Circadian ADCY3 Ser107Pro variant bridges difficulty awakening in the morning and adiposity
Cynthia Tchio, Matthew Maher, Christopher Moth, Jens Meiler, Jacqueline M. Lane, Herman A. Taylor, Jonathan S. Williams, Richa Saxena

TL;DR
A genetic variant in ADCY3 is linked to morning difficulty awakening and increased body fat, connecting circadian rhythms to metabolic health.
Contribution
Identifies a novel ADCY3 variant that links circadian preference and adiposity through genetic and molecular mechanisms.
Findings
The ADCY3 Ser107Pro variant increases eveningness, BMI, and fat mass in multiple ancestry groups.
The variant alters adipose-specific splicing and destabilizes ADCY3 protein structure.
BMAL1 binds near residue 107 in mouse adipose tissue, suggesting a conserved regulatory mechanism.
Abstract
Modern lifestyles often disturb circadian rhythms, yet the genetic circuits that convert this stress into metabolic dysfunction remain poorly defined. Here, we identify a missense variant in ADCY3 (rs11676272; Ser107Pro) as a pleiotropic regulator of circadian preference and adiposity. Using genome-wide pleiotropy analysis in ∼480,000 UK Biobank participants, we show that the G risk allele (Pro107) increases eveningness, BMI, and fat mass in European (n = 451,324) and African (n = 8,738) ancestry groups, with behavioral amplification by morning difficulty awakening in Europeans and power-limited modeling in other populations. Structural modeling and transcriptomic analysis suggest this allele alters adipose-specific splicing and expression and destabilizes ADCY3 protein. In mice, Adcy3 is rhythmically expressed in adipose tissue, with BMAL1 binding near the orthologous residue 107 site.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
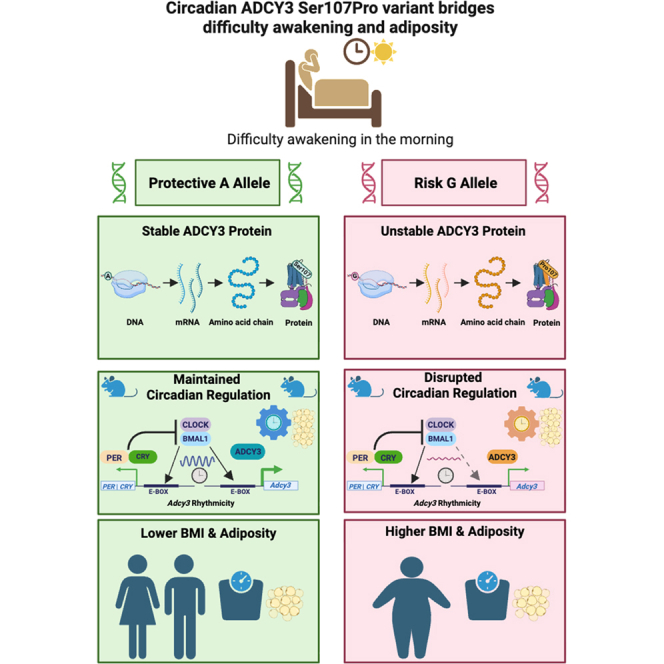 Figure 1
Figure 1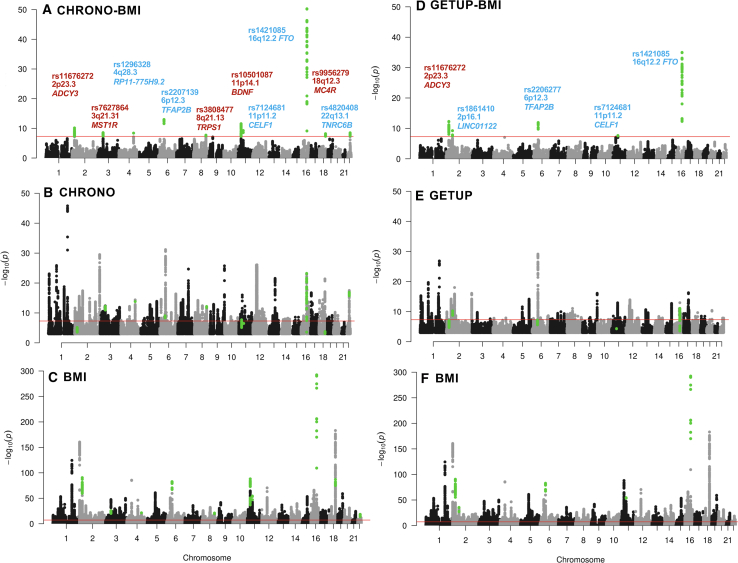 Figure 2
Figure 2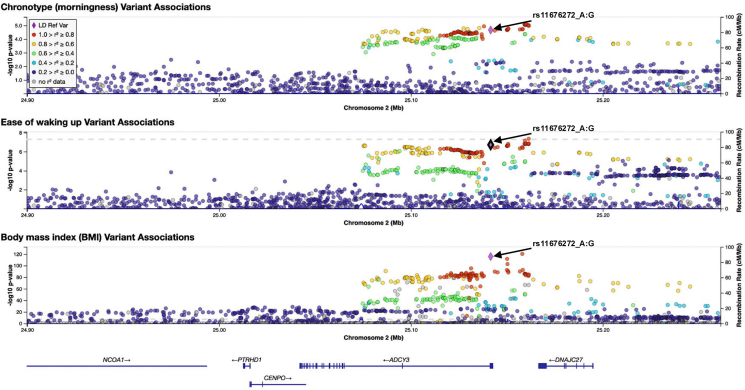 Figure 3
Figure 3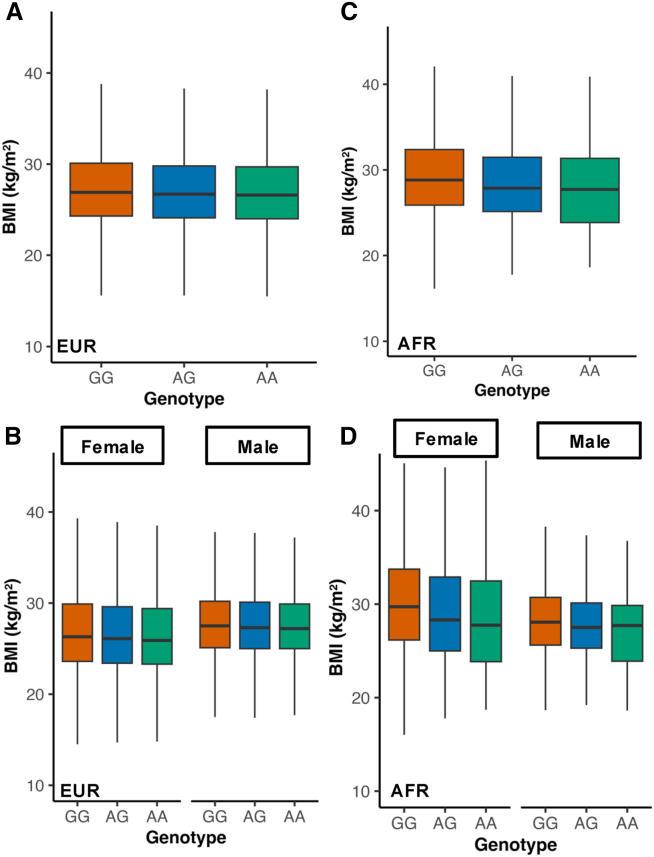 Figure 4
Figure 4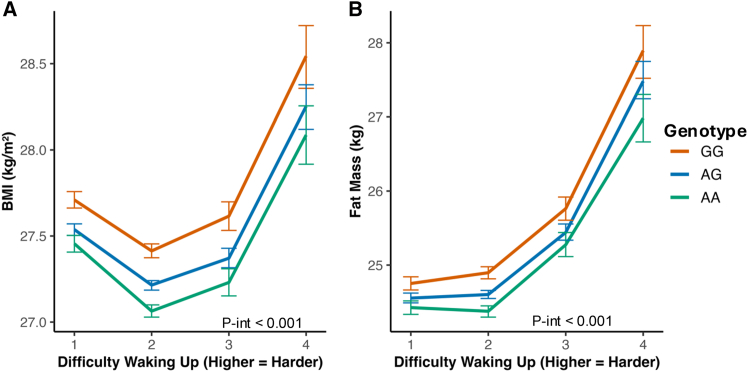 Figure 5
Figure 5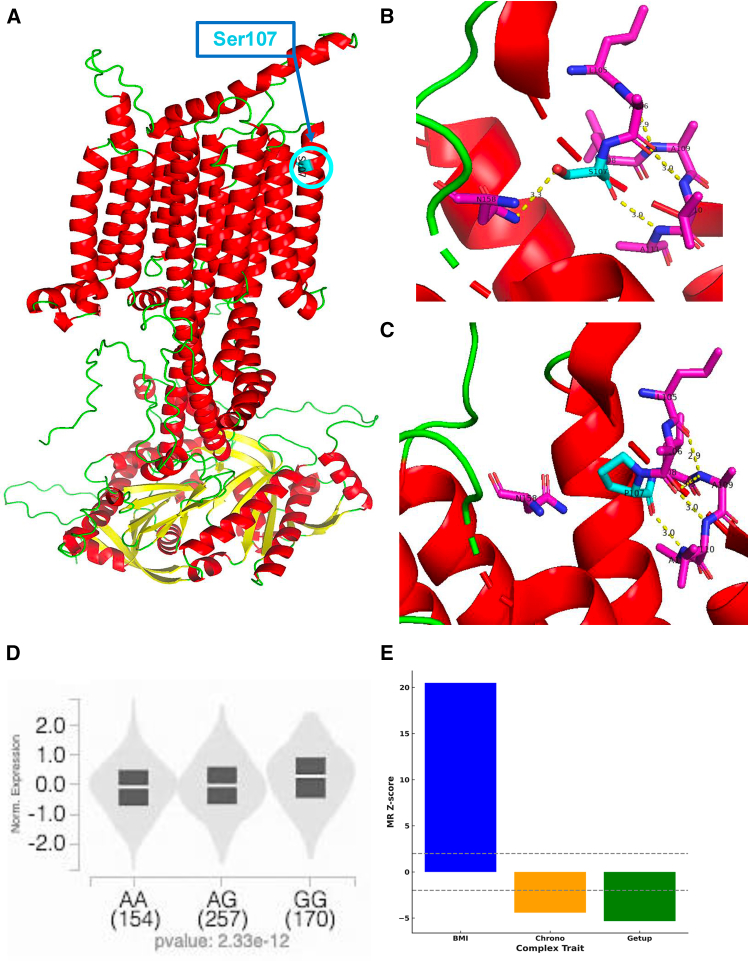 Figure 6
Figure 6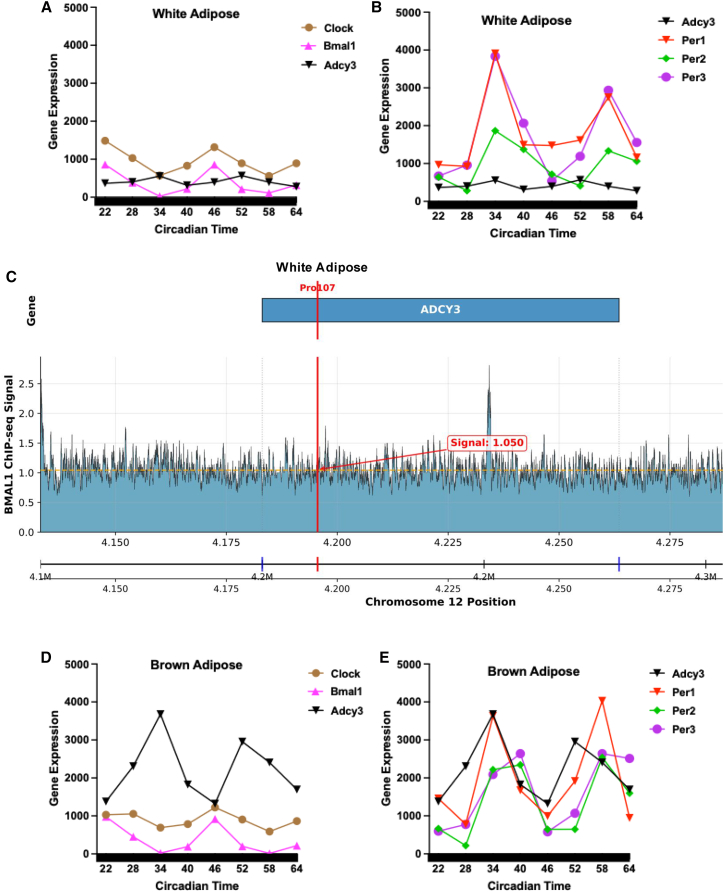 Figure 7
Figure 7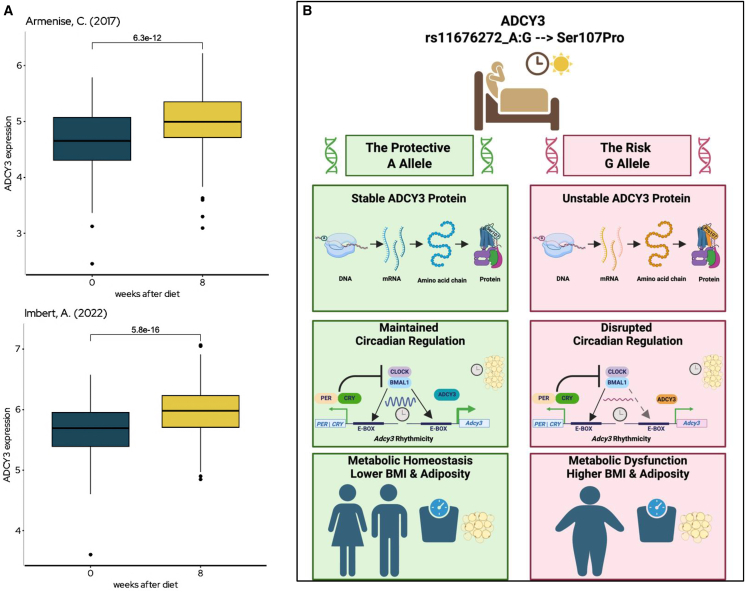 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircadian rhythm and melatonin · Dietary Effects on Health · Genetic Associations and Epidemiology
Introduction
The circadian clock governs a wide range of physiological processes, including sleep-wake cycles, metabolism, and energy homeostasis.1^,^2 Chronotype reflects an individual’s behavioral preference for morning or evening activity, summarizing the phase of the circadian system as shaped by both environmental and genetic factors.3 Genome-wide association studies (GWASs) have identified 351 loci for chronotype; many of which colocalize with loci for body mass index (BMI) and fat mass, pointing to molecular crosstalk between circadian and metabolic regulation.1^,^3^,^4^,^5^,^6^,^7
