Improved oncolytic and immunostimulatory activity of the spontaneous jin-3 reovirus mutant in preclinical bladder cancer models
Arjanneke F. van de Merbel, Maaike H. van der Mark, Lobke C.M. Hensen, Diana J.M. van den Wollenberg, Rob C. Hoeben, Willemijn C.G. Zonneveld, Rob C.M. Pelger, Maxime T.M. Kummeling, Geertje van der Horst, Gabri van der Pluijm

TL;DR
A mutant reovirus called jin-3 shows stronger cancer-killing and immune-boosting effects in bladder cancer models compared to the wild-type virus.
Contribution
The study introduces jin-3, a reovirus mutant with enhanced oncolytic and immunostimulatory activity in preclinical bladder cancer models.
Findings
jin-3 reovirus effectively infects and kills human bladder cancer cells in 2D and 3D models.
jin-3 induces stronger immunogenic cell death and immune mediator production compared to wild-type reovirus.
Co-culture experiments show jin-3 enhances cancer cell death and immune activation in a dose-dependent manner.
Abstract
Immunotherapy has emerged as a promising strategy for treatment of urothelial carcinoma of the bladder (UCB) but not all patients show clinically desirable responses upon treatment with immune-checkpoint inhibition. Oncolytic viruses may unleash the full potential of immunotherapy. Besides tumor-lysing properties, oncolytic viruses can induce durable and systemic anti-viral and anti-tumor immune responses. Here, we evaluated and compared oncolytic and immunostimulatory properties of wild-type reovirus (R124) and the junction-adhesion molecule-independent mutant reovirus (jin-3) in preclinical models of human UCB. Both reovirus variants effectively infected and replicated in human UCB cells and induced cell lysis in flatbottom or 3D cultures and in ex vivo cultured human tumor tissue slices. However, jin-3 reovirus demonstrated greater efficacy in a dose-dependent manner, effectively…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1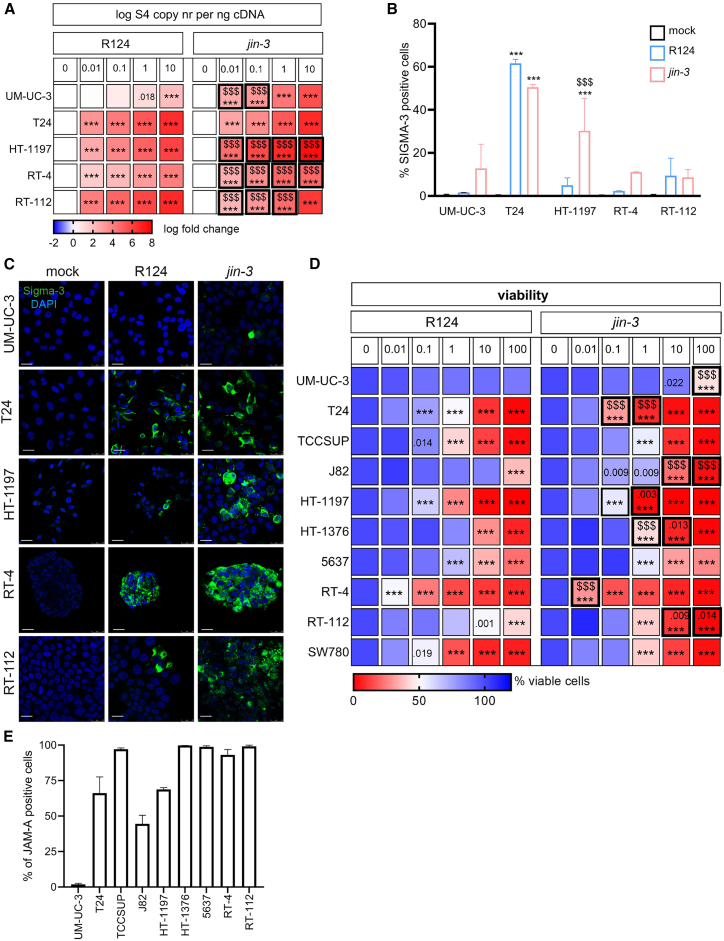 Figure 2
Figure 2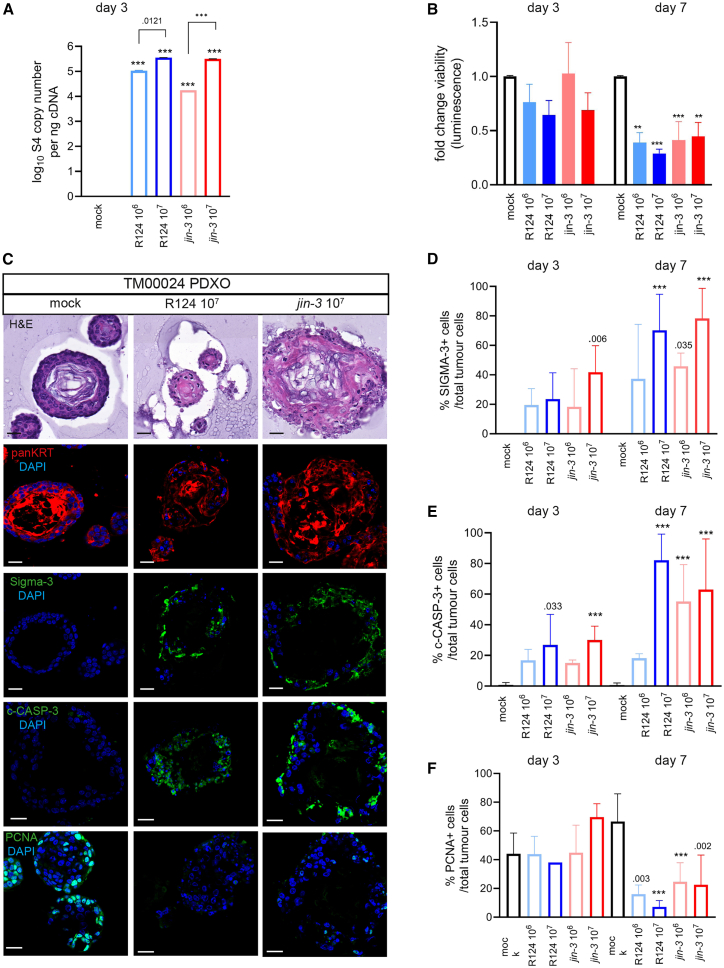 Figure 3
Figure 3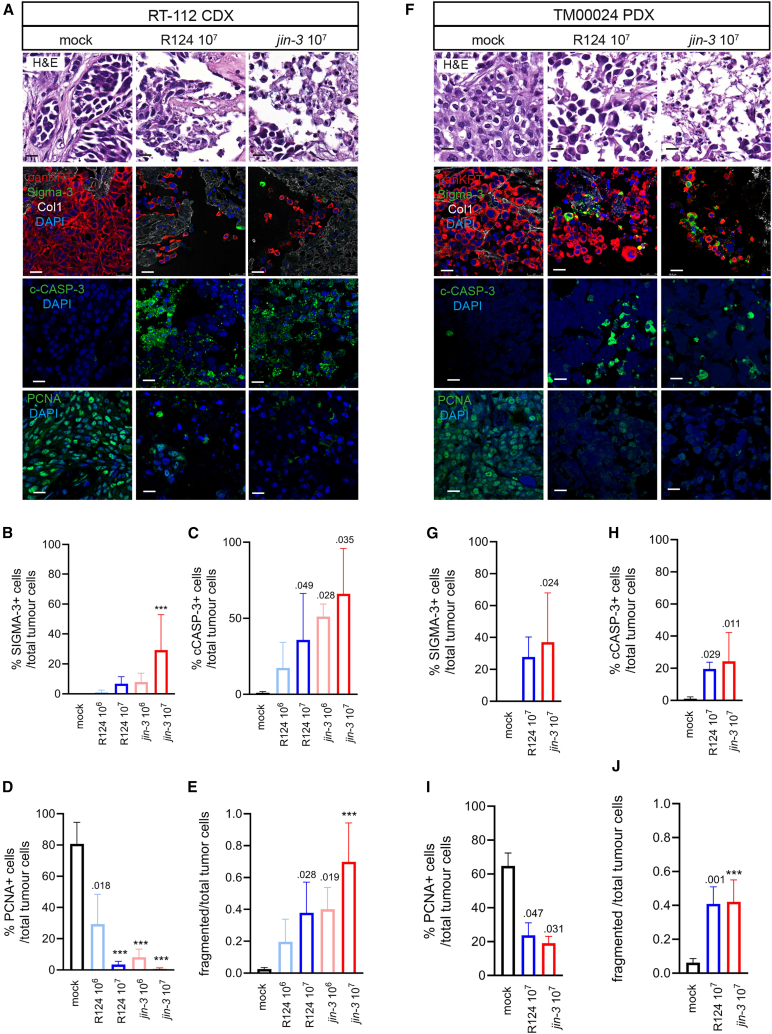 Figure 4
Figure 4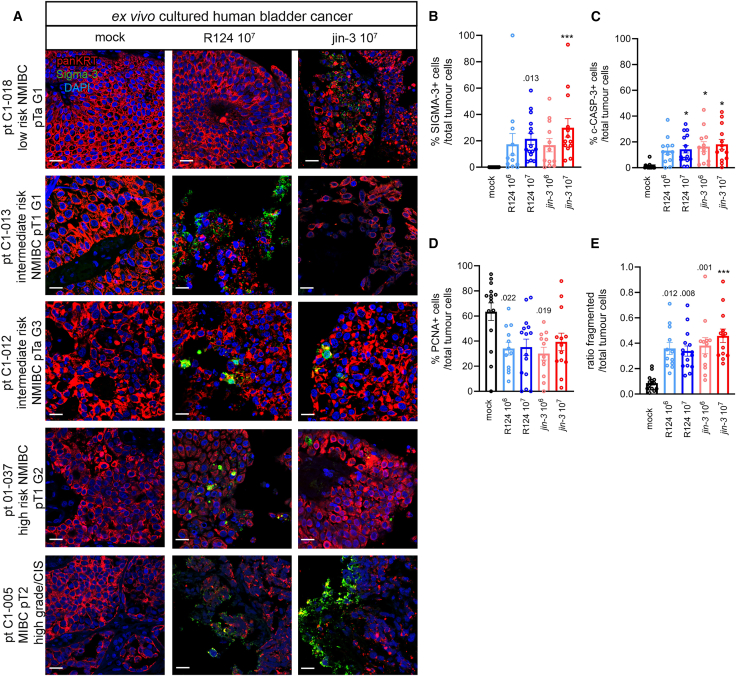 Figure 5
Figure 5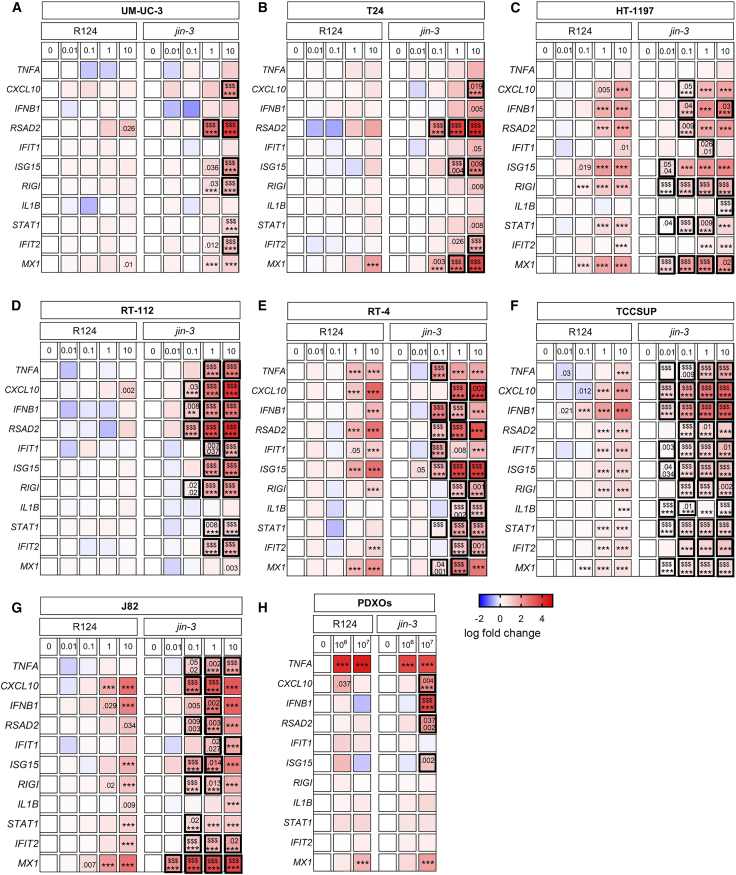 Figure 6
Figure 6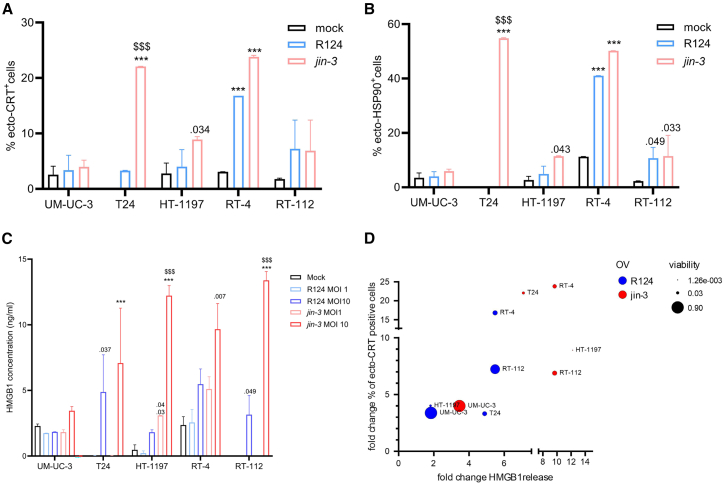 Figure 7
Figure 7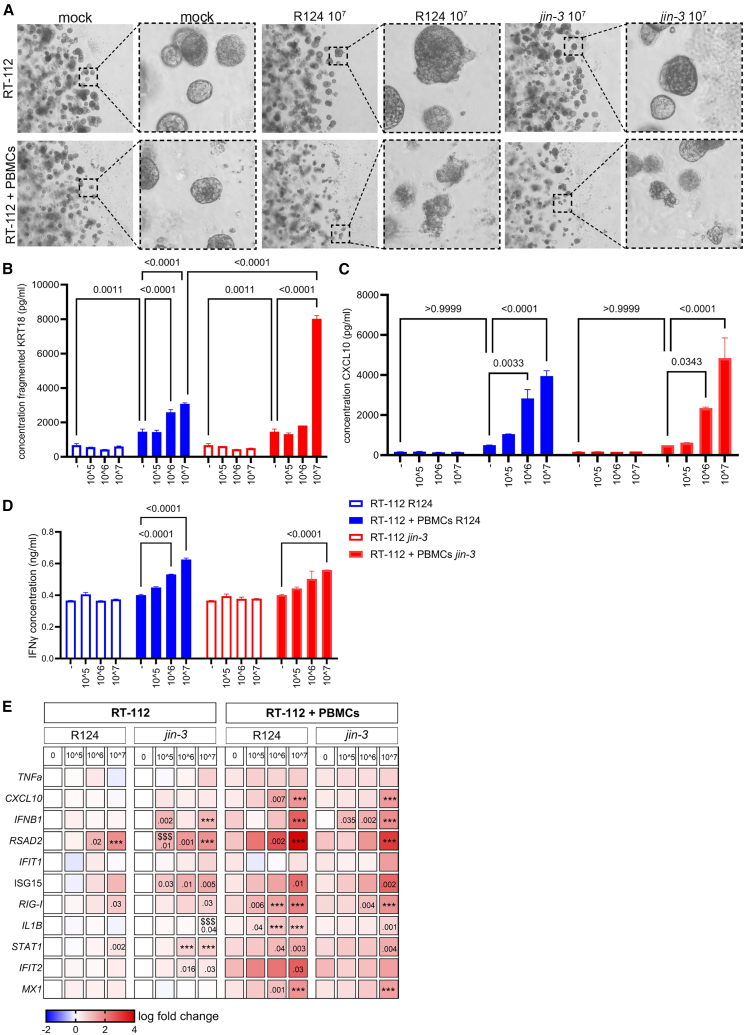 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVirus-based gene therapy research · Viral gastroenteritis research and epidemiology · Cancer Research and Treatments
Introduction
