Treatment of Metastatic Cancer by Conferring Immunogenicity to the Apoptotic Bodies of the Primary Tumor
So-Jung Kim, Hae-Bin Park, Eun-Koung An, Dayoung Ryu, Da Young Kim, Daeun Lim, Wei Zhang, Xiaoyan Zhang, Jianqing Xu, Peter Chang-Whan Lee, Jun-O Jin

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
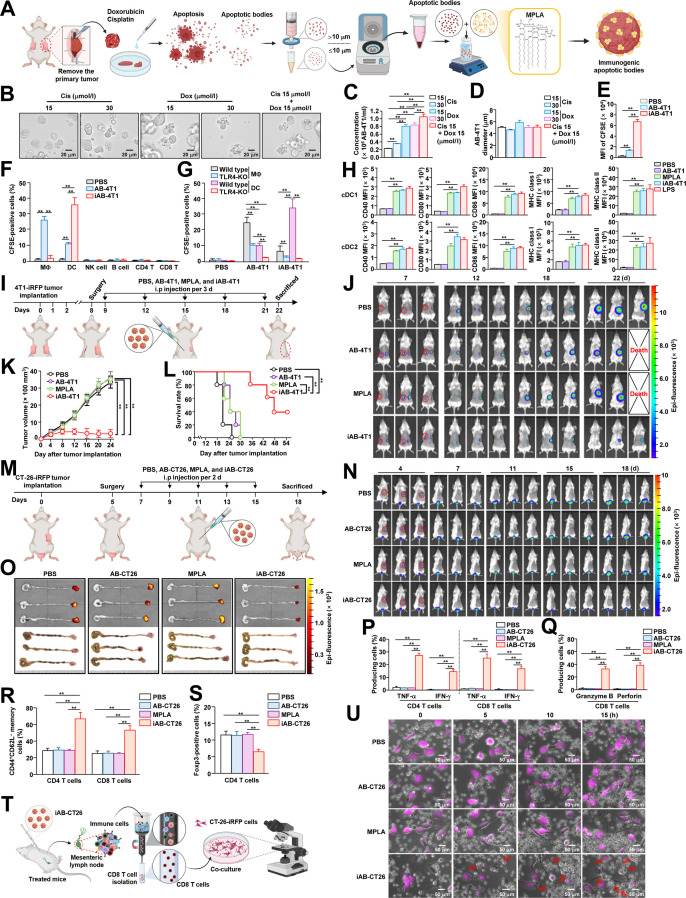 Figure 1
Figure 1- —National Research Foundation of Korea
- —National Research Foundation of Korea
- —National Research Foundation of Korea
- —Asan Institute for Life Sciences, Asan Medical Centerhttp://dx.doi.org/10.13039/501100005006
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhagocytosis and Immune Regulation · Immunotherapy and Immune Responses · Cancer Research and Treatments
The cure rate of cancer is increasing with the development of new treatment methods, including surgical treatments and immunotherapy [1]. Nevertheless, treatment of metastatic cancer remains challenging [2]. Immunotherapy is thought to suppress metastatic cancer, as it promotes specific killing of tumor cells via recognition of cancer antigens [3]. This antigen-specific immunity involving helper T cells and cytotoxic T lymphocyte activation is mediated by antigen presentation and stimulation of dendritic cells (DCs) [4]. Nonetheless, the strategies aimed at treating metastatic cancer by inducing the activation of DCs have not achieved substantial results [5]. Notably, inducing apoptosis in tumor cells has also been explored as a potential therapeutic strategy [6]. Apoptotic bodies (ABs), which are small membrane-bound vesicles released by dying cells during apoptosis, have been investigated as drug delivery vehicles to specifically target and destroy tumors [7]. However, by inducing transforming growth factor-β and interleukin-10, ABs could inhibit macrophage activation, thereby suppressing immune responses to antigens and promoting immune tolerance [8]. Consequently, the therapeutic application of ABs in tumor immunotherapy remains limited due to their potential to promote immune tolerance. Nonetheless, ABs derived from tumor cells contain abundant tumor antigens [9]. If sufficient immune activity can be elicited, these ABs may serve as a potential therapeutic approach for metastatic cancer. Therefore, we hypothesized that generating ABs using tumors extracted from a bilateral tumor model that mimics metastatic/residual disease and enhancing their immunogenicity could provide an effective strategy for treating metastatic cancer (Fig. 1A).
