Metabolic alteration in oxylipins and endocannabinoids point to an important role for soluble epoxide hydrolase and inflammation in Alzheimer’s disease—finding from Alzheimer’s Disease Neuroimaging Initiative
Kamil Borkowski, Chunyuan Yin, Alida Kindt, Nuanyi Liang, Elizabeth de Lange, Colette Blach, John W. Newman, Rima Kaddurah-Daouk, Thomas Hankemeier

TL;DR
The study finds that inflammation-related metabolic changes, especially involving soluble epoxide hydrolase, are linked to Alzheimer's disease progression and cognitive decline.
Contribution
The study provides new insights into inflammation pathways in Alzheimer's disease, highlighting sex and genetic differences in metabolic profiles.
Findings
Disruption in inflammation pathways and sEH dysregulation is confirmed in Alzheimer's disease with sex and disease stage differences.
Metabolic markers of disease progression and cognitive resilience are identified, emphasizing bile acids, lipid peroxidation, and cortisol.
Findings suggest therapeutic approaches targeting inflammation tailored to subgroups based on sex, APOE genotype, and metabolic profiles.
Abstract
Mounting evidence implicates inflammation as a key factor in Alzheimer’s disease (AD) development. We previously identified pro-inflammatory soluble epoxide hydrolase (sEH) metabolites to be elevated in plasma and CSF of AD participants and to be associated with lower cognition in non-AD subjects. Soluble epoxide hydrolase is a key enzyme converting anti-inflammatory epoxy fatty acids to pro-inflammatory diols, reported to be elevated in multiple cardiometabolic disorders. Here we analyzed over 700 fasting plasma samples from the baseline of Alzheimer’s Disease Neuroimaging Initiative (ADNI) 2/GO study. We applied targeted mass spectrometry method to provide absolute quantifications of over 150 metabolites from oxylipin and endocannabinoids pathway, interrogating the role for inflammation/immune dysregulation and the key enzyme soluble epoxide hydrolase in AD. We provide further…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
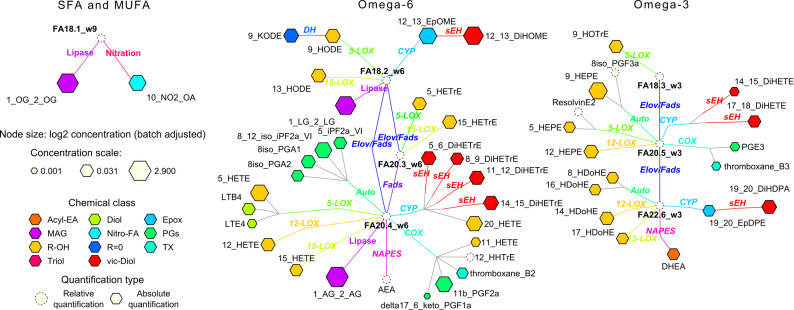 Figure 1
Figure 1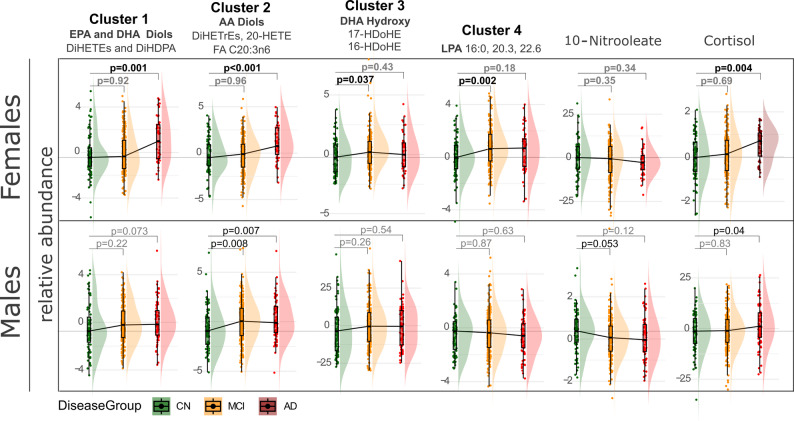 Figure 2
Figure 2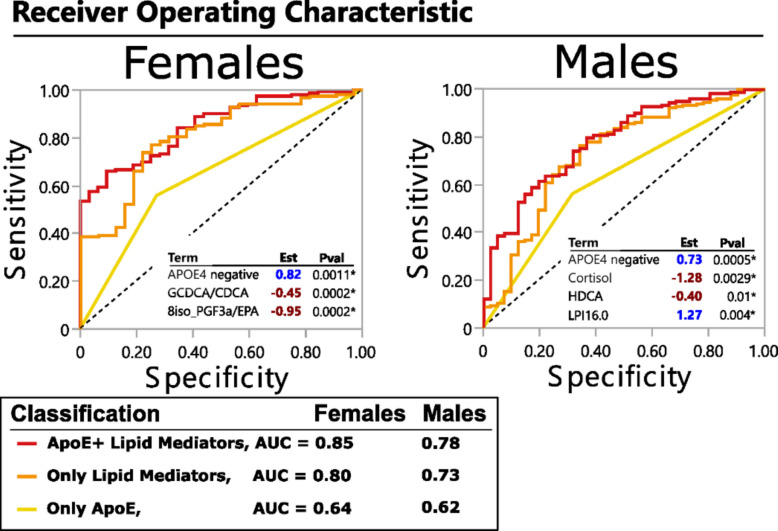 Figure 3
Figure 3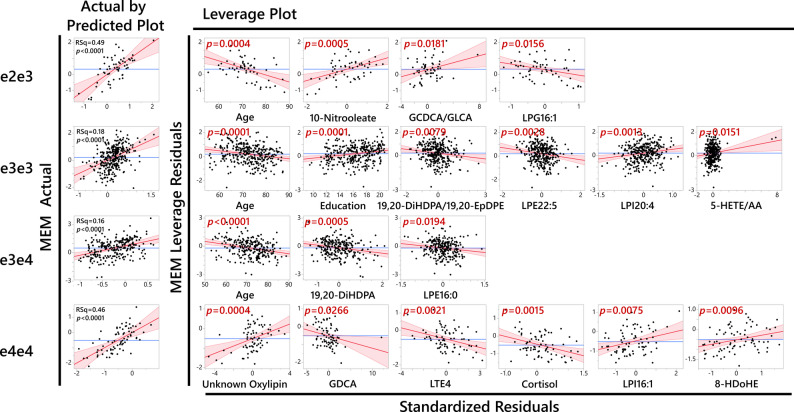 Figure 4
Figure 4- —https://doi.org/10.13039/501100004543China Scholarship Council
- —https://doi.org/10.13039/100000049National Institute on Aging
- —National institute of aging
- —https://doi.org/10.13039/100000009Foundation for the National Institutes of Health
- —https://doi.org/10.13039/501100003246Nederlandse Organisatie voor Wetenschappelijk Onderzoek
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEicosanoids and Hypertension Pharmacology · Fatty Acid Research and Health · Cannabis and Cannabinoid Research
Introduction
Inflammation is recognized as an important contributor to Alzheimer’s disease (AD) pathology and cognitive decline and as a crucial risk factor for AD development that exacerbates disease progression [1, 2].Oxylipin and endocannabinoid (OxyL-EC) pathways are key regulators of inflammation, with changes in their levels being a hallmark of inflammation-related disorders [3–13]. The oxylipins including fatty acid alcohols, diols, epoxides, ketones, and prostanoids are derived from multiple polyunsaturated fatty acids (PUFA) by the action of cyclooxygenases (COX), lipoxygenases (LOX), cytochrome P450 (CYP), soluble epoxide hydrolase (sEH) or reactive oxygen species (ROS) and various downstream enzymatic processes [14]. Circulating endocannabinoids are produced either by acylation and release of acyl ethanolamides from phosphatidylethanolamine and further degraded by fatty acid amide hydrolase (FAAH), or as a product of glycerol-lipid metabolism like monoacylglycerols (MAGL) [15].
