Discovery of an aquaporin (CrAQP2) in the freshwater larval midge, Chironomus riparius and its role in response to road de-icers
Britney N. Picinic, Amber Reinsborough, Sima Jonusaite, Jean-Paul V. Paluzzi, Andrew Donini

TL;DR
Researchers discovered a water channel protein in midge larvae that helps them survive in salty water caused by road de-icers.
Contribution
The study identifies and characterizes CrAQP2, a novel aquaporin in Chironomus riparius, and its role in osmoregulation under de-icer exposure.
Findings
CrAQP2 is highly expressed in osmoregulatory organs like Malpighian tubules.
Knockdown of CrAQP2 reduces midge survival in de-icer and salt-contaminated water.
Beet juice de-icer affects midges similarly to NaCl in terms of osmoregulation.
Abstract
•CrAQP2 is expressed in anal papillae and throughout the alimentary canal of the midge.•CrAQP2 expression is altered in response to NaCl in various osmoregulatory organs.•Knockdown of CrAQP2 reduces survival of midges in de-icer contaminated water.•Beet juice de-icer causes similar effects on C. riparius, relative to NaCl in water. CrAQP2 is expressed in anal papillae and throughout the alimentary canal of the midge. CrAQP2 expression is altered in response to NaCl in various osmoregulatory organs. Knockdown of CrAQP2 reduces survival of midges in de-icer contaminated water. Beet juice de-icer causes similar effects on C. riparius, relative to NaCl in water. Chironomus riparius midge larvae are ubiquitous in freshwater ecosystems throughout the Northern Hemisphere. Recently, freshwater in temperate regions has experienced a threat from salinization due to the use of road de-icers.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1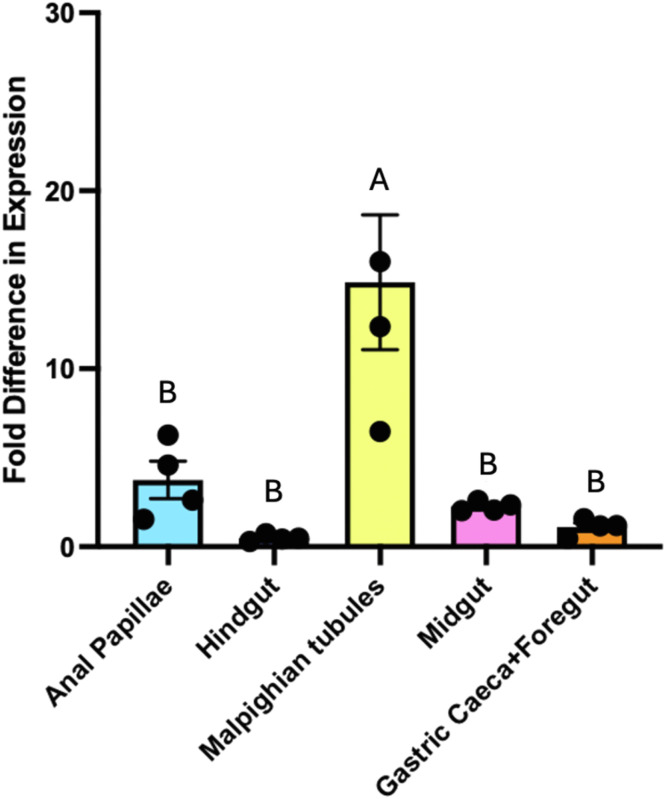 Figure 2
Figure 2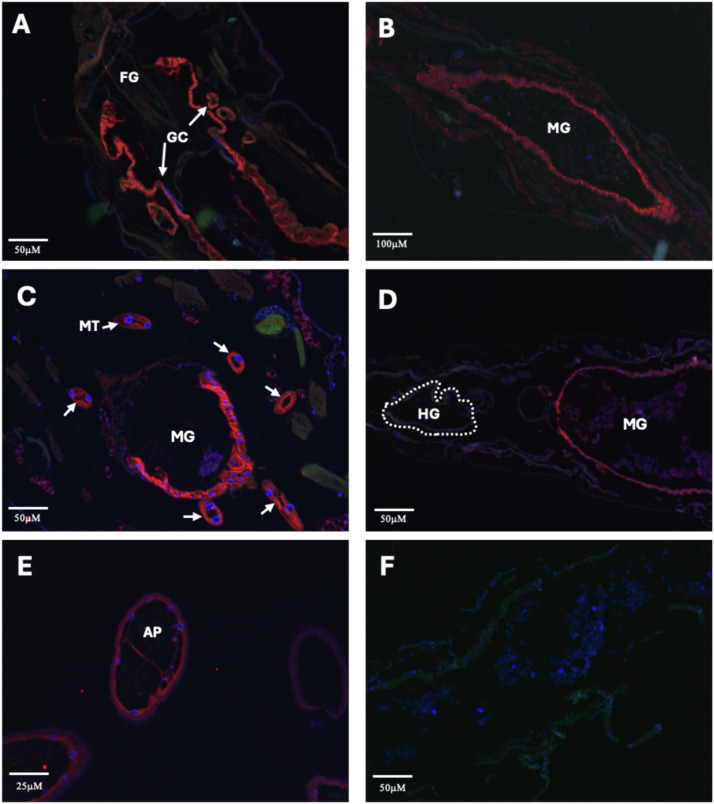 Figure 3
Figure 3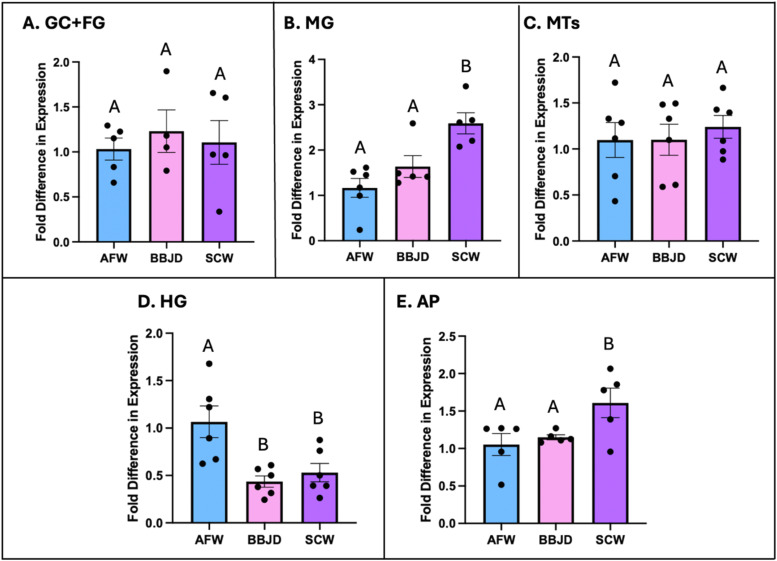 Figure 4
Figure 4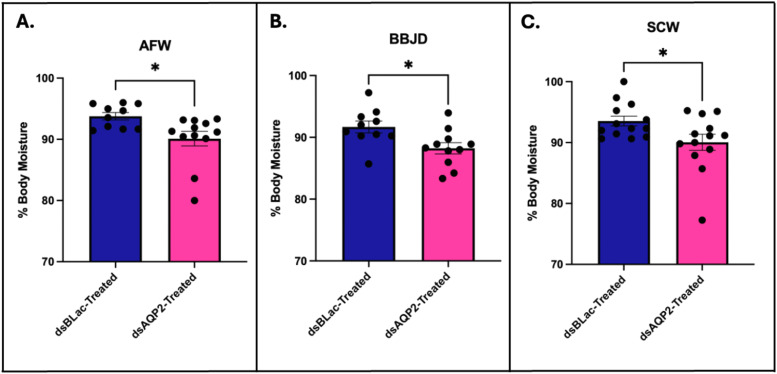 Figure 5
Figure 5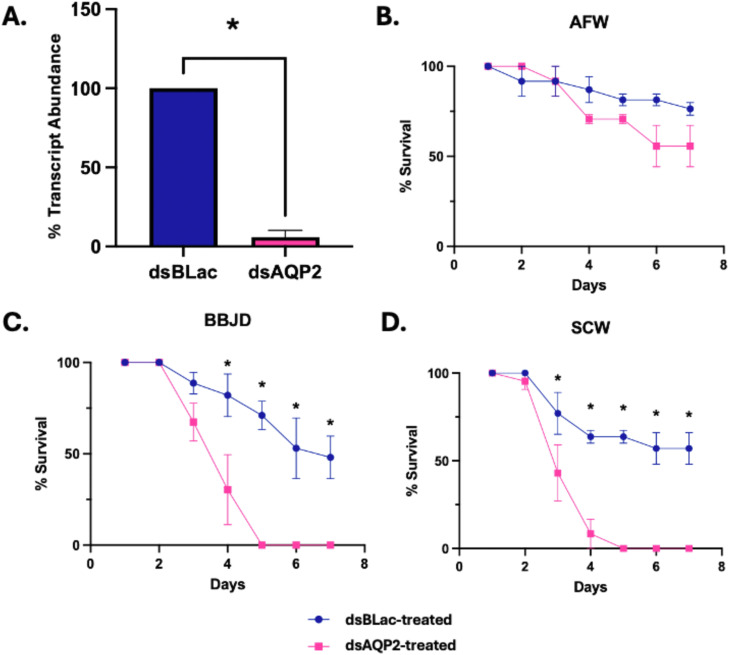 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhysiological and biochemical adaptations · Smart Materials for Construction · Freshwater macroinvertebrate diversity and ecology
Introduction
1
The study of freshwater fauna has been a popular topic due to the unique diversity seen in freshwater ecosystems. Inland waters harbour a wide variety of animals, with over 60 % of those animals being aquatic insects (Dijkstra et al., 2014). The ecological role of aquatic insects is important as they act as primary consumers, detritivores, and predators (Dijkstra et al., 2014). In particular, the presence of aquatic detritivorous insects is essential to freshwater ecosystems because they feed on decaying matter and play a critical role in decomposition and nutrient cycling within their ecosystems. One of many detritivores and primary consumers is the dipteran Chironomidae species, Chironomus riparius, which is a non-biting freshwater midge that is widely distributed throughout the Northern Hemisphere (Pinder, 1986; Nemec et al., 2012).