Urothelial carcinoma of the bladder (UCB) is the seventh most prevalent cancer globally, with a 5-year prevalence surpassing 5 million cases.1 Approximately 75% of newly diagnosed cases encompass non-muscle invasive bladder cancer (NMIBC), which are usually removed by transurethral resection.2 Patients are then categorized as low, intermediate, or high risk for recurrence and progression according to the guidelines of the European Urological Association.3 Current therapy for patients with intermediate- and high-risk NMIBC includes immunotherapy with Bacillus Calmette-Guérin (BCG) or chemotherapy. To date, BCG instillations have proven to be the most successful adjuvant therapy.3^,^4 However, 30%–50% of the patients fail to respond to BCG therapy, and 15% of patients show progression to the muscle invasive disease, MIBC.2^,^5^,^6 These MIBC patients have a relatively poor prognosis (5-year survival of 40%–60%). UCB recurs frequently after initial treatment and requires long-term clinical monitoring, causing substantial morbidity and diminished quality of life.4^,^7^,^8 Immunotherapy (including immune checkpoint inhibition [ICI], cell-based therapy, and cancer vaccines) is most effective in tumors with relatively high somatic mutation rates as is the case for UCB.9^,^10^,^11^,^12^,^13^,^14^,^15
ICIs, including pembrolizumab, nivolumab, atezolizumab, durvalumab, and avelumab, have improved the treatment landscape for bladder cancer, particularly for metastatic urothelial carcinoma.16
These ICIs have shown impressive and durable responses in bladder cancer patients.17^,^18^,^19^,^20^,^21^,^22^,^23^,^24^,^25^,^26^,^27^,^28 In high-risk NMIBC patients who are unresponsive to BCG, 41% of patients display a clinically desirable response to pembrolizumab (i.e., absence of high-risk NMIBC or progressive disease after 3 months).29 About 30% of patients with metastatic MIBC will respond to ICI treatment (objective response rate and overall survival),30 although these benefits appear to be limited to patients with pre-existing anti-tumor responses.25^,^31^,^32^,^33
Currently, ICIs are standard-of-care options in multiple clinical scenarios: as first-line therapy for cisplatin-ineligible patients, second-line therapy after platinum-based chemotherapy failure, and in novel combination regimens.
Despite these encouraging results in a subgroup of UCB patients, treatment modalities that can further improve immunotherapy are urgently required for unresponsive bladder cancer patients.
