Rapid Analysis of Pacific Ciguatoxins in Fish Extracts with a Lateral Flow Assay
Ulises G. Díaz-Avello, Vasso Skouridou, Takeshi Tsumuraya, Masahiro Hirama, Naomasa Oshiro, Mònica Campàs, Ciara K. O’Sullivan

TL;DR
This paper introduces a fast and reliable test for detecting Pacific ciguatoxins in fish, which can help prevent poisoning in non-endemic regions.
Contribution
The first lateral flow assay for detecting major Pacific ciguatoxin congeners using monoclonal antibodies.
Findings
The LFA detects Pacific CTXs with a visual limit of 400 pg/mL and no cross-reactivity with other marine toxins.
The assay successfully identified CTXs in both spiked and naturally contaminated fish samples.
The test provides visual results in 30 minutes and is suitable for use in resource-limited settings.
Abstract
The increasing spread of toxin-producing Gambierdiscus dinoflagellates has led to cases of ciguatera poisoning in previously nonendemic regions. Ciguatoxins (CTXs) are potent, heat-stable compounds responsible for ciguatera, and their rapid and reliable analysis in seafood is essential to prevent illness. This study presents the development of the first lateral flow assay (LFA) for the detection of major Pacific CTXs congeners. The assay employs previously developed monoclonal antibodies in a sandwich format: specific antibodies against CTX1B and CTX3C are immobilized on a single test line, while the reporter antibody is a single antibody recognizing major Pacific CTX congeners that is conjugated to gold nanoparticles. The LFA provides one-step visual results in 30 min, making it suitable for resource-limited settings. It exhibits a visual limit of detection of 400 pg/mL and the assay…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
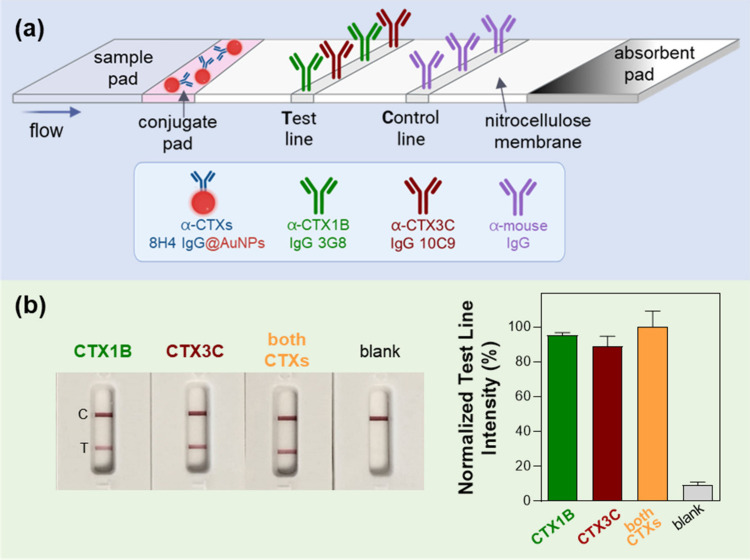 Figure 1
Figure 1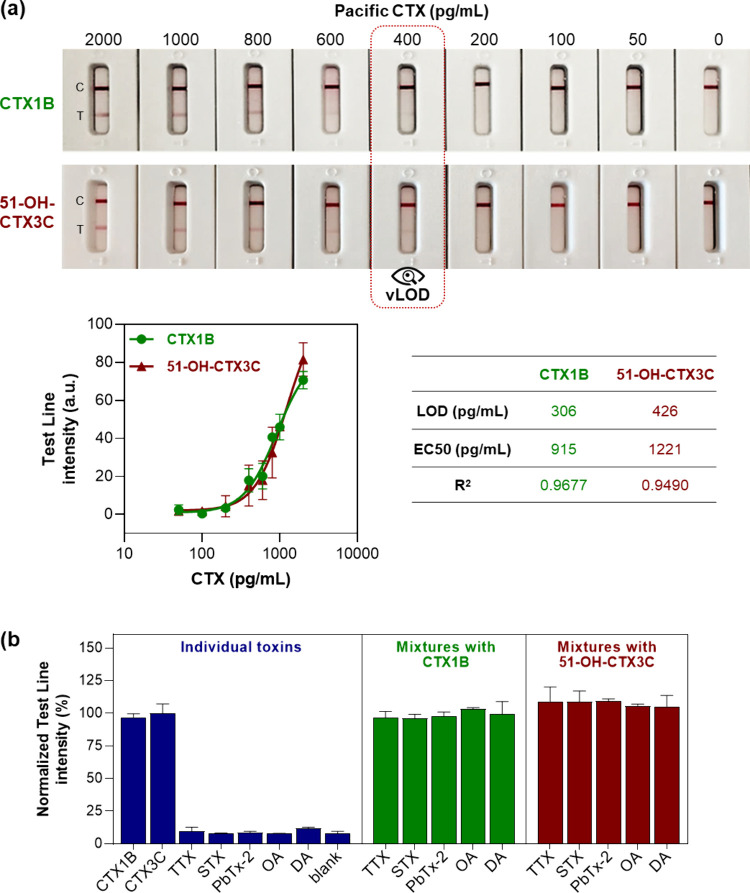 Figure 2