Larval C. riparius reside in freshwater aquatic environments as benthic organisms and their post-embryonic development involves four larval instar phases before pupating and emerging as terrestrial adults. The larvae have been effectively used in toxicology studies for freshwater ecosystems (Vogt et al., 2011; Nicacio and Juen, 2015; Manfrin et al., 2018). C. riparius larvae have been shown to tolerate moderate increases in salinity (Jonusaite et al., 2010). This is significant because in recent decades, anthropogenic activities including but not limited to, the application of fertilizers, resource mining, and the use of road de-icers have resulted in increased levels of salts in freshwater ecosystems. These activities have thus been identified as deadly threats to freshwater ecosystems (Shuler and Relyea, 2018). In the Northern Hemisphere where C. riparius is widely distributed, the use of road de-icing agents through the winter months is important in preventing ice build-up on roadways but leads to freshwater salinization. Typically, the de-icers used are in the form of NaCl pellets or NaCl brine. However, as temperatures fluctuate in these northern regions, the snow and ice melt carries the NaCl with it into the nearby freshwater ecosystems. It has been shown that road salt runoff and increased salinity can have detrimental effects on freshwater organisms that inhabit these aquatic ecosystems due to elevated levels of Na^+^ and Cl^-^ (Meter et al., 2011; Nowghani et al., 2019; Hintz et al., 2022; Dugan and Arnott, 2022). As a potentially eco-friendly alternative to road salt, brine-beet juice de-icers (BBJD) containing relatively lower levels of NaCl are being implemented in limited amounts as replacement to the traditional NaCl de-icers (Nutile and Solan, 2019; Picinic et al., 2022; Reinsborough and Donini, 2025). However, as the BBJD is derived from sugar beets, it is naturally high in K^+^-levels, with reportedly at least 7-fold greater levels of K^+^ in comparison to what is found in NaCl-based de-icers (Fay and Shi, 2012; Wruss et al., 2015; Picinic et al., 2022; Reinsborough and Donini, 2025). The impact of K^+^ on freshwater animals warrants further investigation before widespread use of BBJD as a de-icing agent since it has been documented that half the concentration of KCl compared to NaCl is lethal to at least one chironomid species (Hamilton et al., 1975). In previous studies, it has been shown that increasing concentrations of BBJD in the surrounding media of a freshwater bivalve causes body K^+^ levels to drastically rise (Gillis et al., 2021). More recently, it was shown that BBJD may be equally harmful compared to the traditional NaCl-based de-icer to the amphipod crustacean, Hyalella azteca, due to increasing levels of ions in the water which impacted the activity of important enzymatic ion-transporters (Picinic et al., 2022). Furthermore, BBJD caused significant alterations in the osmoregulatory physiology of C. riparius, which reduced K^+^ uptake and favoured K^+^ secretion (Reinsborough and Donini, 2025).
Aquatic insects regulate salt and water balance through the function of several specialized osmoregulatory organs found in the alimentary canal that work collectively. The gastric caeca (GC) are implicated in ion and water absorption into the haemolymph (Dow, 1987; Clark et al., 1999; Jonusaite et al., 2013; Linser and Dinglasan, 2014; D’Silva et al., 2017). The midgut (MG) is primarily involved in food digestion, nutrient absorption, but also plays a role in ion and water absorption (Dow, 1987; Clark et al., 1999; Jonusaite et al., 2013; Linser and Dinglasan, 2014). The Malpighian tubules (MTs) function in conjunction with the hindgut (HG) to produce urine that favours water secretion and ion conservation in freshwater aquatic insects, driven by the active transport of ions and movement of water via osmosis (Clark and Bradley, 1998; Donini et al., 2006; Zadeh-Tahmasebi et al., 2016). To achieve this, the ion rich primary urine secreted by the MTs is passed to the HG where final alterations of the urine such as the reabsorption of ions can occur, especially in a freshwater environment (Bradley and Phillips, 1977; Jonusaite et al., 2013; Linser and Dinglasan, 2014). Some larval-stage insects including C. riparius and the mosquito Aedes aegypti, possess externally localized osmoregulatory organs known as the anal papillae (AP). The AP are balloon-shaped organs comprised of a single-cell layer thick epithelium that is primarily involved in active ion uptake from the environment in freshwater aquatic insects (Jarial, 1995; Nguyen and Donini, 2010; Surendran et al., 2018). In addition, many aquatic insects also possess ionocytes whether on their gills or their integument, which function similarly to the AP for ion uptake in freshwater environments (Cochran et al., 2024). All together, these osmoregulatory organs work to regulate the ionic composition and osmolarity of body fluids and play an important role in the ability of aquatic insects to tolerate changes in environmental salinity.
To date, the effects of salinization on ion transport in freshwater insects has been studied and reviewed (Silver and Donini, 2021); however, comparatively much less is known about how salinization affects the transport of water. A key component in regulating and maintaining water levels in freshwater insects is the presence of water channel proteins, known as aquaporins (AQPs). AQPs are characterized as transmembrane domain proteins that form a channel in which water can traverse cellular membranes according to the osmotic gradient (Campbell et al., 2008; Spring et al., 2009; Drake et al., 2015; Picinic et al., 2024). AQPs have been characterized in other dipteran insects such as the fruit fly Drosophila melanogaster (Dow and Davies, 2001; Kaufmann et al., 2005; Campbell et al., 2008) and the mosquito Ae. aegypti (Duchesne et al., 2003; Drake et al., 2010, 2015; Akhter et al., 2017; Misyura et al., 2017, 2020; Picinic et al., 2024). Additionally, some AQP genes have been identified in midges such as the sleeping chironomid Polypedilum vanderplanki (Kikawada et al., 2008) and the non-flying Antarctic midge Belgica antarctica (Yi et al., 2011); however, there is currently no information about AQPs in the freshwater midge, C. riparius.
Of the six known insect AQP genes, AQP1 and AQP2, which are named following nomenclature established previously (Drake et al., 2010), have been identified as water-specific AQPs that have a high selectivity for water transport (Drake et al., 2015). AaAQP1 has been localized in larval Ae. aegypti to key osmoregulatory organs including the GC, MG, MTs, HG, and AP (Marusalin et al., 2012; Misyura et al., 2020). The AQP1 homolog in B. antartica was also found to be localized to the FG, MG, and MTs (Yi et al., 2011), alluding to its important function in water transport in these various osmoregulatory organs. Additionally, AaAQP2 has also been localized to the GC, MG, MT, HG, and AP of larval Ae. aegypti (Marusalin et al., 2012; Misyura et al., 2020) and its homolog has been immunolocalized in the FG, MG, and HG of the midge B. antarctica (Yi et al., 2011). Furthermore, AaAQP2 has been shown to be salinity responsive in the AP of larval Ae. aegypti, where increased salinity in brackish water conditions caused a significant increase in AaAQP2 transcript and protein levels relative to freshwater conditions (Durant et al., 2021).