Oncolytic viruses represent a possible new approach since these viruses can infect, replicate, and lyse malignant tumor cells, while minimizing harm to normal cells.34 Furthermore, oncolytic viruses can activate the adaptive and innate immune system35^,^36^,^37^,^38 via the induction of immunogenic cell death (ICD), a type of cell death that triggers immunity against tumor antigens of dying cells, resulting in the exposure of damage-associated molecular patterns (DAMPs), e.g., HMGB1 release.39^,^40 DAMPs can activate antigen-presenting cells, which subsequently can stimulate the expansion and activation of effector T cells, including cytotoxic CD8+, helper CD4+ T cells, and natural killer (NK) cells.41^,^42
Reoviruses are double-stranded RNA viruses and have not been associated with severe disease in humans.43 Mutant reovirus jin-3 harbors a mutation in the spike protein Sigma-1 and can infect cells via the high-affinity receptor junction adhesion molecule A (JAM-A) but also independently of JAM-A on the tumor cell surface by high-affinity binding to negatively charged sialic acids. We previously demonstrated that jin-3 reoviruses display extended tumor tropism compared to the wild-type reovirus.44^,^45 Interestingly, this JAM-A protein is involved in various biological processes including tight junctions involved in epithelial barrier function.46 Expression levels of JAM-A in various cancers have been analyzed using multiple databases, including TIMER2.0, GEPIA2, and the UALCAN portal and was shown to be upregulated in multiple cancers.47 JAM-A was significantly upregulated in urothelial carcinoma compared to normal bladder. However, bladder cancer patients with low JAM-A expression showed decreased overall survival.47 Potentially, this reduced overall survival could be due to the acquisition of a more mesenchymal invasive phenotype (EMT) by the cancer cells due to the shedding of epithelial tight junctions.47 Mutational profile analysis (cBioportal based on TCGA datasets) revealed that JAM-A was altered in more than 15% of bladder cancer patients experiencing either amplification or mutation of JAM-A.47
In this study, the direct oncolytic and indirect immunomodulatory effects of reovirus mutant jin-3 were determined and compared to those of the wild-type T3D reovirus R124 in state-of-the-art preclinical bladder cancer models, including monolayer cultures and three-dimensional cell (co-) cultures and ex vivo cultured bladder tumor tissue slices derived from patients with disease ranging from low-grade NMIBC to MIBC.
