModelTest2 (Darriba et al., 2012; Guindon & Gascuel, 2003), respectively. Single partition and general time reversible (GTR) with invariant sites (+I) were therefore selected for further analyses. BEAST V2.5.2 was used for Bayesian inference, with two runs of 10 million generations with four independent chains and sampling every 1000 generations. Convergence of the analyses was addressed with TRACER.1.7.0 (Rambaut et al., 2018). The final consensus tree was obtained using a posterior probability of 0.9, discarding the first 25% of sampled trees, and generated in TreeAnnotator v2.4.5. The tree was then visualized with FigTree v.1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/).
The ML analysis was conducted through the CIPRES portal using the IQ‐tree tool on XSEDE (version 2.4.0), with 1000 rapid bootstrap replicates to provide statistical support for the nodes (Miller et al., 2010). For the PTP analysis, the tree with the highest likelihood from the ML analysis was selected as the input.
Species delimitation
2.6
Several DNA delimitation analyses were conducted to assess the taxonomic status of the family Parazenidae. First, the distance‐based Barcode Index Number (BIN) was retrieved from BOLD Systems (https://www.boldsystems.org/) (Ratnasingham & Hebert, 2013). The second distance‐based method applied was the Assemble Species by Automatic Partitioning (ASAP) (Puillandre et al., 2021), which is available via a web server (https://bioinfo.mnhn.fr/abi/public/asap/). In this method, p‐distance was calculated for the COI alignment without an outgroup, and the optimal partition was selected based on the ASAP score and threshold distance.
Four tree‐based delimitation approaches were used for analysis. First, two modifications of the PTP algorithm (Zhang et al., 2013) were applied: bPTP, which incorporates Bayesian support values to the delimited species, and mPTP, which uses a multi‐rate criterion to account for varying levels of intraspecific genetic diversity (Kapli et al., 2017). These analyses were conducted on web servers: bPTP was run at (https://species.h-its.org/ptp/) and mPTP at (https://mptp.h-its.org/#/tree). In both cases, the ML tree was used as input, after removing the outgroup, to achieve optimized results (Zhang et al., 2013).
Next, the GMYC model was applied, which identifies the time threshold defined by coalescent processes on ultrametric trees (Fujisawa & Barraclough, 2013; Pons et al., 2006). A variation of this model, the multi‐rate GMYC (mGMYC), was also used, which assumes multiple independent coalescent processes across the tree's topology (Monaghan et al., 2009). Both analyses were performed using the R package ‘splits,’ with the ‘gmyc’ function, specifying ‘method = single’ for GMYC and ‘method = multiple’ for mGMYC (Fujisawa & Barraclough, 2013).
RESULTS
3
Systematic account
3.1
Cyttopsis rosea (Lowe, 1843)
3.1.1
Zeus roseus Lowe, 1843: 85, off Madeira, northeastern Atlantic, Syntypes: (2) BMNH 1852.9.13.104; Cyttus roseus Günther, 1860: 396 (description); Cyttus roseus Vaillant, 1888: 349 (description); Cyttopsis roseus Goode & Bean, 1895: 227 (description); Cyttus (Cyttopsis) roseus Cadenat, 1953: 1079 (description); Cyttus roseus Poll, 1954: 19 (description); Cyttus roseus Furnestin et al., 1958: 427 (description); Cyttopsis roseus Heemstra, 1980: 5 (description); Cyttopsis roseus Heemstra, 1986: 436 (description. key); Cyttopsis rosea Roa‐Varón et al., 2003: 11 (description); Heemstra, 2016: 2227 (description, key).
