Independent, ongoing clade-specific expansions of IS5 elements in Pseudomonas syringae
David A. Baltrus, Audrey Sweten, Thomas Conomos, Nathaniel Ponvert, Jesse D. Woodson, Zachary Konkel, Jonathan Jacobs

TL;DR
This study shows that IS5 elements in Pseudomonas syringae have expanded independently in different clades, affecting gene expression and genome dynamics.
Contribution
The paper reveals independent, recent expansions of a specific IS5 subclass in multiple P. syringae clades.
Findings
A subclass of IS5 elements has undergone independent expansions in multiple P. syringae clades.
Most IS5 insertion sites are not conserved across closely related P. syringae pv. lachrymans genomes.
These IS5 elements can influence the expression of downstream genes in P. syringae.
Abstract
Insertion sequence (IS) elements are transposable regions of DNA present in a majority of bacterial genomes. It is hypothesized that differences in distributions of IS elements across bacterial strains and species reflect underlying differences in population biology. Therefore, shifts in IS element distributions between closely related strains may be proxies for and reflective of changes in population dynamics. Here, we investigate the presence and distribution of a subclass of IS5 elements throughout genomes of Pseudomonas syringae by querying complete genomes for the presence of InsH (the main transposase found within these IS5 elements). We report that this one subclass of IS5 elements appears to have recently undergone independent expansions in multiple P. syringae clades and find that a majority of IS5 insertion sites are not conserved across three closely related P. syringae pv.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
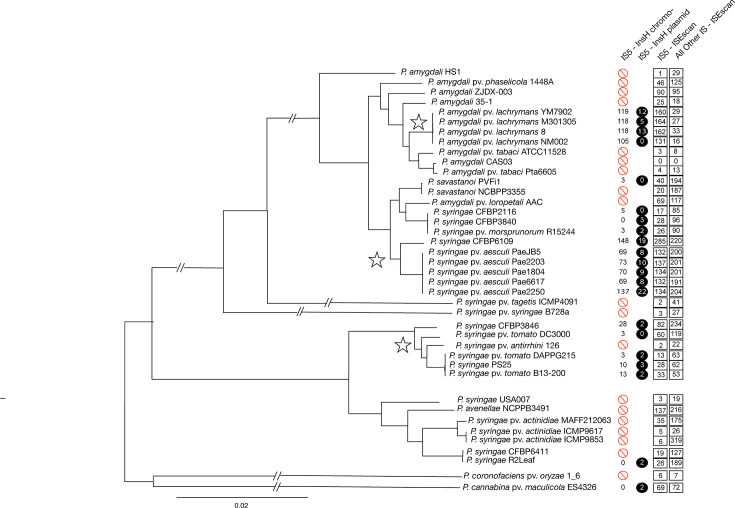 Fig. 1
Fig. 1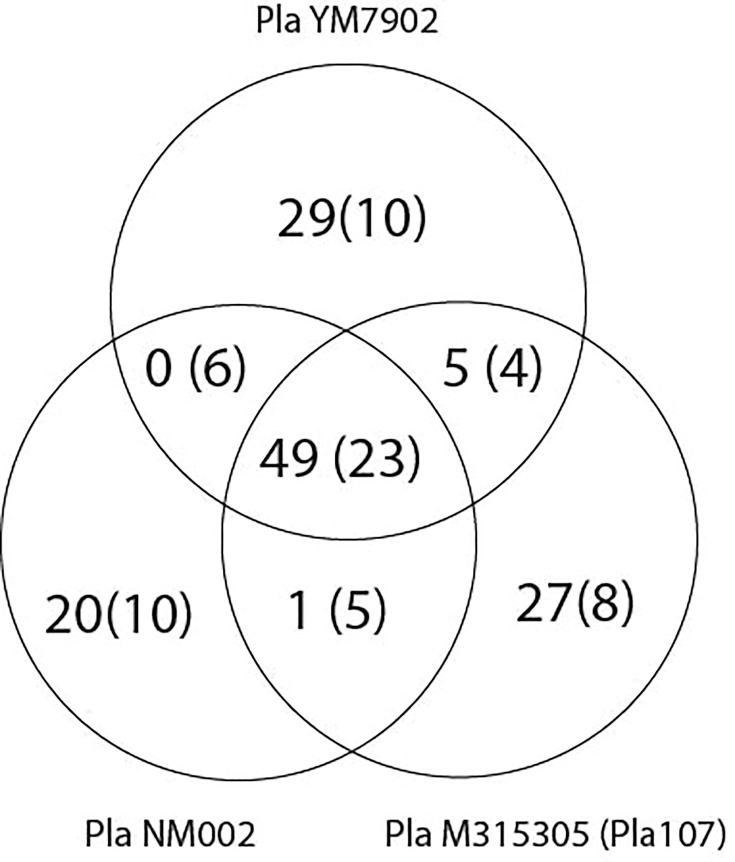 Fig. 2
Fig. 2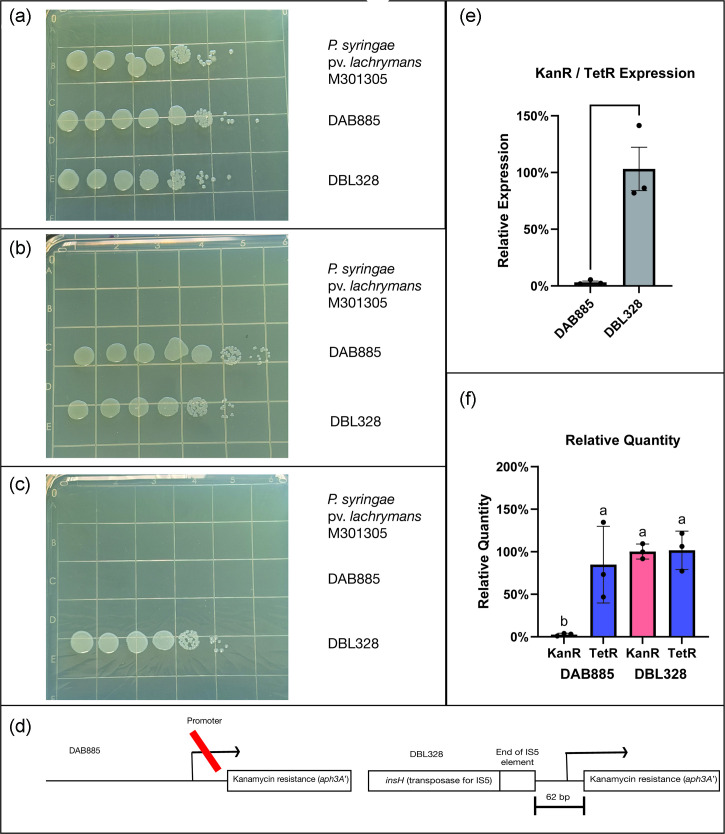 Fig. 3
Fig. 3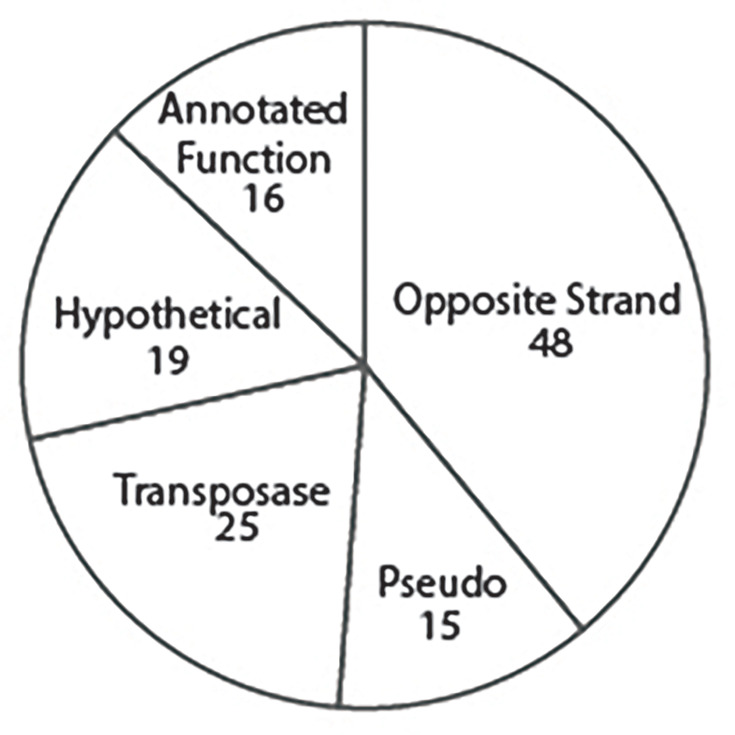 Fig. 4
Fig. 4| Locus tag of IS | RefSeq ID of downstream gene | Distance between InsH and gene (bp) | Functional annotation |
|---|---|---|---|
| PLA107_RS09245 | 285 | GO:0006935 – chemotaxis | |
| PLA107_RS09315 | 184 | GO:0003700 – DNA-binding transcription | |
| PLA107_RS10475 | 513 | GO:0008168 – methyltransferase activity | |
| PLA107_RS11585 | 260 | GO:0016740 – transferase activity | |
| PLA107_RS15180 | 264 | GO:0004222 – metalloendopeptidase activity | |
| PLA107_RS18200 | 228 | Alpha/beta fold hydrolase MenH | |
| PLA107_RS20965 | 802 | GO:0016746 – acyltransferase activity | |
