Experimental susceptibility of Macrobrachium rosenbergii (de Man) subadults and adults to infectious myonecrosis virus (IMNV)
Scarlatt Paloma Alves da Silva, Fernando Leandro dos Santos, Suzianny Maria Bezerra Cabral da Silva

TL;DR
This study shows that Macrobrachium rosenbergii can be infected with a virus that affects shrimp, which could impact freshwater prawn farming.
Contribution
This is the first study to demonstrate that Macrobrachium rosenbergii is susceptible to infectious myonecrosis virus.
Findings
Macrobrachium rosenbergii subadults and adults showed positive PCR results and typical clinical signs of IMNV infection.
Histopathological changes like hemocyte infiltration and muscle sinus dilation were observed in infected prawns.
No massive mortalities occurred, but the prawns can act as a viral source in farming environments.
Abstract
Infectious myonecrosis virus (IMNV) is considered one of the pathogens that most affect the production of Penaeus vannamei, causing economic losses estimated at billions of dollars. P. vannamei rearing is expanding to inland waters, which are commonly used for the farming of Macrobrachium rosenbergii, increasing the probability of exposure of this freshwater prawn to viruses often reported in penaeids, including IMNV. Therefore, the aim of the present study was to investigate the susceptibility of M. rosenbergii to IMNV through an intramuscular challenge. M. rosenbergii subadults and adults were inoculated intramuscularly with IMNV obtained from the abdominal muscle tissue of naturally infected P. vannamei. The viral challenge lasted 21 days. All samples had positive nested-PCR results as well as clinical signs and histopathological findings typical of the virus, such as hemocyte…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
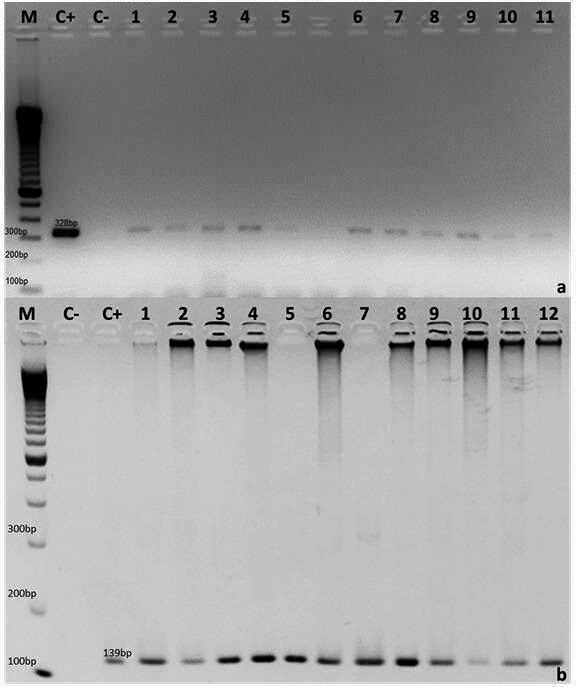 Figure 1
Figure 1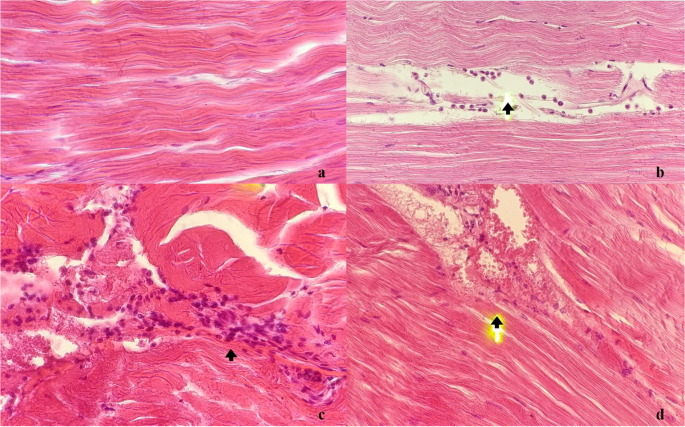 Figure 2
Figure 2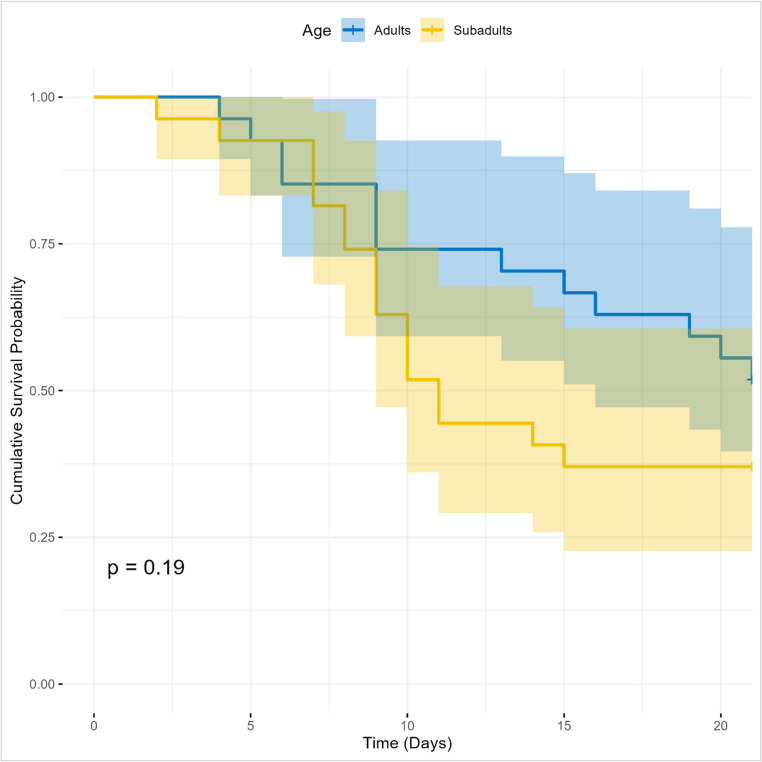 Figure 3
Figure 3- —Universidade Federal Rural De Pernambuco
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Crustacean biology and ecology · Marine Bivalve and Aquaculture Studies
Introduction