Material examined
3.1.2
MHN USC 25228‐1, 137 mm TL, 9 October 2022, Cantabrian Sea; 43.843°N, 6.211°W, 407 m depth; MHN USC 25228‐2, 136 mm TL, 9 October 2022, Cantabrian Sea; 43.843°N, 6.211°W, 407 m depth; MHN USC 25228‐3, 77 mm TL, 10 November 2024, Gulf of Cádiz, south Spain; 36.321°N, 7.071°W, 519 m depth; MHN USC 25228‐4, 135 mm TL, 13 September 2024, Porcupine Bank, western Ireland; 51.855°N, 14.309°W, 356 m depth. MHN USC 25228‐5, 91 mm TL, 5 November 2022, Gulf of Cadiz, south Spain; 36.638°N, 6.943°W, 409 m depth. MHN USC 25228‐6, 150 mm TL, 6 November 2022, Gulf of Cadiz, south Spain; 36.795°N, 7.278°W, 475 m depth.
Description
3.1.3
Body oval and compressed (Figures 2a and 3), body depth greater than head length and contained 1.7–1.9 times in the standard length; head large, contained 2.3 to 2.6 times in the standard length; snout slightly longer than the eye diameter; eye very large, its diameter 2.5 to 3 times in the head; edge of bony orbit with small spines anteriorly (Figure 2b); mouth large, oblique with a protruding upper jaw; with dentaries with two small spines at the symphysis below (Figure 2c); interorbital width 1.5–1.9 in eye diameter; pectoral fin inserted before the spinous dorsal fin, its length much shorter than head length; pelvic fin long, inserted nearly in the vertical from the insertion of the pectorals, its length 1–1.3 times in the head length; first anal spine flattened and immovable (Figure 2d); ventral midline between pelvic and anal fins broad, flattened, with large spiny bucklers; a row of low bony ridges on each side at the base of soft dorsal and anal fins; 66–72 scales in lateral line; caudal peduncle length 4.9–5.8 times in the head length, number of vertebrae 32. The key morphometric and meristic characteristics are provided in Table 1. A revision of the morphological characters documented in the ichthyological literature for the Atlantic and Pacific specimens is presented in Table 2.
Cyttopsis rosea from the northeast Atlantic. Arrows show the described character: (a) whole specimen MHN USC 25228‐4, 135 mm L T showing the protusible mouth, (b) detail of eye with supraorbital crest, (c) small spine on the lower jaw and (d) anal fin with two anterior spines.
Specimen of Cyttopsis rosea Museo Luis Iglesias de Ciencias Naturais in Santiago de Compostela MHN USC 25228‐6, 150 mm L T, caught in the Bay of Cádiz (northeast Atlantic): (a) lateral view of the specimen and (b) radiograph of the specimen.
Species delimitation
3.1.4
In the resulting consensus phylogenetic tree, all sequences formed monophyletic groups based on their morphological identification with high statistical support in both posterior probability and bootstrap, with the exception of FMVIC922‐08 (C. rosea), from New Zealand, whose position in the tree remains unresolved (Figure 4). Species delimitation analyses showed a remarkable consistency among them. Four out of six, including distance (BIN, ASAP) and phylogenetic tree (bPTP, mPTP) methods, gave exactly the same results with a total of seven clades, while both GMYC analyses agreed on eight. For C. rosea, all delimitation analyses found the same three clusters in its monophyletic clade with remarkable geographic patterns. The first clade comprises seven sequences (KP244533‐39) from the Indian Ocean; the second includes another seven sequences from northwestern Pacific Ocean (China, Taiwan, Japan) while the largest clade contained 21 sequences representing northeastern and northwestern Atlantic Ocean and two sequences (KP266756, BCOLL243‐06) of unconfirmed origin (Figure 4). The three sequences belonging to C. cypho were divided in two clusters for all the delimitation analyses, the first with JQ681446 and KP266759 from the northwestern Pacific Ocean (South China) and the second with FOAF455‐07 from the Indian Ocean (Western Australia). The monophyletic group with Parazen Parazen pacificus (Kamohara, 1935) sequences was statistically well supported, but the two subgroups into which it is split were not. Four out of six delimitation analyses considered P. pacificus a single species, while both GMYC split it in two. The first included sequences from the northwestern Pacific Ocean, Taiwan (FTW655‐09 and FTW656‐09) and China (FMVIC284‐08), while the second comprised sequences from Indian Ocean, Western Australia (FOAF452‐07; FOAF454‐07), South Africa (GU804929) and South China (MH638808; KP266779).