| PLA107_RS23725 | 254 | DABB family protein | |
| PLA107_RS24535 | 328 | GO:0003987 – acetate-CoA ligase activity | |
| PLA107_RS25360 | 413 | GO:0016432 – tRNA-uridine aminocarboxypropyltransferase activity | |
| PLA107_RS25860 | 154 | GO:0003677 – DNA binding | |
| PLA107_RS27250 | 320 | GO:0009116 – nucleoside metabolic process | |
| PLA107_RS28260 | 264 | GO:0008754 – O antigen ligase activity | |
| PLA107_RS28390 | 850 | GO:0003677 – DNA binding | |
| PLA107_RS28465 | 272 | GO:0022857 – transmembrane transporter | |
| PLA107_RS32845 | 290 | 3-oxoacyl-[acyl-carrier-protein] synthase III |
- —http://dx.doi.org/10.13039/100000001 National Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · Genomics and Phylogenetic Studies · Bacteriophages and microbial interactions
Data Summary
Sequencing reads used for this assembly have been deposited in the SRA at accession SRR30528424. The genomic assembly of strain DBL328, generated from these reads, has been deposited at Figshare [1]. Other supplementary files and data are also available at this Figshare link.
Introduction
Insertion sequence (IS) elements are a diverse class of transposable gene regions found throughout bacterial genomes, but are largely thought of as genomic parasites that proliferate within a genome [25]. Although IS elements are widespread throughout bacterial genomes and their importance in the creation of genetic and genomic novelty is clear, there have been relatively few investigations of their proliferation and evolutionary dynamics within and between lineages and species. Multiple factors contribute to this relative lack of studies; for instance, repetitive sequences the length of IS elements are often among the most fragmented regions when draft genomes are assembled from ‘short’ read sequences alone and thus it can be difficult to generate accurate counts and place IS elements in proper genomic context. Moreover, even when genomes are completely assembled, it can be challenging to compare syntenic relationships of IS elements across closely related strains. With these ideas in mind and with an interest in highlighting the evolutionary importance of IS elements, we present an analysis of the evolutionary plasticity of an IS5 element across complete genomes of relatively closely related strains of the plant-associated and sometimes pathogenic Pseudomonas syringae and demonstrate the capability of this element to drive downstream gene expression in this bacterium.
