Developmental dynamics of cellular specialization during proanthocyanidin accumulation in persimmon fruit
Yosuke Fujiwara, Soichiro Nishiyama, Akane Kusumi, Keiko Okamoto-Furuta, Hisayo Yamane, Keizo Yonemori, Ryutaro Tao

TL;DR
The paper explores how persimmon fruit cells specialize to store large amounts of proanthocyanidins, using gene activity and structural analysis.
Contribution
The study reveals a novel intercellular sequestration mechanism for proanthocyanidin accumulation in persimmon fruit.
Findings
Spatial gene expression patterns are linked to proanthocyanidin accumulation.
Ultrastructural analysis shows unique cell specialization during fruit development.
Abstract
Spatial gene expression and 2D/3D ultrastructural analyses reveal a unique intercellular sequestration mechanism underlying massive proanthocyanidin accumulation in persimmon fruit.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
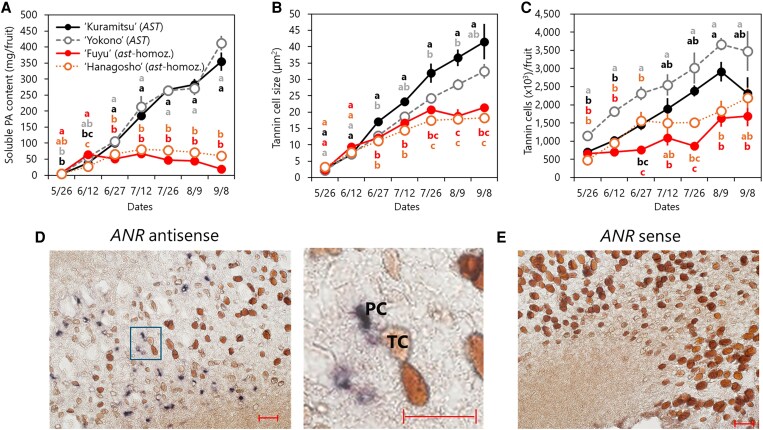 Figure 1
Figure 1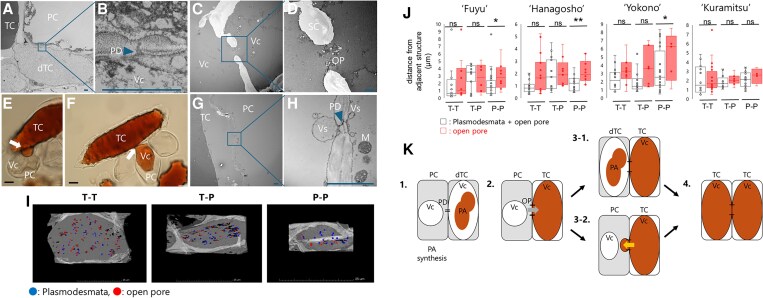 Figure 2
Figure 2- —Japan Society for the Promotion of Science KAKENHI
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Postharvest Quality and Shelf Life Management · Plant Molecular Biology Research
Dear Editor,
Persimmon (Diospyros kaki) fruit accumulates exceptionally large amounts of proanthocyanidins (PAs). Because PAs are highly reactive and potentially toxic to the cell, they are often sequestered into compartmentalized structures (Dixon and Sarnala 2020; Lu et al. 2022). In persimmon, PAs are stored in vacuoles of specialized “tannin cells” (TCs), yet the developmental origin and regulation of these cells have remained unclear. Here, we propose an intercellular partitioning mechanism of PA synthesis and sequestration as a basis for massive PA accumulation.