Figure 2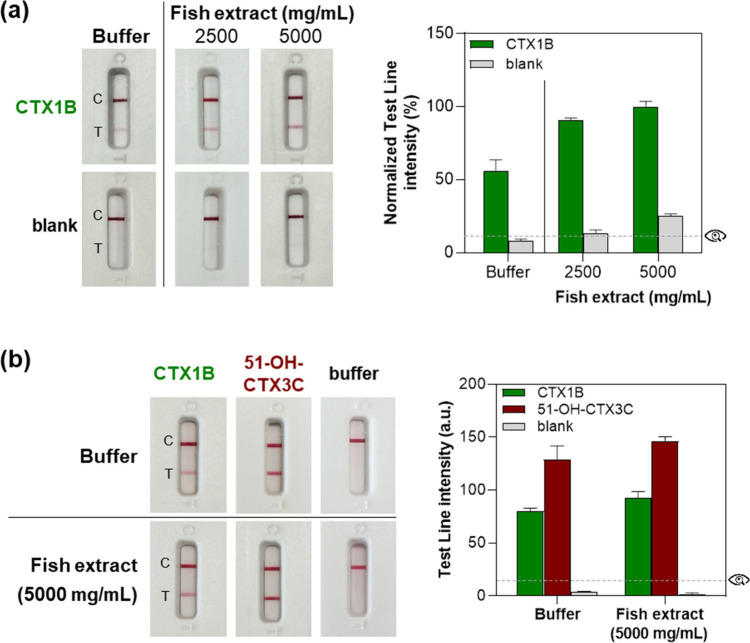 Figure 3
Figure 3 Figure 4
Figure 4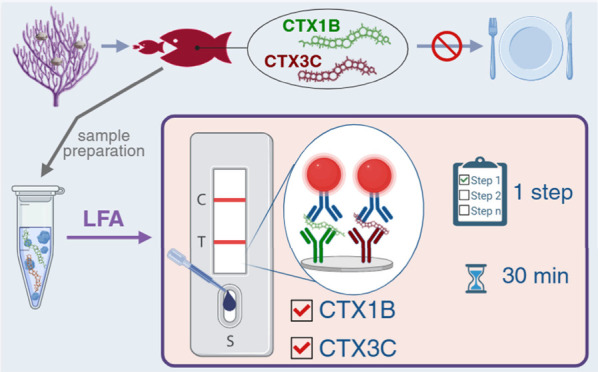 Figure 5
Figure 5 Figure 6
Figure 6- —Ministerio de Ciencia, Innovación y Universidades10.13039/100014440
- —Ministerio de Ciencia, Innovación y Universidades10.13039/100014440
- —Agencia Estatal de Investigación10.13039/501100011033
- —Agencia Estatal de Investigación10.13039/501100011033
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMarine Toxins and Detection Methods · Vibrio bacteria research studies · Melamine detection and toxicity
Marine toxins are naturally occurring compounds primarily produced by certain types of algae, cyanobacteria, diatoms, and dinoflagellates.? They can cause mild to severe illness in humans following the ingestion of contaminated seafood, presenting a wide spectrum of gastrointestinal and neurological symptoms.? Ciguatoxins (CTXs) are potent neurotoxins responsible for ciguatera poisoning (CP), the most frequent seafood-borne illness worldwide,? and they or their precursors are produced by benthic dinoflagellate species of Gambierdiscus.? Ciguatera can cause a variety of gastrointestinal, neurological, and cardiovascular symptoms, often complicating diagnosis due to their nonspecific nature.? It is estimated to affect up to 50,000 people annually, although challenges related to accurate diagnosis and the lack of accessible detection tools are believed to result in significant under-reporting.? Ciguatera was traditionally associated with tropical and subtropical regions; however, the rising water temperatures, disturbance of coral reefs and the globalization of the seafood trade may have contributed to its expansion to nonendemic areas. ?,?,?,? It is now considered an emerging risk in Europe due to the reported intoxication cases, as well as the presence of Gambierdiscus in coastal areas and the detection of CTXs in fishes. ?,?,?