Results
Direct cytolytic effects of jin-3 and R124 reoviruses
A panel of human bladder cancer cell lines of different molecular subtypes was exposed to a dose range of either R124 or jin-3, and viral expression levels were detected by RT-PCR after 24 h. In the UM-UC-3, HT-1197, and RT-4 cell lines, more viral transcripts were detected in the cells exposed to jin-3, whereas in the T24 and RT-112 cell lines either no difference or less viral transcripts were found (Figures 1A and S1A–S1C; Table S1). In addition, viral protein levels of Sigma-3 were determined in various tumor cells by flow cytometry (Figure 1B) or immunofluorescence (Figure 1C) after 72 h of treatment. A similar Sigma-3 pattern was observed, i.e., increased level of expression in jin-3-treated UM-UC3, HT-1197, and RT-4 and a similar expression level of Sigma-3 in T24 and RT-112 cell lines.Figure 1. Comparison of the oncolytic effects of mutant jin-3 versus R124 reovirus in bladder cancer cell lines in vitro(A) Mean viral S4Q copy number (log fold change S4Q mRNA expression vs. mock treated cells) in bladder cancer cell lines exposed to a range of MOI (0.01–0.1–1–10) of either R124 or jin-3 reoviruses after 24 h. In red, the highest infection with virus is shown. Two-way ANOVA followed by Tukey’s post hoc test. n = 3 (2 replicates). UM-UC-3 cells R124 reovirus MOI1 vs. mock p = 0.0184; ∗∗∗p < .001, p < .001, asterisks indicate mock versus reovirus infection, and dollar signs indicate R124 versus jin-3. (C) Confocal images of viral protein (Sigma-3, green) and DAPI (blue)-stained bladder cancer cells treated with MOI10 of R124 or jin-3 for 72h. Scale bars, 25 μm. (D) Mean percentage of viable cells after exposure to a range of MOI of either R124 or jin-3 for 6 days. n = 3 (6 replicates). p values are depicted (vs. mock; when 2 depicted upper p value is R124 vs. jin-3); ∗∗∗p < .001, $$$ p < .001, asterisks indicate mock versus reovirus infection, and dollar signs indicate R124 versus jin-3. (E) Percentage of single, viable, JAM-A protein expressing cells. Mean (SD), n = 3. MOI = multiplicity of infection.
Both jin-3 and R124 effectively killed the majority of bladder cancer cells in a dose- and time-dependent manner (Figures 1D and S1D). In cell lines of different type (MIBC or NMIBC; see Table S1) and molecular cancer subtypes, jin-3 and R124 displayed comparable anti-tumor effects in 5637 (mixed phenotype), RT-4 (luminal phenotype), and SW780 (basal phenotype). In the HT-1197, HT-1376 (both mixed phenotype), and RT-112 cells (luminal phenotype), jin-3 was found to be more effective.48 In the basal molecular subtype bladder cancer cell panel, T24 cells were significantly more effectively killed by jin-3 compared with wild-type reovirus R124, while both viruses were equally effective in TCCSUP cells. Strikingly, while R124 viruses did not affect the viability of the basal type UM-UC-3 cells, jin-3 reduced their viability. Moreover, in J82 cells, R124 viruses resulted in decreased viability only after exposure to high MOI, whereas exposure to jin-3 resulted in reduced viability at lower MOI.
R124 reoviruses enter cancer cells via its high-affinity receptor junction adhesion molecule A (JAM-A),44^,^49^,^50 while the spontaneous mutant jin-3 virus has a broader tropism due to its ability to also infect cancer cells independently of JAM-A, i.e., by binding negatively charged cell surface sialic acids.44
As expected, the expression levels of the canonical viral entry receptor JAM-A in tested cancer cell lines were in line with the ability of the reovirus variants to infect and/or replicate in the tumor cells (Figures 1E and S1G).
Upon challenge with R124 and jin-3, a dose-dependent increase in viral copy number was detected in 3D cultures of RT-112 cells (Figure S1H), coinciding with reduced viability of the treated tumoroids after 3 days (Figure S1I).
In organoids cultured from the patient-derived muscle-invasive bladder cancer xenograft model PDX TM00024, R124 and jin-3 showed dose-dependent infection and replication (Figure 2A).51 Furthermore, a significant reduction in cancer cell viability was observed after reovirus exposure for 7 days (Figure 2B). Viral protein (Sigma-3) was detected in the outer cell layers of the PDXOs, indicating active viral infection and replication after 3 days (Figures 2D and S2A).52^,^53 Moreover, an increased number of c-CASP-3-positive cells was measured (Figures 2E and S2A). Further increase of the Sigma-3- and c-CASP-3-positive cells was observed after 7 days (Figures 2C–2E and S2B for dose range), coinciding with a decrease in cell proliferation (nuclear PCNA; Figures 2C, 2F, and S2B) and loss of structural integrity of the PDXOs (Figures 2C and S2B). No significant differences were observed between the viruses.Figure 2. Comparison of the oncolytic effects of reovirus mutant jin-3 versus wild-type reovirus R124 in 3D cultures in vitroTM00024 bladder PDXOs were exposed to the indicated plaque-forming units (PFUs) of reoviruses for 3 or 7 days.(A) Mean (SD) viral copy number of TM00024 PDXOs treated for 3 days with the indicated PFUs of the reoviruses. Two-way ANOVA followed by Tukey’s post hoc comparison (n = 3) ∗∗∗p < .001. (B–F) Viability (mean [SD] was measured using cell titer glo 3D after days 3 and 7 of reovirus exposure. N = 3 (3 replicates). Two-way ANOVA followed by Tukey’s post hoc comparison. ∗∗∗p < .001. (C) Confocal images of PDXOs (day 7) stained for respectively H&E, panKRT (red); Sigma-3, c-CASP3, or PCNA (green); and DAPI (blue). Scale bars, 20 μm. Cells expressing the respective proteins Sigma-3 (D), c-CASP-3 (E), and nuclear PCNA (F) were counted with ImageJ and divided by the amount of panKRT+_DAPI+ cells. At least five fields were scored per technical replicate. Mean (SD) of N = 3 (3 replicates). ∗∗∗p < .001, asterisks indicate reovirus infection versus mock same day. Two-way ANOVA followed by Tukey’s post hoc comparison.