A related but less-studied trait is self-reported difficulty waking up in the morning, which reflects a person’s ability to arouse from sleep on schedule. Although genetically correlated with chronotype,6 recent work suggests that this trait captures a distinct dimension of circadian strain, potentially linked to misalignment and impaired arousal regulation.8 Understanding how genetic variation modifies this trait could reveal clock-controlled metabolic pathways that are particularly sensitive to behavioral disruption.
The canonical clock genes offer proof of principle, where the deletion of Clock or Bmal1 (Arntl) in mice leads to obesity and glucose intolerance9^,^10 and clock output in brown and white adipose tissue gates lipid mobilization and thermogenesis.11 However, for the vast majority of human GWAS signals, the causal alleles, effector genes, and tissue-specific mechanisms remain unresolved. Notably, no prior study has identified a single coding variant that links a circadian behavioral trait and an adipose-centric metabolic phenotype through functional validation in relevant tissues.
Here, we use a genome-wide pleiotropy approach to bridge this gap. By jointly analyzing GWAS summary statistics for morning chronotype, difficulty waking in the morning, and BMI in ∼480,000 UK Biobank participants of European, African, and south Asian ancestry, we identify a shared missense signal at ADCY3 (rs11676272; Ser107Pro). We show that the BMI-raising G allele (Pro107) has a stronger effect in individuals who report difficulty waking, suggesting a gene-by-behavior interaction. To dissect the mechanism, we integrate in silico protein modeling, human eQTL/splice-QTL/RNA-seq, and mouse circadian RNA-seq into our analytical pipeline. In humans, the G allele destabilizes ADCY3 protein and also acts as an eQTL/sQTL in adipose tissue, where it is paradoxically associated with higher expression, potentially reflecting compensatory regulation. In mice, Adcy3 is rhythmically expressed in adipose, and the genomic region orthologous to residue 107, a conserved position, is a BMAL1-binding target. Furthermore, human adipose RNA sequencing (RNA-seq) from weight-loss interventions shows that ADCY3 expression increases in metabolically improved states, reinforcing its role in energy balance.
Together, these findings position ADCY3 Ser107Pro as a molecular bridge between a daily arousal trait and adipose physiology, illustrating how common genetic variation may contribute to the metabolic consequences of circadian strain.