Increasing evidence has demonstrated OxyL-EC dysregulation in AD participants. Disturbances at the protein expression and metabolome level of these pathways are observed in both the periphery [16, 17] and central nervous system [18–20] in humans. In rodent models of AD, AD-related pathologies can be altered by manipulating OxyL-EC related enzymes, including 5-LOX [21, 22], 12/15-LOX [23–25], COX [26, 27], sEH [28, 29], CYP [30, 31], FAAH [32], MAGL [33], and their combination [34]. Moreover, genetic polymorphisms in many of these pathways have been linked to AD risk, supporting causative effects from these pathways [35–37].
The Alzheimer’s Disease Metabolomics Consortium (ADMC) has shown that peripheral metabolic changes informed about cognitive changes, brain imaging changes, and amyloid plaques, tau tangles, and neurodegeneration (ATN) markers for disease [38–41]. We have recently generated a comprehensive profile of OxyL-EC pathways in subsets of well-characterized AD cohorts, including the Religious Orders Study and the Rush Memory and Aging Project (ROS-MAP) [42] and the Emory cohort (the Emory Healthy Brain Study, Cognitive Neurology Research, and Memory) [43]. We have used a state-of-the-art validated, quantitative, targeted mass spectrometry platform, providing absolute quantification of ~ 150 OxyL-EC, covering multiple metabolic pathways and possible substrates [44, 45]. This pioneer work identified metabolites of CYP/sEH and acyl ethanolamides in plasma and cerebrospinal fluid (CSF) to be associated with AD pathology, and plasma sEH metabolites to be associated with perceptual speed in cognitively normal and mild cognitively impaired subjects. Furthermore, utilizing the metabolomic-proteomic integration approach, we have shown association of the sEH pathway with CSF proteins related to glycolysis, vascular inflammation and neuronal outgrowth [46], with changes in CSF levels of those proteins being reflective of AD-related changes in the brain [47].
In the current manuscript we seek to confirm and expand on our previous findings of the dysregulation in OxyL-EC pathways in AD, utilizing 763 subjects from the Alzheimer’s Disease Neuroimaging Initiative (ADNI) 2/GO cohort. The ADNI cohort enables us to further investigate the dysregulation of OxyL-EC pathways at different disease stages, ranging from cognitively normal (CN), mild cognitively impaired (MCI) to AD, and the association of baseline OxyL-EC levels with future cognitive decline. Additionally, to uncover disease heterogeneity, we investigate the influence of sex and APOE genotype on the OxyL-EC interaction with AD-related outcomes.
Methods
Study participants
The Alzheimer's Disease Neuroimaging Initiative (ADNI) study recruited 763 individuals over the age of 70 years into longitudinal study. At baseline (the time when participants enrolled in the project), this comprised 178 cognitively normal controls (CN), 445 with mild cognitive impairment (MCI; 290 early-stage and 155 late-stage MCI) and 136 with Alzheimer’s Disease (AD). The diagnostic evaluation criteria of ADNI participants, including CN, early/late-MCI and AD were detailed in the ADNI documentation website (https://adni.loni.usc.edu/data-samples/adni-data/study-cohort-information/). In brief, the criteria taking into account evaluation of Mini-Mental State Exam (MMSE) scores, Clinical Dementia Rating scale (CDR), the presence of memory complaint, the presence of cognition and functional performance impairment (measured via the education-adjusted Wechsler Memory Scale-Revised Logical Memory II subscale), and whether the participants met the National Institute of Neurological and Communicative Disorders and Stroke/the Alzheimer’s Disease and Related Disorders Association criteria for probable AD. Additional exclusion criteria, such as the presence of major depression and other neurological disease, also applied and detailed in the abovementioned ADNI documentation website. Originally in ADNI 2/GO, early/late-MCI was differentiated by the cut-offs values of education-adjusted Wechsler Memory Scale-Revised Logical Memory II subscale, but these 2 groups are no longer differentiated here following the new ADNI 3 guidelines and therefore the 2 groups are combined. Baseline clinical diagnoses (CN, MCI, AD) were assigned using standardized neuropsychological assessments and clinical consensus; fluid (CSF/plasma) or PET biomarkers were not used to establish these diagnoses. The early-stage and late-stage MCI participants were grouped together as the distinction between these two groups is minimal. Follow-up cognition tests were performed at 1, 2 and 4 years after baseline. We analyzed 763 plasma samples from baseline, from which 23 non-fasting participants were excluded. The non-fasting participants were present in each diagnosis group. Additional participant group classification was based on the experienced diagnostic change, i.e., progression from CN to MCI or AD, or from MCI to AD within 1, 2 or 4 years after baseline. “Stable CN” comprises CN baseline samples that did not convert to either MCI or AD within 4 years follow up, and “stable MCI” comprises MCI baseline participants that did not convert to AD within 2 years follow up. Participants whose diagnosis reverted, i.e., changed from AD to MCI or CN, or from MCI to CN were excluded (n = 39). See Table 1 and Table 2 for more details. Some key biometrics stratified by sex are