Subsequently, viral infection and replication was determined in tumor tissue slices derived from cell-line-derived xenograft (CDX) and patient-derived xenograft (PDX) models after reovirus exposure. Upon exposure of tumor tissue slices of RT-112 bladder cancer cells54 to R124 and jin-3 ex vivo for 3 days, the viruses were capable of infecting the cancer cells and replicated in viral factories in the cell (Figures 3A and 3B). Furthermore, apoptosis was stimulated upon reovirus exposure as assessed by a relative increase of c-CASP-3-positive cells coinciding with a decline in tumor cell proliferation (Figures 3C–3E and S3A for dose range). In another CDX model J82 (Figure S4), similar results were obtained. Moreover, in ex vivo cultured tumor slices obtained from the PDX model TM00024, which displayed cell membranous JAM-A protein expression (Figure S3C), reovirus exposure also induced increased apoptosis and impaired tumor cell proliferation (Figures 3F–3J and S3B). No significant changes were observed between the two viruses.Figure 3. Comparison of jin-3 and R124 reovirus infection in ex vivo cultured tumor tissueExplanted tissue slices from either RT-112 CDX (A–E) or TM00024 PDX (F–J) were exposed to the indicated PFU of R124 or jin-3 reovirus for 3 days. Tissues were stained for H&E, panKRT (red); Sigma-3, c-CASP3 or PCNA (green), type I collagen (white), and DAPI (blue). Representative confocal images are shown. Scale bars, 20 μm. Cells expressing the respective proteins were counted with ImageJ and divided by the number of panKRT+_DAPI+ cells. At least four fields were scored per technical replicate. Mean (SD) of N = 2 (4 replicates). The ratio fragmented tumor cells was measured by the number of fragmented cells/total tumor cells (E and J). ∗∗∗p < .001, reovirus infection versus mock. Mean (SD), n = 2 (4 replicates). Two-way ANOVA followed by Tukey’s post hoc comparison.
Consistent with earlier findings,47 JAM-A expression was upregulated in bladder cancer compared to normal bladder (TCGA-BLCA database), although low levels of JAM-A are observed in 103/412 samples (Figures S1E and S1F). Next, tumor tissue slices were generated from freshly obtained tissues from patients diagnosed with various stages of bladder cancer (n = 15; Table 1). Tissue slices were exposed to mock, R124, or jin-3 reoviruses for 3 days (Figures 4 and S5). Immunolocalization of JAM-A was determined in the patient tissues, and membranous localization was observed in all the patients, with one patient displaying lower levels of staining (pt C1-005; Figure S5B). Explanted tumor tissue was stained for c-CASP-3, PCNA (proliferation) and integrity of the cells (H&E staining and panKRT/DAPI staining). In the reovirus-treated patient-derived ex vivo cultured tumor explants, interpatient heterogeneity in Sigma-3 protein levels was observed, with Sigma-3 expression found in regions in multiple tissue slices per patient (Figures 4A and 4B). Exposure to reoviruses resulted in increased number of c-CASP-3-positive cells (Figures 4C and S5C), whereas the number of proliferating tumor cells was reduced (Figures 4D and S5C) as well as the integrity of the cells as indicated by the H&E staining and the increased ratio of fragmented cells (Figures 4E and S5A–S5C).Table 1. Patient information: clinical histopathological characteristics of the patients included. Disease stage and grade at time of sampling of the tumor materialPatient identifierDiseaseRisk group55Disease stageDisease gradeTreatment historyAgeGenderCISC1-005MIBC–pT2high-grade CISBCG bladder instillations70maleyes01–032MIBC–pT2G3unknown69malenoC1-015NMIBCvery highpT1focal G3none61maleno01–037NMIBChighpT1G2none82femalenoC1-020NMIBChighpTa/pT1G3none76malenoC1-013NMIBCintermediatepT1low gradenone87malenoC1-012NMIBCintermediatepTaG3none77maleno01–034NMIBCintermediatepTaG2none80malenoC1-008NMIBCintermediatepT1G1none62malenoC1-022NMIBCintermediatepTaG1none79malenoC1-047NMIBCintermediatepTaG1none64malenoC1-086NMIBClowpTaG1none67malenoC1-088NMIBClowpTaG1none73malenoC1-018NMIBClowpTaG1none67malenoC1-014NMIBClowpTalow gradenone70femaleNopTa, p(pathological) non-invasive papillary carcinoma; pT1, tumor invades subepithelial connective tissue; pT2, tumor invades muscle. Grading was scored using the WHO grading 1973 (G1: well differentiated; G2: moderately differentiated; G3: poorly differentiated) or WHO 2004 (low/high grade) MIBC, muscle-invasive bladder carcinoma; NMIBC, non-muscle invasive bladder carcinoma; CIS, carcinoma in situ; BCG, Bacillus Calmette-Guerin.Figure 4. Comparison of jin-3 and R124 reovirus infection in ex vivo cultured tumor tissue slices from human bladder cancer patientsExplanted tissue slices from patients (n = 15) diagnosed with bladder cancer were exposed to either mock, R124, or jin-3 reovirus for 3 days and stained for viral protein, c-CASP-3, PCNA (proliferation), and integrity of the cell (nuclear DAPI and panKRT tumor marker). (A–E) Representative confocal images of the mock and 10^7^ PFU condition of both viruses from five patients ranging from low-risk NMIBC to MIBC (viral protein [green, Sigma-3], tumor markers [red, panKRT], and nuclei [DAPI, blue]. Scale bars, 20 μm). Cells expressing the respective proteins [(B) Sigma-3, (C) c-CASP3, or (D) nuclear PCNA] were counted with ImageJ and divided by the number of panKRT+_DAPI+ cells. At least four fields were scored per technical replicate. Mean (SD) (3–4 replicates) with each dot representing one bladder cancer patient. The ratio of fragmented tumor cells was measured by the number of fragmented cells/total tumor cells (E). ∗∗∗p < 0.001, reovirus infection versus mock, one-way ANOVA followed by Tukey’ post hoc comparison.