IS elements are, at a minimum, composed of regions of DNA bracketed on both sides by inverted repeat sequences recognized by a transposase enzyme enabling excision/insertion of the region from/into a position in the genome [5]. In many cases, the IS element itself encodes a transposase enzyme that can recognize and excise the inverted repeat regions, enabling the entire gene to autonomously ‘jump’ or transpose to other regions of DNA within the same cell. Depending on the type of element, this jump can occur by a zero-sum ‘cut and paste mechanism’ or can be replicative and allow for proliferation of this IS element to higher copy numbers throughout the genome. Notably, multiple IS elements can combine to form compound transposons, which can carry and transfer additional beneficial genes such as those involved in antibiotic resistance [6]. Despite numerous examples where IS element disruption is highly beneficial, the overwhelming majority of evidence suggests that IS element transpositions are predominantly either neutral or deleterious [5,711]. Proliferation of IS elements within and across genomes is therefore thought to be governed by a combination of selection against transposition jumps that are deleterious, coupled with genetic drift (potentially through population bottlenecks) enabling proliferation and expansion of multiple copies of these genes [12], with occasional but rare selection for strong beneficial effects. For instance, in extreme cases with populations of parasitic bacteria that are bottlenecked frequently during host transmission, the number of IS elements within a genome can explode because of excess genetic drift coupled with reduced selection pressures [13]. Due to their perceived neutrality, changes in distributions of IS elements across genomes are therefore hypothesized to correlate with and potentially indicate changes in the population biology of lineages of interest.
Lastly, elements classified in the IS5 family are a particularly interesting group because they have been relatively well-studied for their ability to ‘hotwire’ expression of genes and operons within their proximity [1417]. Although there have been numerous examples of transcriptional upregulation due to IS5 elements, the specific molecular events and genomic contexts enabling them to act as promoters remain unclear [141618].
P. syringae (sensu lato) is a bacterial species complex largely considered to be facultative phytopathogens commonly found associated with plants, but whose environmental persistence and transmission may also be closely tied to the water cycle [1920]. There have now been thousands of genomes sequenced for strains designated within the P. syringae species complex*.* IS elements are often found within these genomes and often disrupt genes that can contribute to virulence. However, given that repetitive regions like IS elements are also the most poorly assembled parts of the genome, there have been few large-scale assessments or general comparisons of IS element compositions across P. syringae. Here, we present an in-depth comparison of one subclass of IS5 elements across complete (or nearly complete) genomes, and we show that copy numbers for this particular element appear to have substantially increased independently in strains of pathovar (pv.) aesculi (a pathogen of horse chestnut) and pv. lachrymans (pathogens of cucumber). While these patterns are somewhat reflective of general IS element proliferation throughout these genomes, there is no clear general signal for proliferation across all genomes. We present further evidence comparing IS5 element positioning across three complete pv. lachrymans genomes, highlighting recent and ongoing changes in the number and position of IS5 elements. Since IS5 elements have the potential to act as promoters enabling transcription of downstream regions in other species, we further demonstrate the ability of this element to drive gene expression of an otherwise silent antibiotic resistance gene in one of these pv. lachrymans strains. Taken together, our results suggest that multiple lineages of P. syringae have experienced extensive proliferation of an IS5 element, and we suggest that this genomic signature could reflect changes in population biology for this lineage compared to other P. syringae clades. However, it is also possible that gene expression changes and disruptions caused by this element are uniquely beneficial in the context of these pv. lachrymans and pv. aesculi pathogens.
Methods
IS5 element characterization across complete genomes
The IS5 element of P. syringae pv. lachrymans was originally identified from the PGAP annotation [21] of the Pla107 genome. The complete protein sequence of the transposase (InsH) of this IS5 element was used as a search query in using the DiamondblastP function against all complete genomes at https://www.pseudomonas.com/ (>1,000 at the time of query) with retention of hits that were >70% length of the original IS element and >98% protein identity [22]. All other comparative genomic data (specifically blastP hits) were acquired by accessing the DiamondblastP option at Pseudomonas.com (database version 22.1, accessed on 24 November 2023). We note that DiamondblastP will only identify IS elements with annotated InsH sequences and therefore that overall numbers represented for each strain represent a minimum number of IS5 elements within each genome. Query sequences and a spreadsheet file of the blastP results can be found at Figshare [1].
Characterization of all IS elements across P. syringae genomes
To further investigate IS element composition across P. syringae genomes, we queried all IS element classes within complete genome sequences which contained sequences for the InsH protein from blast searches as described above, as well as additional representative genomes (from phylogenetically informative strains). IS elements were predicted using Pseudomonas genome assemblies via ISEscan v1.7.2.3 with default parameters [23]. GenBank accessions for each genome queried can be found as an additional file at Figshare [1].
Phylogenetic comparisons
We inferred phylogenetic relationships across all complete P. syringae genomes that contained an IS5 element, as well as representative complete genomes from phylogenetically informative or important clades, using Realphy [24] and by designating Pph1448a, PsyB728a, PtoDC3000, Por1_4, PmaES426 as references and with default parameters. Genome accessions for each of these assemblies and Realphy outputs can be found at Figshare [1].