In order to begin to understand how both NaCl and BBJD may affect the regulation of water transport in freshwater insects we identified an important putative water channel protein CrAQP2, a homolog of the insect AQP2, and studied its expression in the osmoregulatory organs of C. riparius exposed to NaCl or BBJD. We hypothesized that CrAQP2 transcript and protein would be abundant in each of the osmoregulatory organs of C. riparius, with highest expression expected in the MTs and AP where ion and water transport are constant in hypoosmotic conditions. Additionally, we predicted that in the presence of the de-icers, Craqp2 would be upregulated in the organs that typically function in water absorption such as the MG and AP; whereas Craqp2 was predicted to be downregulated in organs that typically function in water secretion such as the GC and MTs. Finally, we hypothesized that the reduction of Craqp2 transcript expression in the midge larvae by RNA interference would result in increased mortality when larvae were faced with abrupt transfer to salinity on account that they would have difficulty regulating body water levels.
Materials and methods
2
Animal rearing
2.1
Chironomus riparius third and fourth instar larvae were taken from a laboratory colony that is maintained at York University (Jonusaite et al., 2013). Larvae developed in ∼6 L tanks containing ∼3 cm of sand (K&E Industrial Sand, ON, Canada) and 3–4 L of artificial fresh water (AFW: 300μ M NaCl, 300μ M NaHCO_3_, 380μ M CaCl_2_, 43μ M KCl; pH 7.6). The tanks were aerated with air stones connected to a constant air supply. The larvae were fed every 2–3 days with crushed TetraFin Goldfish Flake Food (Tetra Holding US, VA, USA) and kept at 21 to 23 °C on a 12h:12 h light: dark schedule. The water in the tanks was topped up once a week with AFW and water changes were completed every 3–4 weeks. Any potential accumulation of salts from the dilute AFW was measured to be negligible. For treatment with de-icers, 2 % brine beet juice de-icer (BBJD) and 7 g/L NaCl-contaminated water (SCW) were chosen based on previously established LC_50_ curves, which determined 50 % survival at each of those concentrations for late-stage C. riparius larvae (Reinsborough and Donini, 2025; Jonusaite et al., 2013). To the best of our knowledge, there is no method for measuring the amount of BBJD that enters waterways but, in the case of NaCl, chloride levels ranging from ∼ 4 to 7 g/L and as high as 10.3 g/L have been recorded in bodies of water near roads (Corsi et al., 2010; Hintz et al., 2022; Kaushal et al., 2005).
Identification of CrAQP2 tissue collection, and quantification of CrAQP2 expression with qPCR
2.2
To identify Craqp2 in C. riparius larvae, the C. riparius reference genome PGI_CHIRRI_v4 (GCA_917,627,325.4; Schmidt et al., 2020) was searched by tblastn operation using the AQP2 (PRIP) deduced protein sequences of Aedes aegypti (AY433444), Polypedilum vanderplanki (AB281619), and Belgica antarctica (AB602340) through the National Center for Biotechnology Information (NCBI) database. Primers were then designed to target a large portion of the putative Craqp2 gene (Table 1). To obtain total RNA samples, 3–4 fourth instar larval C. riparius were taken from the lab colony and transferred to a chilled 300μl aliquot of lysis buffer, prepared from the PureLink RNA minikit (Invitrogen, MA, USA). RNA extraction on whole body samples was completed following the protocol provided in the PureLink RNA minikit (Invitrogen). RNA samples were purified using the TURBO DNAfree^࣪^ Kit (Applied Biosystems, ON, Canada) and cDNA was then synthesized using the iScript cDNA synthesis kit (BioRad, CA, USA) consisting of both oligo(dT) nucleotides and random hexamer primers. The cDNA template was then amplified with Q5® High-Fidelity DNA Polymerase (New England Biolabs, ON, Canada) using specific Craqp2 primers (Table 1) and amplicon band size was confirmed on an agarose gel. The amplified product was then purified using the Monarch PCR Purification Kit (New England Biolabs, ON, Canada), concentration was determined using a NanoDrop 2000 spectrophotometer (Thermo Scientific, DE, USA) and then samples were sent for Sanger sequencing (The Centre for Applied Genomics, Sick Kids, ON, Canada). Sequencing quality and base accuracy was analyzed using Geneious Software (Dotmatics, MA, USA). Following this, quantitative real time PCR (qPCR) primers were designed from the confirmed partial gene sequence, targeting exon-exon boundaries to ensure cDNA amplicon specificity (Table 1).Table 1. Oligonucleotides used for Chironomus riparius AQP2 gene amplification.Table 1 dummy alt textOligo NameForward Primer SequenceReverse Primer SequenceExpected Band SizeExperiment Used For****CrAQP1CACTGGGTGTTGACGAGATCTAAAGAGATGGGCGCACTTC780bpPCRCrAQP2 qPCRTGCTATGACTATTGGACATGACATGTAGTGCTGCAGTTCC153bpqPCR40S Reference GeneTGTACAGAAGAAGAGGTCGAGAAAGCATACCACGATGAGCACG230bpqPCRRP11 Reference GeneCCGTACATTGCACAGTTCGTCATTATAGCCAGGTCTTCCC216bpqPCRElf1 Reference GeneGCTCCTGGACATCGTGATTTTGACTCCCAATGTGAAAGCC159bpqPCRTubulin Reference GeneACTTCTGCTTCAAAATGCGTGCTACGGTTGGTTCCAGATCG237bpqPCRCrAQP2 dsRNACACTGGGTGTTGACGAGATCGCAACACTTCCAAAACATTGAGC304bpdsRNA SynthesisdsAQP2 Reverse T7CACTGGGTGTTGACGAGATCTAATACGACTCACTATAGGGGCAACACTTCCAAAACATTGAGC320bpdsRNA SynthesisdsAQP2 Forward T7TAATACGACTCACTATAGGGCACTGGGTGTTGACGAGATCGCAACACTTCCAAAACATTGAGC320bpdsRNA SynthesisBLac dsRNAATTTCCGTGTCGCCCTTATTCCGTTCATCCATAGTTGCCTGAC800bpdsRNA SynthesisEach oligo name is listed with the corresponding forward and reverse primer sequences used to produce each amplicon. Oligonucleotides used for AQP2 gene identification and amplification are shown and were used for PCR, qPCR (RT-PCR), and dsRNA synthesis.
Craqp2 transcript levels were then measured in the alimentary canal of AFW-reared larval C. riparius including the foregut with gastric caeca (FG+GC), midgut (MG), Malpighian tubules (MTs), hindgut (HG), and anal papillae (AP). The organs were isolated and pooled from ∼40 larvae on ice in 200μl of chironomid saline (5 mM KCl, 74 mM NaCl, 1 mM CaCl_2_, 8.5 mM MgCl_2_, 10.2 mM NaHCO_3_, 8.6 mM HEPES, 20 mM glucose, 10 mM glutamine), which were dissected in a Sylgard®-lined petri dish filled with the same saline, constituting one biological sample. Following collection, organ samples were centrifuged at 10,000 g for 1 min at RT, the saline was removed, and 1 mL of TRIzol® reagent (Invitrogen, ON, Canada) was added. Samples were sonicated on ice using an XL 2000 Ultrasonic Liquid Processor (QSONICA - LLC, CT, USA) and total RNA was isolated from the lysed organ samples following the manufacturer’s guidelines. The resulting RNA samples were treated with the TURBO DNAfree^࣪^ Kit for removal of any potential genomic DNA (Applied Biosystems, ON, Canada). RNA sample concentrations and purity values were measured using a NanoDrop 2000 spectrophotometer (Thermo Scientific, DE, USA). cDNA for each organ was synthesized using the iScript cDNA synthesis kit (BioRad, CA, USA), as previously stated and following the manufacturer’s instructions. For examining of the effects of de-icers on Craqp2 transcript abundance in each of the osmoregulatory organs previously listed, midge larvae were taken from the lab colony and placed in 100 mL of either BBJD, SCW or AFW (control) for 24hr acute exposures. Following that, all samples were treated identically for RNA isolation and cDNA synthesis (as described above), and for subsequent qPCR. Craqp2 transcript abundance in AFW, BBJD, and SCW-treated midge larvae was determined with qPCR conducted using a CFX Duet Real-Time PCR System (Bio-Rad Laboratories, ON, Canada) in combination with the SsoAdvanced Universal SYBR Green Supermix (Bio-Rad Laboratories, ON, Canada), following the manufacturer’s instructions. Relative Craqp2 transcript abundance was calculated using the previously established ΔΔC_T_ method (Livak and Schmittgen, 2001) and for the expression profile, values were normalized to the geometric mean of Rp11 and 40S reference genes. For individual Craqp2 expression in organs exposed to the de-icers, Rp11 and 40S reference genes were used for the GC+FG, MG, HG, and AP, whereas tubulin and Elf1 were used for the MTs as a result of variations in gene transcript in the MTs with BBJD and SCW (Table 1). All reference genes were confirmed to show no changes in transcript abundance with each treatment including AFW, BBJD, and SCW, in each of the organs studied with the respective sets of reference genes. The expression profile across the osmoregulatory organs in freshwater involved 3–4 biological replicates, while the individual Craqp2 expression analyses for each organ in response to road de-icers involved 4–6 biological replicates, all loaded as technical triplicates for quantitative accuracy.