Ciguatoxins are lipid-soluble polycyclic ether compounds with more than 30 congeners, which can be classified into different groups based on their chemical structure. ?,? Pacific CTXs are generally considered as the most potent ones, exhibiting high intraperitoneal toxicity in mice, with half-lethal-dose (LD_50_) values of 0.25–0.9 μg/kg of body weight.? CTX1B (known as P-CTX-1), in particular, is one of the major congeners in the Pacific and the most potent one among its regional analogues, and it has been extensively characterized since the 1990s. ?,? The United States Food and Drug Administration (FDA) has established guidance levels of 0.01 μg/kg CTX1B equivalents and 0.1 μg/kg Caribbean CTX-1 (C-CTX1).? There is no explicit regulatory limit in Europe, but it is prohibited to sell fish with detectable levels of CTXs.? CTXs are heat-stable, cannot be eliminated by cooking or freezing, and have no known antidotes. Consequently, the most effective strategy to mitigate the risk of CP is to avoid consumption of contaminated seafood, which relies on early and reliable detection of the toxins.
Different methods exist for the detection of CTXs which can be broadly classified into biological, chemical and biochemical. ?−? ? Biological methods include the mouse bioassay, cell-based cytotoxicity assays,? and receptor binding assays.? While they offer several advantages, limitations in sensitivity and specificity, costs, and ethical concerns have driven the development of alternative assays. Chemical methods, and specifically liquid chromatography-tandem mass spectroscopy (LC-MS/MS), are the leading techniques for not only detecting CTXs but also providing toxin profiles, and they exhibit very high sensitivity and specificity.? Biochemical assays rely on the use of specific antibodies that recognize CTXs in Enzyme-Linked Immunosorbent Assays (ELISA) and different biosensor formats.
The efforts in developing CTX-specific antibodies began in the late 1970s,? but it was not until the beginning of the 2000s that high-affinity and specific antibodies against Pacific CTXs were reported. Using rationally designed synthetic haptens mimicking parts of the CTXs molecules, mouse monoclonal antibodies were developed against the left wings of CTX1B (3G8 IgG)? and CTX3C (10C9 IgG),? and an antibody against the right wing of CTX1B, also able to bind to the right wing of CTX3C, even though it lacks the M-ring hydroxyl group (8H4 IgG).? By combining the three antibodies 3G8, 10C9, and 8H4 in one single sandwich ELISA, they demonstrated the detection of any of the four main congeners of Pacific CTXs (CTX1B, 54-deoxyCTX1B, CTX3C and 51-OH-CTX3C) with a limit of detection (LOD) of less than 1 pg/mL and the successful detection of CTX1B spiked in fish flesh at the FDA guidance level of 0.01 μg/kg.? The same antibodies were later exploited for the development of magnetic bead-based colorimetric assays and electrochemical biosensors with LODs at the low pg/mL level. ?−? ? ?
Lateral flow assays (LFA) have garnered a lot of interest over the last years for the detection of marine toxins. ?,? These simple, rapid, and low-cost tests can be used in resource-limited settings and can provide visual results in less than 30 min, facilitating timely decision-making and ensuring food safety. The majority of LFAs reported in the literature to date have employed antibodies in a competitive format with signal-off colorimetric visual detection. They targeted primarily tetrodotoxin (TTX), as well as okadaic acid (OA), domoic acid (DA), saxitoxins (STXs), dinophysistoxins (DTXs), brevetoxin B (PbTx-2) and cyclic imines (CIs), for single analyte or multiplex detection of several toxins simultaneously (Supporting Information Table S1).