Consensus phylogenetic tree of 54 COI sequences belonging to the family Parazenidae plus one outgroup species (Aphredoderus sayanus). Node numbers represent posterior probability (>0.9) and bootstrap (>75). Sequences obtained in the present study are highlighted in bold. Each sequence includes its GenBank/Bold unique label, species identification and abbreviation of geographical origin: NWP, northwestern Pacific; SWP, southwestern Pacific; INO, Indian Ocean; NEA, northeastern Atlantic; NWA, northwestern Atlantic. The species delimitation results include distance‐based (Barcode Index Number [BIN] and Assemble Species by Automatic Partitioning [ASAP]) and tree‐based (Bayesian Poisson tree processes [bPTP], multi‐rate Poisson tree processes [mPTP], generalized mixed yule‐coalescent [GMYC] and multiple threshold generalized mixed yule‐coalescent [mGMYC]) methods.
Key to Parazenidae species
3.2
Adapted from Tyler et al. (2003) and Mizumachi et al. (2022); taxonomic characters of C. rosea limited to those of the Atlantic Ocean.
1a. Body elongated, body depth 2.2–3 times in L S; 8 dorsal‐fin spines; 7 pelvic‐fin rays; pelvic‐fin positioned about midway between pectoral‐fin base and anus; pectoral‐fin rays usually 16, rarely 15; procurrent caudal‐fin rays 7 or 8; scales spinoid; no large buckler scales along ventral midline; low sheath of scales along soft dorsal‐ and anal‐fin bases…………… Subfamily Parazeninae, Parazen pacificus (Kamohara, 1935).
1b. Body oval, body depth 1.3–2.5 times in L S; dorsal‐fin spines usually 6 or 7, rarely 8; pelvic‐fin rays 9; pelvic‐fin positioned under or slightly in front of pectoral‐fin base; pectoral‐fin rays usually 13 or 14, rarely 15; procurrent caudal‐fin rays 3 or 4; scales cycloid; large buckler scales along ventral midline; no sheath of scales along soft dorsal‐ and anal‐fin bases ………………………Subfamily Cyttopsinae (2).
2a. Dorsal‐fin spines usually 6, sometimes 7; pectoral‐fin rays usually 13, sometimes 14; large buckler scales along ventral midline of isthmus forming a continuous, sharp‐edged median spiny keel (nearly all buckler scales strongly overlapping); honeycomb pattern on bones on top of head visible externally…………………………… Genus Stethopristes Gilbert, 1905, Stethopristes eos (Gilbert, 1905).
2b. Dorsal‐fin spines usually 7, sometimes 8 or 9; pectoral‐fin rays usually 14, sometimes 13 or 15; large buckler scales along ventral midline of isthmus forming a partially separated spiny keel (a few bucklers not overlapping, especially first, on isthmus, and second, between pelvic‐fin bases); no honey comb pattern on bones on top of head, opercles, and lachrymal‐infraorbitals, only long ridges……………………………. Genus Cyttopsis Gill, 1862 (3).
3a. A faint dark lateral spot posteriorly on the body; 55–64 scales in lateral line; interspace between spines on 4th and 5th abdominal narrower; orbit diameter 13.6%–16.6% L S; interorbital width 5.8%–6.6% L S; snout length 21.1%–27.4% L S; mandible length 22.6%–25.6% …………………Cyttopsis cypho (Fowler, 1934).
3b. Without dark lateral spot posteriorly on the body; 66–72 scales in lateral line; interspace between spines on 4th and 5th abdominal scutes wide; orbit diameter 13%–15.9% L S; interorbital width 6.6–9.1% L S; snout length 16.3%–23.3% of L S; mandible length 22.1%–26.9% of L S………… Cyttopsis rosea (Lowe, 1843) Atlantic Ocean.