Results
Genome-wide pleiotropy analysis identifies shared loci between circadian preference traits and BMI
To identify genetic signals influencing both circadian rhythms and BMI, we conducted a genome-wide pleiotropy analysis using the Pleiotropic Analysis under Composite Null Hypothesis v0.1.1 (PLACO) composite-null test12 with a genome-wide pleiotropy significance threshold of P.PLACO <5 × 10^−8^; and HyPrColoc colocalization13 starting from GWAS summary statistics for morningness chronotype4 (n = 449,734); difficulty waking up in the morning (n = 451,872; both UKB Europeans), and BMI (n = 694,649; UKB and GIANT consortium;14 Table S1). We identified ten pleiotropic loci for morningness chronotype (Chrono) and BMI (Figure 1A; Table S2) and five loci for difficulty waking up in the morning (Getup) and BMI (Figure 1D; Table S3). Four loci (in or near ADCY3, CELF1, TFAP2B, and FTO) were common to both analyses. Regional colocalization analysis using a different approach (HyPrColoc13) confirmed that the same single-nucleotide polymorphisms (SNPs) underlie associations across all three traits, with evidence of shared regulation across all three traits: morningness (Chrono), difficulty waking (Getup), and BMI (posterior probability, PP > 0.80, Table 1). Notably, pleiotropic variants associated with higher BMI are associated with either evening chronotype (ADCY3) or morning chronotype (TFAP2B, CELF1, and FTO). Phenome-wide association analysis (PheWAS) revealed that all four pleiotropic loci are also significantly associated with fat mass, highlighting their role in adiposity regulation (Table S4). Furthermore, in silico assessment revealed that the lead loci reside in active regulatory regions, demonstrated by tissue-specific chromatin interactions (Figure S2) and significant eQTL effects (Table S5).Figure 1. Genome-wide analysis identifies shared genetic loci between circadian traits and BMIManhattan plots showing genome-wide pleiotropy for (A) morningness chronotype × BMI and (D) difficulty waking up in the morning × BMI. Single-trait GWAS plots shown for (B) chronotype, (E) difficulty waking up in the morning, and (C and F) BMI. The red line marks the genome-wide significance threshold (p < 5 × 10^−8^). In (A and D), pleiotropic loci are annotated with the nearest gene; red highlights loci with opposite directions of effect across traits, while blue indicates shared directionality.Table 1. Colocalization identifies shared candidate causal loci for morning circadian preference traits and BMIGeneTraitsPosterior PRegional PCandidate SNPAnnotationP by SNPADCY3Chrono, Getup, BMI0.8430.924rs11676272Missense1.000CELF1Chrono, Getup, BMI0.6150.662rs7124681Intronic1.000FTOChrono, Getup, BMI0.9721.000rs1558902Intronic0.917TFAP2BChrono, Getup, BMI0.9910.999rs2206277Intronic0.828Colocalization analyses were conducted using HyPrColoc (Hypothesis Prioritization in Multi-Trait Colocalization) and GWAS summary statistics (Chrono: Morningness Chronotype; Getup: difficulty waking up in the morning; BMI: body mass index). “Posterior P” (posterior probability of colocalization) assumes that a locus contains 1 causal variant, whereas “Regional P” assumes that a locus contains >1 causal variant. “P by SNP” is the % contribution of the candidate (causal) variant to the posterior p.
A missense variant in ADCY3 is the top candidate for shared causality
Of the four pleiotropic loci, we focus on rs11676272 in ADCY3, a missense variant (Ser107Pro), because it showed a strong colocalization across all three traits, with a PP of 0.84, with the SNP contributing to 100% of the PP (Table 1). Specifically, the G allele, which encodes for proline-107, associates with eveningness (Chrono β = −0.0118 ± 0.0026), more difficulty waking up in the morning (Getup β = −0.0084 ± 0.00157), and higher BMI (β = 0.0328 ± 0.002); the regional plots for all three traits are shown in Figure 2. This SNP was prioritized not only for its robust statistical signal but also because its dual potential to alter ADCY3 protein structure while also regulating gene expression provided a unique opportunity to investigate multi-level gene function. Population frequency analysis revealed that rs11676272 varies across ancestries (33%–85% in GnomAD), with the ancestral G allele being most prevalent in African populations (Figure S11A). The A allele frequency varies across populations and appears to have undergone recent positive selection (Figure S11B), as indicated by Tajima’s D statistics (AFR, −1.33; EAS, −1.44; EUR, −0.71), consistent with adaptation to cold environments.Figure 2. Colocalization analysis supports rs11676272 as a shared causal variant for circadian traits and BMI at the ADCY3 locusRegional plots for (top) chronotype, (middle) difficulty waking, and (bottom) BMI. The lead pleiotropic missense SNP rs11676272 is marked with a purple diamond. Colors reflect linkage disequilibrium (LD, measured as r^2^) with rs11676272, based on the 1,000 Genomes EUR reference panel. Colocalization analysis (HyPrColoc) supports a shared causal variant across all traits (posterior probability, PP = 0.84; regional PP = 0.92; SNP PP = 1.00). Genomic region spans chr2:24,992,038–25,193,106 (GRCh37).
The ADCY3 risk allele’s effect on adiposity is sexually dimorphic and is amplified by difficulty awakening in the morning
To dissect the lead pleiotropic variant, rs11676272, we performed linear mixed models using lme415 in the UK-Biobank cohort. The G risk allele, previously associated with eveningness, showed a dose-response with higher BMI (Figures 3A and 3C), fat mass, and body-fat percentage (Figures S3A and S3B). The effect was largest in Europeans (AA vs. GG β = −0.32 kg m^−2^; p < 2 × 10^−16^) and consistent in Africans, though less precise in south Asians (Table S8). Thus, each A allele lowers BMI relative to the G risk allele.Figure 3. The ADCY3 risk allele G associates with higher BMI in both Europeans and Africans, with stronger effect in women(A) In Europeans (n = 451,324), BMI decreases with each additional A allele (AA vs. GG = −0.32 kg/m^2^, p < 2 × 10^−16^).(B) Sex-stratified analysis shows a stronger protective effect in females (AA vs. GG = −0.38) than males (−0.24), with significant gene-by-sex interaction (p = 2.4 × 10^−14^).(C) In Africans (n = 8,738), AG carriers have lower BMI than GG (effect size β = −0.38, p = 0.009); AA is underpowered.(D) Sex-stratified analysis in Africans shows a similar female-greater trend. Boxplots: center = median; box = interquartile range IQR; whiskers = 1.5 × IQR. Colors, GG (blue), AG (green), AA (orange).
A clear sex-dimorphic pattern emerged: the protective A allele produced a larger reduction in females (AA vs. GG β = −0.38 kg m^−2^) than in males (β = −0.24 kg m^−2^; p for interaction = 2.4 × 10^−14^; Figure 3B). The same female-greater trend was observed in Africans (Figure 3D).
We next asked whether self-reported “difficulty awakening in the morning” modifies this genetic effect. In Europeans, we observed a significant interaction for both BMI and fat mass (Figures 4A and 4B). The positive association between difficulty waking and higher adiposity was strongest in G-allele carriers and attenuated in A-allele homozygotes. No significant interaction was observed for chronotype (Figure S4; Table S8) or for night shift work (n = 42,486; all p > 0.1), underscoring the specificity of the morning-waking trait.Figure 4. The obesogenic effect of the ADCY3 risk allele is amplified by difficulty waking up in the morningInteraction analysis between rs11676272 genotype and waking difficulty in UK Biobank Europeans (n = 451,324).(A) Effect on BMI.(B) Effect on fat mass. Genotype-stratified slopes show that the association between waking difficulty and adiposity is strongest in GG carriers and attenuated in AA homozygotes. Interaction analyses are reported in Table S8, confirming significant genotype × waking-difficulty interactions in AA homozygotes for BMI (p = 8.0 × 10^−4^) and fat mass (p = 2.1 × 10^−3^). Models adjusted for age, sex, PC1-10, and kinship. Error bars = 95% CI.
For African and south Asian participants, the allele showed the same protective direction in heterozygotes (AG vs. GG, β = −0.38 kg m^−2^, p = 0.009 in Africans; β = −0.07 kg m^−2^, p = 0.568 in south Asians) (Table S8). Sample-size differences explain the wider confidence intervals in these groups: Europeans contribute 119,499 AA homozygotes, 141 for Africans, and 2,062 for south Asians (Figure S5).
Together, these data demonstrate that rs11676272 influences adiposity in a sex-dimorphic manner, with AG individuals showing intermediate effects, and that in Europeans the G allele obesogenic impact is exacerbated by difficulty waking in the morning, a behaviorally tractable facet of circadian physiology.