Immunohistochemistry
2.3
Immunohistochemistry (IHC) was carried out on whole body (WB) C. riparius midge larvae to localize CrAQP2 in the alimentary canal. Midges were taken from the lab established colony and placed in 100 mL of AFW, BBJD, or SCW for 24hr acute exposures. Midges were placed in Bouin’s fixative for 2hr at room temperature followed by a replacement of fixative with 70 % ethanol and kept at 4 °C until ready for dehydration. For each treatment, 3–4 biological replicates were used, in addition to ∼4 technical replicates for confirmation of immunostaining consistency. The IHC procedures carried out in this study followed previously established protocols (Akhter et al., 2017; Misyura et al., 2020; Sajadi et al., 2023; Picinic et al., 2024). Briefly, fixed WB samples were embedded in paraffin wax and then ∼5μ m sections were cut using an Epredia HM325 manual microtome (Epredia, MI, USA). Sectioned tissues were then placed on Fisherbrand࣪ ColorFrost࣪ Plus Adhesion Microscope slides, with each slide having tissue sections from the AFW, BBJD, and SCW samples, adjacent to each other. Slides were then processed in a two-day procedure where tissue sections were rehydrated and repeatedly washed with 1xPBS. All slides were probed with a previously described affinity purified polyclonal antibody raised against a peptide representing a small fragment of the Aedes aegypti AQP2 homolog (AaAQP2) (1:50 rabbit polyclonal antibody raised against the peptide CNGLGNTGLKENVQD, Genscript, NJ, USA) (Durant et al., 2021; Picinic et al., 2024). An alignment between the custom AaAQP2 antibody antigen sequence and the putative CrAQP2 antigen sequence revealed ∼50 % similarity (see Supplementary Figure S1). The mouse monoclonal anti-a5 antibody for Na^+^/K^+^-ATPase (NKA) (Douglas Fambrough, Developmental Studies Hybridoma Bank, IA, USA, 1:5 dilution) was used as a membrane marker for all tissue sections (Picinic et al., 2024). The secondary antibody used to detect AaAQP2-like protein was a goat anti-rabbit AlexaFluor 594 conjugated (Jackson Immunoresearch) antibody used at a 1:400 dilution and the secondary antibody used to detect NKA was a goat anti-mouse secondary antibody conjugated to Cy2 (Jackson Immunoresearch) used at a 1:500 dilution. To determine antibody specificity, control slides were prepared, where the AaAQP2 primary antibody was omitted from the processing step and only secondary antibody was added. Control and experimental slides were treated identically, where tissue sections from AFW, BBJD, and SCW samples were placed adjacent to each other on each slide and ran through the same IHC processing protocol together. Slides were then mounted with 1:1 of glycerol:1xPBS mounting media containing 4μg/mL DAPI and left to dry for at least 24hr before imaging. Fluorescent microscopy was completed using an Olympus IX81 fluorescent microscope (Olympus Canada, ON, Canada) and images were captured using CellSense® 1.12 Digital Imaging software (Olympus Canada). Identical acquisition settings were utilized for all tissue sections on each individual slide, to ensure comparisons could be made between AFW, BBJD, and SCW tissue samples. Images were overlayed using ImageJ 1.53k software.
dsRNA synthesis and confirmation of CrAQP2 knockdown
2.4
To obtain whole body RNA samples, 3–4 fourth instar larval midges were transferred to a chilled 300μl aliquot of lysis buffer, prepared from the PureLink RNA minikit (Invitrogen, MA, USA) and RNA extraction, purification, and cDNA synthesis was completed as described above. To obtain a large amount of Craqp2 cDNA serving as template for dsRNA synthesis*,* PCR reactions with the Q5 Polymerase (New England Biolabs, ON, Canada) using specific Craqp2 primers designed to target a relatively large portion of the gene (Table 1). The amplified product was then purified using the Monarch PCR Purification Kit (New England Biolabs, ON, Canada) and its concentration was determined using a NanoDrop 2000 spectrophotometer (Thermo Scientific, DE, USA). dsRNA synthesis was completed following a previously established protocol (Chasiotis et al., 2016; Misyura et al., 2017). The T7 promoter sequence was added to the 5′ end of forward and reverse primers used to amplify a portion of the Craqp2 gene. In addition, as a knockdown control gene, a fragment of the bacterial β-lactamase (βLac) gene was amplified from a pGEM-T-Easy vector using previously established primers (Chasiotis et al., 2016) and βLac primers with T7 promoter sequence were used (Table 1). Separate reactions were run which included the dsRNA cDNA template with either the appropriate forward primer and the reverse primer with a T7 promoter sequence or the forward primer with T7 promoter sequence and the appropriate reverse primer. Following amplification, matching reactions were pooled and then purified with the Monarch PCR Purification kit (New England Biolabs, ON, Canada). The amplicons containing T7 promoter sequences were then used to make dsRNA for Craqp2 and βLac with the Promega T7 RiboMAX Express RNAi System Kit (Promega, WI, USA), following the manufacturer’s instructions. Delivery of dsRNA for Craqp2 or βLac involved feeding larval midges following a protocol previously established for larval Ae. aegypti knockdown (Singh et al., 2013; Chasiotis et al., 2016; Durant and Donini, 2018) with minor modifications. Specifically, for each biological replicate, ∼10–12 third and fourth instar larvae were taken from the lab colony and placed in 150μl of RNAse-free water containing 0.5μg/μl dsRNA (Craqp2 or βLac), left to incubate in an open microcentrifuge tube for 2hr at room temperature, before being transferred to 100 mL of aerated AFW. Larvae were left in the aerated AFW containers for 48hr and 72hr post dsRNA feeding before 4–5 whole body larvae were sonicated in chilled lysis buffer, total RNA was extracted, and cDNA synthesized as described above. To confirm knockdown, qPCR on the cDNA samples was performed as described above and the relative transcript abundance in 48hr post Craqp2 dsRNA feeding was normalized to the dsβLac treatment group.
Body water
2.5
Potential effects of Craqp2 knockdown on total body water of C. riparius larvae under osmotic challenges was measured following a previously established protocol (Nguyen and Donini, 2010). Larvae were placed in microcentrifuge tubes at a density of ∼10 larvae/150μl of 0.5μg/μl dsAQP2 or dsβLac for 2hr at room temperature, as described above and then transferred into aerated containers of AFW for 48hr. Larvae were equally distributed into containers containing 100 mL of AFW, BBJD, or SCW for a 24hr acute exposure. Following this exposure, individual midges were dried on a KimTech wipe and placed into individual pre-weighed PCR tubes where their wet weight was measured using a microbalance (model UMXP; Mettler-Toledo International, ON, Canada). Larvae in their individual PCR tubes were placed in an oven set to 60 °C for ∼5 days. Larvae were weighed again in their individual PCR tubes to obtain their dry weight and the difference between wet weight and dry weight was calculated to determine total body water. For AFW n = 6–8; BBJD n = 7–10; SCW n = 7–10.
Midge survival
2.6
To assess the survivability of larval C. riparius under osmotic challenges post Craqp2 knockdown, three rounds of survival assays were completed. For each round, 30 larvae were placed in microcentrifuge tubes at a density of ∼10 larvae/150μl of 0.5μg/μl dsAQP2 or dsβLac. The microcentrifuge tubes were kept open at room temperature for 2hr. Midges were transferred to containers with 100 mL of aerated AFW, for dsAQP2 and dsβLac individuals, respectively and held for 48 hrs. Then, the group of larvae were evenly distributed into to 200 mL of AFW as the control, or the treatment groups with either BBJD or SCW, all with constant aeration. The midge larvae were then left in their treatments for 7 days, with daily survival counts completed. For dsβLac-treated individuals, the total number of larvae assessed was n = 17 for AFW, n = 18 for BBJD, and n = 17 for SCW. For dsAQP2-treated individuals, the total number of larvae assessed was, n = 18 for AFW, n = 18 for BBJD, and n = 19 for SCW. All treatment groups were fed identically, with each container receiving 0.1 g of crushed TetraFin Goldfish Flake Food (Tetra Holding US, VA, USA) ∼48hr post transfer into AFW, BBJD, or SCW.