presented in the Supplemental Table S1. The differences between cognitive groups in the levels of key biometrics presented in Table 1 were assessed using ANOVA. All cognitive measures were different in each group with the following distribution: stable CN > Stable MCI > MCI converter > AD. For CSF biomarkers, the Ttau/AB42 was different in each group (CN < Stable MCI < MCI converter < AD). PTAu181 showed the following distribution: CN < Stable MCI < (MCI converter = AD).Table 1. Demographics and clinical data of studied ADNI subjects at baselineGroupStable CNStable MCIMCI converterADSample number (n)15831381133Male:Females48%: 52%54%: 46%55%: 45%59%:41%Age, mean ± SD72.3 ± 6.0871.3 ± 7.4771.8 ± 7.1273.7 ± 8.13BMI, mean ± SD27.3 ± 4.6227.8 ± 5.1127.4 ± 5.4326.4 ± 5.50Education y, mean ± SD16.8 ± 2.5016.0 ± 2.6516.0 ± 2.6615.8 ± 2.73APOE genotype %APOE4 (e4 pos/e4 neg)28%: 72%41%: 59%74%: 26%65%:35%E2E2 (n = 3)0.6%0.3%00.8%E2E3 (n = 60)12.3%8.3%3.7%3.8%E3E3 (n = 334)58%50.5%22.2%30.1%E2E4 (n = 12)0.6%1.9%2.5%1.5%E3E4 (n = 254)24.7%30.4%50.6%43.6%E4E4 (n = 77)3.8%8.6%21%20.2%Cognitive biomarkersMOCA, mean ± SD25.8 ± 2.3823.6 ± 3.1421.5 ± 2.6517.3 ± 4.55ADNI MEM, mean ± SD1.1 ± 0.600.5 ± 0.66-0.2 ± 0.54-0.9 ± 0.53ADNI EF, mean ± SD1.0 ± 0.840.4 ± 0.84-0.08 ± 0.85-0.9 ± 0.94ADNI LAN, mean ± SD0.9 ± 0.710.4 ± 0.75-0.06 ± 0.69-0.8 ± 0.97ADNI VS, mean ± SD0.3 ± 0.560.0 ± 0.70-0.2 ± 0.78-0.6 ± 0.95ADAS13, mean ± SD8.7 ± 4.5613.8 ± 6.1021.1 ± 6.3031.0 ± 8.51MMSE, mean ± SD29.1 ± 1.2228.2 ± 1.6627.1 ± 1.6723.1 ± 2.03CSF biomarkersTtau/AB420.21 ± 0.160.3 ± 0.230.56 ± 0.290.63 ± 0.32pTau (181)21.1 ± 8.924.3 ± 12.635.3 ± 15.936.9 ± 15.9Abbreviations: CN Cognitive Normal, MCI Mild Cognitive Impairment, SD Standard Deviation, MOCA Montreal Cognitive Assessment, ADNI Alzheimer’s Disease Neuroimaging Initiative, MEM Memory, EF Executive Functioning, LAN Language, VS Visuospatial Functioning, ADAS13 Alzheimer’s Disease Assessment Scale 13, MMSE Mini-Mental State Examination. Stable CN represents a subset of CN group that does not change cognitive status over 4 years follow upTable 2Diagnoses for subjects with available CSF measurements in each visitBaseline CNBaseline MCIBaseline ADBaseline178 CN429 MCI133 AD2 years137 CN, 13 MCI, 2 AD, 26 NA19 CN, 266 MCI, 60 AD, 84 NA1 MCI, 28 AD, 104 NA4 years94 CN, 11 MCI, 2 AD, 71 NA27 CN, 156 MCI, 48 AD, 198 NA133 NAAbbreviations: AD Alzheimer’s disease, CN Cognitive Normal, MCI Mild Cognitive Impairment, NA cognitive test result not available
Metabolomics profiling
All baseline plasma samples (n = 763) were assessed using a targeted ultra-high pressure liquid chromatography (UHPLC)-MS/MS lipidomic profiling strategy as previously described [44]. Briefly, liquid–liquid extraction with 1:1 methyl tertbutyl ether/n-butanol was performed on plasma samples after including isotopically labeled internal standards and targets were measured using reversed-phase liquid chromatography-mass spectrometry (RPLC-MS/MS) under high and low pH conditions after solvent exchange. Specific UHPLC systems and columns were employed for optimal separation and detection and data preprocessing was performed with vendor-specific software to ensure accurate peak integration and internal standard matching. Briefly, for the high pH measurements a SCIEX 6500 + QTRAP mass spectrometer was used with a Kinetex EVO column by Phenomenex followed by analyst data acquisition software (Version V1.7.2, AB Sciex). The triple quadrupole mass spectrometer operated in polarity switching mode and all analytes were monitored in dynamic Multiple Reaction Monitoring (dMRM) mode. While for the low pH analysis, a Sciex 7500 QTRAP mass spectrometer was used with an Acquity UPLC BEH C18 column (Waters) followed by Sciex OS Software V2.0.0.45330 (AB Sciex). This platform enables analyses of 260 metabolites covering oxylipins and their free fatty acid precursors, lysophospholipids, sphingoid bases, endocannabinoids and bile acids. All samples were processed with rigorous quality control measures including case/control randomization over all batches, inclusion of solvent blanks, pooled sample quality control aliquots, and National Institute of Standards and Technology (NIST) Standard Reference Material 1950 – Metabolites in Human Plasma (Merck, Darmstadt, Germany). These batches were re-randomized for measurements where method blanks, reference and solvent calibration samples were included at regular intervals. The majority of analytes were quantified against analytical standards, where area counts were recorded, adjusted for deuterated surrogate responses and then calculated into nanomolar concentrations using solvent calibration lines from which the lowest calibration point was subtracted from other calibration points. Next, the calibration lines were checked for outliers which were identified using an outlier test, where only calibration points at the ends were allowed to be removed. A calibration point within the calibration line was removed if the ratio was lower than its previous (lower) calibration point. After these corrections, the model was recalculated. Reported monoacylglycerols (MAGs) are the sum of 1- and 2-acyl isomers and 10-Nitrooleate is the sum of 9-Nitrooleate and 10-Nitrooleate, due to isomerization during sample processing. Fatty acids, arachidonyl ethanolamide (AEA) and 12-HHTrE were reported as ratios of their peak area to the peak area of corresponding surrogate.