The rs11676272 variant has multi-level functional consequences
To elucidate the molecular mechanisms of rs11676272, we first used AlphaFold-based structural modeling to predict the impact of the Ser107Pro missense mutation on ADCY3 protein architecture (Figure 5A). The protective A allele encodes serine (Ser107), which form stable hydrogen bonds with asparagine at residue 158 (N158 or Asn 158) and supports a more stable protein conformation (Figure 5B). In contrast, the G allele, which encodes proline at residue 107 (Pro107) is predicted to disrupt these stabilizing hydrogen bond with Asn 158 (N158) and destabilizes the ADCY3 protein structure (Figure 5C) as indicated by ΔΔG = 3.61 kcal/mol. In addition to the protein-altering effect, we found that rs11676272 is a significant expression and splicing quantitative trait locus (eQTL and sQTL) for ADCY3 in subcutaneous adipose tissue (Figures 5D and S7A). Consistent with this, tissue-level isoform analysis across GTEx v8 (Figure S10A) shows that adipose tissue expresses a distinct set of ADCY3 isoforms compared to other tissues such as liver, hypothalamus, and cerebellum. This provides a biological basis for the tissue-specific splicing effects associated with rs11676272. The risk allele G was paradoxically associated with higher ADCY3 expression, potentially reflecting a compensatory transcriptional response to impaired protein stability or altered feedback regulation. Mendelian randomization Wald ratio test supported a causal relationship between this altered ADCY3 expression and the observed traits (Figure 5E). The variant’s location within an active regulatory element in adipose tissue, confirmed by histone markers H3K4me1 and H3K4me3 (Figure S6), and binding site to multiple transcription factors, including RNA Polymerase II POLR2A (Table S7), provides a strong mechanistic basis for its regulatory effects.Figure 5. The rs11676272 variant has multi-level functional consequences on protein stability and gene expression(A) The AlphaFold predicted the full protein structure of ADCY3, with the location of serine 107 (S107) highlighted in a transmembrane (TM2) helix.(B) A zoomed-in view of Ser107 shows that the hydroxyl oxygen of S107 forms a hydrogen bond (3.3 Å) with the backbone amide nitrogen of asparagine 158 (N158), and the backbone amide nitrogen of A111 forms a hydrogen bond (3.0 Å) with the hydroxyl oxygen of S107.(C) The rs11676272 G allele encodes Pro107, disrupting the stabilizing bond with N158 and increasing predicted protein instability (ΔΔG = 3.61 kcal/mol). The protective A allele encodes Ser107, which preserves these interactions.(D) In addition to its effect on the protein, the rs11676272 G allele is a significant expression quantitative trait locus (eQTL) associated with higher ADCY3 expression in subcutaneous adipose tissue (p = 2.33 × 10^−12^).(E) Mendelian randomization Wald ratio method analysis supports a causal relationship between this altered ADCY3 expression and higher BMI, eveningness chronotype, and more difficulty waking up.
Adcy3 is rhythmically expressed in mice adipose tissue, and BMAL1 binds the genomic region orthologous to residue 107 of Adcy3
Given ADCY3’s link to circadian and adiposity traits in humans, we queried the human ADCY3 sequence and identified multiple canonical BMAL1 E-box motifs (CACGTG) across the promoter and gene body. As shown in Figure S10B, the NCBI Variation Viewer highlights these BMAL1-binding motifs (blue lines) together with the rs11676272 (Ser107Pro) variant (pink line) within the coding region, supporting the presence of potential circadian regulatory elements at this locus.
We then investigated the physiological regulation of Adcy3 in mouse adipose tissue. Using publicly available RNA-seq datasets, we found that Adcy3 expression was robustly rhythmic in both white (WAT; Figures 6A and 6B) and brown (BAT; Figures 6D and 6E) adipose tissue, oscillating in antiphase to the core clock activator Bmal1 and in phase with the Per1-3 repressors (Figures 6B and 6D). This expression pattern is consistent with the canonical circadian feedback loop, in which BMAL1 and CLOCK activate transcription of the Per genes, whose protein products accumulate and inhibit BMAL1/CLOCK activity in a near 24-h cycle. In contrast, Adcy3 expression was not rhythmic in the hypothalamus, cerebellum, or liver (Figure S8), underscoring the adipose-specific nature of its circadian regulation.Figure 6Adcy3 shows rhythmic expression in mouse white and brown adipose tissue and BMAL1 binding at the orthologous locus(A and B) Adcy3 rhythmically oscillates (meta2d p = 3.8 × 10^−4^) in antiphase to Clock and Bmal1 and in phase with Per1/Per2/Per3 over a 42-h circadian cycle in mouse white adipose (n = 1/timepoint) (C) BMAL1 ChIP-seq in inguinal white adipose tissue shows binding at the Adcy3 locus, near the orthologous residue 107 site which harbors the Ser107Pro variant from human.(D and E) Rhythmic gene expression of Adcy3 and core clock genes in brown adipose tissue over a 42-h circadian cycle (meta2d p = 6.1 × 10^−4^, n = 1/timepoint) shows a similar phase relationship to Bmal1 and Per genes, indicating coordinated rhythmic regulation across adipose depots.
To establish a direct mechanistic link, we analyzed BMAL1 ChIP-seq data from mouse WAT, which revealed binding peaks at the Adcy3 locus (Figure 6C), including one overlapping the genomic region orthologous to residue 107. This residue position is evolutionarily conserved across mammals, although human populations vary in allele frequency at this site (the ancestral G allele encodes proline, while the derived A allele encodes serine). These data establish Adcy3 as a direct transcriptional target of BMAL1. Additionally, we found that Adcy3 expression is cold-induced in iWAT of control mice (10°C vs. 22°C). In Bmal1 knockout mice, baseline expression is elevated and the relative cold response appears attenuated, suggesting BMAL1 may contribute to cold-induced regulation of Adcy3 (Figure S9). Consistent with this, recent work has demonstrated that ADCY3 isoforms regulate brown adipose thermogenesis directly.16 Together, these findings suggest that Adcy3 is a rhythmic clock output gene in adipose tissue and may contribute to cold-induction thermogenic pathways.
ADCY3 expression in human adipose tissue is a marker of metabolic health
Finally, to assess the clinical relevance of these findings in humans, we analyzed data from the Adipose Tissue Knowledge Portal.17 This analysis revealed that ADCY3 expression is significantly upregulated in human subcutaneous adipose tissue following successful diet-induced weight loss (Figure 7A), consistent with improved metabolic state. This pattern contrasts with the adipose eQTL signal of the G allele (Figure 5D), where the increased expression of ADCY3 might likely reflect compensatory upregulation in response to reduced protein stability (Figure 5C) rather than enhanced functional activity.Figure 7A unified model for ADCY3 in circadian-metabolic integration and human adipose health(A) ADCY3 expression increases in subcutaneous adipose tissue following diet-induced weight loss (800-1,000 kcal/day) in two independent studies (Imbert et al., n = 558, p = 5.8e-12; Armenise et al., n = 191, p = 6.3e-12), consistent with improved metabolic state.(B) Schematic model, in AA homozygotes (left), ADCY3 is stable and functional, conferring resilience to circadian strain and supporting maintained circadian regulation of adipose function. In GG homozygotes (right), protein instability and impaired rhythmic response increase vulnerability to lifestyle stressors and promote higher adiposity.
These two independent observations18^,^19 suggest that functional ADCY3 activity, rather than transcript abundance alone, is a hallmark of healthy adipose tissue. Combined with our genetic and circadian data, these findings position ADCY3 as a circadian-regulated metabolic effector that integrates genetic, behavioral, and environmental cues to influence obesity risk.
Discussion
Obesity arises from a complex interplay of genetic factors, behavioral influences, and tissue-specific physiology. This study identifies the circadian clock-associated gene ADCY3 as a key genetic mediator of the metabolic consequences of lifestyle factors. We provide evidence from large-scale human genetics and mechanistic mouse studies that a common missense variant in ADCY3, rs11676272, interacts with difficulty waking up to significantly increase an individual’s risk of obesity. Our findings reveal a sex-dimorphic effect, a novel mechanism involving protein destabilization with compensatory transcript upregulation and a potential link to thermogenic regulation in adipose tissue, establishing ADCY3 as a critical node at the interface of circadian biology and metabolic health. Importantly, our study provides the first evidence that a common coding variant links a circadian behavioral trait with adipose regulation, thereby establishing a novel mechanistic connection between circadian strain and obesity risk.