Statistics
2.7
All data sets were analyzed using Prism^ࣨ^ 9 software (GraphPad Software Inc., CA, USA). Each qPCR data set, including the Craqp2 expression profile and effects of de-icers on Craqp2 transcript in each organ, was analyzed with a one-way ANOVA with multiple comparisons following log-transformation. An unpaired t-test was used to determine significance between dsβLac-treated and dsAQP2-treated midges to determine significance of knockdown efficiency of Craqp2 transcript. For survival and body water data, a one-way ANOVA with multiple comparisons was completed to determine significant differences between dsβLac-treated and dsAQP2-treated midge larvae.
Results
3
Characterization of CrAQP2
3.1
The deduced amino acid sequence of CrAQP2 contains the hallmark features of aquaporins with the two NPA motifs that form the pore of the channel and the selective filter-forming aromatic-arginine residues (Fig. 1). The CrAQP2 sequence is similar to the sequences of characterized PRIP aquaporins as shown by sequence alignment with several PRIP aquaporin homologs from other insects.Fig. 1Multiple sequence alignment of the Chironomus riparius aquaporin CrAQP2 (PRIP) deduced protein sequence with select orthologs from other insects. CrAQP2 shares sequence similarity with characterized insect PRIPs including: Apis mellifera (XP_394,391.1), Drosophila melanogaster (NP_610,686.1), Anopheles gambiae (BAI60044.1), Aedes aegypti (XP_001656932.1), Polypedilum vanderplanki (BAF62090.1), Bombyx mori (NP_001153661.1). Dashed boxes identify the two NPA motifs and solid boxes identify the selectivity filter aromatic-arginine residues (ar/R).Fig 1 dummy alt text
Craqp2 transcript abundance in osmoregulatory organs
3.2
Craqp2 transcript was detectable in each of the osmoregulatory organs examined (Fig. 2). The MTs had the highest levels of Craqp2 transcript, which was significantly higher than all other osmoregulatory organs analyzed with at least 5 times greater abundance. Comparatively, Craqp2 transcript abundance in all other examined osmoregulatory organs showed no statistical difference, although the trend indicated moderate levels in the AP and MG, while HG and GC/FG had the lowest abundance.Fig. 2CrAQP2 expression profile in the osmoregulatory organs of Chironomus riparius larvae in freshwater. Larval freshwater Chironomus riparius aquaporin-2 (CrAQP2) transcript abundance in the osmoregulatory organs, including the gastric caeca with foregut (GC+FG), midgut (MG), Malpighian tubules (MTs), hindgut (HG), and anal papillae (AP). CrAQP2 was detected in each of the organs studied, with highest transcript abundance in the MTs and lowest abundance seen in the HG tissue relative to the GC+FG as the calibrator. Bars show mean (± SEM) values with different letters denoting significantly different CrAQP2 transcript abundances between organs as determined by one-way ANOVA with Tukey’s multiple comparison’s test (p < 0.05), n = 3–4.Fig 2 dummy alt text
Immunolocalization of CrAQP2 in larval C. riparius
3.3
We utilized a custom antibody raised against a peptide antigen representing a small portion of AaAQP2, the AQP2 homolog of Aedes aegypti (Durant et al., 2021). This antigen sequence is similar to the putative CrAQP2 sequence (see Supplementary Figure S1). Using this antibody, AaAQP2-like immunoreactivity was detected in the osmoregulatory organs, including the GC+FG, MG, MTs, and AP (Fig. 3). Although immunohistochemistry was completed for organs of larvae exposed to AFW, BBJD and SCW, no differences were observed in AaAQP2-like immunoreactive staining between treatments, therefore representative images from AFW larvae were chosen to summarize the localization of AaAQP2-like immunoreactivity in osmoregulatory organs (Fig. 3). Samples processed from BBJD and SCW-treated midges had similar relative AaAQP2-like immunoreactive staining (see Supplementary Figure S2). Beginning with the anterior portion of the alimentary canal, AaAQP2-like immunoreactivity was detected in the GC but not in the FG region, with staining appearing uniform within the basolateral membrane of the GC (Fig. 3A). AaAQP2-like immunolocalization was also visible in the MG tissue, with staining mostly appearing at the apical or lumen facing membrane of the gut cells (Fig. 3B). Transverse cross sections of the MTs show mainly cytosolic as well as apically localized AaAQP2-like immunoreactive staining (Fig. 3C). In the HG, there was no AaAQP2-like immunoreactivity detected (Fig. 3D). Finally, AaAQP2-like immunoreactivity was detected within the basal membrane of the externally located AP organ, with no staining visible at the apical membrane (Fig. 3E). The negative control, which excluded the primary antibody, had no AaAQP2-like immunoreactivity in any region of the whole-body midge (Fig. 3F).Fig. 3Immunolocalization of AQP2 in the osmoregulatory organs of 4th instar larval Chironomus riparius midges reared in freshwater. A. Gastric caeca (GC) and foregut (FG), B. midgut (MG), C. Malpighian tubules (MT), D. hindgut (HG), and E. anal papillae (AP). Panel F. shows the negative control with primary antibody omitted, where no AQP2 staining was observed over the whole-body midge section. For each organ, at least 3–4 biological replicates were completed each with 4–5 technical replicates run. Red staining indicates AQP2 immunoreactivity; green staining indicates the Na^+^/K^+^-ATPase (NKA) membrane marker immunoreactivity; and blue (DAPI) staining for nuclei.Fig 3 dummy alt text
effects of de-icers on CrAQP2 transcript abundance in midge osmoregulatory organs
3.4
Acute exposure (24 hr) to BBJD or SCW did not alter the CrAQP2 transcript abundance in the GC+FG and MTs (Fig. 4A& 4C). In the MG and AP, SCW treatment significantly increased the CrAQP2 transcript abundance relative to AFW; however, BBJD did not affect CrAQP2 transcript levels in these organs (Fig. 4B& 4E). In the HG, CrAQP2 transcript abundance was significantly reduced with both BBJD and SCW treatment compared to AFW (Fig. 4D).Fig. 4Analysis of CrAQP2 expression in the osmoregulatory organs of larval Chironomus riparius in response to road de-icers. Transcript abundance of CrAQP2 in the osmoregulatory organs of the midge following acute (24hr) exposures to artificial freshwater (AFW), brine beet juice de-icer (BBJD), and NaCl-contaminated water (SCW). **A.**CrAQP2 abundance in the gastric caeca with foregut (GC+FG) tissue, showing no changes in transcript abundance with de-icer treatment (n = 4–5). **B.**CrAQP2 abundance in the midgut (MG), which increased in SCW relative to the AFW and BBJD groups (n = 5–6). **C.**CrAQP2 abundance in the Malpighian tubules (MTs), showing no changes in transcript abundance with de-icer treatment (n = 6). **D.**CrAQP2 abundance in the hindgut (HG), which decreased in BBJD and SCW relative to the AFW group (n = 6). **E.**CrAQP2 abundance in the anal papillae (AP), which increased in SCW relative to the AFW and BBJD groups (n = 5). For each graph, values are displayed as mean±SEM and differences analyzed by one-way ANOVA with Tukey’s multiple comparisons where different letters denote significantly different transcript abundance (p < 0.05).Fig 4 dummy alt text
Influence of CrAQP2 knockdown on total body water content under exposure to de-icers
3.5
Total body water following CrAQP2 knockdown in C. riparius larvae exposed to de-icers was measured. Body water percentage was determined following CrAQP2 knockdown with acute (24hr) exposures to AFW, BBJD, or SCW. Larvae exposed to AFW with CrAQP2 knockdown experienced a significant decrease in body water in comparison to the dsßLac-treated individuals (Fig. 5A). Similarly, CrAQP2 knockdown resulted in a significant decrease in body water in comparison to dsßLac-treated controls following both BBJD (Fig. 5B) and SCW exposure (Fig. 5C).Fig. 5Total body water post CrAQP2 knockdown in larval Chironomus riparius post transfer to AFW, BBJD, and SCW. Body water percentage ( %) was measured after CrAQP2 knockdown and a subsequent acute (24hr) exposure to artificial freshwater (AFW), brine beet juice de-icer (BBJD), or NaCl-contaminated water (SCW). As a knockdown control, dsβLac-treatment was used for all conditions. A. Body water % post AFW transfer, showing significant decrease in body water with CrAQP2 knockdown. B. Body water % post BBJD transfer, showing a significant decrease in body water with CrAQP2 knockdown. C. Body water % post SCW transfer, showing significant decrease in body water with CrAQP2 knockdown. For all panels (A-C), individual unpaired t-tests were completed and an * represents p < 0.05 (n = 10–13).Fig 5 dummy alt text
Influence of CrAQP2 knockdown on midge survival under prolonged exposure to de-icers
3.6