The production of farmed crustaceans reached a total of 11.2 million tons in 2020, corresponding in monetary terms to US$81.5 billion and approximately 13% of all aquiculture production for human consumption (FAO, 2022). Among species produced in shrimp farming activities, the giant freshwater prawn, Macrobrachium rosenbergii, is the fifth most widely farmed crustation in the world, with global production of 294 thousand tons in 2020 (FAO, 2021), to which Brazil contributed an estimated 150 thousand tons (Valenti et al. 2021).
Although M. rosenbergii is an important source of aquicultural income, economic losses, especially those caused by infectious diseases, result in fluctuations in production (Bonami and Sri Widada 2011; Chen et al. 2021). There are frequent reports of the incidence of viruses, particularly white tail disease (WTD) or white muscle disease (WMD), which is the greatest threat to the viability of M. rosenbergii production due to the combination of M. rosenbergii nodavirus (MrNV) and extra small virus (XSV) (Hooper et al. 2022). The main clinical signs of WTD are the whitish coloration of the muscles, which begins in the abdomen and affects the entire musculature of the animal in more advanced stages, as well as lethargy and anorexia (Bonami and Sri Widada 2011).
Besides M. rosenbergii, infection by MrNV has been reported in Penaeus vannamei farmed in Asia (Senapin et al. 2012). In the study cited, after the screening of samples negative for infectious myonecrosis virus (IMNV) and Penaeus vannamei nodavirus (PvNV) via real-time polymerase chain reaction (RT-PCR), a reassessment was performed of this material from grow-out farms with high mortality rates and reports of animals with muscle opacity as well as positive RT-PCR results for MrNV; moreover, infectiousness was demonstrated when P. vannamei was fed chopped tissue from M. rosenbergii infected by MrNV at low temperatures and salinity of the cultivation water (Senapin et al. 2012). In another study, Senapin et al. (2013) reported coinfection by IMNV and MrNV in P. vannamei subadults farmed in Indonesia detected by nested RT-PCR and immunohistochemical analysis, with the occurrence of muscle lesions in the coinfected animals.