Reovirus-induced immune activation
Inflammatory cytokines, type I interferons (IFNs), and IFN-stimulated genes (ISGs) are of key significance in oncolytic virotherapy that largely relies on the stimulation of anti-tumor and anti-viral immune cell activation.55^,^56^,^57 Exposure to R124 and jin-3 reovirus resulted in a significant and dose-dependent upregulation of IFN-β gene expression (IFNB1) after 24 h in multiple bladder cancer cell lines (Figures 5A–5G). jin-3 treatment resulted in a significantly stronger upregulation in IFNB1 (6/7) compared to R124 (4/7) in all the tested cell lines (Figures 5C–5G), except for UM-UC-3 and T24 (Figures 5A and 5B). Moreover, exposure to jin-3 led to significantly stronger induction of ISGs (i.e., ISG15, IFIT1, IFIT2, RSAD2, and MX1) in most of the tested cell lines compared to R124 (Figure 5). Exposure to jin-3 led to a superior and significant expression of inflammatory cytokines CXCL10 (in 7/7 cell lines), TNF (4/7), and IL1B (2/7) compared with R124 (Figure 5). Moreover, the cytosolic RNA sensor RIG-I was significantly upregulated after jin-3 exposure compared to R124 in 6/7 cell lines (Figure 5). In the TM00024 PDXOs, exposure to jin-3 reovirus resulted in significant upregulation of TNF, CXCL10, IFNB1, and RSAD2 gene expression, whereas R124 reovirus only induced TNF gene expression (Figure 5H).Figure 5. Comparison of immune modulation induced by jin-3 and R124 reovirus treatment of bladder cancer cell linesHeat maps of various inflammatory cytokines and interferon-stimulated genes (log fold change mRNA expression [2^−ΔΔCt^] vs. mock treated cells) in bladder cancer cell lines UM-UC-3 (A), T24 (B), HT-1197 (C), RT-112 (D), RT-4 (E), TCCSUP (F), J82 (G), and PDXOs (H) exposed to a range of either R124 or jin-3 reoviruses after respectively 24 h (cell lines) and 3 days (PDXOs). p values are depicted (vs. mock; when 2 depicted upper p value is R124 vs. jin-3) ∗∗∗p < .001, $$$ p < .001, asterisks indicate mock versus reovirus infection, and dollar signs indicate R124 versus jin-3.
Cell surface expression of the DAMPs ecto-calreticulin and ecto-HSP90 was significantly increased upon exposure to reoviruses (Figures 6A and 6B). Strikingly, jin-3 caused a more pronounced increase in HMGB1 release, a well-established marker of immunogenic cell death42 (Figure 6C). Correlation between these markers at an MOI of 10 is depicted in Figure 6D.Figure 6. Comparison of immunogenic cell death induced by jin-3 and R124 reovirus treatment of bladder cancer cell lines(A–C) Cell lines were treated with oncolytic viruses for 48 h at an MOI of 10, and the DAMPs ecto-calreticulin (A) and ecto-HSP90 (B) were measured using flow cytometry. Secreted HMGB1 protein was measured with an ELISA after 48 h of treatment with a dose range of oncolytic viruses (C). ∗∗∗p < 0.001, $$$ p < 0.001, asterisks indicate reovirus infection versus mock same day, and dollar signs indicate R124 vs. jin-3. Mean (SD), N = 3 (2 replicates). Two-way ANOVA followed by Tukey’s posthoc comparison. (D) Correlation graph of the HMGB1 release (x axis), percentage of viable cells (size of the dots), and fold change of ecto-CRT (y axis)-positive cells at MOI 10 of the indicated virus.
Subsequently, co-culturing of 3D tumoroids of human RT-112 cells with partially HLA-matched peripheral blood mononuclear cells (PBMCs) led to increased tumoroid killing (Figure S7A). This was accompanied with significantly increased release of fragmented cytokeratin-18 (KRT18; Figure S7B), CXCL10 (Figure S7C), and IFN-γ (Figure S7D). Addition of reoviruses to the 3D cultures resulted in expression of viral transcripts in the reovirus-treated RT-112 cells, with a lower copy number when the 3D cultures were co-cultured with PBMCs (Figure S7E). Reovirus treatment in the co-cultures resulted in augmented tumoroid killing compared to the mock-treated conditions, with jin-3 displaying significantly more pronounced effects (Figures 7A and 7B). Moreover, chemo- and cytokine production (CXCL10 and IFN-γ) and mRNA expression of type I IFN response genes (e.g., IFNB1, RSAD2, MX1, IFIT2, and ISG15 mRNA levels) was significantly enhanced upon reovirus treatment in these cultures (Figures 7C–7E and S7F–S7P).Figure 7. Activation of PBMCs in RT-112 co-cultures(A) Brightfield images of RT-112 bladder cancer cells that were cultured in 60% Matrigel and allowed to form 3D structures for approximately 3 days. Subsequently, PBMCs were added at an effector: target ratio of 20:1 in absence or presence of either R124 or jin-3 reovirus at the indicated PFU for an additional 3 days. (B) Fragmented KRT18 was determined as an outcome measure for tumor cell killing 3 days after OV exposure. (C–E) CXCL10 levels and (D) IFN-γ levels were measured 3 days after OV exposure. (n = 3, 2 replicates). Mean (SD). Two-way ANOVA with Tukey’s post hoc comparison. (E) Heatmap of various inflammatory cytokines and interferon-stimulated genes (log fold change mRNA expression [2^−ΔΔCt^] vs. mock treated cells) in 3D cultured RT-112 cells or RT-112 cells co-cultured with PBMCs exposed to either R124 or jin-3 reoviruses. Mean (SD). p values are depicted (vs. mock; when two depicted upper p value is R124 vs. jin-3) ∗∗∗p < .001, $$$ p < .001, asterisks indicate mock versus reovirus infection and dollar signs R124 versus jin-3., n = 2 (2 replicates). Two-way ANOVA followed by Tukey’s post hoc comparison.
Discussion
In this study, we evaluated the direct oncolytic and immunomodulatory effects of reoviruses R124 and jin-3 in preclinical human bladder cancer models.
We observed that reoviruses induce potent oncolytic effects in flatbottom 2D and 3D cultures of human bladder cancer. Despite the differences in oncolytic responses, which are likely to be due to well-documented clinical inter-tumoral heterogeneity in bladder cancer,58^,^59 we observed viral infection upon reovirus administration in most human bladder cancer cells. Moreover, we detected viral infection and replication in near-patient ex vivo cultured bladder cancer tissue slices derived from CDX and PDX models and primary bladder cancer patient material.