To that end, tumors from 4T1 cells expressing infrared fluorescent protein (iRFP), approximately 100 mm^3^ in size, were excised from tumor-bearing mice and processed into single-cell suspensions (Supplementary Materials and Table S1). The 4T1-iRFP tumor cells were then subjected to cisplatin (Cis) and/or doxorubicin (Dox), leading to apoptosis (Fig. S1A and B) and the production of AB (AB-4T1) within 24 h (Fig. 1B). Compared with each agent alone, AB-4T1 production was more efficient when a combination of Cis and Dox was used (Fig. 1C and Table S2). The harvested AB-4T1 was approximately 5 μm in diameter (Fig. 1D). To confer immunogenicity to AB-4T1, monophosphoryl lipid A (MPLA), a detoxified lipopolysaccharide derivative and Toll-like receptor 4 (TLR4) agonist [10], was selected as an immune adjuvant. Lipid insertion into AB-4T1 to produce immunogenic apoptotic bodies (iABs) was assessed using DSPE–PEG2000–fluorescein isothiocyanate, revealing a concentration-dependent increase in insertion, with fluorescence rising stepwise up to 50 μg (Fig. S1C and D). Importantly, MPLA insertion did not induce marked changes in AB size, morphology, or protein composition over the 10-d observation period (Fig. S1E to G).
We then verified DC targeting, showing that iAB-4T1 was effectively phagocytosed by bone-marrow-derived DCs (Fig. 1E and Fig. S2A and B). To assess in vivo uptake, immune cells from the spleen were isolated and analyzed (Fig. S2C). While AB-4T1 was efficiently internalized by F4/80^+^ macrophages, iAB-4T1 exhibited a higher uptake efficiency by DCs (Fig. 1F and Fig. S2D). As previously reported [8], AB-4T1 administered in vivo was predominantly phagocytosed by macrophages, contributing to their clearance and potential immune suppression. Notably, iAB-4T1 uptake by DCs was significantly reduced in TLR4-knockout mice, indicating that this process occurred in a TLR4-dependent manner (Fig. 1G).
To conduct experiments under conditions similar to those in patients with metastatic cancer or residual disease, 4T1-iRFP tumors were implanted on both sides of the mammary gland; the tumors on one side were surgically removed, and iAB-4T1 was generated. Of note, iAB-4T1 containing 40 μg of MPLA markedly enhanced activation marker (CD40, CD80, and CD86) expressions in conventional DC 1 (cDC1) and cDC2 cells within the spleen and tumor-draining lymph nodes, with no additional effect at higher doses. Compared to MPLA alone, iAB-4T1 showed a splenic DC activation efficiency of 97.2% ± 1.2% (Fig. 1H and Figs. S3 and S4).
Then, we evaluated the therapeutic effect of iAB-4T1 in our in vivo tumor model. Eight days after 4T1-iRFP tumor injection on the left side of the mammary gland, iAB-4T1 was administered to the mice 5 times at 3-d intervals (Fig. 1I). Tumor growth was suppressed in iAB-4T1-treated mice compared to that in controls, with some mice showing complete remission (Fig. 1J and K and Fig. S5A). Mice administered iAB-4T1 survived longer than the control groups (Fig. 1L), and the tumor weight of mice administered iAB-4T1 was significantly lower than that of control mice (Fig. S5B).
Other metastatic cancer models, CT-26-iRFP and B16-iRFP, were also constructed to evaluate the anticancer effect of iABs. The in vivo cancer models were created by administering CT-26-iRFP cells to the rectum and left flank, and B16-iRFP melanoma cells to both flanks. When the left flank of the CT-26-iRFP tumor and the right flank of the B16-iRFP tumors reached approximately 100 mm^3^ in size, the tumors were excised, and iABs were generated. Specifically, ABs were produced by treating cancer cells obtained from CT-26-iRFP and B16-iRFP tumors with Cis or Dox (Tables S3 to S6 and Figs. S6 and S7), and MPLA was inserted into ABs to form iABs. Notably, mice treated with CT-26-iRFP tumor-generated iABs (iAB-CT26) showed suppression of CT-26-iRFP tumor growth in the rectal area compared to that of tumors in control mice (Fig. 1M to O and Fig. S8). Similarly, iABs from B16-iRFP tumor cells (iAB-B16) effectively inhibited the growth of metastatic B16-iRFP melanoma (Fig. S9).