PAs, common secondary metabolites in land plants, are flavan-3-ol polymers that accumulate in vacuoles and confer astringency while providing antioxidant benefits (Dixon et al. 2005; Aron and Kennedy 2008). Although their biosynthesis pathway is well characterized, intracellular trafficking to the vacuole and polymerization remain incompletely resolved (Zhao 2015; Dixon and Sarnala 2020). Oriental persimmon shows extreme PA accumulation, with sequestration in idioblast TCs dispersed in the parenchyma and often clustered. TCs were documented more than a century ago, and their size is known to correlate with fruit PA content (Howard 1906; Tokugawa and Yuasa 1936; Yonemori and Matsushima 1985, 1987; Ikegami et al. 2004; Hamada et al. 2009; Fig. 1A and B). A distinctive feature is the presence of large open pores (OPs) in cell walls (Yonemori and Matsushima 1987). Astringency strongly affects persimmon's market value; pollination-constant and nonastringent (PCNA) mutants (ie ‘Fuyu’ and ‘Hanagosho’) cease PA accumulation early to produce nonastringent fruit (Fig. 1A) and are widely used in cultivation and modern breeding. PCNA is conferred by a recessive allele at the ASTRINGENCY (AST) locus (Ikeda et al. 1985; Yamada and Sato 2002; Nishiyama et al. 2018), but the causal gene remains unknown.
Here, using an enzymatic and hydraulic workflow (Supplementary Figure S1), we quantified TCs per fruit and observed a steady increase toward the onset of ripening in all cultivars (Fig. 1C). We also observed a positive association between the TC number and the soluble PA content in non-PCNA (‘Kuramitsu’ and ‘Yokono’) and in the relatively PA-rich PCNA cultivar ‘Hanagosho’ (Supplementary Figure S2). These patterns suggest that storage capacity expands via continued TC differentiation during the PA-accumulation window.
In situ hybridization revealed that anthocyanidin reductase (ANR), a key enzyme that supplies the PA extension unit epigallocatechin in persimmon fruit (Akagi et al. 2009), was expressed in a scattered pattern across the mesocarp and was frequently strongest in parenchyma cells (PCs) directly adjacent to TCs (Fig. 1d and e; Supplementary Figure S3). This pattern indicates functional partitioning: PCs synthesize PA components, while TCs specialize in sequestration. This contrasts with systems where biosynthesis and storage colocalize, such as the Arabidopsis seed coat, in which PAs accumulate in the inner integument and BANYULS (ANR homolog) is expressed in those layers (Debeaujon et al. 2003). Similar colocalization can be seen in different organs across species (Tsai et al. 2006; Abeynayake et al. 2011; Westley et al. 2024). In Kalanchoe and Acorus, chalcone synthase is restricted to PA-containing idioblasts (Karwatzki et al. 1993), unlike the adjacent parenchyma expression seen here. These examples highlight a distinctive sequestration system in persimmon.
Microscopic observation of OPs in persimmon is challenging: 2D imaging is sensitive to sectioning geometry (ie sectioning must be performed precisely at their location), and ultrathin sectioning is extremely difficult because TCs and PCs differ in physical properties after fixation. We therefore used focused ion beam–scanning electron microscopy (FIB-SEM) to reconstruct interfaces in 3D (Supplementary Videos S1 and S2), complemented by transmission electron microscopy (TEM) to resolve ultrastructure. Methods are provided in the Supplementary Material (Supplementary Table S1).
We observed plasmodesmata (PD) clustered at interfaces between developing TCs and PCs, with the TC vacuole attached to the wall (Fig. 2A and B). Organelles, such as the Golgi apparatus, endoplasmic reticulum (ER), mitochondria, and plastids, are enriched at these interfaces (Supplementary Figure S4). ER abundance varied among TCs (Supplementary Figure S5), consistent with the absence of ANR in mature TCs (Fig. 1D) and suggesting that ER-poor cells are storage-oriented. At later stages, OPs interrupting the wall were frequently observed, and vacuoles of adjacent cells were often continuous through OPs (Fig. 2C to F). Vesicle-like structures were frequently associated with PD at TC–PC interfaces (Fig. 2G and H). FIB-SEM enabled OP quantification and showed that OPs were most frequent at TC–TC interfaces and least at PC–PC interfaces, whereas total PD + OP counts were comparable among interface types (Fig. 2I). Nearest-neighbor distances further supported OP enrichment at TC cell walls, since OP spacing was significantly larger in PC–PC interfaces than in interfaces that included TCs in most cultivars tested (Fig. 2J). All these features were observed in both non-PCNA and PCNA (Supplementary Figure S6).