Amyloid and Tau measurement in CSF: The levels of Amyloid beta 1–42 (Aβ42) peptide and tau proteins in CSF samples were quantified using a Roche Elecsys immunoassay. The measurement processes were fully automated and the sample preparation procedure involving 2 incubation steps were detailed in previous publications [48, 49]. The cut-off value of Tau/Aβ42 ratio of 0.24 was derived from previous studies using Gaussian mixture modeling to define asymptomatic AD with the presence of AD-related biomarkers [50, 51].
Data preprocessing
After peak integration an in-house quality assessment tool was used to perform correction of target compounds with internal standards to obtain relative ratios, batch correction, and filtering of targets with low signal to noise ratios. To obtain absolute concentrations, linear regression models were calculated per batch between known concentrations and relative ratios for each target. The ratio of the lowest calibration point (CAL0) was subtracted from all ratios in the calibration line. Baseline data with detectable coverage of over 70% were further analyzed. All data were log2-transformed. The following data pre-processing procedure was performed in JMP® Pro 17.2.0 (SAS Institute Inc, Cary NC): outliers were detected and removed using the Robust Fit Outliers method and missing values were imputed with the Multivariate Normal Imputation method [52]. Ratios of metabolites were calculated based on the base-2 exponential-transformed data after imputation, followed by log2-transformation before further analysis. Non-fasted participants (n = 23) and participants of Significant Memory Concern (SMC) were excluded from further analysis. Medication adjustments were made based on the stepwise AIC-backward method previously described [53, 54].
Statistical analysis
Data reduction
To reduce the dimensionality and collinearity of the data, gain additional information about associations among variables and to facilitate result interpretation variables showing significant associations with the outcomes (Determined by the analysis of variance (ANOVA)) were converted into cluster components. Variables were clustered using the Jmp Pro implementation of the VARCLUS Procedure, a principal components analysis for variable grouping and cluster component (the linear combination of all variables in each cluster) calculation. This is a data-driven approach where the algorithm defines the optimal number of clusters, with resulting clusters composed of highly correlated variables. Those correlations in terms of metabolomic data mostly align with biological pathways, further confirming integrity of the data.
Comparison of lipid mediator levels between diagnosis groups
For lipid mediator levels, ANOVA followed by Dunnett’s post hoc test was performed to compare means between CN and other diagnosis groups, stratified by sex. Lipid mediators exhibiting significant differences between groups were grouped into clusters as described above. Cluster components were used in ANOVA analysis with Dunnett post-test, comparing the difference in lipid mediator levels between CN and other diagnosis groups. Supplemental Table S2 and Supplemental Table S3 describe cluster members as well as their correlations within the cluster, least square means (standard error range) and ANOVA p-values. ANOVA p-values for individual metabolites are presented in the Supplemental Table S4. Presence of the APOE 4 allele, sex, body mass index (BMI), age and years of education were included as covariates in all analyses. This analysis was performed in R studio software (Version 4.3.1). Figures were created in R, cytoscape 3.10.2. Multiple comparison control was accomplished with the false discovery rate (FDR) correction method of Benjamini and Hochberg [55].
MCI conversion and memory (ADNI MEM) predictive minimal models
Using JMP 17.0 pro (JMP, SAS institute, Cary, NC), we applied a combination of bootstrap forest and stepwise linear (for ADNI MEM) and logistic (for MCI converters vs stable MCI) regression modeling, with Bayesian Information Criterion (BIC) cutoff. Variable selection by bootstrap forest minimized outlier effects. Variables most frequently appearing in the models were identified by bootstrap forest (logistic or regression, respectively): trees in forest = 100; terms sampled per split = 5; bootstrap sample rate = 1. A variable contribution scree plot was generated using variable rank and the likelihood ratio of chi-square (for categorical fasted/non-fasted prediction) or sum of squares (for continues cognitive scores). The scree plot was used to determine a likelihood ratio of chi-square or sum of squares cutoff value for variables contributing to the model. Selected variables were then subjected to forward stepwise logistic or linear regression modelling. Stepwise regression was used to highlight independent predictors of outcomes, to create the minimal model.
Results
In the current study we have analyzed plasma from the 763 participants from the ADNI 2/GO, providing absolute quantification of 45 eicosanoids, 5 endocannabinoids, 49 lysophospholipids, 13 monounsaturated and polyunsaturated fatty acid (MUFA&PUFA), 4 sphingolipids, and 1 steroids lipids (Fig. 1 and Supplemental Table S3). In addition, our data included 18 bile acids (Supplemental Figure S1). The bile acid levels in ADNI cohort were previously reported [40] and are used in the current analysis only to show their interaction with lipid mediators.Fig. 1. Quantified oxylipins, endocannabinoids and polyunsaturated fatty acids in plasma samples of ADNI 2/GO participants, projected onto their metabolic pathway. Nod size indicates the average concentration observed across the samples, with chemical classes indicated by node color
Sex-specific differences in sEH metabolites along disease trajectories
To assess the changes in oxylipin metabolism along disease trajectory, we compared the differences in their levels between diagnosis groups, in the analysis stratified by sex. The following groups were used: cognitively normal (CN), mild cognitively impaired (MCI) and dementia (AD). We excluded subjects who were cognitively normal at baseline who change their status at the 4-year follow-up. The differences between the groups were compared using ANOVA with significant metabolites further converted into cluster components (Fig. 2). Factorial analysis with sex x group interaction was applied to identify metabolites affected differently by diagnosis group between males and females. Out of 224 metabolites and informative ratios and summations, 21 showed differences between groups. The 21 significant variables were condensed into 6 cluster components by variable clustering and reanalyzed by ANOVA with a Dunnett post-test against CN in a sex-stratified analysis (Fig. 2). Individual cluster members, their contribution to the cluster and their mean differences between groups are described in the Supplemental Table S2 and Supplemental Table S3. The main differences were observed in fatty acid vicinal diols (products of sEH metabolism, Clusters 1 for eicosapentanoic acid (EPA) and docosahexanoic acid (DHA) and Cluster 2 for the arachidonic acid (AA) derivatives) with sex differences. In Females, both omega-3 (EPA and DHA) and omega-6 (AA) derivatives were higher in AD when compared to the CN. However, the MCI was not different from the CN. Additionally, differences were observed in the levels of DHA-derived LOX metabolites (HDoHEs) and LPAs (Clusters 3 and 4) with MCI showing higher levels than CN and cortisol with AD showing higher levels than CN.Fig. 2. Soluble epoxide hydrolase metabolites are elevated along AD trajectory in a sex specific manner. Differences in means of lipid mediators between diagnosis groups, stratified by sex. To reduce data dimensionality and to facilitate interpretation and presentation, lipid mediators manifesting significant diagnosis group differences were converted into cluster components. Individual cluster members are defined in the cluster description, with the more detailed description of individual contribution to the cluster and differences in means between the treatment groups provided in the Supplemental Table S2 and Supplemental Table S3. Colors indicate diagnosis group: green – cognitively normal (CN; 83 females, 75 males) defined as healthy subjects who remind healthy after 4 years follow up; yellow—mild cognitively impaired (MCI; 179 females, 215 males); red – Alzheimer’s disease (AD; 53 females, 79 males). P values are derived from ANOVA with Dunnett post-test and indicate the differences from the CN
In males, omega-3 diols differences did not reach significance (e.g., AD vs stable CN p = 0.07). On the other hand, AA diols were higher in MCI and AD, compared to CN. This diagnosis group effect for both omega-3 and omega-6 vicinal diols was different from the one observed in females, with the diagnosis group x sex interaction p = 0.078 (for cluster 1, omega-3 diols) and p = 0.022 (for cluster 2, omega-6 diols) in the diagnosis group and sex factorial analysis. Additionally, 10-Nitrooleate was lower in MCI than CN and cortisol was higher in AD then CN.