To confirm the treatment mechanism of residual metastatic cancer by iAB-CT26, mesenteric lymph nodes (mLNs) were harvested from CT-26-iRFP tumor-bearing mice. CD4 and CD8 T cells in the mLNs of mice treated with iAB-CT26 secreted tumor necrosis factor-α and interferon-γ with high efficiency in response to the tumor antigens (Fig. 1P and Fig. S10). Production of cytotoxic mediators in iAB-CT26-treated mLN CD8 T cells was also increased by tumor antigens (Fig. 1Q and Fig. S11). Furthermore, CD44^+^CD62L^−^ memory CD4 and CD8 T cells exhibited an increasing trend, whereas Foxp3^+^ regulatory CD4 T cells tended to decrease after iAB-CT26 treatment (Fig. 1R and S and Figs. S12 and S13). To assess cytotoxic T lymphocyte activation, CD8 T cells were isolated from the mLNs of iAB-CT26-treated mice and co-cultured with CT-26-iRFP cells (Fig. 1T), revealing that iAB-CT26-treated CD8 T cells effectively attacked CT-26-iRFP cells within a short time, whereas other control CD8 T cells did not (Fig. 1U and Videos S1 to S4). Additionally, administration of iAB-CT26 did not induce hepatotoxicity or inflammation of peripheral tissues (Figs. S14 and S15). Together, these data demonstrate that iAB treatment could promote antigen-specific T cell immunity.
To advance clinical trials, effective methods for selectively removing metastatic cancer from patients and for the large-scale production and storage of iABs will be essential. Particularly, in cases of deep-seated tumors or micrometastatic lesions, it is necessary to consider tumor acquisition strategies and methods for sufficient production of iABs from those tumors. Despite these limitations, iABs could offer several advantages. By leveraging patient-derived tumor material, it may be less affected by tumor heterogeneity and adaptable for tailored metastatic cancer treatment. Moreover, iABs could reduce the cost and time required to identify neo-antigens for developing cancer vaccines. Furthermore, by generating antigen-specific immune responses, iABs might elicit fewer acute inflammatory reactions than nonspecific immune checkpoint blockade. Collectively, these features suggest that iABs could represent a promising personalized therapeutic approach for metastatic cancer.
Ethical Approval
All animal studies were performed with approval from the Institutional Animal Care and Use Committee of the ASAN Medical Center (approval no. 2023-04-041), and all experiments were performed after approval by a local ethics committee at the Laboratory Animals Center of ASAN Medical Center.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Oliveira G, Wu CJ. Dynamics and specificities of T cells in cancer immunotherapy. Nat Rev Cancer. 2023;23(5):295–316.37046001 10.1038/s 41568-023-00560-y PMC 10773171 · doi ↗ · pubmed ↗
- 2Liu Z, Chen J, Ren Y, Liu S, Ba Y, Zuo A, Luo P, Cheng Q, Xu H, Han X. Multi-stage mechanisms of tumor metastasis and therapeutic strategies. Signal Transduct Target Ther. 2024;9(1):270.39389953 10.1038/s 41392-024-01955-5PMC 11467208 · doi ↗ · pubmed ↗
- 3Butterfield LH, Najjar YG. Immunotherapy combination approaches: Mechanisms, biomarkers and clinical observations. Nat Rev Immunol. 2024;24(6):399–416.38057451 10.1038/s 41577-023-00973-8PMC 11460566 · doi ↗ · pubmed ↗
- 4Kim SJ, Park HB, An EK, Eom HY, Zhang W, Kim J, Choi I, Kwak M, Lee PCW, Jin J-O. Artificial immunogenic cell death lipid nanoparticle functions as a therapeutic vaccine for cancer. Adv Funct Mater. 2023;33(31):2302825.
- 5Gong N, Alameh M-G, El-Mayta R, Xue L, Weissman D, Mitchell MJ. Enhancing in situ cancer vaccines using delivery technologies. Nat Rev Drug Discov. 2024;23(8):607–625.38951662 10.1038/s 41573-024-00974-9 · doi ↗ · pubmed ↗
- 6Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020;17(7):395–417.32203277 10.1038/s 41571-020-0341-y PMC 8211386 · doi ↗ · pubmed ↗
- 7Miao X, Wu X, You W, He K, Chen C, Pathak JL, Zhang Q. Tailoring of apoptotic bodies for diagnostic and therapeutic applications: Advances, challenges, and prospects. J Transl Med. 2024;22(1):810.39218900 10.1186/s 12967-024-05451-w PMC 11367938 · doi ↗ · pubmed ↗
- 8Ravichandran KS. Find-me and eat-me signals in apoptotic cell clearance: Progress and conundrums. J Exp Med. 2010;207(9):1807–1817.20805564 10.1084/jem.20101157 PMC 2931173 · doi ↗ · pubmed ↗