Besides oncolytic properties, oncolytic viruses can induce both anti-viral immunity and anti-tumor immune responses, which is required for long-lasting and systemic tumor control.60 Effective oncolytic virotherapy relies on the production of pro-inflammatory cytokines and type I IFNs and the induction of interferon-stimulated genes (ISGs). Pro-inflammatory chemokines and cytokines are released and attract lymphocytes toward the tumor.60^,^61^,^62 These anti-viral immunological events could turn immunological “cold” tumors into “hot” tumors. Our study demonstrated that jin-3 reovirus led to elevated expression of ISG and inflammatory cytokine production by infected bladder cancer cells compared to R124 wild-type reovirus. This is further substantiated by the augmented release of immunogenic cell death markers HMGB1 and DAMPs (i.e., ectopic cell surface expression of calreticulin and HSP90 after exposure to jin-3 reovirus). Co-culturing of bladder tumoroids with PBMCs resulted in a significant, dose-dependent increase in tumor cell death and elevated production of CXCL10 and IFN-γ after reovirus exposure. This is consistent with findings in a colorectal cancer co-culture system in which autologous PBMCs acquire tumor-targeting capabilities, which correlated with clinical responses to checkpoint inhibitors.63
Overall, our preclinical results support the notion that infection of the human bladder cancer cells with reoviruses, especially jin-3, induces oncolysis and may contribute to a switch from an immunologically “cold’’ toward an immunological “hot’’ inflammatory phenotype. Additionally, jin-3 has an extended tropism compared to the wild-type reovirus because these viruses can infect cells independently of JAM-A expression on the tumor cell surface. These results are in line with previous reports in prostate cancer cells showing that intra-tumoral injection of wild-type reovirus induced pro-inflammatory cytokines production.45^,^64
Intravesical administration of reovirus in an orthotopic model of superficial bladder cancer revealed minor side effects of therapy with significantly higher tumor-free survival compared to BCG treatment.65 Moreover, in a syngeneic mouse model, intravesical therapy of reovirus in combination with an anti-programmed cell death protein 1 (PD-1) antibody improved survival and induced a more immune-active (hot) tumor microenvironment.66
Previous clinical studies have also demonstrated the potential of oncolytic viruses in cancer therapy, highlighting their ability to target and destroy cancer cells selectively. For example, T-VEC is a Food and Drug Administration (FDA)-approved herpes-simplex-virus-based therapy for advanced melanoma.67 Other oncolytic viruses including vesicular stomatitis virus, adenovirus, and vaccinia virus have been studied in multiple cancer types, including bladder cancer.68^,^69^,^70^,^71^,^72^,^73^,^74^,^75^,^76
Some oncolytic viruses, for example, ICAM-1 targeted Coxsackievirus A21 (CVA21) and vaccinia virus, have shown acceptable safety profiles and proof of viral targeting and replication in the bladder tumor cells in clinical trials.68^,^71^,^77 In addition, in early stage bladder cancer, oncolytic adenoviruses have been efficacious in BCG-refractory patients.78^,^79^,^80^,^81^,^82^,^83^,^84^,^85
Importantly, clinical trials have revealed that pelareorep, also known as Reolysin, an isolate of wild-type T3 Dearing reovirus, can be safely administered to cancer patients, including patients with advanced cancer such as multiple melanoma, prostate, and esophagus cancer, with relatively few adverse effects.43^,^49^,^86^,^87^,^88^,^89 However, the clinical benefit of wild-type reovirus administration is limited, and phase II studies have revealed no survival benefit.90 No clinical trials with reovirus have been reported for bladder cancer yet. Clinical benefit may be enhanced by using our reovirus mutant jin-3 with an expanded tropism, beyond the canonical reovirus entry receptor JAM-A.44
Due to the significant variability between and within tumors—including genetic mutations and differences in the tumor microenvironment—it is essential to develop diverse and improved oncolytic virotherapy strategies using multiple types of oncolytic viruses.
Matching the most effective oncolytic virus(es) to an individual patient is key for virotherapy to become more effective. Stratification of patients who are more likely to respond to a specific oncolytic virus is required but—to date—not yet accomplished. If proven, this personalized approach will enhance treatment outcomes by matching the best candidate virus to an individual tumor based on its unique biological characteristics.
Accumulating evidence shows that oncolytic virotherapy, when used as a monotherapy, is unlikely to fully overcome the immunosuppressive tumor microenvironment in a clinical setting. Several clinical studies are with oncolytic reovirus in combination with immune checkpoint inhibition in various cancers including pancreatic adenocarcinoma, metastatic non-small lung cancer, multiple myeloma, and metastatic colorectal cancer.91^,^92^,^93^,^94^,^95^,^96^,^97 For example, in the GOBLET multi-cohort phase 1/2 study in pancreatic and anal cancer, intravenous pelareorep, in combination with the PD-L1 inhibitor atezolizumab, displayed promising efficacy (respectively 62% and 37.5% objective response rate for metastatic pancreatic and anal cancer).98 Furthermore, combination of pelareorep with the PD-1 inhibitor pembrolizumab modestly promoted anti-tumor activity in advanced pancreatic cancer.99
To better understand the interactions between oncolytic viruses and the immune system, tumor-immune cell co-culture systems—as the ones described here—can be exploited. By incorporating both tumor cells and relevant immune cell populations (e.g., T cell populations and dendritic cells), these systems can be used to assess how different OVs modulate immune cell activation, suppression, or recruitment, to monitor cytokine and chemokine production in response to viral infection and to evaluate the potential for synergy or antagonism when combining oncolytic viruses with current immunotherapy (immune checkpoint inhibition or cell-based immunotherapy). Correct timing, optimal oncolytic virus (hence the repertoire of OVs), optimal dosing, and treatment combinations are key.
Collectively, our findings underscore the oncolytic potential of reoviruses in diverse preclinical models of human bladder cancer. Notably, the mutant reovirus jin-3 exhibits a more robust immunomodulatory response in preclinical bladder cancer models compared to the wild-type reovirus R124. This promising result positions jin-3 as a compelling candidate for future clinical translation.
Material and methods
Reovirus production
Wild-type T3D reovirus strain R124 was plaque-purified from the wild-type reovirus T3D (ATCC) on HER911 cells.44^,^100 Reovirus mutant jin-3 was isolated from JAM-A-deficient U118MG cells after passaging of the wild-type T3D strain R124.44 Both R124 and jin-3 reoviruses were propagated, purified, and titrated on human HER911 cells as previously described.44
Preclinical models and culture conditions
Cell lines as monolayers: a panel of human bladder cancer cell lines was cultured in monolayers with the respective culture medium (Table S1).
Three-dimensional bladder cancer tumoroids or patient-derived xenograft (PDX) organoids (PDXOs) were cultured from cells isolated from subcutaneously growing PDX TM00024 tumors51 (Jackson Laboratory; ethical permit AVD16605) or from the RT-112 and J82 cell lines as described previously.52
Three-dimensional tumor-immune cell co-cultures: 10,000 RT-112 cells were seeded in 40 μL domes in 60% Matrigel. After 3 days, partially HLA-matched PBMCs (Sanquin NVT0631.01) were added at a 20:1 effector target ratio to the 3D cultures. After 3 days, conditioned medium was collected, and RNA was isolated as described below.
Ex vivo cultured tissue explants were sliced and cultured as previously described.54 Bladder cancer tissue was derived from either CDX, PDX, or directly from patient tumor material (Biobank Urology protocol B20.060, animal ethical protocol AVD16605; Table 1). Multiple tissue slices were used per condition and cultured for 4 days.