A central finding of our work is the potentiation of genetic risk by a behavioral factor. The obesogenic effect of the G risk allele was most pronounced in individuals who reported difficulty waking up in the morning, a behavioral marker that correlates with BMI. To test the specificity of this finding, we examined other, broader measures of circadian life. Importantly, we observed no significant obesogenic interaction for innate chronotype preference or for the occupational category of night-shift work. This crucial distinction suggests that the biological pathway involving ADCY3 is not uniformly sensitive to all forms of circadian disruption. Rather, our results indicate that its vulnerability is selectively unmasked by increased adiposity, due to difficulty awakening in the morning.
The functional impact of the Ser107Pro mutation underscores the regulatory consequences of rs11676272. Structural modeling shows that the proline substitution leads to the loss of a stabilizing polar bond and predicts significant destabilization of the ADCY3 protein structure. While this may not impair catalytic function directly, it could influence ADCY3’s role in signaling complexes or membrane dynamics, consistent with prior in vitro assays showing a modest, though non-significant, reduction in cAMP production in Hek293 cell line.20 Notably, the variant also resides within an active promoter and enhancer region in adipose tissue and is associated with a paradoxical increase in ADCY3 expression. This raises the key possibility that rs11676272 functions as a dual-threat variant, simultaneously altering protein structure while its cis-regulatory effect drives a compensatory increase in transcript levels that is ultimately insufficient to restore normal protein function. This complexity is compounded by tissue-specific isoform usage; we find that adipose tissue expresses a distinct set of ADCY3 transcripts not found in other key metabolic organs, providing a compelling mechanism by which the rs11676272 sQTL can exert its tissue-restricted effects.
Our findings in mice provide a direct physiological context for this mechanism. We show that Adcy3 is a rhythmic gene specifically in adipose tissues, oscillating in antiphase to its direct transcriptional regulator, BMAL1. Importantly, BMAL1 ChIP-seq reveals a binding peak exactly over the murine Adcy3 segment orthologous to the human Ser107/Pro107 codon, indicating that the same cis-element harboring the missense variant is itself a bona-fide clock-controlled enhancer. This positions Adcy3 as a direct, rhythmic output of the core clock machinery in adipose tissue, tasked with integrating circadian signals with environmental cues. Furthermore, our analyses suggest a functional link to energy expenditure, as Adcy3 expression is cold-inducible in a BMAL1-dependent manner. While this might provide indirect support for circadian-thermogenic integration, additional functional studies will be required. We also note that the Prx1-Cre line used in these datasets drives recombination in mesenchymal precursor cells across multiple tissues, and is, therefore, not strictly adipose-specific.21^,^22 Thus, some of the observed effects in Bmal1fl/fl;Prx1-Cre mice could reflect influences outside adipose tissue. In addition, cold exposure in WAT may influence pathways beyond thermogenesis, including lipolysis, and therefore these findings should be interpreted with caution. Our use of these secondary mouse data was intended to provide mechanistic context for the human phenotype rather than definitive functional validation. Future studies using adipose-specific Cre lines or promoter-reporter assays will be required to directly test BMAL1 regulation of ADCY3 in adipose tissue. Notably, independent experimental work has demonstrated that ADCY3 isoforms modulate brown adipose thermogenesis,16 supporting the relevance of ADCY3 to thermogenic regulation. This regulatory mechanism parallels prior findings at the FTO locus, where obesity-associated variants regulate thermogenesis in a tissue-autonomous manner.23 Our discovery that full-length Adcy3 oscillates introduces a crucial circadian dimension to prior “rheostat” models of its function, such as that of the truncated isoform Adcy3-at,16 suggesting that the temporal gating of ADCY3 expression is a key mechanism for coordinating thermogenic sensitivity with daily metabolic rhythms.
The role of ADCY3 as a circadian-regulated metabolic effector involved in thermogenic adaptation is further illuminated by its place in human history, which speaks to the broader debate on the origins of obesity-related genetic variation. This debate often centers on two competing ideas: the “thrifty gene” theory,24 which posits that variants were positively selected for energy storage in environments of food scarcity, and the “drifty gene” theory,25 which argues that variants accumulated neutrally after predation pressures on fat regulation were relaxed. While neither has been definitively proven, our findings for ADCY3 align more closely with a model of positive, adaptive selection akin to the thrifty gene concept. Although residue 107 is conserved across mammals, human populations show variation in allele frequency: the ancestral G (Pro) allele is most common in African populations, whereas the derived A (Ser) allele is more frequent in Europeans and south Asians, consistent with genetic drift and/or recent selection. Specifically, the protective A allele of rs11676272, which we associate with favorable metabolic outcomes in humans, is more prevalent in non-equatorial populations and exhibits strong signatures of recent positive selection. Its associated gene, ADCY3, plays a critical role in thermogenic regulation in mice, further supporting its relevance to cold-adaptive energy balance. This pattern is consistent with prior studies showing that metabolic variants often bear signatures of selection linked to lifestyle and energy balance throughout human history.26^,^27^,^28 This suggests that the variant may have conferred an adaptive metabolic advantage in response to specific environmental pressures, such as colder climates. The A allele may have conferred an adaptive metabolic advantage in response to colder climates, consistent with the hypothesis that seasonal or climate-related thermal stress can shape long-term thermogenic programming. Supporting this idea, recent human research by Yoneshiro et al. showed that individuals conceived during colder seasons exhibited higher BAT activity and greater thermogenic capacity later in life.29 Taken together, the population genetics of rs11676272 and its functional role in BMAL1-dependent thermoregulation strongly suggest that ADCY3 is a gene that has been shaped across human history to fine-tune energy expenditure in response to environmental demands, providing a modern example of adaptive metabolic genetics.
Our findings consistently point to the conclusion that high levels of functional ADCY3 activity are a key feature of a healthy metabolic state in adipose tissue. This principle is supported by two independent lines of evidence. First, our genetic data reveals that the protective A allele of rs11676272, which is predicted to produce a more stable ADCY3 protein, is robustly associated with lower BMI and fat mass. Second, and in perfect alignment with this, independent clinical data show that successful diet-induced weight loss triggers a significant physiological upregulation of ADCY3 expression in human adipose tissue.18^,^19 This convergence of genetic and clinical evidence solidifies the importance of the peripheral adipose pathway and establishes ADCY3 as a key marker and potential mediator of metabolic health in humans.
In conclusion, we identify ADCY3 as a key integrator of circadian and metabolic signals. We show that a common variant interacts with difficulty awakening to increase obesity risk, linking behavioral circadian strain to adipose regulation through both molecular and physiological mechanisms. These findings establish a new framework for understanding gene-environment interactions in metabolic disease and highlight ADCY3 as a tractable target for rhythm-based interventions aimed at improving metabolic resilience and reducing obesity risk in genetically susceptible individuals. To our knowledge, this is the first coding variant shown to connect a circadian behavioral trait with adipose regulation. This novelty underscores the significance of our findings for understanding how genetics, circadian behavior, and adipose biology intersect to shape metabolic disease risk.
Limitations of the study
While our data strongly support a peripheral adipose-centric mechanism, we acknowledge the well-established role of ADCY3 in the central nervous system. ADCY3 is highly expressed in the brain, and rare, severe loss-of-function mutations were associated with monogenic obesity, highlighting its critical function in central energy homeostasis pathways.20 Thus, it is plausible that the pleiotropic effects of this common variant reflect a combined influence in both central and peripheral tissues. In the brain, ADCY3 localizes to primary cilia of hypothalamic nuclei, including the arcuate nucleus (ARC), suprachiasmatic nucleus (SCN), and paraventricular nucleus (PVN), which are involved in metabolic and circadian regulation.30^,^31^,^32 The SCN, recognized as the master circadian clock, coordinates daily rhythms in physiology and behavior.33^,^34^,^35^,^36 Notably, glutamate stimulation of the SCN increases BAT thermogenesis and core body temperature in rats, demonstrating that central circadian circuits can modulate peripheral energy expenditure.37 Furthermore, the melanocortin-4 receptor (MC4R), a Gs-coupled GPCR involved in appetite suppression, co-localizes with ADCY3 at PVN neuron cilia, and disruption of either protein’s ciliary localization impairs energy homeostasis.31 Selective inhibition of ADCY3 signaling at MC4R neuronal cilia increases food intake and body weight,31 underscoring the importance of ADCY3-mediated cAMP signaling in central metabolic control. Future studies will be necessary to elucidate the relative contributions of central versus adipose ADCY3 signaling to pleiotropic trait associations. Another limitation of our work is that the difficulty waking in the morning is based on a single self-report question; objective measures such as polysomnography or actigraphy would offer more granular insight into circadian arousal phenotypes. Additionally, the African and south Asian subsets were underpowered for interaction analyses; therefore, future studies with larger multi-ancestry samples will be necessary to evaluate population-specific effects. Notably, our findings for ADCY3 do not exist in isolation. Other pleiotropic loci identified in our genome-wide pleiotropy screen, such as CELF1, are also known to regulate thermogenic output, albeit through different post-transcriptional mechanisms.38^,^39 This convergence suggests a broader principle in which multiple circadian-metabolic loci modulate energy expenditure through complementary molecular pathways.
