The complete chloroplast genome and phylogenetic analysis of Schisandra henryi C. B. Clarke
Xiuying Yang, Xuefei Qiu, Xianglan Liang, Song Guo, Guoan Shen

TL;DR
This paper reports the full chloroplast genome of Schisandra henryi and its phylogenetic relationships with other species in the Schisandra genus.
Contribution
The study provides the first complete chloroplast genome sequence and phylogenetic analysis for Schisandra henryi.
Findings
The complete chloroplast genome of S. henryi is 146,882 bp long with 39.45% GC content.
124 genes were annotated, including 81 protein coding genes, 35 tRNA genes, and 8 rRNA genes.
Phylogenetic analysis shows S. henryi is most closely related to S. sphenanthera within the Schisandra genus.
Abstract
Schisandra henryi C.B. Clarke belongs to the Schisandraceae family and is known to have important medicinal benefits. The complete chloroplast genome of S. henryi was 146,882 bp in length, with the overall GC content of 39.45%. 124 genes were annotated, which consist of 81 protein coding genes, 35 tRNA genes, and 8 rRNA genes. The phylogenetic tree constructed based on chloroplast genomes indicates that the genus Schisandra is closely related, with S. henryi being most closely related to S. sphenanthera within this group. This study provides further insights into the chloroplast genome database of S. henryi.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1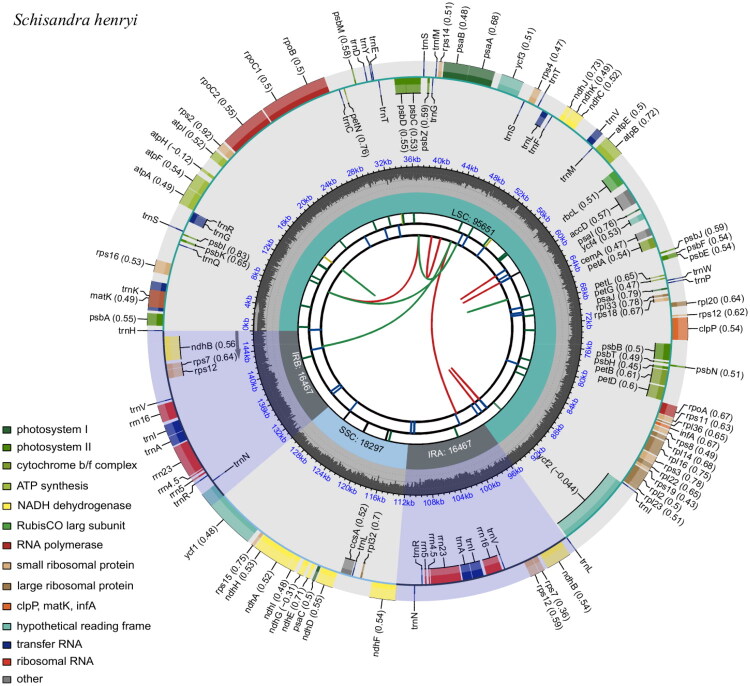 Figure 2
Figure 2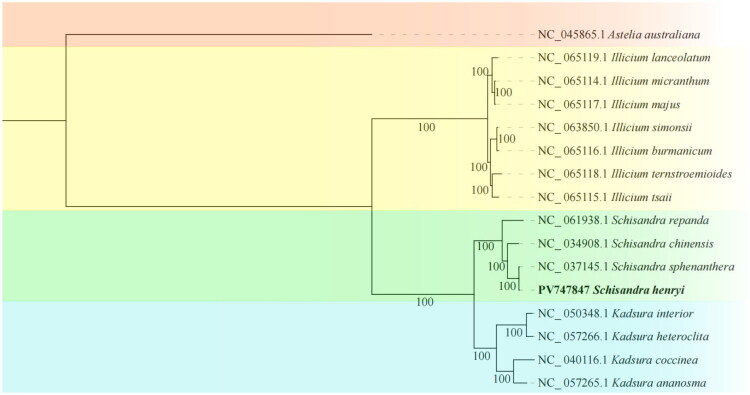 Figure 3
Figure 3- —Guangxi Special Project for Science and Technology Bases and Talents
- —The Scientific Research and Technology Development Plan of Guangxi Laibin, China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-derived Lignans Synthesis and Bioactivity · Medicinal Plants and Bioactive Compounds · Genomics and Phylogenetic Studies
Introduction
Schisandra henryi C.B. Clarke belongs to the Schisandraceae family and is mainly distributed in Yunnan Province, China (Jafernik et al. 2023). In traditional medicine, the roots and stems of S. henryi exhibit wind-damp-dispelling, blood-activating, and analgesic properties, whereas the fruits demonstrate lung-astringing, antitussive, antiperspirant, and essence-stabilizing effects (Ye et al. 2021). It contains various medicinal components, including lignans, polyphenols, triterpenoids, and nortriterpenoids, among others (Jafernik et al. 2023). Schiprolactone A, schisanlactone B, nigranoic acid, and schisandronic acid, which were first isolated from the stems of S. henryi, exhibit certain cytotoxic activity (Chen et al. 2003; Xue et al. 2011). S. henryi extracts show anti-inflammatory and antioxidant activities (Jafernik et al. 2024). Additionally, compounds such as henridilactones E, H, N, and O and schinortriterpenoids display neuroprotective effects and promote neurite outgrowth (He et al. 2020; Jafernik et al. 2024). Studies have shown that extracts from in vitro-cultured microshoots and leaves of parent plants of S. henryi demonstrated significant antiproliferative activity against human cancer cell lines (including Jurkat, MCF-7, HT-29, and HeLa), along with certain antimicrobial activities (Jafernik et al. 2024).
