Species diversity of Derxomyces (Bulleribasidiaceae, Tremellales) in China, with descriptions of two new species

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
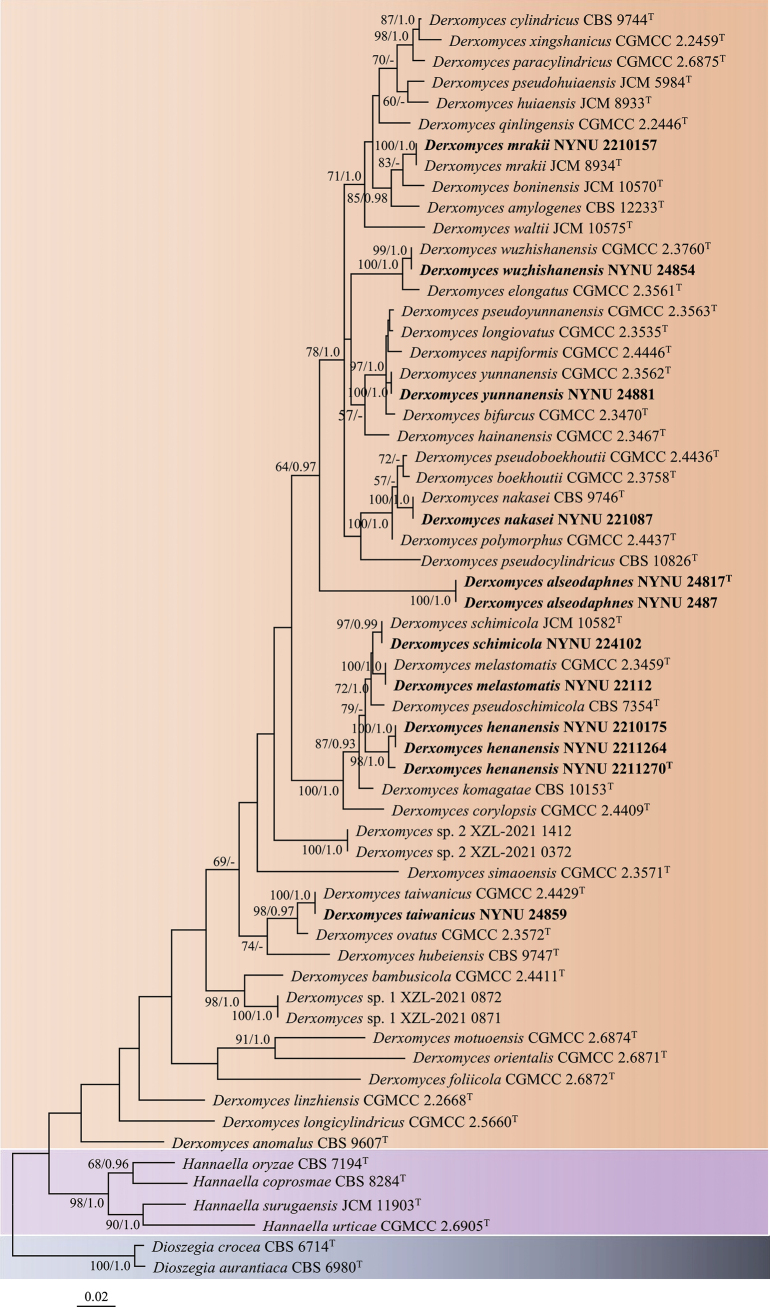 Figure 1
Figure 1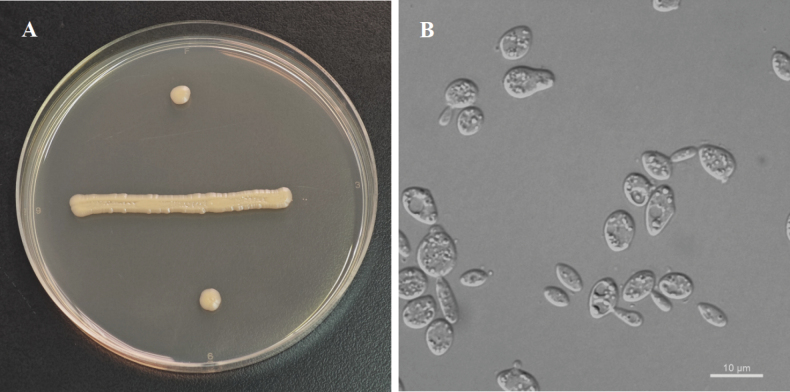 Figure 2
Figure 2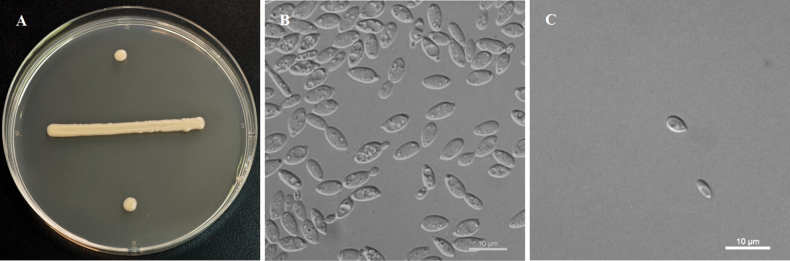 Figure 3
Figure 3| Species | Strain no. | Locality | GenBank accession no. | |
|---|---|---|---|---|
| ITS | LSU D1/D2 | |||
|
|
|
|
|
|
|
|
|
|
|
|
|
| CBS 12233T | China |
|
|
|
| CBS 9607T | China |
|
|
|
| CGMCC 2.4411T | China |
|
|
|
| CGMCC 2.3470T | China |
|
|
|
| CGMCC 2.3758T | China |
|
|
|
| JCM 10570T | Japan |
|
|
|
| CGMCC 2.4409T | China |
|
|
|
| CBS 9744T | China |
|
|
|
| CGMCC 2.3561T | China |
|
|
|
| CGMCC 2.6872T | China |
|
|
|
| CGMCC 2.3467T | China |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| JCM 8933T | New Zealand |
|
|
|
| CBS 9747T | China |
|
|
|
| CBS 10153T | Japan |
|
|
|
| CGMCC 2.5660T | China |
|
|
|
| CGMCC 2.3535T | China |
|
|
|
| CGMCC 2.2668T | China |
|
|
|
| CGMCC 2.3459T | China |
|
|
|
|
| China |
|
|
|