AD-impacted individual oxylipins projected onto their metabolic pathways are presented in Supplemental Figure S2.
Lipid mediators and APOE4 discriminate between stable MCI and MCI converters
Metabolic signatures associated with AD resilience, i.e., remaining MCI in comparison to MCI that progresses further (within 2 years) into AD, are poorly understood. Utilizing ADNI longitudinal information about patient diagnosis status, we defined the minimal number of factors (both clinical outcomes and lipid mediators, including bile acids) that can describe the difference between converters and non-converters in a sex-stratified analysis (Fig. 3). In both sexes, combining APOE4 status with the plasma lipid mediator levels yielded the best discriminatory model: only APOE4 AUC = 64 for females and 62 for males; only lipid mediators AUC = 80 for females and 73 for males; APOE4 and lipid mediators AUC = 85 for females and 78 for males.Fig. 3. Lipid mediators and APOE genotype define AD resilient phenotype. Minimal model discriminating between stable MCI and MCI-AD converters (within 2 years), generated using stepwise logistic regression, stratified by sex. Biometrics (BMI, age,), presence of APOE4 allele and lipid mediators, including oxylipins, endocannabinoids and bile acids were used. The Receiver operation characteristics (ROC) curve is shown for the best model consisting of APOE4 information together with lipid mediators (red line) as well as for APOE4 alone (yellow line) and lipid mediators alone (orange line). The estimate and p values for each factor in the model are displayed in the ROC graph. The number of subjects: stable MCI = 153 females and 182 males; MCI converters = 32 females and 41 males
In females, the likelihood of remaining MCI without converting to AD (AD resilience) was defined by the lack of an APOE4 allele, lower glycine conjugation of chenodeoxycholic acid (define as GCDCA/CDCA ratio) and lower levels of EPA-derived 8iso-prostaglanding F3 alpha (8-iso PGF3a). In males, AD resilience was defined by the lack of APOE4 allele, lower cortisol and hyodeoxycholic acid (HDCA) and higher levels of lysophosphatidylinostitol palmitate (i.e., LPI16.0). Further factorial analysis with group x sex interaction (model adjusted by APOE, age and BMI) confirm sex-specific effect for GCDCA/CDCA (p_interaction_ = 0.04), 8iso_PGF3a (p_interaction_ = 0.003) and cortisol (p_interaction_ = 0.04). The interaction terms for HDCA and LPI16:0 were not significant.
The derived minimal models had similar statistical power to discriminate converter vs non-converters whether the model was applied for the 1, 2 or 4 years follow up conversion criteria.
Additionally, we did not include CSF biomarkers of amyloid or AD-related pathology in the current model and acknowledge that it is the main determinant of the following transition to AD [56]. After addition of CSF Tau/AB42 into the model: 1) in males, the minimal model would include Tau/AB42 (est = −2.78, p < 0.0001) and 14,15- dihydroxy eicosatrienoic acid (DiHETrE)/AA (est = −1>, p = 0.01) resulting with the ROC of 0.79; in females Tau/AB42 is the only component of the models (est = −2.8, p < 0.0001) with the ROC of 0.81. It is important to notice that among stable MCI group, the bimodal distribution of the CSF Tau/AB42 was observed, however we do not have sufficient number of participants to perform additional stratification by CSF Tau/AB42 in addition to sex.
Markers of cognitive resilience among differential Tau and amyloid pathology, mediated by APOE genotypes
Next, we sought to define peripheral markers of memory function, measured by ADNI composite memory score[52] within the cohort. We are particularly interested in those markers in subjects manifesting high Tau and amyloid pathology, as indicators of cognitive resilience (understood as preserving cognitive functions despite AD pathological signatures). AD related pathologies are measured by the ratio of total Tau to AB42 in CSF. In the ADNI GO/2 population, the Tau/AB42 ratio in CSF is influenced by APOE genotype, with e2 allele manifesting the least and e4 allele manifesting the highest level (Supplemental Figure S3). Each genotype manifested a range of memory scores, although e2e2 and e2e4 genotypes did not have sufficient number of participants for further analysis (n = 3 and = 14, respectively). To define the relationship between peripheral metabolism, Tau and amyloid pathology and memory, we applied stratification strategies for correlation analysis between plasma lipid mediators and the memory score. For correlation analysis, we applied a combination of random forest for features selection and stepwise linear regression to define a minimal model that explains the relation between memory and plasma lipid mediators and clinical outcomes including age, sex, BMI, APOE4 status, years of education (see materials and methods).