The survival of C. riparius larvae was assessed in CrAQP2 knockdown larvae under prolonged exposure to de-icers. Larvae exposed to CrAQP2 dsRNA showed significantly reduced transcript abundance (∼90 %) after 48 hr exposure to dsRNA where levels were just 10 % of those in the ßlac dsRNA control group (Fig. 6A). Survivability was assessed over one week post CrAQP2 knockdown. When maintained in AFW, midge survival decreased over the course of 7 days in both dsßLac-treated and dsAQP2-treated larvae, with no significant differences observed between the two groups (Fig. 6B). When midges were transferred from AFW into BBJD, survival of dsAQP2-treated larvae was stable for the first 2–3 days post transfer, but significantly decreased relative to the dsßLac-treated larvae 4 days post transfer, before no survival was seen by day 5 (Fig. 6C). Similarly, in the SCW exposure group, dsAQP2-treated larvae had survival comparable to dsßLac-treated larvae for the first 2 days; however, a rapid decline in survivability was seen at 3-days post transfer with ∼50 % surviving, followed by <10 % survival at 4-days post transfer and no survival by day 5 post transfer (Fig. 6D).Fig. 6Chironomus riparius survival post CrAQP2 knockdown and following prolonged exposure to road de-icers. Midge larvae survival was observed over a 7-day period, following CrAQP2 knockdown and exposure to artificial freshwater (AFW), brine beet juice de-icer (BBJD), or NaCl-contaminated water (SCW), with mean±SEM values reported. All treatments were completed on both dsAQP2-treated and dsβLac-treated larvae, with the latter serving as a knockdown control with larvae treated identically. A. Verification of CrAQP2 gene knockdown in midge larvae, 48hr post dsRNA treatment. B. Survival ( %) of midge larvae in AFW for dsβLac-treated (n = 17) and dsAQP2-treated (n = 18) individuals. Survival between control and knockdown larvae was statistically similar over the 7-day period. C. Survival ( %) of midge larvae in BBJD for dsβLac-treated (n = 18) and dsAQP2-treated (n = 18) individuals. Survival in the CrAQP2 knockdown group was significantly reduced compared to the control larvae by 4-days post transfer to BBJD. D. Survival ( %) of midge larvae in SCW for dsβLac-treated (n = 17) and dsAQP2-treated (n = 19) individuals. Survival in the CrAQP2 knockdown group was significantly reduced compared to the control larvae by 3-days post transfer to SCW. A two-way ANOVA with multiple comparisons was run for each set of values and data points with an asterisk () denote statistically significant differences (p < 0.05) between the dsβLac*-treated and dsAQP2-treated larvae at each time point.Fig 6 dummy alt text
Discussion
4
Overview
4.1
Salinization of freshwater is a serious global issue and, in temperate climates, the use of de-icing agents during the winter season contributes to the problem. Thus, salinization of aquatic environments can have detrimental consequences to organisms living in freshwater, including aquatic insects, which are important in maintaining a healthy ecosystem. Although a number of studies have examined the effect of salinization on ion transport in freshwater insects (Kapoor et al., 1978; Jonusaite et al., 2011, 2013; Scheibener et al., 2016; Jonusaite et al., 2017; Nowghani et al., 2019; Orr and Buchwalter, 2020; Lambret et al., 2021; Picinic et al., 2022), comparatively little work has been done on water transport. In this study, we identified a putative water selective aquaporin (AQP) in C. riparius larvae which we named CrAQP2 and shares ∼70 % sequence identity with PRIP AQPs of other insects. We conclude that CrAQP2 is a homolog of the Pyrocoelia rufa integral protein (PRIP) (Cabrero et al., 2020; Drake et al., 2010; Yang and Piermarini, 2017; Tan and Chen, 2023). We then examined the effects of de-icing agents including NaCl and brine-beet juice de-icer (BBJD) on CrAQP2 expression and function.
We found CrAQP2 gene transcript and protein to be present in almost all of the osmoregulatory organs of midge larvae that were raised in their typical freshwater environment simulated in the laboratory by artificial freshwater (AFW). Studies on these PRIP channels have determined that these aquaporins transport water and some, but not all, can also transport urea or glycerol and therefore this suggests that the function of CrAQP2 is primarily for water transport (Azuma et al., 2012; Drake et al., 2015; Goto et al., 2011; Kikawada et al., 2008; Fabrick et al., 2014). This assumption is consistent with CrAQP2 having the highly conserved residues, phenylalanine, histidine, and arginine of the selectivity filter (see Fig. 1). Therefore, under this assumption that CrAQP2 transports water, we propose some functional roles for this transporter when C. riparius larvae are exposed to de-icing agents.
Expression of CrAQP2 in osmoregulatory organs of larvae in AFW, SCW and BBJD
4.2
Malpighian tubules
4.2.1
The highest abundance of CrAQP2 transcript was observed in the MTs of larval C. riparius and we found putative CrAQP2 (AaAQP2-like staining) protein dispersed throughout the cytosol and along the apical membrane of the MT cells (Fig. 2C). This observed staining pattern is consistent with CrAQP2 localization in the larger more abundant principal cells of MTs. In D. melanogaster, PRIP is highly expressed in the stellate cells of the MTs, aiding in transepithelial water entry into the lumen (Cabrero et al., 2020); whereas in adult A. aegypti, AaAQP2 (PRIP homolog) is expressed in both the principal and stellate cells of the MTs (Drake et al., 2010; Picinic et al., 2024). The MTs play an important role in both aquatic and terrestrial insects to produce primary urine, while maintaining haemolymph osmolarity. For C. riparius larvae that inhabit freshwater, the MTs must consistently produce dilute urine to clear excess water as a result of their hypoosmotic environment and CrAQP2 is positioned to participate in this process by transporting water into the lumen of the MTs. With de-icer treatment (BBJD and SCW), which decreases the gradient for water entry into the body but increases salt uptake, we found no changes to CrAQP2 transcript abundance, nor did we find any qualitative differences in AaAQP2-like staining in the MTs. In A. aegypti larvae reared in brackish water (30 % seawater; NaCl the predominant salts), transcript abundance of aquaporins (including AaAQP2, the PRIP homolog) in MTs was no different than the levels measured in freshwater reared larvae (Misyura et al., 2020). Furthermore, the protein abundance of at least three distinct aquaporins in the MTs, excluding AaAQP2 which was not assessed, were consistent between freshwater and brackish water reared larvae (Misyura et al., 2020). Relatedly, fluid secretion rates of MTs from brackish water mosquito larvae were found to be comparable to those from freshwater larvae (Donini et al., 2006). Similarly, the MTs from C. riparius larvae reared in SCW (NaCl) had fluid secretion rates similar to those of AFW-reared larvae (Reinsborough and Donini, 2025). Interestingly, MTs from C. riparius larvae exposed to BBJD secreted fluid at rates 1.5 times those of MTs from AFW and SCW exposed larvae and there is some evidence that this aids in removing the excess K^+^ which the larvae encounter in the BBJD (Reinsborough and Donini, 2025). Collectively, the results from the mosquito and midge larvae suggest that the underlying expression of AQPs is sufficient to support different rates of fluid secretion; that other AQPs that have yet to be discovered may play a role in the increased fluid secretion rates observed in BBJD midge larvae; or that AQPs (including CrAQP2) may be regulated through post translational modifications. In terms of the latter, the phosphorylation state of AQPs in adult female mosquito MTs did not change after a blood meal despite the dramatic increase in fluid secretion that ensues (Kandel et al., 2022).
Anal papillae (AP)
4.2.2
CrAQP2 transcript as well as AaAQP2-like immunoreactive staining in the AP of midge larvae were detected. The AP are important osmoregulatory organs in mosquito and midge larvae, responsible for the active uptake of ions and osmotically obliged water from the external freshwater environment (Jarial, 1995; Nguyen and Donini, 2010; Marusalin et al., 2012). Transcript levels of several AQPs were measured in the AP of A. aegypti and AaAQP2 transcript abundance was high compared to other AQPs when larvae were held in freshwater (Marusalin et al., 2012; Akhter et al., 2017; Durant et al., 2021). Since AaAQP2 is the predominantly expressed water transporting AQP in the AP it is likely to be, at the very least, partially responsible for the observed water flux across the AP epithelium (Marusalin et al., 2012; Akhter et al., 2017; Durant et al., 2021). Therefore, CrAQP2 is also likely to be important in mediating water transport across the AP epithelium of C. riparius, since we found AaAQP2-like immunolocalization on the basal membrane of the midge AP (Fig. 2E).