Comparison of IS5 synteny and position across three complete P. syringae pv. lachrymans genomes
For comparison across three strains of P. syringae pv. lachrymans, completely sequenced chromosomes for P. syringae pv. lachrymans M301350, P. syringae pv. lachrymans NM002 and P. syringae pv. lachrymans YM7902 were queried for positions of annotated InsH sequences as above using https://www.pseudomonas.com/. We note that all three genome sequences were annotated using the NCBI PGAP pipeline, and thus annotations should be adequately comparable across all three strains. blastP was further used to search annotated protein sequences across all three genomes using loci upstream and downstream of each identified IS5 element sequence to identify proximate regions of synteny across the three chromosomes of these strains. If a blast search failed to retrieve hits >98% for proteins of interest (or if the query was itself an IS5 element), we queried at least three additional loci upstream or downstream of the focal IS5 element to establish synteny. If syntenic insertion of an IS5 element could be clearly established by hand annotation across two or three genomes or if it was obvious that the focal copy of an IS5 element was not present within a genome, we placed this IS5 element into a ‘clear’ group. If synteny could not easily be established due to rearrangements or other genomic variation, and the surrounding genomic region was likely present within each genome in some context, we place the focal element into an ‘unclear’ annotation group. Since we performed this search by hand, we note that there were a small number of cases where there were full-length IS5 elements and pseudogenized IS5 elements at syntenic positions across the genomes and we counted these as syntenic positions.
Selection of an IS5 element driving kanamycin gene expression
Pseudomonas amgydali pv. lachrymans 107 (Pla107, also known as MAFF301315 and PlaN7512) was originally isolated from diseased cucumbers (Cucumis sativus) in Japan in 1975 and deposited at the Ministry of Agriculture, Fisheries and Forestry, Japan (MAFF no. 301315). The isolate used to derive strains for this report was directly acquired from MAFF, and the complete genome assembly of this strain was originally reported in Smith et al. [25] found at GenBank at accession GCA_000146005.2. Originally, a phenotypically marked version of strain Pla107 was created (named DAB885) through the integration of the vector pMTN1907 into a region on megaplasmid pMPPla107 and using positive selection for recombinants through tetracycline resistance [2627]. Although pMTN1907 also contains an aph3A′ locus from Campylobacter coli with expression driven by its native promoter, and this locus enables kanamycin resistance in Escherichia coli, this gene as constituted does not enable Pla107 to grow on plates supplemented with kanamycin [28]. An isolate of this strain was grown overnight in King’s B medium (KB) supplemented with tetracycline (10 µg ml^−1^) on a shaking incubator at 27 °C at 220 r.p.m. After growth overnight, 200 µl of this culture was spread on KB agar plates supplemented with kanamycin (25 µg ml^−1^) and a single kanamycin-resistant colony was picked to KB liquid media. This culture was grown overnight and frozen down in the Baltrus Lab stock collection as strain DBL328. For genome sequencing, a subsample of this frozen stock was streaked to KB agar plates containing tetracycline and kanamycin, and subsequently a single colony was picked to KB liquid media supplemented with kanamycin. Genomic DNA from DBL328 was isolated from this liquid culture using a Promega (Madison, WI) Wizard kit, sequenced at Plasmidsaurus using standard protocols and assembled using their standard pipeline involving Flye v. 2.1 [29]. Sequencing reads used for this assembly have been deposited in the SRA (Sequence Read Archive) at accession SRR30528424. The genomic assembly of strain DBL328, generated from these reads, has been deposited at Figshare [1]. The coding sequence of the aph3A′ locus on the megaplasmid of pMPPla107 in DBL328 was queried by blastN in this assembly, with the region upstream of this locus analysed for promoter insertions.
RNA isolation and RT-qPCR amplification
To analyse the expression of the kanamycin resistance locus in DAB885 and DBL328, three single colonies of each strain were individually picked to 2 ml KB media supplemented with tetracycline (10 µg ml^−1^), grown overnight at 27 °C with shaking and diluted 1:100 the next morning in KB media. Cultures were then grown at 27 °C with shaking for 5 h (until mid-log phase). Cells were spun for 3 min at 3,000 g, the supernatant was removed and pellets were flash frozen in liquid nitrogen. Pellets were randomized and blinded for subsequent RNA isolation and RT-qPCR analysis. Pellets were resuspended by pipetting in 1 ml TRIzol (ThermoFisher Cat. No. 15596018) and held at room temperature for 5 min. Briefly, 200 µl chloroform was added, samples were shaken for 20 s, held at room temperature for 3 min, and then spun at 10,000 g for 18 min. 350 µl of the upper aqueous layer was transferred to a new tube, and 350 µl of 100% ethanol was added and mixed by pipetting. The mixture was then bound to an RNeasy column from a Qiagen RNeasy Mini kit (Cat. No. 74104) by centrifuging at 8,000 g for 30 s. The flow-through was discarded, and the sample was washed once with 700 µl Buffer RW1 from the RNeasy kit by centrifuging at 8,000 g for 30 s, then twice using 500 µl of Buffer RPE by centrifuging at 8,000 g for 30 s and 2 min, respectively, then eluted from the column in 40 µl ultrapure water. cDNA was synthesized from 1 µg of RNA using the ThermoFisher Maxima First Strand cDNA Synthesis Kit (Cat. No. K1671). cDNA was diluted 1:50 in nuclease-free water and used as template for RT-qPCR. A Bio-Rad CFX96 C1000 Touch thermocycler was used for RT-qPCR with the following programme: 95 °C 3 min, (95 °C 10 s, 60 °C 30 s)×40 cycles. RT-qPCR results were analysed using the Bio-Rad CFX Maestro software package. Statistical analysis and graphing were performed using GraphPad Prism. Primers used for kanamycin resistance locus RT-qPCR reaction were F: 5′-GGCTAAAATGAGAATATCACCGG-3′, R: 5′-CTTTAAAAAATCATACAGCTCGCG-3′ [30]. Primers used for tetracycline resistance RT-qPCR reaction were F: 5′-GCGGGATATCGTCCATTCCG-3′, R: 5′-GCGTAGAGGATCCACAGGACG-3′ [31].