We identified APOE genotype stratification to yield significant models for associations of plasma metabolites and memory scores (Fig. 4) in comparison to stratification by CSF Tau/AB42 (using previously published cutoff value [57]), which did not produce significant models. In addition to the Tau/AB42, we applied pTau181 (pos/neg) stratification to reflect other markers for amyloid pathology (Cutoff values for pTau181 was provided by ADNI with value of 19.2 pg/mL). For pTau181 negative subjects, the memory was mostly predicted by age, sex and education (the three factors gave Rsq value of 0.15). The final minimal model did include 17-HDoHE increasing the Rsq to 0.17. For pTau181 positive subjects, the minimal model included 7 factors and gave Rsq of 0.22. The model included presence of ApoE4 allele, the ratio of 19_20_DiHDPA/19_20_EpDPE, age, sex, education, 14_15_DiHETrE and LPI 20.4.Fig. 4. Lipid mediators reveal heterogeneity among APOE genotypes and predictors of the memory function. Multilinear model showing differential contribution of key peripheral metabolites and other factors such as age and education to memory scores (MEM) across APOE genotypes. The number of subjects: APOE2/3 = 60; APOE3/3 = 334; APOE3/4 = 254; APOE4/4 = 77. More details about the model are presented in the Supplemental Table S5
Each APOE genotype manifested unique sets of variables contributing to the model. Memory in APOE 2/3 was mainly defined by the age (negative association, p = 0.0004) and 10-Nitrooleate (positive association, p = 0.0005), in addition to lesser contribution of the ratio of glycochenodeoxycholic acid (GCDCA) to glycolithocholic acid (GLCA) (positive association, p = 0.018) and LPG16:1 (negative association, p = 0.015). Those 4 variables generated a memory model with the RSq = 0.49 with the n = 71. Memory in APOE3/3 genotype subjects was defined by age, the ratio of 19,20-DiHDPA/19,20-EpDPE (DiHDPA – dihydroxy docosapentaenoic aicd; EpDPE – epoxy docosapentaenoic acid), LPE22:5 (LPE – lysophosphatidylethanolamine. Negative association, p = 0.0001, 0.0079, 0.0028 respectively) and the years of education, LPI20:4 (LPI – lysophosphatidylinositol) and the ratio of 5-HETE/AA (HETE – hydroxy eicosatrienoic acid; AA – arachidonic acid. Positive association, p = 0.0001, 0.0013 and 0.015 respectively). Those 6 variables generated a memory model with the RSq = 0.18 with the n = 387. Memory in APOE3/4 genotype subjects was defined by age, 19,20-DiHDPA and LPE16:0 (negative association, p < 0.0001, 0.005, and 0.019 respectively). Those 3 variables generated a memory model with the RSq = 0.16 with the n = 283. Memory in APOE4/4 genotype subjects was defined by an unknown (unidentified) oxylipin, LPI16:1, 8HDoHE (HDoHE – hydroxydocosahexaenoic acid. Positive association, p = 0.0004, 0.0075 and 0.0096 respectively) and cortisol, leukotriene E4 (LTE4) and glycodeoxycholic acid GDCA (negative association, p = 0.0015, 0.0021, 0.0027 respectively). Those 6 variables generated a memory model with the RSq = 0.46 with the n = 78. The unknown (unidentified) oxylipin manifested mass transition of Resolvin E2 (m/z 333—> m/z 115, the 115 fragment is for carboxyl group until 4’ carbon, as it breaks by 5’ hydroxyl group), but the chromatography retention time was different from the analytical standard. Additional correlative analysis shows a high correlation of this unknown oxylipin with EPA- derived vicinal diols (data not shown). Considering the importance of this metabolite in our predictive model and poor understanding of Resolvin biology, the full identification of this compound is of interest and is underway.
To further demonstrate the APOE genotype specificity of those associations, we applied a factorial analysis for the key factors identified in the stepwise model (age, education, 10-Nitrooleate, 19,20-DiHDPA, unknown oxylipin, LTE4 and cortisol) to test the memory score interaction with APOE genotype. Following p_interactions_ were achieved with other key factors used as covariates in the model: age p_interaction_ = 0.06 for e4e4 vs e3e4; 0.35 for e4e4 vs e3e3 and 0.09 for e4e4 vs e2e3; education p_interaction_ = 0.67 for e3e3 vs e3e4; 0.45 for e3e3 vs e4e4 and 0.8 for e3e3 vs e2e3; 10-Nitrooleate p_interaction_ = 0.037 for e2e3 vs e3e3; 0.098 for e2e3 vs e3e4 and 0.74 for e2e3 vs e4e4; 19,20-DiHDPA p_interaction_ = 0.04 for e3e4 vs e4e4; 0.015 for e3e4 vs e3e3 and 0.05 for e3e4 vs e2e3; unknown oxylipin p_interaction_ = 0.12 for e4e4 vs e3e4; 0.07 for e4e4 vs e3e3 and 0.28 for e4e4 vs e2e3; LTE4 p_interaction_ = 0.05 for e4e4 vs e3e4; 0.0052 for e4e4 vs e3e3 and 0.0043 for e4e4 vs e2e3; cortisol p_interaction_ = 0.11 for e4e4 vs e3e4; 0.062 for e4e4 vs e3e3 and 0.17 for e4e4 vs e2e3;
Stepwise regression model identifies a minimal number of factors for predicted values and eliminates collinear variables. For example, 19,20-DiHDPA is highly correlated with other omega-3 diols, like EPA derived dihydroxy eicosatetraenoic acid (DiHETE)s, and replacing 19,20-DiHDPA with closely correlated DiHETEs would yield similar predictive power. To illustrate the collinear relationship between measured lipid mediators and to facilitate minimal model interpretation, metabolite partial correlation network is presented in Figure S3.
Following on those findings we investigated whether GlycA, a marker of systemic inflammation, is differentially associated with cognition among ApoE genotypes. The interaction analysis indeed identified significant GlyC ApoE genotype interaction for the ApoE3/3 and ApoE4/4 genotypes (p = 0.02). The strength of the association between GlycA and cognition was increasing as the number of ApoE4 copies increased (i.e. ApoE2/3 est = −0.001, p = 1; ApoE3/3 est = −0.42, p = 0.007; ApoE3/4 est = −0.51, p = 0.01; ApoE4/4 est = −1.1, p = 0.0004).
Of note, the small N prevented us from investigating the impact of sex on those interactions in all genotypes. However, we were able to analyze the sex x MEM interaction for metabolites selected in the e3e3 and e3e4 genotypes. We found significant interaction for 5-HETE in e3e3 group (the est for males was 0.06 and for females −0.7). In the e3e4 group two interactions were found, for LPE22.5 (the est for males was −0.12 and for females −0.0002) and for LPI20.4 (the est for males was 0.09 and for females −0.05).