P. vannamei is the only marine shrimp farmed in Brazil (IBGE, 2021), and its farming in brackish and freshwater environments as it expands inland (Pimentel et al. 2022), increases the probability of approaching areas of M. rosenbergii production and exposing this species to other viruses, including IMNV, which is considered endemic in northeast Brazil (Prasad et al. 2017). IMN results in muscular opacity of the abdominal segments due to tissue necrosis, and is responsible for 40% to 80% of mortality in shrimp farming, causing estimated US$ 440 million in losses in Brazil between 2002 and 2005 (Andrade et al. 2007). Therefore, the aim of the present study was to investigate the susceptibility of M. rosenbergii to IMNV through a viral challenge.
Materials and methods
Preparation of viral inoculum
The viral inoculum used came from a strain endemic to Northeast Brazil, obtained from a naturally infected animal during an outbreak on a commercial farm. The presence of IMNV in the tissue used for the viral inoculum (100 g) was confirmed by nested PCR (Poulos and Lightner 2006) and the absence of infection by Taura Syndrome Virus (TSV), White Spot Syndrome Virus (WSSV), Infectious Hypodermal and Hematopoietic Necrosis Virus (IHHNV), and Necrotizing Hepatopancreatitis Bacterium (NHPB) was confirmed by nested PCR, via IQ2000TM kits, according to the manufacturer’s intructions (Farming IntelliGene Tech. Corp., Taiwan) (data not shown). The protocol for preparing the IMNV viral inoculum was carried out according to Silva et al. (2015) and, for this purpose, the tissue was then homogenized in 300 mL of 2% sterile saline solution (1:3, v: v). The homogenized material was successively filtered (300, 210 and 70 μm) to obtain the inoculum, which was diluted at 1:10. The viral load was determined using the method described by Silva et al. (2011) (1.1 × 10³ copies of IMNV/µg of RNA), followed by storage at − 80 °C until use in the viral. challenge.
Experimental conditions
Fifty-four healthy individuals of M. rosenbergii were used in this experiment, including subadults (5–15 g) and adults (16–26 g) (Valenti 1998). The adults were randomly distributed, with no separation between males and females for the experiment. All specimens were from the Johei Koike Mainland Aquiculture Station of Universidade Federal Rural de Pernambuco (UFRPE). The prawns were distributed according to weight and kept in experimental units with 15 L of useful volume at a density of one animal per 5 L at 28 °C in freshwater. The animals were fed a commercial ration (35% of crude protein; Guabitech Active 35, Brazil) corresponding to 3% of the biomass twice per day, as described by D’Abramo and Sheen (1994).
The water used in the system was previously filtered (30 μm) and treated with chlorine (30 ppm). Alkalinity and hardness were maintained above 50 mg of CaCO_3_/L by the addition of sodium carbonate. The volume of the experimental units was renewed daily at a rate of 20% of the total volume. Daily cleaning of the experimental units was performed through siphoning, with the replacement of the volume of water lost during the procedure.
Temperature, pH, and oxygen were monitored twice per day. Water samples were collected weekly for the analysis of nitrite, nitrate, ammoniacal nitrogen, alkaline and hardness using a commercial colorimetric kit (Alcon Labcon, Camboriú, Brazil) to ensure that these parameters were within the ideal limits for carids proposed by New et al. (2010).
All experimental conditions were conducted in accordance with ethical and experimental precepts, authorized by the Ethics Committee on the Use of Animals (CEUA) of the Universidade Federal Rural de Pernambuco (UFRPE).
Viral challenge
The susceptibility of M. rosenbergii subadults and adults to IMNV was determined via an intramuscular injection, following the method described by Silva et al. (2015). The treatments were (1) M. rosenbergii subadults experimentally challenged with IMNV, (2) M. rosenbergii adults experimentally challenged with IMNV, (3) unchallenged M. rosenbergii subadults (control group 1), and (4) unchallenged M. rosenbergii adults (control group 2).