Statistics
Data for comparisons within Fig. 3(e) were analysed with a Student’s t-test, assuming equal variances. Data for comparisons within Fig. 3(f) were analysed with a one-way ANOVA, which produced an F=10.17, with groups ‘a’ and ‘b’ statistically significantly found to be different with a P value of 0.05 and a P=0.0042 by Tukey’s post-hoc test. Raw data for each analysis can be found at Figshare [1].
Results
Variability in presence and copy number of IS5 elements throughout P. syringae complete genomes
We used DiamondblastP to search all complete P. syringae (and related species) genomes through https://www.pseudomonas.com/ and to evaluate the presence and copy numbers of annotated InsH protein (the transposase of IS5, encoded within this IS element). Active IS5 elements (with intact InsH) are not universally present throughout all P. syringae genomes, with their presence largely localized to three phylogenetic clades (with stars differentiating these clades highlighted in Fig. 1) in addition to a handful of other scattered strains. Two of these clades are found within phylogroup 3, while one clade is found within phylogroup 1. Copy numbers within each strain are quite variable, but we note that there has been a uniform explosion to >100 copies within two clades comprising two pathovars of P. syringae (pv. aesculi, a pathogen of horse chestnut; pv. lachrymans, a pathogen of cucurbits with these strains largely causative of disease in cucumber). Judging by phylogenetic relationships, and because of the absence of IS5 elements within strains and clades separating P. syringae pv. aesculi and pv. lachrymans, a parsimonious explanation is that these IS elements independently expanded within each of these clades because there are numerous complete genome sequences that lack IS5 elements interspersed between these two clades in the phylogeny.
IS5 elements are found throughout P. syringae strains, with independent expansions across multiple clades. We used Realphy to infer a phylogeny across all complete P. syringae genomes containing the InsH transposase of the IS5 element, while also including relevant representative complete genome sequences to adequately represent genomic diversity in this species. The first column displays either the number of IS5 elements found on the chromosome of each strain (black numbers) or displays whether a genome lacks this element entirely (red circle). The second column (black circle with white number) displays the number of IS5 elements that were localized to plasmids for most strains and to either plasmids or other fragments for P. amygdali pv. aesculi 2250. The third column displays the number of all IS5 elements identified by ISEscan, while the fourth column displays the number of additional IS elements identified by ISEscan. Stars denote clades of interest for IS element expansions.
We also note that IS5 elements are often present on both the chromosome and plasmids of strains, although there are strains where the IS5 element is present solely on the chromosome (P. syringae pv. lachrymans NM002, Pseudomonas savastanoi, P. syringae pv. tomato DC3000, P. syringae CFBP2116) as well as strains where this element is only present on the plasmid (P. syringae R2Leaf, P. syringae CFBP3840, Pseudomonas cannabina ES4326). Additionally, we acknowledge that listed numbers of IS5 elements represent a minimum, as we counted only full-length and active (e.g. annotated) copies, and thus, pseudogenized versions will not be represented in this dataset. However, as an additional step, we searched genomes with no annotated IS5 elements using tblastN with Pla107 InsH as a query and found zero cases where a genome only possessed pseudogenized versions of InsH (data not shown). If hits were present for InsH, they were weak (<50% sequenced identity) or internal fragments within annotated IS elements from alternative families. We therefore believe that the zero counts are an accurate representation of the number of these specific Pla107 IS5 elements present within this dataset.
Variability of IS element copy numbers throughout P. syringae complete genomes
IS element proliferation within a genome is hypothesized to potentially reflect trends in population biology for the lineages of interest [23233]. In this way, an explosion of IS5 elements within a genome might reflect smaller effective population sizes for the lineages and clades highlighted by stars in Fig. 1. Since overall population dynamics should impact copy numbers of all IS element families (rather than just IS5), one clear prediction is that if IS5 element expansion is driven by changes in population biology of the strains then all IS element families should be impacted and should therefore expand in concert within these genomes. To follow up on our initial investigation of the presence of intact insH (and IS5 elements) across P. syringae, we further queried for the presence and copy numbers of all IS element families from complete genomes containing insH as well as additional phylogenetically informative strains. This query also allows independent enumeration of IS5 elements, including those that are too divergent in InsH sequence to have been included in above analyses. Although InsH protein sequences from IS5 elements used in queries above appear to be nearly identical in sequence within and across all genomes where present, with only one or two amino acid substitutions differentiating alleles of insH, it is likely that there are a variety of distinct and highly diverged IS5 elements also found within some genomes. Judging by distinct allelic classes of these IS5 elements, it is highly likely that divergent copies have arisen from a separate horizontal gene transfer event into these strain backgrounds. From the data, there is no clear signal of correlation between IS5 copy number and copy numbers of other IS element families within each of the lineages. Strains in P. syringae pv. lachrymans with large numbers of IS5 elements contain a relatively small number of additional IS element families. Conversely, strains in P. syringae pv. aesculi contain large numbers of both IS5 elements and other additional IS elements. Strains in phylogroup 1 with subtle increases in IS5 copy numbers (between 13 and 30) can contain relatively few (53) to many (234) additional IS element copies. Thus, we do not find a clear general signal that can explain IS5 expansion. This hints that lineage level complexities in population and genome dynamics could be the main drivers of proliferation of IS elements across strains.