In this work, we report the first LFA test for the detection of the main congeners of CTX1B and CTX3C with one single test, based on a signal-on sandwich format employing the antibodies previously developed by Tsumuraya and co-workers.? The design of the LFA test is illustrated in Figurea: the 3G8 IgG against the left wing of CTX1B and the 10C9 IgG against the left wing of CTX3C are immobilized at equal concentrations at the test line, whereas the 8H4 IgG against the common right wing of CTX1B and CTX3C is conjugated to 40 nm gold nanoparticles (8H4@AuNPs conjugate) and deposited on the conjugate pad. Once the sample is applied to the sample pad, it flows through the strip by capillary forces. If any of the main congeners of CTXs is present in the sample (CTX1B, 54-deoxyCTX1B, CTX3C or 51-OH-CTX3C), it will be bound by the 8H4 antibody immobilized on the AuNPs and the complex will be captured at the test line by one of the two antibodies, depending on the specific CTX present in the sample. This will ultimately result in the formation of a red line. Remaining 8H4@AuNPs conjugate will be captured at the control line by an anti-mouse antibody, forming a red line and indicating correct function of the test (Figure S1).
The 8H4@AuNPs conjugate was prepared by first optimizing the concentration of the antibody required to provide colloidal stability (Figure S2a), and the final conjugate was characterized by UV–vis spectroscopy (Figure S2b). The tests were prepared and assembled as detailed in the Supporting Information. The design of the test was evaluated by analyzing CTX1B and 51-OH-CTX3C individually and in combination. Due to the lack of high-quality and accurately quantified CTX3C standards, the 51-OH-CTX3C congener was used as a representative model for CTX3C-type congeners, since it is recognized by the corresponding antibodies to the same extent as CTX3C. ?,? Each sample (50 μL) was applied to the LFA test, followed by the addition of running buffer (100 μL). Test line intensities were measured after 30 min with a portable LFA reader. As shown in Figureb, the relative intensities obtained for each individual toxin at 1000 pg/mL were comparable to that of the mixture containing both toxins at 500 pg/mL each (total concentration of 1000 pg/mL). These results confirm that the test is capable of detecting both toxins simultaneously without signal interference.
Several test parameters were subsequently optimized to enable the sensitive detection of CTXs. First, LFAs incorporating different nanoparticle conjugates were fabricated to identify the one providing the most sensitive and specific detection. To this end, the reporter 8H4 antibody was conjugated to (a) spherical AuNPs (40 nm), (b) gold nanourchins (AuNUs; 90 nm), (c) streptavidin immobilized on AuNPs (SA@AuNPs; 40 nm) and (d) carbon nanoparticles (CNPs; 25 nm average primary particle size) (Supporting Information). AuNUs are gold nanoparticles with spiky and uneven surfaces, exhibiting unique plasmonic properties with increased surface area compared to spherical AuNPs.? SA@AuNPs are often combined with biotinylated antibodies to achieve higher loading and potentially improve orientation and sensitivity.? Finally, CNPs are small, black, amorphous nanoparticles that can be efficiently functionalized with proteins. They are considerably more cost-effective than AuNPs and provide high visual contrast on white membranes.? CTX1B was analyzed at varying concentrations with each LFA, and the highest signal and signal-to-noise ratio were observed with spherical 40 nm AuNPs (Figure S3). Therefore, this conjugate was selected for further test development. The effect of nitrocellulose (NC) membrane type on signal intensity and assay run time was then evaluated. Two commonly used NC membranes, FF120HP and FF170HP, were compared, with FF170HP having smaller pores and a slower flow rate. When FF170HP was used, higher test line intensities were observed within a shorter assay time (25–30 min) as compared to FF120HP (Figure S4), and it was therefore selected for further development. The concentrations of antibodies used for constructing the test line were subsequently optimized. In parallel, the potential application of a silver enhancement step was assessed, based on previous studies demonstrating its effectiveness in improving assay sensitivity.? The highest test line intensity was observed when 1.5 mg/mL of each antibody (3G8 and 10C9) was used instead of 1 mg/mL (Figure S5). A slight increase in the signal was observed for an assay run time longer than 20 min, while silver enhancement had no significant effect. Therefore, a concentration of 1.5 mg/mL for both antibodies and an assay time of 30 min without silver enhancement were selected as the final test conditions.