Treatments 1 and 2 comprised six repetitions with three animals each, whereas treatments 3 and 4 (control groups 1 and 2) comprised three repetitions with three animals each. All animals in treatments 1 and 2 were challenged with 100 µL of viral inoculum (dilution: 1:10) using an insulin syringe and an intramuscular injection of the third abdominal segment, as described by Silva et al. (2015). In the negative control group, the animals were injected with 100 µL of 0.85% sterile saline solution.
Throughout the challenge, the animals were monitored daily for the observation of clinical signs of IMNV and mortality. The experiment lasted 21 days, during which time dead animals were collected and stored at − 80 °C for subsequent nested-PCR analysis. Survivors were sacrificed on Day 21 and also stored at − 80 °C for subsequent molecular analysis.
Molecular analysis
RNA extraction and RT-PCR
Total RNA was extracted from the muscle (60 mg) of the second abdominal segment in 1 mL of TRIzol (Invitrogen, USA), following the manufacturer’s instructions. After extraction, RNA was qualitatively and quantitatively assessed via spectrophotometry at 260 and 280 ηm (NanoVue Plus™ spectrophotometer, GE Healthcare, USA) and stored at − 80 °C. RT-PCR was then performed using the Improm-II™ Reverse Transcription System (Promega, Madison, WI, USA) in a final volume of 20 µL, with 300 ηg/µL of total RNA and 0.5 µg of oligo(dT)15, following the manufacturer’s instructions. cDNA was stored at − 20 °C until the nested-PCR analysis.
Nested PCR
Infection by IMNV in M. rosenbergii subadults and adults was determined by nested PCR using the specific primers described by Poulos and Lightner (2006). This is the confirmatory diagnosis recommended for IMNV by the World Organization for Animal Health (OIE, 2023). The reaction conditions and thermal cycles were those described by Poulos and Lightner (2006). The products of the first (328 bp) and second (139 bp) PCR were submitted to electrophoresis in 2% agarose gel stained with ethidium bromide, and fragment size was estimated using a 100-bp molecular weight marker (Invitrogen, USA). In each set of nested PCR reactions, a positive control was used, with P. vannamei tissues positive for IMNV, as well as a negative control, replacing the cDNA with ultrapure water (Sigma, USA).
Histopathological analysis
All animals in the different treatments were submitted to histopathological analysis for the identification of lesions suggestive of IMNV. For such, the prawns were fixed in a Davidson’s alcohol-formalin-acetic acid solution for 72 h and transferred to a 70% ethanol solution. The samples were then dehydrated, clarified, embedded in paraffin, cut to a thickness of 3 to 5 μm and stained with hematoxylin and eosin, as described by Lightner (1996).
All slides were examined under an optical microscope for the determination of coagulative necrosis among the muscle fibers of the abdominal segments, hemocyte infiltration among the muscle fibers, and the occurrence of cytoplasmic inclusion bodies (Tang et al. 2005).
Statistical analysis
As normal distribution and equality of variances were confirmed by the Shapiro-Wilk test and Bartlett’s test, respectively, analysis of variance (ANOVA) and the Student’s t-test were used to determine statistically significant differences between treatments (p ≤ 0.05). The data from the nested-PCR analysis and data on mortality were submitted to descriptive statistics. The cumulative survival probability for each group was estimated using the Kaplan-Meier method. Curves were plotted to visualize the mortality dynamics of both groups over time. Confidence intervals of 95% were calculated and plotted to indicate the precision of the estimates. To determine whether the observed difference in survival curves was statistically significant, the log-rank test was used. This test compares the survival distribution between groups, with the null hypothesis (H0) that there is no difference in mortality between adults and subadults. A p-value less than 0.05 was considered statistically significant. All data were analyzed using R software (version 4.5.1) and the survival and survminer packages.
Results and discussion
All samples of M. rosenbergii subadults and adults challenged with IMNV (1.1 × 10³ copies of IMNV/µg of RNA) had positive nested-PCR results (Fig. 1), showing clinical signs of infection such as multifocal opacity in the muscles of the abdominal segment, lethargy, and a reduction in food intake. No massive mortalities occurred during the period of the experiment (Table 1).