Viability assays
Monolayer experiments: 1,500 cells per well were seeded in a 96-well plates. After 24 h, these cells were exposed to a range of either R124 or jin-3 (MOI 0–100 plaque-forming units [PFUs]/cell). Cells were incubated for 72 or 144 h, after which a CellTiter 96 AQueous One Solution Cell Proliferation Assay (Promega) was performed according to manufacturer’s protocol. After 2 h of incubation time, optical density was measured at 490 nm using SpectraMax iD3 plate reader to assess cell viability.45
The 3D tumoroids or PDXOs were exposed to a range of R124 and jin-3 reoviruses. After 3–7 days, viability was determined using the Cell Titer Glo assay, according to the manufacturer’s protocol (Promega).45
Flow cytometry
Four hundred thousand cells/well were seeded in a 6-well plate. Each cell line was exposed to either R124 or jin-3 (mock solution only, MOI 10) and incubated for 48 h. Subsequently, levels of viral protein expression (Sigma-3), apoptosis (cleaved caspase-3 [c-CASP-3]), and protein expression of several DAMPs (cell surface expression of calreticulin and HSP90) for each cell line/virus combination were acquired using LSR Fortessa X-20 Cell Analyzer and analyzed using FlowJo software (Table S1).
Histology and immunofluorescence
H&E and immunofluorescent stainings were executed as previously described.54 H&E and immunofluorescent stainings were visualized using the Pannoramic MIDI slide scanner (3DHISTECH). All fluorescent stainings were visualized by confocal microscopy (63× magnification, resolution 1,024 × 1,024) (Leica SP8). At least four fields of the fluorescent staining were scored per TS by two independent reviewers. The average of these 4 fields was shown for each of the technical replicates. Cells expressing the respective proteins were counted with ImageJ and divided by the number of pancytokeratin-positive and DAPI-positive (panKRT+_DAPI+) cells (i.e., intact tumor cells with intact nuclei). The ratio of fragmented tumor cells was measured by the number of fragmented cells divided by the number of total tumor cells.
Real-time quantitative polymerase chain reaction
Cells were infected with the viruses and after the indicated time RNA was isolated according to the manufacturer’s protocol (Nucleospin RNA kit Macherey-Nagel). cDNA was generated by using random primers (Promega), and real-time qPCR was performed with GoTaq Mastermix (Promega) according to the manufacturer’s protocol in technical duplicates and biological triplicates (Promega). Gene expression was normalized to GAPDH. Primer sequences can be found in Table S1.
ELISAs
Cells were exposed to R124 or jin-3 with an MOI of 10. At the indicated time post exposure, HMGB1 (high mobility group box 1) release was measured by performing the Lumit HMGB1 Human/Mouse Immunoassay (Promega).
Three-dimensional (co-) cultures were exposed to R124 or jin-3. At the indicated time points post exposure, conditioned medium was collected, and either fragmented KRT18 (Abcam), IFN-γ (Abcam), or CXCL10 (Invitrogen) levels were measured as indicated by the manufacturers protocol.
Statistical analysis
Statistical analyses were performed by using GraphPad Prism 8.0. Two-way ANOVA was performed for viability and ELISA experiments followed by Tukey post hoc test for multiple comparison. For ex vivo cultures, sample size was calculated using proportional odds models as described.101 Multiple slices were scored per condition per patient (∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001).
Data and code availability
Data supporting the findings of this study are available within the paper and its supplemental information.
Acknowledgments
This study was supported by the OAK Foundation (www.supportcasper.nl;STRATIVIR and SOAK18.01), the 10.13039/501100004622KWF Dutch Cancer Society (KWF15932), and a kind gift from the Franje Foundation.
Author contributions
Conceptualization, R.H., G.P., and G.H.; data curation, A.F., M.M., and G.H.; formal analysis, A.F., M.M., L.H., D.W., and G.H.; funding acquisition, R.H., G.P., and G.H.; investigation, A.F., M.M., L.H., D.W., and G.H.; methodology, A.F., M.M., D.W., and G.H.; project administration, G.P. and G.H.; resources, R.H., D.W., R.F., and W.Z.; supervision, R.H., G.P., and G.H.; validation, A.F., M.M., G.P., and G.H.; visualization, A.F., M.M., and G.H.; writing—original draft, A.F. and G.H.; writing—review and editing, A.F., R.H., M.K., G.P., and G.H.
Declaration of interests
The authors declare no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wéber A.Vignat J.Shah R.Morgan E.Laversanne M.Nagy P.Kenessey I.Znaor A.Global burden of bladder cancer mortality in 2020 and 2040 according to GLOBOCAN estimates World J. Urol.4220242373862541710.1007/s 00345-024-04949-8PMC 11021283 · doi ↗ · pubmed ↗
- 2Babjuk M.Burger M.Compérat E.M.Gontero P.Mostafid A.H.Palou J.van Rhijn B.W.G.Rouprêt M.Shariat S.F.Sylvester R.European Association of Urology Guidelines on Non-muscle-invasive Bladder Cancer (Ta T 1 and Carcinoma In Situ) - 2019 Update Eur. Urol.7620196396573144396010.1016/j.eururo.2019.08.016 · doi ↗ · pubmed ↗
- 3Zhang Y.Zhang X.Huang X.Tang X.Zhang M.Li Z.Hu X.Zhang M.Wang X.Yan Y.Tumor stemness score to estimate epithelial-to-mesenchymal transition (EMT) and cancer stem cells (CS Cs) characterization and to predict the prognosis and immunotherapy response in bladder urothelial carcinoma Stem Cell Res. Ther.142023153672121710.1186/s 13287-023-03239-1PMC 9890713 · doi ↗ · pubmed ↗
- 4Schmidt S.Kunath F.Coles B.Draeger D.L.Krabbe L.M.Dersch R.Kilian S.Jensen K.Dahm P.Meerpohl J.J.Intravesical Bacillus Calmette-Guérin versus mitomycin C for Ta and T 1 bladder cancer: Abridged summary of the Cochrane Review Investig. Clin. Urol.61202034935410.4111/icu.2020.61.4.349PMC 732964532665991 · doi ↗ · pubmed ↗
- 5Chan K.S.Volkmer J.P.Weissman I.Cancer stem cells in bladder cancer: a revisited and evolving concept Curr. Opin. Urol.2020103933972065728810.1097/MOU.0b 013e 32833 cc 9df PMC 2997612 · doi ↗ · pubmed ↗
- 6Chang J.T.Mani S.A.Sheep, wolf, or werewolf: cancer stem cells and the epithelial-to-mesenchymal transition Cancer Lett.341201316232349989010.1016/j.canlet.2013.03.004PMC 3762892 · doi ↗ · pubmed ↗
- 7Ferlay J.Steliarova-Foucher E.Lortet-Tieulent J.Rosso S.Coebergh J.W.W.Comber H.Forman D.Bray F.Cancer incidence and mortality patterns in Europe: estimates for 40 countries in 2012 Eur. J. Cancer 492013137414032348523110.1016/j.ejca.2012.12.027 · doi ↗ · pubmed ↗
- 8Dyba T.Randi G.Bray F.Martos C.Giusti F.Nicholson N.Gavin A.Flego M.Neamtiu L.Dimitrova N.The European cancer burden in 2020: Incidence and mortality estimates for 40 countries and 25 major cancers Eur. J. Cancer 15720213083473456037110.1016/j.ejca.2021.07.039PMC 8568058 · doi ↗ · pubmed ↗