While the medicinal properties of S. henryi have been well studied, research on its genetic and genomic aspects remains scarce. Schisandraceae is a small family of basal angiosperms within the order Austrobaileyales, comprising several genera including Schisandra. The chloroplast (cp) genome, as a highly conserved and informative DNA molecule, provides valuable information for phylogenetic and evolutionary studies and can serve as a ‘super-barcode’ for species identification and germplasm authentication (Wang et al. 2018; Milarska et al. 2023).
Materials and methods
Fresh leaves of one individual were collected from Rongshui Miao Autonomous County, Liuzhou City, Guangxi Zhuang Autonomous Region, China (N23°47′07″, E109°11′46″) and preserved in desiccant. The collection of plant materials (Figure 1) was authorized by Dr. Song Guo ([email protected]). A voucher specimen of S. henryi (voucher number: JXHC031) is preserved at the College of Biology and Food Engineering, Guangxi Science and Technology Normal University, under the supervision of Dr. Song Guo. Species identification was performed by Dr. Song Guo based on morphological characteristics in accordance with Flora of China.
Plant image of S. henryi. S. henryi is a climbing vine with thin, glossy, elliptic-ovate leaves with slightly serrated margins, alternate inflorescences, and a reticulate vein structure. The photo of the species was taken by the authors (Song Guo).
Total genomic DNA was extracted from fresh leaf tissue using the DNeasy Plant Mini Kit (Cat. No. 69104, Qiagen, Hilden, Germany). A DNA library was then created using insert fragments of approximately 300 base pairs in size, followed by paired-end sequencing (2 × 150 bp) on the Illumina HiSeq 2500 platform (San Diego, CA). The quality control of the raw sequencing reads was performed using the fastp v0.2 (Chen et al. 2018), resulting in the production of high-quality filtered reads. Subsequently, the reads were utilized for the de novo assembly of complete plastid genomes, employing the GetOrganelle v1.7.7.1 (Jin et al. 2020). The assembly graph was visualized using Bandage v0.8.1 (Wick et al. 2015). The assembled cp genome was annotated using CPGAVAS2 (Shi et al. 2019). The annotated genome was visualized using CPGView (Liu et al. 2023). Subsequently, cis-splicing and trans-splicing genes were also identified and analyzed based on the annotation results and visualized using the CPGView.