Fig. 1. Detection of IMNV in Macrobrachium rosenbergii subadults and adults experimentally infected with intramuscular injection. M: 100-bp molecular weight marker (Invitrogen, USA); C+: positive control; C−: negative control (ultrapure water); (a) 1 to 5: samples of M. rosenbergii subadults positive for IMNV; 6 to 11: samples of M. rosenbergii adults positive for IMNV; (b) 1 to 6: samples of M. rosenbergii subadults positive for IMNV; 7 to 12: samples of M. rosenbergii adults positive for IMNV
Table 1. Cumulative mortality in different treatments of M. rosenbergii challenged with IMNVTreatmentNumber of individualsCumulative mortalityDay 1Day 7Day 14Day 21Day 22^^118028172180243939033034902025^^Sacrificed animals. Treatment: 1- M. rosenbergii subadults experimentally challenged with IMNV; 2- M. rosenbergii adults experimentally challenged with IMNV; 3- Unchallenged M. rosenbergii subadults (control group 1); 4- Unchallenged M. rosenbergii adults (control group 2)
In the first PCR, nine animals were positive in treatment 1 (M. rosenbergii subadults experimentally challenged with IMNV) and 13 animals were positive in treatment 2 (M. rosenbergii adults experimentally challenged with IMNV) (Table 2). All samples were positive in the second PCR (Table 2). All animals in treatments 3 and 4 (control groups 1 and 2) had negative results in both nested-PCR analyses (Table 2) and had no signs of infection.
Table 2. Number of individuals infected in four different treatments of M. rosenbergii challenged with IMNVTreatmentNumber analyzedPositive1st PCRPositive2nd PCR1- M. rosenbergii subadults experimentally challenged with IMNV189182- M. rosenbergii adults experimentally challenged with IMNV1813183- Unchallenged M. rosenbergii subadults (control group 1)9004- Unchallenged M. rosenbergii adults (control group 2)900
IMNV has been detected in other species. Investigating the susceptibility of juveniles of P. vannamei, L. stylirostris, and Penaeus monodon for four weeks through an intramuscular challenge of an inoculum containing purified virions, Tang et al. (2005) demonstrated infection by IMNV on the 14th day post-inoculation (dpi) via in situ hybridization. The authors also found clinical signs typical of IMNV (whitish lesions in the muscles of the abdomen and tail) in all challenged individuals of P. vannamei on the sixth dpi, whereas such lesions were observed in L. stylirostris only on the 13th day and no clinical signs were found in P. monodon. In the present study, clinical signs (muscle opacity) were found in M. rosenbergii subadults and adults experimentally infected with IMNV.
Infection by IMNV was also found in wild breeders of P. monodon caught in the northeastern Indian Ocean. Among the six lots analyzed using nested-PCR, only two had infected animals, with a total of six positive samples among the 90 analyzed in these lots, with no detection of the basophilic cytoplasmic inclusion bodies characteristic of IMNV reported for penaeids (Srisala et al. 2021).
In a study involving the challenge of Fenneropenaeus merguiensis and Penaeus esculentus through noninvasive procedures that reproduced natural infection routes, such as transmission through water and predation (ingestion of tissue of P. vannamei contaminated with IMNV), Gudkovs et al. (2015) performed a set of molecular tests (PCR, sequencing and in situ hybridization) and histological analysis and determined that both species are susceptible to IMNV, with the confirmation of infection and the demonstration of the replication of the virus in both species.
Besides the occurrence of IMNV in diverse species, viral coinfection has also been reported. After the collection of 30 ill specimens of P. vannamei (10 ± 2 g) from a farm in the state of Ceará in northeast Brazil, Feijó et al. (2013) found that 12 had positive qPCR results for both IMNV and WSSV, and also described the following histopathological lesions in the animals: (i) hemocyte infiltration and coagulative necrosis (16/30 animals); (ii) hypertrophy of lymphoid and spheroid organs (9/30 animals); and (iii) ectopic spheroids, mainly close to the antennal gland and in the skeletal muscle (6/30 animals).