DISCUSSION
4
Fish taxonomy is far from complete. The taxonomic trend throughout the history of ichthyology has been to reduce the number of species within a given taxon through successive taxonomic revisions (synonymous species). As a result, many fish species are now widely distributed and morphological differences, when known, are attributed to natural intraspecific variation. In contrast, the use of molecular techniques seems to indicate the presence of a large hidden diversity and putative cryptic species with a more reduced distribution area (Bañón et al., 2024; Pavan‐Kumar et al., 2016).
Morphological and meristic characteristics of C. rosea specimens examined were consistent with previous descriptions of the species. However, differences were found between the Atlantic and Pacific specimens. The most remarkable are the caudal peduncle, which is longer in the Pacific than in the Atlantic specimens (10.8%–13.7% L S vs. 6.6%–9% L S), the number of vertebrae (32 in the Atlantic vs. 30–31 in the Pacific) and the lower number of scales in the lateral line of the Atlantic (66–72) compared to the Pacific specimens (73–82). According to Heemstra (1980), the number of lateral line scales in C. rosea ranges from 73 to 84, in the same range as the 73–82 found in the Pacific specimens by Mizumachi et al. (2022). However, Heemstra's counts are based on the examination of six specimens from three different geographical areas, Japan, South Africa and the Northeast Atlantic (Heemstra, 1980), so it is not possible to establish geographical differences. Vaillant (1888) reported 53 scales in the lateral line of an Atlantic specimen. Although this number is anomalous and outside the known range of the species, it could support the lower number of scales in Atlantic specimens.
More striking is the presence of two small spines on the lower jaw. Although these spines are shown in radiographs and drawings of the species (Peters et al., 2024; Rosen, 1984; Tyler et al., 2003), to our knowledge this feature has only been recorded by Jordan and Fowler (1902) in the description of C. itea, a synonym of C. rosea. The taxonomic significance of this character needs to be reviewed.
Single‐gene species delimitation methods have been widely used for biodiversity studies (Ramirez et al., 2023). However, taking into account their different assumptions and principles, they can be affected differently by the data set features (Magoga et al., 2021). For instance, the barcoding gap estimation can vary due to a higher number of sampled haplotypes, which correlates with an increased probability of shared haplotypes between closely related species, or because of a mismatch between the number of intra‐ and interspecific sequences, therefore altering the results obtained through genetic distances (Phillips et al., 2019). Similarly, tree‐based methods rely on the upstream correct preparation of phylogenetic trees (Goulpeau et al., 2022). GMYC can overestimate the number of taxa, particularly in species with a strong intraspecific genetic structure (Hilário et al., 2021). However, approaches like GMYC and PTP are considered robust when properly used since they make stronger assumptions about speciation and distribution of genetic diversity because they use a speciation model and require tree‐building with an a priori model of nucleotide substitution (Hubert et al., 2024). Therefore, each delimitation tool has its own advantages and limitations, and by combining multiple delimitation algorithms results interpreted with the help of other sources (morphology, ecology, biogeography, etc.) limits the impacts of biased sampling or particular evolutionary histories (Carstens et al., 2013).
Our study revealed discrepancies between molecular taxonomy and morphological identification. The results indicate the presence of three distinct C. rosea lineages, suggesting the possibility of cryptic genetic differentiation and/or misidentification.
This taxonomic inconsistency was previously observed by Costa et al. (2012) for Atlantic specimens and Teramura et al. (2022) for Pacific specimens. These authors noted that C. rosea sequences in BOLD fall into different BINs, indicating divergent and overlapping lineages. Now we can say that these BINs are fully supported by our results, which combine several independent approaches. Therefore C. rosea is a complex of cryptic species, classified as a single species on the basis of morphological similarities, but with distinct genetic lineages.
Interestingly, C. rosea complex shows a clear geographic pattern, clustering together the samples from the Indian Ocean, the northwestern Pacific and the North Atlantic. Similar results for marine fish species with a supposed global distribution, which turned out to be several geographically distinct species, have been found in Shortfin neoscopelid Neoscopelus microchir Matsubara, 1943 (Bañón et al., 2024), Reef croaker Odontoscion dentex (Cuvier, 1830) (da Silva et al., 2023) or in the genus Monacoa Whitley, 1943 (Poulsen et al., 2016).