Specific proliferation and movement of IS5 elements in three P. syringae pv. lachrymans genomes
To evaluate the variability in the position of IS5 elements across closely related strains, we compared IS5 element distributions from the subclass queried above (represented by InsH sequences) of three P. syringae pv. lachrymans chromosomes. These three strains are closely related according to phylogenetic relationships (Fig. 1) and display Average Nucleotide Identity (ANI) values >99% in all pairwise comparisons (see heatmap in Fig. S1, calculated by FastANI at GTDB [3435]). Although we sought to classify conservation of each IS5 element according to synteny across genomes, genomic variability in regions containing IS5 elements across strains made many of these comparisons challenging. Thus, we split IS5 elements into classes based on distributions across all three genomes and further split each locus within each class into groups (within the presence/absence split) depending on whether synteny was ‘clear’ or ‘unclear’.
As shown in Fig. 2, we found that 49 of the IS elements are found in the same positions across each of the 3 genomes, while 23 additional loci appear to be potentially conserved in position across the 3 strains, but where syntenic relationships are unclear. Given that there are >100 IS5 elements within each strain, a minority of all IS5 elements in each genome are present in conserved locations across these three strains. Further, each strain possessed between 20 and 30 copies of the IS5 element which were present in unique locations across their genomes with an additional ~10 copies per strain where it was likely that the loci were uniquely present. Many of the unique insertions were in regions of the chromosome that were variable mobile elements (predicted to be integrated conjugative elements and phage regions), while numerous others were the product of local tandem duplications of a conserved IS5 element (data not shown).
Diversity in IS5 element insertion sites across three P. amygdali pv. lachrymans. We identified whether IS5 element insertion points (represented by annotated InsH protein sequences) were conserved or divergent across the chromosomes of three strains within pv. lachrymans: M315305 (Pla107), NM002, YM7902. The number outside of parentheses indicates situations where IS5 elements are shared or divergent between strains with high confidence (the clear annotation group), while the number inside parentheses indicates the number of lower confidence instances (the unclear annotation group).
IS5 elements within Pla107 are capable of driving downstream gene expression
IS5 elements have previously been shown to be able to drive gene of downstream genes in other bacterial species. We therefore took advantage of a previously constructed strain to act as a promoter trap to select this ability in IS5 elements from P. syringae. The genome of Pla107 contains a mobilizable megaplasmid that we have previously tagged using the suicide vector pMTN1907. pMTN1907 contains ORFs capable of encoding resistance to both kanamycin (aph3A′) and tetracycline (tetA), and replication of this plasmid within E. coli provides resistance to both of these antibiotics. However, whereas the tetracycline resistance locus is expressed and provides resistance when this vector is incorporated into the genome of Pseudomonas strains, the aph3A′ gene is not expressed in Pseudomonas and therefore strains with pMTN1907 recombined into their chromosomes (or other replicons) remain kanamycin sensitive. We hypothesized that differential functions of the promoter for kanamycin resistance within this plasmid could explain differential resistance phenotypes in E. coli and Pseudomonas strains. Therefore, we selected for kanamycin-resistant mutants of our previously constructed Pla107::pMTN1907 strain (DAB885) with the hopes that we could identify a transposition event of IS5 elements in kanamycin-resistant isolates of this strain. We found that kanamycin resistant mutants occurred quite frequently within this strain background, and sequencing and assembly of one of these revertants demonstrated insertion of an IS5 element upstream of the kanamycin resistance gene within the version of pMTN907 found on the megaplasmid. Specifically, it appears as though a 1,210 bp IS5 family element transposed into a position 62 bp upstream of the start codon for kanamycin resistance in the integrated version of pMTN1907 (Fig. 3a).
IS5 elements can drive downstream gene expression. Strain DAB885 is an isolate of P. amygdali pv. lachrymans M301305, in which pMTN1907 has been recombined into a megaplasmid found in the strain. pMTN1907 codes for an enzyme that provides kanamycin resistance, but which is not expressed in P. syringae. We selected for a kanamycin-resistant version of DAB885 by plating cells onto KB media containing 25 µg ml−1 kanamycin to generate strain DBL328. (a)–(c) We show growth after dilution plating overnight cultures of either P. amygdali pv. lachrymans M3013015, DAB885 and DBL328 onto KB media, KB media containing 10 µg ml−1 tetracycline or KB media containing 25 µg ml−1 kanamycin. (d) Genome sequencing demonstrated that an IS5 element transposed upstream of the kanamycin resistance gene within DBL328 to enable kanamycin resistance. (e) Relative expression of the aph3A′ locus compared to the tetA locus in strains DAB885 and DBL328. A Student’s t-test was used for statistical comparisons across three biological replicates. (f) Relative quantity of mRNA for aph3A′ (KanR) and tetA (TetR) in strains DAB885 and DBL328 at mid-log phase. Data from three biological replicates are shown. Different letters indicate statistical differences between groups by XXXX.
To confirm our prediction that the IS5 element did indeed drive expression in strain DBL328, we performed RT-qPCR using primers anchored in both the tetA locus and aph3A′ in both the previously constructed Pla107::pMTN1907 strain (DAB885) as well as the kanamycin-resistant derivative DBL328. Our data support the hypothesis that the IS5 element can drive gene expression of aph3A′ in this context since the tetA locus is equally upregulated in both backgrounds in mid-log phase, but aph3A′ is only expressed in strain DBL328 (Fig. 3e and f).
Discussion
Ecological and population-level inferences of IS element proliferation
We present data demonstrating that IS5 elements are undergoing ongoing and independent expansion across multiple phylogenetic clades of P. syringae. What is less clear are the evolutionary and ecological forces that enable such expansions within these specific clades. One possibility is that IS5 elements have not expanded within other clades simply because these elements have been introduced to genomes relatively recently and there therefore hasn’t been enough time for expansions to occur. As an argument against this relatively simple explanation, we highlight that clades which have undergone expansion (highlighted by stars in Fig. 1) contain equal or less diversity across strains (are similarly aged or ‘younger’) than another clade with more moderate IS5 element copy numbers as judged by phylogenetic distance.