Assessing the impact of infectious hypodermal and hematopoietic necrosis virus (IHHNV) and IMNV through an oral challenge in wild juveniles of Farfantepenaeus subtilis (2.56 ± 0.44 g), Coelho et al. (2009) found that only 10% of the challenged animals were positive for both viruses (determined by PCR). The authors also reported signs of tissue changes in the specimens infected with IMNV, in which all positive animals had low levels of hemocyte infiltration and mild coagulation.
After the screening of 15 specimens of P. vannamei with signs of whitish muscles and a suspicion of infection by IMNV collected from out-growth nurseries of eight farms in three provinces of Indonesia, Senapin et al. (2013) found that 10 animals were positive for IMNV and six of these animals were also positive for Macrobrachium rosenbergii nodavirus (MrNV). MrNV and extra small virus (XSV) are the etiological agents of white tail disease (WTD) in Macrobrachium rosenbergii, the most common symptoms of which are opacity of the abdominal muscle and degeneration of the telson and uropods (Chen et al. 2021).
The histopathological analysis revealed hemocyte infiltration (Fig. 2b, c and d) and sinuses dilation in the muscles of the animals experimentally inoculated with IMNV in both treatments 1 and 2 (Fig. 2d). In contrast, no tissue changes were found in the control groups (M. rosenbergii subadults and adults inoculated with 0.85% saline solution) (Fig. 2a).
Fig. 2. Photomicrographs of M. rosenbergii tissue examined for lesions suggestive of IMNV (a, b, c and d): (a) Adult from control group with no apparent change in muscle (arrow) in 6th right abdominal segment. HE staining. Magnification: 400×; (b) Infected subadult (treatment 1) with discrete hemocyte infiltration among muscle bundles (arrow) in 6th right abdominal segment. HE staining. Magnification: 400×; (c) Infected adult (treatment 2) with intense hemocyte infiltration among muscle bundles (arrow) and coagulative necrosis in 1st right abdominal segment. HE staining. Magnification: 400×; (d) Infected adult (treatment 2) with dilation in sinus and discrete hemocyte infiltrate (arrow) in 6th right abdominal segment. HE staining. Magnification: 400×
Tang et al. (2005) reported the occurrence of skeletal muscle lesions, including coagulative necrosis, fibrotic inflammation, fibrosis, and cytoplasmic inclusion bodies, when the penaeid species P. vannamei, L. stylirostris, and Penaeus monodon were experimentally challenged with IMNV. These histopathological findings are compatible with those in M. rosenbergii subadults and adults in the present study (Fig. 2).
Fig. 3. Cumulative survival probability of adults and subadults throughout the experiment
Regarding mortality rates, there is no statistically significant evidence that they differed between the adult and subadult groups (p-value of 0.19) (Fig. 3). Furthermore, although the subadult curve appears to have a higher mortality rate than the adults, this difference may simply be a random variation in the sample and not a true difference in the population.
The combination of data from the nested-PCR and histopathological analyses shows that M. rosenbergii is sensitive to IMNV. This is the first study to demonstrate the susceptibility of M. rosenbergii subadults and adults to IMNV.
Conclusions
The present study demonstrated the susceptibility of M. rosenbergii subadults and adults to infection by IMNV under experimental conditions. Although no massive mortality occurred during the 21 days of the experiment among the challenged prawns, one should not underestimate the presence of M. rosenbergii as a viral source. Future studies are suggested to assess the role of M. rosenbergii as a vector in the horizontal transmission of IMNV in P. vannamei.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1FAO (Food and Agriculture Organization of the United Nations) (2021) Global aquaculture production Quantity (1950–2021). Fisheries and Aquaculture. https://www.fao.org/fishery/statistics-query/en/aquaculture/aquaculture_quantity. Accessed 25 Sept 2022
- 2OIE (World Organization for Animal Health) (2023) Manual of Diagnostic Tests for Aquatic Animals: Infection with infectious myonecrosis virus. Aquatic Manual Online Access. https://www.woah.org/en/what-we-do/standards/codes-and-manuals/aquatic-manual-online-access/. Accessed 14 October 2022
- 3Pimentel OALF, Roy LA, Santos EP, Oliveira VQ, Brito LO (2022) Penaeus vannamei inland farming: perspectives and opinions. Rev Aquac 1–7. 10.1111/raq.12782