The morphological differences found in C. rosea between Atlantic and Pacific specimens may represent allopatric speciation between specimens with large geographical separations, as indicated by molecular analyses. According to these results, C. rosea forms a species complex consisting of at least three putative species. Based on the type locality, C. rosea would be an Atlantic species, while there may be up to two other morphologically similar Indo‐Pacific species that have not yet been described. The addition of a species key to the family Parazenidae and the geographical representation of the C. rosea molecular operational taxonomic units will be useful for the future delimitation of this species complex.
Despite being represented by only three sequences, all delimitation analyses showed two independent clusters for C. cypho. This is a remarkable result taking into account the low sample size for this species, which normally hides genetic diversity (Luo et al., 2015). Further analyses are needed to unravel the hidden diversity of C. cypho.
A particular case is the sequence morphologically identified as C. rosea, but which has no clear position in the tree and therefore is not part of the C. rosea complex. All delimitation analyses agreed to consider it as a separate species but the relationships among it, C. cypho and C. rosea complex remain unclear.
Similarly, previous taxonomic studies on P. pacificus have indicated the existence of cryptic species. Based on morphological characters, Kotlyar (2001) suggested that the Atlantic parazenid specimens of P. pacificus may be a separate species from the Indo‐Pacific specimens. Morphological differences and COI gene analysis from multiple specimens of P. pacificus across its geographic range support the hypothesis that this species may comprise at least two cryptic species, Atlantic and Indo‐Pacific (Corcoran & Grande, 2023; Singer et al., 2022). In our analyses, the lack of public sequences from the Atlantic Ocean did not allow us to test this hypothesis. However, while distance‐based methods and PTP both identified P. pacificus as a single species, GMYC split it into two groups (northwestern Pacific vs. northwestern Pacific and Indian Ocean). These results should be interpreted with caution because PTP is generally regarded as providing more accurate species delimitations than GMYC, which has a tendency to overestimate species numbers (Tang et al., 2014). Further analysis including samples from the North Atlantic will be necessary to resolve the taxonomic status of P. pacificus.
Discrepancies between morphological identification and molecular taxonomy are not uncommon in other zeiform fishes such as John Dory Zeus faber L. 1758 (Ward et al., 2008), silvery John Dory (Lowe, 1852) Zenopsis conchifer (Matusevich et al., 2024) or mirror Dory Zenopsis nebulosa (Temminck & Schlegel, 1845) (Wang et al., 2012), suggesting hidden diversity within each nominal species. It is also not uncommon for new zeiform species to be described, such as Zenopsis stabilispinosa Nakabo, Bray & Yamada, 2006 or Zenopsis filamentosa Kai & Tashiro, 2019. All these indications suggest that the taxonomy of zeiforms is far from complete, pending the description of new cryptic species.
The growing number of cryptic fish species indicates that traditional morphological techniques may no longer be adequate for effective species discrimination. However, some species currently classified as cryptic might, on closer morphological and spatial analysis, be reclassified as pseudocryptic (Lajus et al., 2015). Really, the diagnostic characteristics of many fishes, mainly non‐commercial or deep‐sea fishes, are often based on ancient manuscripts by examining only a few specimens, and these descriptions remain valid to this day (Bañón et al., 2022). The lack of fish taxonomists, the difficulty of examining specimens from different geographical areas and the rejection of descriptive work by many scientific journals are behind these gaps in knowledge. The resulting lack of understanding of intraspecific variation complicates the identification of species boundaries, hindering accurate assessments of species diversity and distribution (Frutos et al., 2022).
The challenge for the future is not only to identify fish species with hidden biodiversity, but also to describe these new species. Unfortunately, all indications are that this will be a slow process given the lack of trained taxonomists and institutional support for biodiversity research.