It is also possible that there are particularities about the ecology of pv. lachrymans and pv. aesculi that enable expansion of IS elements compared to other clades. Although IS element insertions can be beneficial under certain contexts and circumstances [31636], it is thought that these events occur in a minority of cases and that the IS element copy number within a genome is driven by a combination of neutral and deleterious transposition events [333738]. At one end of the spectrum, transposon landing sites are restricted to regions of the genome that are not critical for carrying out cellular functions within a given environment because disruption of these regions is lethal. However, if a transposition event is non-lethal to the cell but either partially lowers fitness of the cell or is neutral, the strength of selection against this particular insertion site at a population level will be driven by population size [39]. At relatively large effective population sizes, selection is a powerful force to cull most deleterious mutations from a population, and genetic drift is comparatively weak, thus limiting the fixation of neutral mutations. However, at relatively small effective population sizes, selection is much weaker and genetic drift much stronger, and thus deleterious and neutral mutations can fix more easily [40]. Thus, it may be that pv. lachrymans strains and pv. aesculi clades possess smaller effective population sizes than other clades due to ecological differences of these strains. Perhaps they undergo smaller or more frequent population bottlenecks during transmission and infection.
To this point, we highlight that both pv. lachrymans and pv. aesculi clades are part of phylogroup 3 of P. syringae. Although many P. syringae strains have been isolated from environmental and water sources, and it is often assumed that their life cycles are intimately tied to transmission by the water cycle, it is notable that relatively few phylogroup 3 strains have been isolated from water sources compared to other phylogroups [1941]. Thus, phylogroup 3 strains may have distinct transmission mechanisms which could have cascading effects on their population dynamics and drive smaller effective population sizes and enable IS5 element proliferation. While likely affected by sampling bias, most of the dramatic cases of IS element and transposon expansions in bacterial genomes thus far are found in obligate symbionts and parasites whose population sizes are drastically bottlenecked during transmission from host to host [4042]. Flipping this idea around, differential IS element proliferation in one bacterial lineage compared to a closely related lineage could therefore be an indicator of a change of relevant evolutionary parameters in populations from one lineage compared to the other. However, we note that our data also suggest that collections of IS element families are not impacted equally across each lineage. Therefore, if changes in population dynamics have enabled expansions of IS5 elements in both pvs. lachrymans and aesculi, the population-level phenomena enabling such expansions have not affected all IS element families similarly, as broad IS element expansion has only occurred in pv. aesculi strains. While our data suggest that strains within pv. aesculi may display different population dynamics than other lineages of P. syringae as reflected in IS element copy numbers, changes in population parameters therefore do not cleanly and generally explain expansion of IS5 elements for pv. lachrymans.
To our knowledge, there have been few previous attempts to characterize IS element proliferation across multiple closely related genomes at both the copy numbers and granularity of genome context reported in this manuscript. Previous reports have typically focused on comparing overall numbers of IS elements between genomes of various distances [23], have identified shared and divergent locations of IS elements across genomes with lower transposon numbers overall [43] or have done so in a manner that emphasizes identification of new insertions [44]. Our results, that IS element positions can differ dramatically across highly related genomes, echo narratives arising from these reports. Moreover, our data highlight that IS elements can rapidly proliferate from a very small (<5) to much higher (>100) copy numbers over relatively short periods of time even within closely related bacterial species and that positions of many of these insertions can dramatically differ between strains.
We acknowledge that our reported results on IS5 element lineage-specific expansion could be explained in part through the existence of additional mechanistic phenomena acting in a highly specific manner. A recent manuscript suggested that IS5 elements could undergo a ‘copy/paste’ mechanism of transposition within a genome rather than the zero-sum ‘cut/paste’ mechanism [45]. Through this action, a single copy of an IS5 element may be able to copy itself and jump to a different genomic location in addition to being maintained at the original insertion site, especially if present on a multi-copy replicon. Such a replication mechanism could facilitate rapid and accelerated expansion of these elements over relatively short evolutionary periods, but it is unclear why this effect would be lineage-specific if IS5 elements are already present. Alternatively, it is possible that there is interference of IS5 elements by other proteins or IS elements in a lineage-specific manner, with expansions therefore explained by absence of inhibition in certain strains or lineages. It is well established that enzymes exist which can limit transposon activity [46] and that chromosome structure and/or protein activity can bias transposition [47]. However, there is limited data supporting this hypothesis at present for this system as there are no known enzymes that directly limit IS5 transposon activity [17]. In the absence of additional future experiments this explanation remains intriguing but unfounded.
Influence of IS element proliferation on evolutionary dynamics
Aside from possible ecological correlates of IS5 element proliferation, what are the possible evolutionary consequences of such expansions? When IS elements proliferate throughout bacterial genomes, they seed that genome with many copies of identical ~1,000 bp nucleotide sequences. These sequences can then act as landing sites for homologous recombination events, including duplications, deletions and inversions [24849]. Therefore, regardless of changes in population dynamics that affect the fixation of mutations, P. syringae lineages where specific IS element families have proliferated to hundreds of copies could therefore be prone to a higher frequency of occurrence of these types of events through time and could be considered to have higher evolutionary plasticity than sister lineages without high levels of IS element proliferation. Indeed, it is worth mentioning that one of the difficulties in identifying synteny between IS5 element insertions is that the regions of the genome they are found in are prone to duplications, inversions and translocations.