Discussion
Increased inflammatory responses have been reported in previous AD investigations but how these systems change over the progression of this disease have not been clearly delineated. Here we show a dysregulation of lipid mediators throughout AD progression and uncover disease heterogeneity influenced by sex and APOE genotype. We confirmed and extended our previous findings, from two independent cohorts, of increased soluble epoxide hydrolase activity during AD development. We also provide evidence implicating the APOE4 allele, bile acid metabolism, lipid peroxidation and stress response as sex-specific factors influencing the resistance of MCI subjects to progress to ADAPOE. Additionally, we showed APOE4/4 genotype-specific associations between plasma inflammatory markers and memory, identifying populations for precision medicine approach for potential therapeutic treatment.
Growing evidence points towards the sEH as a novel therapeutic target for AD [28, 58]. Furthermore, plasma sEH metabolites (derived from AA, EPA and DHA) increase with AD pathology [57] and these metabolites are associated with cognition (i.e. perceptual speed) in healthy and MCI individuals [42]. A strong connection of sEH-associated biochemistry with AD-affected vascular inflammation and energy metabolism in the central nervous system was also observed [59]. Increased sEH expression in the AD-afflicted brain at both the transcript and protein levels are also seen [60]. In rodent models of AD, AD-related pathologies can be altered by manipulating both sEH [28, 29] and CYP [30, 31] metabolism. Moreover, several genetic polymorphisms in the CYP/sEH pathway have consistently pointed towards reduced CYP-dependent epoxy fatty acid production and increased sEH activity or expression to be a risk for AD and increase AD-related pathologies [61–64], supporting causative effects of this critical inflammatory switch in the progression of this disease.
The current work provides additional evidence from a third independent AD cohort to support the dysregulation of sEH-dependent metabolism during AD development. Furthermore, the current study provides additional granularity (level of complexity) regarding sex, APOE genotype and pre-AD stages. In particular, we saw an increase in EPA, DHA and AA-derived sEH products in females with AD, but not with MCI, compared to CN participants. In contrast, in males, we only saw an increase in AA-derived sEH metabolites, but they were present in subjects with MCI, suggesting that dysregulation of sEH in males may appear earlier in disease progression than in females. Similarly, differences in sEH metabolism have been reported to contribute to the sex difference in the vulnerability to other dementia risk factors, such as ischemic brain injury [65, 66], high glycemic diet [67–70] and high-fat diet [71, 72]. At the molecular level, the possible involvement of sEH in sex-specific AD development includes a wide range fatty acid diol cellular functions (or depletion of fatty acids epoxide), e.g., ferroptosis-mediated neurodegeneration [73], exciting neurons [74], immune regulation [75, 76], and regulating mitochondrial respiration [77]. For the epoxy fatty acids, their action is beneficial or detrimental depending on factors such as precursor fatty acids, concentration, targeting cells, and biological processes [75, 78]. Sex-specific regulation of sEH, especially in response to sex hormones is broadly in the following review [79]. Together, this evidence suggests that reducing sEH activity may be a promising therapeutic approach in participants with both MCI and AD. These may include the use of sEH inhibitors currently under development [80, 81], but also include current therapies such as estrogen replacement therapy which can both down regulate sEH [82] and reduce AD risk in post-menopausal women [83].
Our results also support a sex-specific dysregulation of cortisol metabolism in AD, as seen by others [84, 85]. Particularly we observed, increased cortisol concentrations in AD subjects when compared to CN in both sexes; increased levels in males MCI subjects that further convert to AD within 2 years, when compared to stable MCI phenotype; and negative association with memory in APOE 4/4 carriers. Cortisol is a stress response hormone that facilitates glucose dependent energy metabolism. This hormone crosses blood brain barrier via P-glycoprotein transporter [86] consistent with the strong correlation between plasma and CSF cortisol levels [57]. Cortisol was previously implicated in neurodegeneration [87] and cognition [88] via regulation of the Hypothalamic–Pituitary–Adrenal (HPA) axis [89, 90] and was suggested by others as potential therapeutic target for AD [91]. Sex-related differences were also reported, with plasma cortisol associations with brain volumes and memory performance being more pronounced in females than in males [92]. We also show greater difference between CN and AD in females then in males. Our analysis here also adds to those findings by extending our understanding of cortisol and stress involvement in cognition and AD in specific subpopulations. In particular, we noted that males with MCI have elevated cortisol prior to conversion to AD, an effect not observed in females. This appears to contradict the previous findings suggesting a greater importance of this stress hormone in females with AD. Additionally in subjects with APOE 4/4 genotype, we show evidence that cortisol may impact memory performance, together with immune cells regulators. Our finding adds granularity to the known associations of stress hormones, cognition and AD development, identifying subpopulations for further potential therapeutic targeting of cortisol pathway in AD.
The current work also indicates the differential relationship between markers of immune cells activation and memory performance among various APOE genotype carriers. Notably in the presence of the APOE 4/4 genotype, memory was negatively associated with the pro-inflammatory cysteinyl leukotriene metabolism reported by LTE4, generated via 5-LOX-dependent metabolism. LTE4 is a monocyte activator with vasoactive properties, activating vascular permeability and edema [93], and its levels were associated with cardiovascular disorders [94, 95]. Previously, 5-LOX expression was found to be upregulated in the peripheral blood mononuclear cells from AD participants [96, 97]. Furthermore, a gain of function 5-LOX polymorphism was also associated with risk of AD [98], indicating a potential causative relationship of the dysregulation of this pathway to AD. Meanwhile in rodent models of AD upregulation of hippocampal and cortex 5-LOX expression was associated with elevated γ-secretase activity and Aβ peptide formation [97, 99–102]. APOE genotypes-specific oxylipin profile in postmortem human dorsolateral prefrontal cortex found that 5-LOX-derived EPA oxylipins LXA5 and LXB5 in brain were negatively associated with cognitive performance, and the association was stronger among APOE4 carriers compared to APOE3 carriers [103]. This research also found that the levels of a 5-LOX product, LXA4 was negatively associated with sEH activity. On the basis of this, our results here further indicate that the dynamics between APOE genotypes, 5-LOX, sEH, fatty acid precursors and cognition can be captured in the blood lipidome, which can be monitored less invasively and more routinely for longitudinal cohorts.