Resource availability
Lead contact
Further information and requests for resources should be directed to and will be fulfilled by the lead contact, Richa Saxena ([email protected]).
Materials availability
This study did not generate new, unique materials.
Data and code availability
- •This study analyzes existing, publicly available data. GWAS summary statistics were obtained from the UK Biobank and published consortia, including the GIANT Consortium and the HUGE-AMP Knowledge Portals, as cited in the manuscript. Mouse circadian RNA-seq data were obtained from GEO (GEO: [GSE54651](GSE54651)), and BMAL1 ChIP-seq data were obtained from GEO (GEO: [GSE181443](GSE181443)). Human adipose RNA-seq data were obtained from the Adipose Tissue Knowledge Portal. GTEx v8 expression quantitative trait loci data were obtained from the GTEx Portal. Human histone ChIP-seq data were obtained from ENCODE, and transcription factor ChIP-seq annotations were obtained from RegulomeDB. Individual-level genotype and phenotype data used for interaction analyses were accessed through the UK Biobank Resource (application number 6818) and are available to qualified researchers upon approved application. All accession numbers and resource links are listed in the key resources table.
- •This study does not report original code. All analyses were performed using publicly available software and R packages as specified in the STAR Methods and key resources table.
- •Any additional information required to reanalyze the data reported in this study is available from the lead contact upon request.
Acknowledgments
This research has been conducted using the UK Biobank Resource (UK Biobank application number 6818). We would like to thank the participants and researchers from the UK Biobank study and Genetic Investigation of ANthropometric Traits (GIANT) Consortium. C.T. is supported by BWF G-1022367 and 10.13039/100009886AASM 358-FP-24. J.M. is supported by a Humboldt Professorship of the 10.13039/100005156Alexander von Humboldt Foundation. J.M. acknowledges funding by the 10.13039/501100001659Deutsche Forschungsgemeinschaft (DFG) through SFB1423 (421152132), SFB 1664 (514901783), TRR 386 (514664767), and SPP 2363 (460865652). J.M. is supported by the 10.13039/501100002347Federal Ministry of Education and Research (BMBF) through the Center for Scalable Data Analytics and Artificial Intelligence (ScaDS.AI), through the 10.13039/501100018929German Network for Bioinformatics Infrastructure (de.NBI), and through the 10.13039/100021828German Academic Exchange Service (DAAD) via the School of Embedded Composite AI (SECAI 15766814). Work in the Meiler laboratory is further supported through the 10.13039/100000002National Institutes of Health (NIH) through R01 HL122010, R01 DA046138, R01 AG068623, R01 LM013434, S10 OD016216, S10 OD020154, and S10 OD032234; R.S. is supported by 10.13039/100000002NIH R01 DK107859, 10.13039/100000002NIH R01 DK102696, 10.13039/100000002NIH R01 HL146751, and the 10.13039/100005294MGH Research Scholar Fund. J.M.L. is supported by 10.13039/100000002NIH/10.13039/100000050NHLBI K01 HL136884 and 10.13039/100000002NIH/10.13039/100000051NHGRI R01 HG012810.
Author contributions
C.T. and R.S. designed the study; J.L. conducted chronotype GWAS for the UKB cohort; C.T. acquired the BMI GWAS summary statistics. C.M. and J.M. built the protein structural biology pipeline, and C.T. conducted the remaining human genetics and in vivo periodicity analyses. All co-authors participated in acquiring, analyzing, and interpreting the data. C.T. wrote the manuscript, and all co-authors reviewed and edited the manuscript before approving its submission. R.S. is the guarantor of the work and, as such, has full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Declaration of interests
The authors have declared no competing interests.
Declaration of generative AI and AI-assisted technologies in the writing process
During the preparation of this work, the authors used Harvard GPT (Educational license) to assist with manuscript organization, scientific writing, and figure legend refinement. In addition, the Stanford Biomni AI agent was used to generate the BMAL1 ChIP-seq visualization presented in Figures 6C and S6. After using these tools, the authors reviewed, edited, and validated all content manually and take full responsibility for the integrity and accuracy of the manuscript.
STAR★Methods
Key resources table
REAGENT or RESOURCESOURCEIDENTIFIERDeposited dataUK Biobank genotype and phenotype dataUK BiobankApplication #6818UK Biobank chronotype GWAS summary statisticsUK Biobank | HUGE-AMP Sleep Portalhttps://sleep.hugeamp.orgUK Biobank BMI GWAS summary statisticsUK Biobank | HUGE-AMP Metabolic Portalhttps://hugeamp.orgGIANT Consortium BMI GWAS summary statisticsGIANT Consortiumhttps://portals.broadinstitute.org/collaboration/giantGTEx v8 eQTL dataGTEx Consortiumhttps://gtexportal.orgMouse circadian RNA-seq (white adipose tissue)Zhang et al.40GEO: [GSE54651](GSE54651)Mouse circadian RNA-seq (inguinal white adipose tissue)Xiong et al.41GEO: [GSE183000](GSE183000)Mouse circadian RNA-seq (liver and adipose)Paschos et al.11GEO: [GSE35026](GSE35026)Mouse BMAL1 ChIP-seq (inguinal white adipose tissue)Hepler et al.42GEO: [GSE181443](GSE181443)Human adipose RNA-seq (weight-loss studies)Adipose Tissue Knowledge Portalhttps://adiposetissue.orgHuman histone ChIP-seq dataENCODE Consortiumhttps://www.encodeproject.orgTranscription factor binding annotationsRegulomeDBhttps://regulomedb.orgSoftware and algorithmsPLACORay and Chatterjee12https://github.com/RayDebashree/PLACOHyPrColocFoley et al.13https://github.com/jrs95/hyprcoloclme4Bates et al.15https://github.com/lme4/lme4MetaCycleWu et al.43https://github.com/gangwug/MetaCycleAlphaFold2DeepMindhttps://alphafold.ebi.ac.ukPyMOL v2.5.5Schrödinger LLChttps://pymol.orgOtherProtein structure visualizationVUStruct Pipelinehttps://staging.meilerlab.org/vu-structBMAL1 ChIP-seq visualizationStanford Biomnihttps://github.com/snap-stanford/biomni
Experimental model and subject details
Human samples
This study analyzed existing, de-identified genome-wide association study (GWAS) summary statistics from the UK Biobank (UKB), a population-based cohort of approximately 500,000 participants aged 40–69 years at recruitment. Chronotype summary statistics were derived from a self-reported morningness phenotype (N = 449,734).4 BMI summary statistics were obtained from a meta-analysis combining UK Biobank and GIANT Consortium data (total N = 694,649).14 A GWAS for difficulty waking up in the morning was conducted de novo in UK Biobank participants (N = 451,872) under UK Biobank application number 6818, with adjustment for age, sex, genotyping array, and the first 10 genetic principal components, and the de-identified genome-wide association study (GWAS) summary statistics have been publicly available in the Sleep Knowledge portal.44
UK Biobank participants include individuals of multiple ancestries; ancestry-specific analyses were conducted for European, African, and South Asian ancestry groups as described in the results. Sex-stratified analyses were performed to assess sex-dependent effects, and a significant gene-by-sex interaction was observed for adiposity traits. No new human participants were recruited for this study. All analyses were performed on publicly available, summary-level data, and no individual-level identifiable information was accessed. Ethical approval and informed consent procedures are described in the original UK Biobank study.
Animal models
All animal data analyzed in this study were obtained from previously published, publicly available datasets. Circadian gene expression analyses were performed using RNA-seq data from adult C57BL/6 male mice (6 weeks of age) entrained to a 12 h:12 h light–dark cycle for one week and subsequently released into constant darkness, with tissues harvested every 6 h across circadian times CT22–CT64 (GEO: [GSE54651](GSE54651)). These data were used to assess endogenous circadian rhythmicity across multiple tissues, including white and brown adipose tissue.