Additionally, one of the most intensely investigated aspects of IS5 element biology is the ability of this family of elements to enable the expression of downstream genes and operons, and this has largely been described as occurring in multiple operons in E. coli strains [161836]. To our knowledge, our data represent one of the clearest demonstrations that IS5 elements can lead to upregulation in systems outside of the well-studied E. coli models and could provide useful evolutionary comparison for dissecting the upregulation phenomenon at a systems level. It is currently unknown how many of the ~>100 copy numbers of IS5 elements actually do drive downstream gene expression in P. syringae pv. lachrymans and aesculi genomes, but we expect future studies to better characterize the potential of these elements to change gene expression patterns.
We have further investigated potential influences of IS5 insertions within the P. syringae pv. lachrymans 107 by categorizing genes immediately downstream of all IS5 insertions in this genome. Our assumption in this analysis is that the IS5 element must be positioned upstream of the gene and on the same strand to enable upregulation (the same position as in DBL328 upstream of aph3A′). We acknowledge that this assumption could be wrong and that alternative positioning of IS5 elements could lead to upregulation of proximate genes, but we feel as though such speculation is out of scope for this current manuscript. As one can see in Fig. 4, a majority of the IS5 insertions are located on the opposite strand of DNA as the next potential downstream gene and thus are not currently candidates for upregulation by these elements if our assumption about position holds. The second largest group of genes downstream of IS5 elements is additional copies of IS elements (with both IS5 and ISPsy19 represented). Given the large number of IS elements clustered in this manner, we believe it is likely that these elements may be jumping and proliferating throughout these genomes as compound transposons or at least in a correlated manner. The third largest class of genes downstream of IS5 elements is annotated as ‘hypothetical’ proteins, while the fourth class likely represents genes with a high likelihood of coding for proteins. Although this fourth class is relatively small, we note that it does appear to be overrepresented in DNA-binding elements and those involved in metabolism (Table 1), and it will be interesting to investigate IS element influences on their expression patterns across strains.
Characterization of IS5 element insertion sites and downstream genes. We have characterized genes downstream of all IS5 element insertions in the genome of P. syringae pv. lachrymans 107. The ‘Wrong context’ category represents situations where the IS5 element is on the opposite DNA strand as a potential proximate downstream gene. The ‘Transposase’ category represents situations where the IS5 element is immediately upstream of another potential IS element on the same strand. The ‘Hypothetical’ category represents situations where there is a potential gene downstream of the IS5 element, but this gene is annotated as hypothetical. The ‘Annotated Function’ category represents genes which are downstream of the IS5 element and are on the same strand of DNA and which have annotated functions other than transposase activity. The ‘Pseudo’ category indicates situations where IS5 elements are either upstream of annotated pseudogenes or have clearly disrupted a previously intact locus (upstream and downstream genes of the IS5 element have the same annotation).
Conclusions
Increasing numbers of complete bacterial genomes provide an exceptional opportunity to take account of IS element variation across strains and within bacterial species and here we evaluate the distribution of IS5 elements across the phytopathogen P. syringae. Our report demonstrates that IS5 elements are prevalent within the genomes of a variety of lineages of P. syringae, that some lineages display independent expansions of this element, that these elements are actively transposing throughout these genomes such that a minority of insertion sites are conserved across closely related strains and that this element has the potential to drive downstream gene expression within these strains. Moreover, many of the insertion sites that are unique to specific strains are present within regions of the genome which are differentially present or absent from closely related genomes, and regions of IS5 insertions are often sites that often show inversions and duplications/deletions. At present, it is unclear whether changes in population-level dynamics can explain the differential expansion of IS5 elements within pvs. aesculi and lachrymans strains. It also remains unknown what fraction of the ~100 copies of IS5 elements within some genomes directly upregulate expression within downstream regions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baltrus D Sweten A Conomos T Ponvert N Woodson JD et al IS 5 element characterization across pseudomonas syringae strains 202510.6084/m 9.figshare.27098338 · doi ↗
- 2Siguier P Gourbeyre E Chandler M Bacterial insertion sequences: their genomic impact and diversity FEMS Microbiol Rev 20143886589110.1111/1574-6976.1206724499397 PMC 7190074 · doi ↗ · pubmed ↗
- 3Consuegra J GafféJ Lenski RE HindréT Barrick JE et al Insertion-sequence-mediated mutations both promote and constrain evolvability during a long-term experiment with bacteria Nat Commun 20211298010.1038/s 41467-021-21210-733579917 PMC 7881107 · doi ↗ · pubmed ↗
- 4Vandecraen J Chandler M Aertsen A Van Houdt R The impact of insertion sequences on bacterial genome plasticity and adaptability Crit Rev Microbiol 201743709730 Epub ahead of print November 210.1080/1040841 X.2017.130366128407717 · doi ↗ · pubmed ↗
- 5Siguier P Gourbeyre E Varani A Ton-Hoang B Chandler M Everyman’s guide to bacterial insertion sequences Microbiol Spectr 20153 Epub ahead of print April 210.1128/microbiolspec.MDNA 3-0030-201426104715 · doi ↗ · pubmed ↗
- 6Wagner A Cooperation is fleeting in the world of transposable elements P Lo S Comput Biol 20062 e 16210.1371/journal.pcbi.002016217140282 PMC 1676026 · doi ↗ · pubmed ↗
- 7Casacuberta E González J The impact of transposable elements in environmental adaptation Mol Ecol 2013221503151710.1111/mec.1217023293987 · doi ↗ · pubmed ↗
- 8Plague GR Intergenic transposable elements are not randomly distributed in bacteria Genome Biol Evol 2010258459010.1093/gbe/evq 04020697140 PMC 2942039 · doi ↗ · pubmed ↗