A total of 15 cp genome sequences of species from the family Schisandraceae were retrieved from the NCBI database for phylogenetic analysis, with Astelia australiana NC_045865.1 (Asteliaceae) used as the outgroup. Sequence alignment and phylogenetic tree construction were performed using PhyloSuite v1.2.3 (Zhang et al. 2020). The multiple sequence alignment was conducted with MAFFT (Katoh et al. 2002). A maximum-likelihood phylogenetic tree was then inferred using IQ-TREE (Nguyen et al. 2015) under the GTR + F + I + G4 substitution model. Branch support was assessed with 1000 replicates of ultrafast bootstrap (UF-bootstrap) and the SH-like approximate likelihood ratio test (SH-aLRT) with 1000 replicates. The resulting tree was visualized using the iTOL (Letunic and Bork 2021) web platform.
Results
Approximately, 4.1 Gb of raw data of S. henryi was obtained. The cp genome exhibited a length of 146,882 bp. The coverage depth ranged from a minimum of 115× to a maximum of 24,373× (Figure S1). All gene features, such as cis-splicing (Figure S2) and trans-splicing (Figure S3), as well as circular map (Figure 2), were visualized using CPGView. The cp genome had a typical quadripartite structure, consisting of a large single-copy (95,651 bp), a small single-copy (18,297 bp), and two inverted repeat (IR) regions (16,467 bp). The average guanine–cytosine (GC) content was 39.45%.
Circular visualization of the plastome architecture. The schematic comprises six concentric tracks: (1) innermost repeat elements (direct/palindromic repeats shown as red/green arcs); (2) long tandem repeats (blue bars); (3) microsatellites (color-coded by unit size: p1–p6); (4) structural boundaries (SSC/IR/LSC regions); (5) GC content profile; and (6) outermost gene features (color-coded by function, with transcriptional orientation indicated). Codon usage bias appears parenthetically after gene names. Species identification is noted in the upper left, with functional legend in the lower left.
A total of 124 functional genes were annotated, among which 81 are protein-coding genes (PCGs), 35 were transfer genes (tRNAs), and eight are ribosomal RNA genes (rRNAs). Within the IR regions, there are duplications of three PCGs (ndhB, rps12, rps7), five tRNAs (trnN-GUU, trnR-ACG, trnA-UGC, trnI-GAU, trnV-GAC), and four rRNAs (rrn5, rrn4.5, rrn23, and rrn16). Furthermore, 10 PCGs contained a single intron (rps16, atpF, rpoC1, petB, petD, rpl16, rpl2, ndhB(2x), ndhA), and two genes (ycf3 and clpP) contained two introns (Figure S2). Meanwhile, eight tRNAs contained single intron (trnK-UUU, trnG-GCC, trnL-UAA, trnV-UAC, trnI-GAU(2x), and trnA-UGC (2x)). The cp genome’s trans-splicing gene rps12 (Figure S3) comprised three unique exons, two of which were duplicated due to their localization in the IR regions.
The ML phylogenetic tree constructed from the Schisandraceae family showed that S. henryi and Schisandra sphenanthera were most closely related (Figure 3). The family Schisandraceae comprises only three genera, among which Schisandra and Kadsura are more closely related to each other than to Illicium (Figure 3).