Additional RNA-seq data from inguinal white adipose tissue (iWAT) were obtained from adult male C57BL/6 mice (10–12 weeks of age) maintained under a 12 h:12 h light–dark cycle and exposed to either ambient (22°C) or cold (10°C) conditions, including wild-type and Bmal1-deficient genotypes (GEO: [GSE183000](GSE183000)). Tissue samples were collected at zeitgeber time ZT3 following a two-week acclimation period. Direct transcriptional regulation was assessed using BMAL1 ChIP-seq data from mouse iWAT (GEO: [GSE181443](GSE181443)).
All animal experiments were conducted by the original study authors in accordance with institutional animal care and use guidelines, as described in the respective source publications. No new animal experiments were performed for this study.
Method details
Genome-wide pleiotropy analysis
To identify shared genetic signals between circadian preference and BMI, we used PLACO (Pleiotropic Analysis under Composite Null Hypothesis v0.1.1), a statistical approach that detects pleiotropic loci based on summary-level GWAS data from two traits.12^,^45 We harmonized effect alleles, removed SNPs with mismatched alleles, and filtered datasets for minor allele frequency (MAF) > 1%. The Pearson correlation of Chrono-BMI and Getup-BMI Z-scores were 0.0758 and 0.0871, respectively. A total of 67,449 SNPs were available for Chrono-BMI pleiotropy analysis and 41,808 SNPs for Getup-BMI pleiotropy. Given the large effect sizes in BMI GWAS, applying the recommended Z^2^ > 80 filtering would have disproportionately removed variants with strong pleiotropic effects. Since the GWAS summary statistics have overlapping samples/individuals from the UK-Biobank, we decorrelate the Z-scores using the correlation matrix. To ensure the retention of biologically meaningful SNPs, we validated our PLACO findings through HyPrColoc (Hypothesis Prioritization in Multi-Trait Colocalization v1.0),13 a robust method prioritizing shared causal variants across multiple traits. To visualize pleiotropic loci, we generated Manhattan and Q-Q plots using the qqman R package46 All the regional association plots were generated using LocusZoom47 from the GWAS summary statistics for circadian preference and BMI.
Colocalization analysis
To validate the PLACO results, we performed colocalization using HyPrColoc (Hypothesis Prioritization in Multi-Trait Colocalization v1.0) default parameters of prior.2 = 0.98, bb.alg = F.
Functional annotation and causal inference
We annotated significant variants using FUMA (v1.5.2).48 Lead SNPs were identified using a P-value threshold of 5 × 10^−8^, and linkage disequilibrium (LD) was assessed using the UKB British 10K reference panel (LD R^2^ threshold: lead SNPs = 0.6, secondary SNPs = 0.1, positional mapping window = 10 kb, locus merge distance = 250 kb).
To investigate regulatory functions, we integrated GTEx v8 cis-eQTLs49 from VannoPortal50 and Roadmap Epigenomics data51 to identify tissue-specific regulatory effects. The human subcutaneous adipose tissue ChIP-Seq for enhancer epigenetic markers H3K4me1 (ENCFF896XMZ, ENCFF806FHF, and ENCFF562LBI) and promoter marker H3K4me3 (ENCFF017XYK, ENCFF078XHS, and ENCFF494FAI) were obtained from ENCODE, and transcription factor binding data from RegulomeDB.52
Tissue-specific colocalization and causal inference
To assess whether pleiotropic signals reflect shared genetic regulation, we applied HyPrColoc to evaluate colocalization between PLACO-identified loci and tissue-specific eQTLs. We prioritized loci within ADCY3, CELF1, FTO, and TFAP2B based on their pleiotropic effects on circadian traits and BMI. HyPrColoc estimates posterior probability (PP) for colocalization, identifying candidate causal SNPs contributing to the total colocalization signal.
To assess the causality and directionality of ADCY3 expression on complex traits, we performed Mendelian Randomization (MR) analysis focused on the missense variant rs11676272 using the TwoSampleMR R package (v0.6.14).53^,^54 The eQTL effect of rs11676272 on ADCY3 expression in subcutaneous adipose tissue (SAT) was used as the exposure, and GWAS summary statistics for BMI, morningness chronotype (Chrono), and difficulty waking up (Getup) were used as outcomes. Causal estimates were derived using the Wald ratio method, which is optimal when a single variant is used as an instrumental variable. The MR Z score was computed as MR Beta/MR SE.
Phenome-wide association study
To explore additional associations, we used OpenTargets55 to conduct a phenome-wide association study (PheWAS) using GWAS summary statistics from FinnGen, UK Biobank, and the GWAS Catalog.
UK Biobank genetic and phenotypic model analysis
To dissect the effects of the lead variant rs11676272, we performed detailed analyses in the UK Biobank cohort. All linear regression models testing the main effect of genotype, sex-dimorphism, and gene-by-behavior interactions were adjusted for age, sex, and the first 10 genetic principal components to control for population stratification. Analyses were performed in R (v4.1.0). Full model summaries are available in Table S8.
Protein structural modeling
We performed calculations and visualization on the ADCY3 full-length AlphaFold 256 model of UniProt: O60266-1, as well as 37% identity Swiss homology Models templated on CryoEM structures PDB: 8sl4.pdb (ADCY5 positions 51 to 1124) and PDB: 8buz.pdb (ADCY8 positions 59 to 1114 positions). We used the ΔΔG Cartesian57^,^58 protocol as implemented in the VUStruct pipeline59 to predict whether S107P might negatively impact ADCY3’s free energy of folding. The PyMOL Molecular Graphics System, Version 2.5.5, Schrödinger, LLC, was used to visualize ADCY3 protein structure with a focus on changes to the hydrogen bonding network around Serine 107 on the mutation to Proline.
ADCY3 circadian rhythmicity in animal models
To assess endogenous circadian rhythmicity, we analyzed publicly available RNA-seq data from C57Bl/6 male mice ([GSE54651](GSE54651)),40 where 6-week-old mice were entrained to a 12h:12h light:dark schedule for 1 week, then placed in constant darkness, and tissues were harvested at circadian time CT 22–64 every 6 h.40 RNA was extracted using Qiagen RNeasy kit, and Illumina HiSeq 2000 was used for sequencing. DeSeq2 was used for normalization. We used the MetaCycle R package43 to assess the rhythmicity of gene expression from white adipose, brown adipose, hypothalamus, cerebellum, liver, and aorta. For periodicity analysis in MetaCycle, we used Meta2d method that integrates the ARS (Autoregressive Spectral analysis), the JTK_CYCLE (Jonckheere-Terpstra-Kendall Cycle), and the LS (Lomb-Scargle periodogram) to analyze rhythms in time-series data.43 This integrated approach improves detection power by combining complementary algorithms for rhythmicity in time-series data without replicates. GraphPad Prism (v.10.2.3) was used to generate the rhythmic gene expression figures.
Due to ADCY3 adipose-specific rhythmicity, we used openly available RNAseq data from NCBI Geo [GSE183000](GSE183000) for C57Bl/6 mice inguinal white adipose tissue (iWAT).41 The mice were kept in a 12:12 light/dark cycle. For adipose-specific Bmal1 target, Bmal1fl/fl and Prx1-Cre transgenic mice alongside CRISPR-engineered ROSA-26 Bmal1 knock-in and knockout mice. Then, the 10-12 week-old male mice (n = 3/group) were placed in an ambient temperature of 22°C and a cold temperature of 10°C with a 2-week acclimation period, and the iWATs were collected at ZT3. Total RNA was extracted using Trizol, and Illumina HiSeq 2500 was used for sequencing. GraphPad Prism was used for the analysis and to generate the figures. Additionally, the binding of BMAL1 to ADCY3 was further confirmed in mice using iWAT openly accessible ChipSeq data ([GSE181443](GSE181443)),42 and we used Biomni Stanford60 to visualize the BMAL1 peak; signal intensities were calculated as mean values across 500-bp genomic bins spanning the ADCY3 locus (chr12:4,133,103-4,313,525, GRCm39 assembly) with 50-kb flanking regions. We then query the ADCY3 homo sapiens sequence from the NCBI Gene: NM_001377130.1 (ADCY3/NM_001377130.1/NP_001364059.1) to scan for the canonical Bmal1-specific E-box motif CACGTG,61 and we identified multiple E-box motifs in human.
Human ADCY3 tissue-specific isoform usage
We used GTex v.8 transcript browser to assess tissue-specific isoform expression of ADCY3 across different tissues, as the splicing regulation and isoform length might explain tissue-specific rhythmicity, as it might result in the removal of BMAL1 binding motifs.