All previous optimizations were performed using unblocked NC membranes, the sample pads were pretreated with a blocking agent (1% w/v BSA) and a detergent (0.05% w/v Tween-20), and the LFA running buffer was PBS. Blocking agents, often used in combination with detergents, are typically employed to prevent nonspecific signals and improve sensitivity by increasing the signal-to-noise ratio. To simplify test fabrication, unblocked membranes are preferred and any blocking agents required are incorporated in the sample pad.? In the case of the CTXs LFA test, nonspecific signals were occasionally observed when unblocked membranes were used in combination with pretreated sample pads. To eliminate these nonspecific signals, several running buffer additives were evaluated. BSA and skimmed milk resulted in the lowest test line intensities in blank samples without CTXs, although the lines were still visible to the naked eye, as any intensity above 10–12 arbitrary units (a.u., per LFA reader) is discernible (Figure S6a). The addition of BSA (1% w/v) to the running buffer was then assessed using both unblocked and blocked NC membranes. Blocking the membranes effectively eliminated nonspecific signals, and supplementation of the running buffer with BSA was not necessary when blocked membranes were used (Figure S6b). However, to prevent nonspecific signals when analyzing complex samples such as fish extracts, blocked membranes and BSA-supplemented running buffer were used.
The analytical sensitivity of the optimized LFA test was finally evaluated by constructing calibration curves for CTX1B and 51-OH-CTX3C separately. As shown in Figurea, test lines with gradually increasing intensities were observed with increasing CTXs concentration, consistent with signal-on tests. The visual limits of detection (vLOD) were 400 pg/mL for both CTX1B and 51-OH-CTX3C. When the portable LFA reader was used to measure the test line intensities, the LODs calculated from the sigmoidal curves were 306 pg/mL for CTX1B and 426 pg/mL for 51-OH-CTX3C. To further demonstrate the sensitivity of the assay, we tested each CTX individually at a concentration below the LOD, as well as a mixture in which the combined concentration corresponded to the vLOD. While the individual toxins at 200 pg/mL produced no visible test lines, their 200 + 200 pg/mL mixture resulted in a detectable positive signal (Figure S7). Overall, the vLOD of this test is superior to that of most LFA tests reported to date for other marine toxins (Table S1), potentially due to the sandwich format of the assay as compared to the competitive format typically employed for small molecules, as well as the high affinity of the monoclonal antibodies.
Furthermore, the test demonstrated high specificity, as no signals were observed for the other marine toxins tested (saxitoxin, domoic acid, okadaic acid, tetrodotoxin, and brevetoxin B) (Figureb). The presence of these toxins in the same sample did not interfere with the detection of CTXs, as indicated by the comparable relative signal intensities observed for CTXs tested alone and in combination with the other marine toxins.
The suitability of the Pacific CTXs LFA test for analyzing fish samples was assessed by using noncontaminated and contaminated extracts, prepared as detailed in the Supporting Information. First, an extract from noncontaminated Lutjanus bohar was spiked with CTX1B (50 μL of 600 pg/mL) at various matrix concentrations. Initial tests were conducted using unblocked NC membranes. CTX1B was successfully detected in all spiked extracts, regardless of matrix concentration, but significant overestimation was observed, with recoveries ranging from 161% to 178% (Figurea). In addition, nonspecific signals were detected with the neat (nonspiked) extracts. These background signals were effectively eliminated by using LFA strips fabricated with blocked NC membranes. When undiluted fish extract (50 μL of 5000 mg flesh equiv./mL, corresponding to 250 mg flesh equiv.) was used, both CTX1B and 51-OH-CTX3C, spiked in the extract at 1000 pg/mL, were efficiently detected with improved accuracy, resulting in recoveries of 115.3% and 113.7%, respectively (Figureb).
The LFA test was finally applied to naturally contaminated fish extracts from L. bohar, L. monostigma and Variola louti specimens (Table S2 and Figure). All extracts were analyzed without dilution, with the exception of samples FE20111 and FE20112, which were diluted 3-fold and 2-fold, respectively, prior to analysis. Out of the ten samples analyzed, nine had been previously confirmed to contain CTXs by LC-MS/MS,? while one was negative (Table S2). The LFA test detected CTXs in seven of the nine positive samples (Figure). False-negative results were observed for two samples: FE24002 and FE24003. Sample FE24003 contained a very low concentration of CTXs (66 pg/mL), below the LFA’s LOD (300–400 pg/mL), hence explaining the negative result. The false-negative result for sample FE24002 by visual inspection remains unclear, considering that LC-MS/MS reported a concentration of 543 pg/mL. It should be noted that CTX1B was the least abundant congener and that the 52-epi-54-deoxyCTX1B is probably less well recognized by the assay. Nevertheless, this sample was correctly identified as CTX-positive when the LFA reader was used to measure the intensity of the test line. The LFA test also correctly identified the negative sample as CTX-free.