With respect to the influence of de-icers, it is already known that when mosquito larvae are exposed to salinity, the AP undergo morphological and ultrastructural changes (Wigglesworth, 1938; Sohal and Copeland, 1966; Surendran et al., 2018). It appears that when larvae are exposed to NaCl, their AP decrease in size and the epithelium has fewer microvilli and membrane folds thereby reducing the surface area for transport (Wigglesworth 1938; Sohal and Copeland, 1966). However, when larvae are raised over at least 20 generations in diluted natural seawater the AP increase in size over subsequent generations (Surendran et al., 2018). Furthermore, the anal papillae can adjust their ion transport functions in response to salinity (Nguyen and Donini, 2010; Donini et al., 2007; Reinsborough and Donini, 2025). In A. aegypti, exposure to NaCl based salinity decreases Na^+^ uptake by AP and in C. riparius this leads to an apparent reversal of transport from Na^+^ uptake to Na^+^ secretion (Donini et al., 2007; Reinsborough and Donini, 2025). When C. riparius were exposed to BBJD which contains relatively less NaCl, the AP responded similarly to those in the mosquito by decreasing, but not reversing, Na^+^ uptake (Reinsborough and Donini, 2025).
In this study, SCW caused a significant increase in CrAQP2 transcript abundance in midge AP while BBJD exposure did not change CrAQP2 transcript levels. The effects of increased NaCl-based salinity such as brackish water exposure caused similar findings in A. aegypti larvae, where a significant increase in AaAQP2 protein was determined in mosquito AP, relative to the freshwater control (Durant et al., 2021). The haemolymph of C. riparius remains hyperosmotic to the surrounding SCW (haemolymph: ∼ 270 mOsmol versus SCW: ∼ 240 mOsmol; Reinsborough and Donini, 2025); however, the gradient at which water entry is favoured is decreased with the increased external osmolarity from a difference of ∼ 160 mOsmol in AFW to ∼30 mOsmol in SCW (Reinsborough and Donini, 2025). In BBJD, the difference between haemolymph (hyperosmotic to BBJD) and the BBJD is ∼ 80 mOsmol (Reinsborough and Donini, 2025). Therefore, it stands to reason that a greater influx of water across the AP is occurring in BBJD relative to SCW conditions, and the level of water influx in BBJD may be sufficient to deal with the increased fluid secretion by the MTs in BBJD (Reinsborough and Donini, 2025). In contrast, the lower influx of water at the AP that would be caused by the reduced osmotic gradient in SCW may not be sufficient to avoid net water loss with the MTs maintaining rates of fluid secretion similar with those in AFW. This may be the reason why larvae in SCW have increased expression of CrAQP2 in the AP, so as to increase the influx of water when faced with a lower osmotic gradient in an attempt to avoid dehydration.
Midgut, gastric caeca, and hindgut
4.2.3
CrAQP2 transcript and AaAQP2-like immunoreactivity were evident in the MG and GC. Given that the foregut mainly serves as a conduit for food passage to the MG, and the absence of AaAQP2-like immunoreactivity in sections of the foregut, we suggest that the CrAQP2 expression in the GC-FG samples is confined to the GC. In the hindgut, CrAQP2 transcript was detectable but at relatively low levels and AaAQP2-like immunoreactivity was not observed.
In A. aegypti, AaAQP2 transcript was detected in the midgut of larvae and adults, and AaAQP2 was immunolocalized to the basolateral epithelial membrane of the posterior midgut in adults (Drake et al., 2010; Marusalin et al., 2012; Picinic et al., 2024). The tissue sections probed in this study showed AaAQP2-like immunoreactivity mostly confined to the apical (lumen facing) side of the C. riparius MG. We believe that most of the sections probed were from the anterior regions of the MG and cannot draw any conclusions about the localization of AaAQP2-like staining in the posterior MG. In some adult mosquitoes such as A. aegypti (Drake et al., 2010; 2015) and Anopheles sinensis (Zhao et al., 2024), entomoglyceroporins that transport solutes preferentially over water are more highly expressed in the MG presumably aiding in absorption of solutes into the haemolymph. At this stage, information on the identity or expression of other AQPs in C. riparius midgut remains unknown, although it is likely that entomoglyceroporins are present and expressed in the midge. The midgut of insects has a peritrophic membrane which isolates the food bolus from direct contact with the epithelial cells. As the food progresses posteriorly, enzymes are secreted, and digestion occurs. Fluid then travels anteriorly within the space between the peritrophic membrane and the epithelium where absorption of nutrients can occur (Ramsay, 1950; Volkmann and Peters, 1989). In C. riparius, CrAQP2 in the MG may contribute to sustaining this flow of fluid, supporting digestive and absorptive processes and/or mediating absorption of water into the haemolymph.
We also found increases in CrAQP2 transcript abundance in the MG with SCW treatment relative to AFW; however, no changes were seen with BBJD treatment. As larvae imbibe their external medium when feeding, the resulting osmotic gradient across the midgut would be greatly reduced in SCW resulting in less water absorption into the haemolymph (Reinsborough and Donini, 2025). Larvae face an accumulation of NaCl in SCW. If CrAQP2 is involved in water uptake across the MG in the midge, it is possible that in response to SCW exposure, the larval midges upregulate CrAQP2 levels in the MG to increase the uptake of water into the haemolymph helping to avoid dehydration caused by excess ion uptake and the reduction in the osmotic gradient that drives water towards the haemolymph. In BBJD, the osmotic gradient is not reduced to the same levels as in SCW and therefore it may be unnecessary to increase CrAQP2 expression to maintain sufficient water uptake into the haemolymph across the MG (Reinsborough and Donini, 2025).
Some of the fluid that moves anteriorly between the peritrophic membrane and epithelium of the midgut enters the gastric caeca, sites that are implicated in the final digestion of proteins and carbohydrates as well as ion and solute exchange with the haemolymph (see Godoy et al., 2023). In the GC of C. riparius, CrAQP2 localized on the basolateral membrane of epithelial cells can regulate water exchange with the haemolymph perhaps facilitating digestion and absorption of nutrients. In larval mosquitoes reared in freshwater, the GC move Na^+^ and K^+^ from the haemolymph into the GC lumen (D’Silva et al., 2017). Along with active ion transporters, AaAQP2 transcript (and other AQPs) are expressed in the GC of freshwater larval A. aegypti (Patrick et al., 2006; Misyura et al., 2020). In light of these facts, it is possible that water from the haemolymph is osmotically obligated to follow ion transport, similar to what occurs at the MTs. If this is the case, the GC may help to rid the larvae of excess water when they are in a freshwater environment. Assuming the GC of C. riparius function similarly to those in the mosquito, CrAQP2 could facilitate this movement of water. However, at this time, we do not know what the osmolality of the GC luminal contents are, and this information is necessary to draw a conclusion on this point.
The BBJD and SCW treatments did not affect the abundance of CrAQP2 transcript in the larval midge GC. It is plausible that the GC consistently secretes ions and water into their lumen irrespective of changes in external salinity. In A. aegypti larvae reared in brackish water, there is a reduction in ATPase activity and overall preference for secreting Na^+^ over K^+^ across the caecal epithelium (D’Silva et al., 2017). However, it was demonstrated that the transepithelial potential, as well as the sum of cation (Na^+^ and K^+^) concentrations, did not change in the caeca of brackish water larvae. Furthermore, AaAQP2 mRNA abundance in larval A. aegypti GC did not change following exposure to brackish water (Misyura et al., 2020). This suggests that similar rates of water secretion occur; however, since the mosquito larvae ingest the water they live in, the osmotic gradient across the GC of brackish water larvae may be significantly lower than freshwater larvae if that brackish water enters the GC lumen without much modification (D’Silva et al., 2017). A similar scenario is likely occurring in the GC of C. riparius larvae exposed to SCW or BBJD, where CrAQP2 expression remains consistent relative to freshwater exposed larvae.
In this study, we did not detect high abundance of CrAQP2 in the HG, which is consistent with previous studies on A. aegypti larvae that reported extremely low levels of AaAQP2 in the HG, relative to the other AQP isoforms expressed in this organ (Marusalin et al., 2012). Due to their freshwater environment, the HG is likely working to actively re-uptake ions while leaving water in the final urine destined for excretion (Bradley and Phillips, 1977; Jonusaite et al., 2013); therefore, it suggests that CrAQP2 may not play a prominent role in the HG of C. riparius larvae. Despite this low baseline transcript abundance and absence of detectable immunoreactivity in AFW, we measured the effects of BBJD and SCW on CrAQP2 expression in the HG of C. riparius. De-icer treatment significantly decreased the already very low levels of CrAQP2 in the HG, relative to AFW and immunohistochemistry revealed little to no presence of CrAQP2 in the HG. Therefore, water flux across the HG epithelium is likely to be low so that excess water remains in the urine for excretion. However, further investigation into potentially other AQPs present is essential to understanding the role of the HG.