Systematic evolutionary tree of S. henryi. Download 15 chloroplast genome sequences from NCBI and construct a phylogenetic tree using the maximum-likelihood method. The Schisandraceae species are Illicium lanceolatum NC_065119.1, Illicium micranthum NC_065114.1, Illicium majus NC_065117.1, Illicium simonsii NC_063850.1 (Tiantian et al. 2025), Illicium burmanicum NC_065116.1, Illicium ternstroemioides NC_065118.1, Illicium tsaii NC_065115.1, Schisandra repanda NC_061938.1, Schisandra sphenanthera NC_037145.1, Kadsura interior NC_050348.1, Kadsura heteroclita NC_057266.1, Kadsura ananosma NC_057265.1 (Liu et al. 2020), Kadsura coccinea NC_040116.1 (Li and Zheng 2018), Schisandra chinensis NC_034908.1, with Astelia australiana NC_045865.1 (Amor et al. 2020) as the outgroup.
Discussion and conclusions
The complete cp genome of S. henryi exhibited a typical circular quadripartite structure and had a complete size of 146,882 bp, containing a total of 124 genes, with a high average sequencing depth of 397.90×, which ensured high-quality assembly and subsequent analysis. The cp genomes of Schisandra species (Li and Zheng 2018; Wei et al. 2020; Chen et al. 2024; Zhang et al. 2024; Yu et al. 2025). Phylogenetic analysis shows that species within the genus cluster together with high support (bootstrap values of 100%), with S. henryi being most closely related to S. sphenanthera. The cp genome of S. sphenanthera (accession number: NC_037145.1) was reported to be 146,853 bp in length (Wei et al. 2020), which was 29 bp smaller than the cp length of S. henryi. The sequence variation among Schisandra species allows differentiation between species, while the conserved structure and gene content of the cp genome make it a reliable ‘super-barcode’ for species identification and germplasm authentication.
Overall, the complete cp genome of S. henryi enriches the genomic resources available for the genus and provides valuable data for future comparative, phylogenetic, and evolutionary studies in Schisandraceae.
Supplementary Material
Supplemental Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amor MD, Holmes GD, James EA. 2020. Characterization of the complete plastid genome of Astelia australiana (JH Willis) LB Moore (Asteliaceae, Asparagales). Mitochondrial DNA B Resour. 5(1):656–657. 10.1080/23802359.2019.171123333366690 PMC 7748721 · doi ↗ · pubmed ↗
- 2Chen H et al. 2024. The complete chloroplast genome and phylogenetic analysis of a rare species Schisandra macrocarpa (Schisandraceae). Mitochondrial DNA B Resour. 9(12):1693–1697. 10.1080/23802359.2024.243709939687441 PMC 11648132 · doi ↗ · pubmed ↗
- 3Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. 10.1093/bioinformatics/bty 56030423086 PMC 6129281 · doi ↗ · pubmed ↗
- 4Chen Y-G et al. 2003. Triterpenoids from Schisandra henryi with cytotoxic effect on leukemia and He La cells in vitro. Arch Pharm Res. 26(11):912–916. 10.1007/bf 0298019914661856 · doi ↗ · pubmed ↗
- 5He T-B et al. 2020. Neuroprotective schinortriterpenoids with diverse scaffolds from Schisandra henryi. Bioorg Chem. 105:104353. 10.1016/j.bioorg.2020.10435333096311 · doi ↗ · pubmed ↗
- 6Jafernik K et al. 2024. Comparative assessment of lignan profiling and biological activities of Schisandra henryi leaf and in vitro Plant Form bioreactor-grown culture extracts. Pharmaceuticals. 17(4):442. 10.3390/ph 1704044238675405 PMC 11053505 · doi ↗ · pubmed ↗
- 7Jafernik K, Ekiert H, Szopa A. 2023. Schisandra henryi—a rare species with high medicinal potential. Molecules. 28(11):4333. 10.3390/molecules 2811433337298808 PMC 10254417 · doi ↗ · pubmed ↗
- 8Jafernik K, Motyka S, Calina D, Sharifi-Rad J, Szopa A. 2024. Comprehensive review of dibenzocyclooctadiene lignans from the Schisandra genus: anticancer potential, mechanistic insights and future prospects in oncology. Chin Med. 19(1):17. 10.1186/s 13020-024-00879-038267965 PMC 10809469 · doi ↗ · pubmed ↗