The functional interaction between APOE, 5-LOX and sEH can be supported by various mechanisms: for example, the higher level of APOE4 carriers in activation of Ca^2+^ influx and thus Ca^2+^-dependent cPLA2 and 5-LOX compared to APOE2 or E3 [103, 104]; for another, the presence of prone-to-aggregation lipid-poor APOE4 protein can affect the lipid efflux functions of ABCA1[105] and change the lipid composition of cells [106], which serve as substrates to release free fatty acids subject to 5-LOX and sEH metabolism [107, 108]. A possibility also remains that APOE4 leads to the altered ability of trafficking inflammation-regulating lipid mediators between lipoprotein and lipoprotein-metabolizing cells, which includes microglia, a critical player in AD pathogenesis [107]. This could be supported by a strong association of memory and GlycA in ApoE4/4 carriers, and less pronounced associations among other genotypes. Further investigation is therefore warranted to clarify if the combination of altering 5-LOX metabolism and fatty acid composition together can be a specific modifiable target for APOE4 carriers to improve cognition. Lipid mediators affected during AD development, projected onto the metabolic pathway are presented in the Supplemental Figure S2.
Limitations
This study is not big enough to explore APOE genotypes and sex combined stratification. In particular, APOE x sex stratification resulted in very small subgroups with insufficient numbers to yield reliable results, and therefore these analyses were not performed. ADNI 1 and GO/2 cohorts do not represent full demographics of US and world population. This shortcoming is addressed in ADNI 3 cohort. Longitudinal data are needed to further connect dysregulation in lipid mediators, sEH among others and disease progression, define subgroups to identify population for targeted interventions. Further, the analysis of converters vs non-converters has a small n and should be validated in the larger cohort. Limited availability of CSF pTau181 and amyloid-PET in this cohort further constrained amyloid-based stratifications. Future studies with larger or pooled longitudinal cohorts will be necessary to enable adequately powered amyloid-defined subgroup analyses alongside clinical diagnoses. Finally, the diols and the ratio of epoxides to diols are our best attempt to approximate sEH activity and the results should be understood as suggestive.
Supplementary Information
Supplementary Material 1. Supplementary Material 2. Supplementary Material 3. Supplementary Material 4. Supplementary Material 5. Supplementary Material 6. Supplementary Material 7. Supplementary Material 8.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Karu N, Kindt A, Lamont L, van Gammeren AJ, Ermens AAM, Harms AC, Portengen L, Vermeulen RCH, Dik WA, Langerak AW, van der Velden VHJ, Hankemeier T. Plasma Oxylipins and Their Precursors Are Strongly Associated with COVID-19 Severity and with Immune Response Markers. Metabolites. 2022;127.10.3390/metabo 12070619 PMC 931989735888743 · doi ↗ · pubmed ↗
- 2Ikonomovic MD, Abrahamson Ee Fau - Uz T, Uz T Fau - Manev H, Manev H Fau - Dekosky ST, Dekosky ST. Increased 5-lipoxygenase immunoreactivity in the hippocampus of patients with Alzheimer's disease(0022–1554 (Print)).10.1369/jhc.2008.951855 PMC 258390718678882 · doi ↗ · pubmed ↗
- 3Xiang Z, Ho L Fau - Yemul S, Yemul S Fau - Zhao Z, Zhao Z Fau - Qing W, Qing W Fau - Pompl P, Pompl P Fau - Kelley K, Kelley K Fau - Dang A, Dang A Fau - Qing W, Qing W Fau - Teplow D, Teplow D Fau - Pasinetti GM, Pasinetti GM. Cyclooxygenase-2 promotes amyloid plaque deposition in a mouse model of Alzheimer's disease neuropathology(1052–2166 (Print)).10.3727/000000002783992352 PMC 597752512450219 · doi ↗ · pubmed ↗
- 4Ghosh AA-O, Comerota MM, Wan D, Chen F, Propson NE, Hwang SA-OX, Hammock BD, Zheng HA-O. An epoxide hydrolase inhibitor reduces neuroinflammation in a mouse model of Alzheimer's disease. LID - 10.1126/scitranslmed.abb 1206 LID - eabb 1206(1946–6242 (Electronic)).10.1126/scitranslmed.abb 1206 PMC 778455533298560 · doi ↗ · pubmed ↗
- 5Ruiz-Pérez G, Ruiz de Martín Esteban S, Marqués S, Aparicio N, Grande MT, Benito-Cuesta I, Martínez-Relimpio AM, Arnanz MA, Tolón RM, Posada-Ayala M, Cravatt BF, Esteban JA, Romero J, Palenzuela R. Potentiation of amyloid beta phagocytosis and amelioration of synaptic dysfunction upon FAAH deletion in a mouse model of Alzheimer's disease. Journal of Neuroinflammation. 2021;18(1):223. 10.1186/s 12974-021-02276-y.10.1186/s 12974-021-02276-y PMC 848261434587978 · doi ↗ · pubmed ↗
- 6Bajaj SA-O, Zameer S, Jain S, Yadav V, Vohora D. Effect of the MAGL/FAAH Dual Inhibitor JZL-195 on Streptozotocin-Induced Alzheimer's Disease-like Sporadic Dementia in Mice with an Emphasis on Aβ, HSP-70, Neuroinflammation, and Oxidative Stress(1948-7193 (Electronic)).10.1021/acschemneuro.1c 0069935316021 · doi ↗ · pubmed ↗
- 7Qu T, Manev R, Manev H. 5-Lipoxygenase (5-LOX) promoter polymorphism in patients with early-onset and late-onset Alzheimer's disease. J Neuropsychiatry Clin Neurosci. 2001;13(2):304-5. 10.1176/jnp.13.2.304. Pub Med PMID: 11449041.10.1176/jnp.13.2.30411449041 · doi ↗ · pubmed ↗
- 8Toledo JB, Arnold M, Kastenmuller G, Chang R, Baillie RA, Han X, Thambisetty M, Tenenbaum JD, Suhre K, Thompson JW, John-Williams LS, Mahmoudian Dehkordi S, Rotroff DM, Jack JR, Motsinger-Reif A, Risacher SL, Blach C, Lucas JE, Massaro T, Louie G, Zhu H, Dallmann G, Klavins K, Koal T, Kim S, Nho K, Shen L, Casanova R, Varma S, Legido-Quigley C, Moseley MA, Zhu K, Henrion MYR, van der Lee SJ, Harms AC, Demirkan A, Hankemeier T, van Duijn CM, Trojanowski JQ, Shaw LM, Saykin AJ, Weiner MW, Doraiswam · doi ↗ · pubmed ↗