AUTHOR CONTRIBUTIONS
R.B. conceived the idea and conducted morphological analyses. D.B.G., F.B., A.S.C. and A.d.C. conducted sequencing and molecular analyses. F.B. conducted the biogeography representation. F.B. and J.C.A. compiled the specimens. J.D.B.V. made the X‐ray images. R.B. wrote the first draft of the manuscript, which was further elaborated by D.B.G., F.B. and A.d.C. All authors critically reviewed and approved the manuscript.
FUNDING INFORMATION
This research was developed thanks to the European Maritime, Fisheries and Aquaculture Fund (PORCUDEM‐20233FMP001). This research was funded by national funds through the Fundação para a Ciência e a Tecnologia within the scope of the Strategic Funding UIDB/04423/2020 (https://doi.org/10.54499/UIDB/04423/2020), UIDP/04423/2020 (https://doi.org/10.54499/UIDP/04423/2020), and LA/P/0101/2020. (https://doi.org/10.54499/LA/P/0101/2020). A.d.C. research was partly funded through a contract for competitive reference research groups of the Xunta de Galicia (ED431C 2019/28).
CONFLICT OF INTEREST STATEMENT
The authors declare that they have no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bañón, R. , Baldó, F. , Serrano, A. , Barros‐García, D. , & de Carlos, A. (2022). Gaidropsarus gallaeciae (Gadiformes: Gaidropsaridae), a new northeast Atlantic rockling fish, with commentary on the taxonomy of the genus. Biology, 11, 860. 10.3390/biology 11060860 35741381 PMC 9219912 · doi ↗ · pubmed ↗
- 2Bañón, R. , Barros‐García, D. , Arronte, J. C. , Rábade, S. , del Río, J. L. , Baldó, F. , & de Carlos, A. (2024). Diving deeper into the taxonomy of the Neoscopelus species complex (Myctophiformes: Neoscopelidae) with the description of Neoscopelus serranoi sp. nov. Zootaxa, 5529(3), 487–510. 10.11646/zootaxa.5529.3.4 39646400 · doi ↗ · pubmed ↗
- 3Bañón, R. , Casas, J. M. , Piñeiro, C. G. , & Covelo, M. (1997). Capturas de peces de afinidades tropicales en aguas atlánticas de Galicia (NO de la península Ibérica). Boletín Del Instituto Español De Oceanografía, 13(1–2), 57–66.
- 4Cadenat, J. (1953). Notes d'lchtyologie ouest africaine, VI. Poissons des campagnes du “Gerard Treca”. Bulletin de l'Institut Français d'Afrique Noire, 15(3), 1051–1102.
- 5Carstens, B. C. , Pelletier, T. A. , Reid, N. M. , & Satler, J. D. (2013). How to fail at species delimitation. Molecular Ecology, 22(17), 4369–4383. 10.1111/mec.12413 23855767 · doi ↗ · pubmed ↗
- 6Corcoran, H. , & Grande, T. C. (2023). Geographic variation and species composition of the Zeiform fish Parazen . https://ecommons.luc.edu/
- 7Costa, F. O. , Landi, M. , Martins, R. , Costa, M. H. , Costa, M. E. , Carneiro, M. , Alves, M. J. , Steinke, D. , & Carvalho, G. R. (2012). A ranking system for reference libraries of DNA barcodes: Application to marine fish species from Portugal. P Lo S One, 7(4), e 35858. 10.1371/journal.pone.0035858 22558244 PMC 3338485 · doi ↗ · pubmed ↗
- 8da Silva, T. F. , Sampaio, I. , Angulo, A. , Domínguez‐Domínguez, O. , Andrade‐Santos, J. , Guimarães‐Costa, A. , & Santos, S. (2023). Species delimitation by DNA barcoding reveals undescribed diversity in Stelliferinae (Sciaenidae). P Lo S One, 18(12), e 0296335. 10.1371/journal.pone.0296335 38153939 PMC 10754464 · doi ↗ · pubmed ↗