Quantification and statistical analysis
Genome-wide pleiotropy analysis
Genome-wide pleiotropy analysis was conducted using PLACO (Pleiotropic Analysis under Composite Null Hypothesis v0.1.1), a statistical framework that identifies shared genetic effects across two traits. PLACO was applied to identify SNPs influencing morning chronotype, difficulty waking up, and BMI, with statistical significance determined using a genome-wide significance threshold of p < 5 × 10^−8^. SNP harmonization was performed by aligning effect alleles across summary statistics, and variants with minor allele frequency (MAF) < 1% were excluded. Pearson correlation of Z-scores was calculated to assess the degree of pleiotropy between Chrono-BMI and Getup-BMI, yielding correlation coefficients of 0.0758 and 0.0871, respectively. To validate our pleiotropy findings, we performed a secondary pleiotropy analysis using HyPrColoc, a Bayesian method that prioritizes shared causal variants across multiple traits. HyPrColoc colocalization results were considered significant when the posterior probability (PP) exceeded 0.60, indicating strong evidence of a shared causal genetic signal.
Colocalization analysis
Colocalization analysis was performed to determine whether pleiotropic loci exhibit a shared causal effect on gene expression and complex traits. We used HyPrColoc (Hypothesis Prioritization in Multi-Trait Colocalization v1.0) to estimate posterior probabilities (PP) of colocalization between PLACO-identified SNPs and expression quantitative trait loci (eQTLs) in functionally relevant tissues obtained from GTEx v8. Colocalization was performed using a ± 1 kb window around each gene of interest based on the GRCh37/hg19 genome build. The genomic regions analyzed were: ADCY3 (chr2:25,041,038–25,144,106), CELF1 (chr11:47,486,489–47,588,091), FTO (chr16:53,736,875–54,156,853), and TFAP2B (chr6:50,785,584–50,816,332). A colocalization signal was considered noteworthy when PP > 0.60, suggesting that the same genetic variant regulates gene expression and the associated trait.
Linear mixed-model analyses (UK Biobank)
All genotype–phenotype associations were tested with linear mixed models in R (lme4 v1.1-34; lmerTest v3.1-3). For each quantitative trait (BMI, fat mass, % body fat, chronotype score, and morning difficulty waking score), we fitted.
where Genotype were coded as a three-level factor (reference = GG; comparison levels AG and AA). Kinship_ID is a random intercept that groups all individuals belonging to the same UK Biobank kinship component, thereby accounting for cryptic relatedness. Sex-stratified models omitted the Sex term; interaction models added the relevant cross-product (e.g., Genotype × Sex or Genotype × GET_UP). Complete model outputs are provided in Table S8.
Mendelian Randomization
Mendelian randomization (MR) was performed to determine whether ADCY3 expression causally influences morning chronotype and BMI. The TwoSampleMR R package (v0.6.14)53^,^54 was used to integrate eQTL and GWAS data. Given that rs11676272 was the only available genetic instrument for ADCY3 expression in adipose tissue, we applied the Wald ratio method, which estimates causal effects by dividing the variant’s effect size on the outcome by its effect on the exposure.
The eQTL effect of rs11676272 on ADCY3 expression in subcutaneous adipose tissue (SAT) was used as the exposure, and GWAS summary statistics for BMI, morningness chronotype (Chrono), and difficulty waking up (Getup) were used as outcomes. Causal estimates were derived using the Wald ratio method, which is optimal when a single variant is used as an instrumental variable. The MR Z score was computed as MR Beta/MR SE and significance was assessed at p < 0.05.
Protein structural modeling and stability analysis
Protein structural analysis was conducted to evaluate the impact of rs11676272 (S107P) on ADCY3 stability. Structural modeling was performed using AlphaFold2 and Swiss homology models, comparing ADCY3 against CryoEM templates of ADCY5 and ADCY8. Structural stability was assessed using Rosetta ddG Cartesian calculations, where ΔΔG values greater than 3 kcal/mol were considered indicative of significant destabilization. The PyMOL Molecular Graphics System (v2.5.5) was used for molecular visualization, specifically examining hydrogen bonding networks, polar interactions, and structural dynamics following the S107P substitution.
Circadian expression analysis
Circadian expression analysis was conducted to assess the rhythmic regulation of ADCY3 in adipose tissue. RNA-seq datasets from mice (C57Bl6, [GSE54651](GSE54651), [GSE183000](GSE183000)) were analyzed using MetaCycle, a statistical tool that integrates ARS (Autoregressive Spectral Analysis), JTK_CYCLE (Jonckheere-Terpstra-Kendall Cycle), and LS (Lomb-Scargle Periodogram) methods to detect oscillatory gene expression patterns. BMAL1-dependent transcriptional regulation was further evaluated using ChIP-Seq data, which revealed BMAL1 binding peaks at the ADCY3 promoter in mouse inguinal white adipose tissue (iWAT).
Tissue-specific isoform regulation and BMAL1 binding motif analysis
Tissue-specific isoform expression of ADCY3 was analyzed using GTEx v8 isoform data. Differential isoform expression was assessed across multiple tissues, with adipose-specific transcripts absent in the liver, whole blood, hypothalamus, pancreas, and heart. To determine whether alternative splicing affects BMAL1 binding and ADCY3 rhythmicity, we examined the presence of BMAL1-specific E-box motifs (CACGTG) within the ADCY3 promoter region. Multiple BMAL1-specific E-box binding sites were identified suggesting that BMAL1 directly regulates ADCY3 in a tissue-specific manner.
Statistical software and significance thresholds
All statistical analyses were performed using R (v4.1.0), Python (v3.9), ggplot2 (v3.5.0), Metacycle (1.2.0) and GraphPad Prism (v10.2.3). The significance threshold for all tests was set at p < 0.05 unless otherwise stated. Full statistical details, including sample sizes, effect sizes, confidence intervals, and P-values, are provided in the figure legends and supplementary tables.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lane J.M.Qian J.Mignot E.Redline S.Scheer F.A.J.L.Saxena R.Genetics of circadian rhythms and sleep in human health and disease Nat. Rev. Genet.24202342010.1038/s 41576-022-00519-z 36028773 PMC 10947799 · doi ↗ · pubmed ↗
- 2Mure L.S.Le H.D.Benegiamo G.Chang M.W.Rios L.Jillani N.Ngotho M.Kariuki T.Dkhissi-Benyahya O.Cooper H.M.Panda S.Diurnal transcriptome atlas of a primate across major neural and peripheral tissues Science 3592018 eaao 031810.1126/SCIENCE.AAO 0318 PMC 592473229439024 · doi ↗ · pubmed ↗
- 3Lane J.M.Vlasac I.Anderson S.G.Kyle S.D.Dixon W.G.Bechtold D.A.Gill S.Little M.A.Luik A.Loudon A.Genome-wide association analysis identifies novel loci for chronotype in 100,420 individuals from the UK Biobank Nat. Commun.720161088910.1038/ncomms 10889 PMC 478686926955885 · doi ↗ · pubmed ↗
- 4Jones S.E.Lane J.M.Wood A.R.van Hees V.T.Tyrrell J.Beaumont R.N.Jeffries A.R.Dashti H.S.Hillsdon M.Ruth K.S.Genome-wide association analyses of chronotype in 697,828 individuals provides insights into circadian rhythms Nat. Commun.10201934310.1038/s 41467-018-08259-730696823 PMC 6351539 · doi ↗ · pubmed ↗
- 5Hu Y.Shmygelska A.Tran D.Eriksson N.Tung J.Y.Hinds D.A.GWAS of 89,283 individuals identifies genetic variants associated with self-reporting of being a morning person Nat. Commun.720161044810.1038/ncomms 10448 PMC 474081726835600 · doi ↗ · pubmed ↗
- 6Tchio C.Williams J.S.Taylor H.Ollila H.M.Saxena R.An integrative approach prioritizes the orphan GPR 61 genomic region in tissue-specific regulation of chronotype Sleep Adv.62025 zpaf 03010.1093/sleepadvances/zpaf 030PMC 1222426140612388 · doi ↗ · pubmed ↗
- 7Kalmbach D.A.Schneider L.D.Cheung J.Bertrand S.J.Kariharan T.Pack A.I.Gehrman P.R.Genetic basis of chronotype in humans: Insights from three landmark GWAS Sleep 402017 zsw 04810.1093/sleep/zsw 048PMC 608475928364486 · doi ↗ · pubmed ↗
- 8Burns A.C.Zellers S.Windred D.P.Daghlas I.Sinnott-Armstrong N.Rutter M.Hublin C.Friligkou E.Polimanti R.Phillips A.J.K.Sleep inertia drives the association of evening chronotype with psychiatric disorders: epidemiological and genetic evidence Preprint atmed Rxiv 202410.1101/2024.09.10.24313197 · doi ↗