It is necessary to mention that, in the present study, the naturally incurred samples were prepared following a multistep extraction and purification protocol, which produced highly purified fish extracts.? This, together with the use of blocked NC membranes, certainly contributed to the good performance observed when applying the LFA to both spiked and naturally contaminated samples, as cleaner extracts reduce potential interferences from matrix components. It is also important to note that matrix effects cannot be fully predicted, as the composition of fish tissues varies considerably between species, individuals, size and origin.? This variability may influence assay performance and could also explain, at least in part, the few inconsistent results. While the extraction protocol used here is effective, it is also relatively labor-intensive. There may be room for simplification of the extraction procedure to make the assay more suitable for field deployment. Future research should therefore focus on the development of rapid, reliable, and efficient extraction methods that ensure high recoveries of CTXs while maintaining minimal matrix interference. Optimizing sample processing and toxin extraction workflows will be essential to fully assess and expand the applicability of this LFA to field settings.
In conclusion, a simple and rapid LFA test was developed for the detection of the main congeners of Pacific CTXs. The test enables simple, one-step visual screening of fish samples within 30 min and operates with a cutoff of approximately 0.1 μg/kg, as demonstrated by the analysis of samples FE20101 and FE20110 without prior dilution. This cutoff is sufficient to detect CTXs in the food remnants of CP patients, which have been reported to contain a minimum of 0.025 MU/g (equivalent to 0.175 μg/kg CTX1B).? Lehane and Lewis? also reported that CTX1B can induce ciguatera symptoms at concentrations as low as 0.1 μg/kg. Other studies have similarly suggested that a threshold of 0.1–0.2 μg/kg could be more pragmatic for CP risk management than the U.S. FDA’s cautionary guidance level of 0.01 μg/kg. ?,? Achieving such sensitivity remains challenging due to the limited availability and high cost of purified CTX standards, which constrain the development of more sensitive detection methods.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Daguer H.Hoff R. B.Molognoni L.Kleemann C. R.Felizardo L. V.Outbreaks, toxicology, and analytical methods of marine toxins in foods Curr. Opin. Food Chem.201824435510.1016/j.cofs.2018.10.006 · doi ↗
- 2Morabito S.Silvestro S.Faggio C.How the marine biotoxins affect human health Nat. Prod. Res.20183262110.1080/14786419.2017.132973428532167 · doi ↗ · pubmed ↗
- 3Loeffler C. R.Tartaglione L.Friedemann M.Spielmeyer A.Kappenstein O.Bodi D.Ciguatera mini review: 21st century environmental challenges and the interdisciplinary research efforts rising to meet them Int. J. Environ. Res. Public Health 202118302710.3390/ijerph 1806302733804281 PMC 7999458 · doi ↗ · pubmed ↗
- 4Perkins J. C.Zenger K. R.Liu Y.Strugnell J. M.Ciguatera poisoning: a review of the ecology and detection methods for Gambierdiscus and Fukuyoa specias Harmful Algae 202413910273510.1016/j.hal.2024.10273539567072 · doi ↗ · pubmed ↗
- 5Friedman M. A.Fernandez M.Backer L. C.Dickey R. W.Bernstein J. M.Schrank K.Kibler S.Un updated review of ciguatera fish poisoning: clinical, epidemiological, environmental, and public health management Mar. Drugs 2017157210.3390/md 1503007228335428 PMC 5367029 · doi ↗ · pubmed ↗
- 6Alvito P.Gago-Martínez A.Ciguatera toxins, a potential health risk emerging in Europe: overview of progress and challenges Curr. Opin. Food Sci.20256310130310.1016/j.cofs.2025.101303 · doi ↗
- 7FAO and WHO . Report of the Expert Meeting on Ciguatera Poisoning. Rome, 19–23 November 2018. Food Safety and Quality 2020, 9. 10.4060/ca 8817 en. · doi ↗
- 8Holmes M. J.Lewis R. J.Reviewing evidence for disturbance to coral reefs increasing the risk of ciguatera Toxins 20251719510.3390/toxins 1704019540278692 PMC 12030847 · doi ↗ · pubmed ↗