Role of CrAQP2 in total body water content with de-icer treatment
4.3
To understand the role of CrAQP2 in the response of C. riparius larvae to de-icer treatment on a whole organism level, we decreased the expression of CrAQP2 in AFW larvae using RNA interference. We then subjected these larvae to an abrupt, acute exposure SCW or BBJD, in comparison to control larvae maintained AFW, in an attempt to mimic what they may encounter in their natural environment as a result of salinization. After the exposure, total body water was measured to assess the contribution of CrAQP2 to the osmotic regulation of larvae. Decreasing the expression of CrAQP2 significantly decreased the total body water of larvae, regardless of treatment (including controls).
In AFW larvae, we reason that the AP are predominantly responsible for the reduction in body water following CrAQP2 knockdown because the AP are sites of water uptake in dilute conditions and moderately express CrAQP2 transcript (Fig. 2; Marusalin et al., 2012). It is likely that other AQPs are expressed in the AP, however; if the AP of C. riparius are similar to the AP of A. aegypti then expression of other AQPs is low (Durant et al., 2021). Therefore, decreased CrAQP2 expression in the AP is expected to significantly reduce water uptake. The MG may also contribute to reduced water uptake when CrAQP2 expression is compromised assuming this transporter mediates water absorption across the MG (see explanations above, Section 4.2.3). On the other hand, any reduction in CrAQP2 expression in the MTs or GCs would likely cause the accumulation of water in the body since both of these organs are normally responsible for secreting water. However, it is possible that the MTs continue to secrete primary urine at a consistent rate even with CrAQP2 knockdown because of the likelihood of other AQPs expressed by the MTs. For example, in the mosquito A. aegypti, four of the six AQP genes have been localized in the MTs (Drake et al., 2010; Marusalin et al., 2012; Misyura et al., 2020; Picinic et al., 2024). This suggests a model where water transport may occur through many AQPs simultaneously and therefore the reduction in CrAQP2 expression alone may not have a significant effect on the overall function of the MTs if this were the case. As a result, this suggests that the lower body water observed with CrAQP2 knockdown could be driven by the decreased capacity for water uptake at the AP.
Larvae exposed to de-icers face an osmotic gradient across the body wall which favours water movement into the haemolymph; however, this gradient is much lower than what they face in freshwater, particularly in SCW (see Section 4.2.2). Therefore, less water is obliged to enter through the AP and/or MG and the larval osmoregulatory physiology responds by increasing transcript abundance of CrAQP2 in AP and MG in SCW exposure (Fig. 4) to increase water uptake (see Sections 4.2.2 & 4.2.3). CrAQP2 knockdown under these conditions attenuates the physiological response resulting in insufficient water being taken up across AP and/or MG, and since the MTs likely continue to produce primary urine at similar levels, the larvae experience dehydration leading to significantly lower total body water content. Indeed, this is supported by recent evidence demonstrating that the MTs of larvae in SCW and BBJD secrete fluid at the same and higher rates than those in AFW, respectively (Reinsborough and Donini, 2025), and the effect of CrAQP2 knockdown in MTs is likely to be blunted by the expression of other AQPs. Thus, the net result is similar to control larvae where dehydration occurs.
Effect of CrAQP2 knockdown on larval survival
4.4
With the knowledge that CrAQP2 knockdown causes dehydration in the midge larvae after just 24 h of exposure to de-icers and control conditions, we tracked survival of midges over 7 days. After 3 days, knockdown larvae in AFW showed lower survival than control larvae but at least half of the larvae survived to 7 days. We suggest that, although uptake of water across the AP would be compromised, as evidenced by lower body water, the osmotic gradient is sufficient to drive enough water into the body to permit survival. In contrast, in SCW where the osmotic gradient is reduced, the data suggests that not enough water enters the body and this manifests in an abrupt drop in survival beginning on day 3. In BBJD, the osmotic gradient may be sufficient to sustain enough water uptake, but this is likely countered by the increase in fluid secretion rates by the MTs causing a more gradual drop in survival beginning on day 3. In both de-icers, all larvae succumb to the salinization treatment by day 5.
In conclusion, this study identified the first AQP in C. riparius, which we named CrAQP2, and also measured its transcript abundance and localized its distribution at the protein level using immunohistochemistry in the alimentary canal of midge larvae. Furthermore, the influence of common road de-icers (BBJD and NaCl) on CrAQP2 expression in each of the osmoregulatory organs was observed and knockdown of CrAQP2 revealed the role of this water channel in mitigating osmotic stress associated with salinization. We found that CrAQP2 is expressed broadly in the midge alimentary canal and overall, NaCl-contaminated water caused differential gene expression in a few of the osmoregulatory organs while BBJD only affected CrAQP2 expression in the HG. However, with CrAQP2 knockdown, treatment of midge larvae with either BBJD or NaCl was lethal after only a few days. A reduction in CrAQP2 functioning associated with knockdown results in reduced body water for midge larvae in freshwater, SCW, and BBJD. From this study, it is evident that CrAQP2 is an important water channel protein in C. riparius, with a role in homeostatic water transport. Furthermore, salinization of freshwater influences CrAQP2 expression and its role in osmoregulation. CrAQP2 appears to be vital in the ability of C. riparius to adapt to and survive in increased salinity caused by road de-icer contamination.
Funding
This study was supported by Natural Sciences and Engineering Research Council of Canada Discovery Grants # RGPIN-2024-06188 to AD; and RGPIN-2020–06130 to JPP, Ontario Ministry of Colleges and Universities, Ontario Graduate Scholarship to BNP.
Credit author statement
B. Picinic: designed experiments, carried out experiments, analyzed data, wrote the manuscript. A. Reinsborough, carried out experiments. S. Jonusaite, carried out experiments. J-P. Paluzzi: designed experiments, edited the manuscript. A. Donini: designed experiments, edited the manuscript, obtained funding
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akhter H.Misyura L.Bui P.Donini A.Salinity responsive aquaporins in the anal papillae of the larval mosquito, Aedes aegypti Comp. Biochem. Physiol.203201714415110.1016/j.cbpa.2016.09.00827642095 · doi ↗ · pubmed ↗
- 2Azuma M.Nagae T.Maruyama M.Kataoka N.Miyake S.Two water-specific aquaporins at the apical and basal plasma membranes of insect epithelia: molecular basis for water recycling through the cryptonephric rectal complex of lepidopteran larvae J. Insect Physiol.58420125235332228568610.1016/j.jinsphys.2012.01.007 · doi ↗ · pubmed ↗
- 3Bradley T.J.Phillips J.E.Regulation of rectal secretion in saline-water mosquito larvae living in waters of diverse ionic composition J. Exp. Biol.661977839687060410.1242/jeb.66.1.83 · doi ↗ · pubmed ↗
- 4Cabrero P.Terhzaz S.Dornan A.J.Ghimire S.Holmes H.L.Turin D.R.Romero M.F.Davies S.A.Dow J.A.T.Specialized stellate cells offer a privledged route for rapid water flux in Drosophila renal tubule Proc. Natl. Acad. Sci. USA 11732020177917873190732110.1073/pnas.1915943117 PMC 6983416 · doi ↗ · pubmed ↗
- 5Campbell E.M.Ball A.Hoppler S.Bowman A.S.Invertebrate aquaporins: a review J. Comp. Physiol.17820089359551859483510.1007/s 00360-008-0288-2 · doi ↗ · pubmed ↗
- 6Chasiotis H.Ionescu A.Misyura L.Bui P.Fazio K.Wang J.Patrick M.Weihrauch D.Donini A.An animal homolog of plant Mep/Amt transporters promotes ammonia excretion by the anal papillae of the disease vector mosquito Aedes aegypti J. Exp. Biol.21992016134613552694449610.1242/jeb.134494 · doi ↗ · pubmed ↗
- 7Clark T.M.Bradley T.J.Malpighian tubules of larval Aedes aegypti are hormonally stimulated by 5-hydroxytryptamine in response to increased salinity Arch. Ins. Biochem. Phys.3421998123141
- 8Clark T.M.Koch A.Moffett D.F.The anterior and posterior ‘stomach’ regions of larval Aedes aegypti midgut: regional specialization of ion transport and stimulation by 5-hydroxytryptamine J. Exp. Biol.2021999247252988263710.1242/jeb.202.3.247 · doi ↗ · pubmed ↗