We propose two complementary routes for TC expansion active during PA accumulation (Fig. 2K). In one route, PCs adjacent to TCs differentiate into new TCs, consistent with the ANR spatial pattern. In the other, TC protrusions penetrate neighboring cells and fuse vacuoles through OPs, yielding clustered TCs with vacuolar continuity. Meanwhile, some ANR signals are isolated from existing TCs (Fig. 1D), indicating a de novo path that does not require direct contact. The presence of OPs and interface features in PCNA fruit (Fig. 2J; Supplementary Figure S6) indicates that the anatomical “containers” for massive PA storage can form even when biosynthesis is curtailed, placing the AST genes more plausibly on biosynthesis or intracellular transport rather than intercellular sequestration.
Several mechanistic questions follow. First, if biosynthesis is localized, TCs may emit positional cues that induce biosynthesis in adjacent cells; this remains untested. Second, the origin of OPs is unclear; some observations suggest physical rupture, whereas the frequent association of vesicle-like structures and ER at PD points to regulated wall remodeling. Third, isolated ANR expression implies an additional initiation path. Addressing these points will clarify how the storage cells are specified and expanded.
We propose that specialized intercellular sequestration enables persimmon to accumulate PAs massively while minimizing cytosolic exposure. This perspective reframes metabolite control as a problem of interface biology as well as pathway flux. The framework suggests targets for breeding or engineering. We anticipate similar strategies may apply to other secondary metabolites that require compartmentalization for plant fitness and product quality.
Supplementary Material
kiaf645_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abeynayake SW, Panter S, Mouradov A, Spangenberg G. A high-resolution method for the localization of proanthocyanidins in plant tissues. Plant Methods. 2011:7:1–6. 10.1186/1746-4811-7-13.21595992 PMC 3117829 · doi ↗ · pubmed ↗
- 2Akagi T et al Expression balances of structural genes in shikimate and flavonoid biosynthesis cause a difference in proanthocyanidin accumulation in persimmon (Diospyros kaki Thunb.) fruit. Planta. 2009:230:899–915. 10.1007/s 00425-009-0991-6.19669159 · doi ↗ · pubmed ↗
- 3Aron PM, Kennedy JA. Flavan-3-ols: nature, occurrence and biological activity. Mol Nutr Food Res. 2008:52:79–104. 10.1002/mnfr.200700137.18081206 · doi ↗ · pubmed ↗
- 4Debeaujon I et al Proanthocyanidin-accumulating cells in Arabidopsis testa: regulation of differentiation and role in seed development. Plant Cell. 2003:15:2514–2531. 10.1105/tpc.014043.14555692 PMC 280558 · doi ↗ · pubmed ↗
- 5Dixon RA, Sarnala S. Proanthocyanidin biosynthesis—a matter of protection. Plant Physiol. 2020:184:579–591. 10.1104/pp.20.00973.32817234 PMC 7536678 · doi ↗ · pubmed ↗
- 6Dixon RA, Xie DY, Sharma SB. Proanthocyanidins—a final frontier in flavonoid research? New Phytol. 2005:165:9–28. 10.1111/j.1469-8137.2004.01217.x.15720617 · doi ↗ · pubmed ↗
- 7Hamada K, Hasegawa K, Ogata T. The growth of tannin cells from before flowering to fruit maturity and astringency in Japanese persimmons. J Hortic Sci Biotechnol. 2009:84:72–76. 10.1080/14620316.2009.11512482. · doi ↗
- 8Howard BJ . Tannin cells of persimmons. Bull Torrey Bot Club. 1906:33:567. 10.2307/2478934. · doi ↗