| CGMCC 2.6874T | China |
|
|
|
| JCM 8934T | New Zealand |
|
|
|
|
| China |
|
|
|
| CBS 9746T | China |
|
|
|
|
| China |
|
|
|
| CGMCC 2.4446T | China |
|
|
|
| CGMCC 2.6871T | China |
|
|
|
| CGMCC 2.3572T | China |
|
|
|
| CGMCC 2.6875T | China |
|
|
|
| CGMCC 2.4437T | China |
|
|
|
| JCM 5984T | Japan |
|
|
|
| CGMCC 2.4436T | China |
|
|
|
| CBS 7354T | Canada |
|
|
|
| CGMCC 2.3563T | China |
|
|
|
| CBS 10826T | China |
|
|
|
| CGMCC 2.2446T | China |
|
|
|
| CBS 9144T | China |
|
|
|
|
|
|
|
|
|
| CGMCC 2.3571T | China |
|
|
|
| CGMCC 2.4429T | China |
|
|
|
|
|
|
|
|
|
| JCM 10575T | Japan |
|
|
|
| CGMCC 2.3760T | China |
|
|
|
|
|
|
|
|
|
| CGMCC 2.2459T | China |
|
|
|
| CGMCC 2.3562T | China |
|
|
|
|
|
|
|
|
| 2 XZL-2021-1412 | China |
|
| |
| 2 XZL-2021-0372 | China |
|
| |
| 1 XZL-2021-0872 | China |
|
| |
| 1 XZL-2021-0871 | China |
|
| |
|
| CBS 6980T | China |
|
|
|
| CBS 6714T | UK |
|
|
|
| CBS 8284T | New Zealand |
|
|
|
| CBS 7194T | Japan |
|
|
|
| JCM 11903T | Japan |
|
|
|
| CGMCC 2.6905T | China |
|
|
| Characteristics | 1 | 2* | 3 | 4* | 5* | 6* |
|---|---|---|---|---|---|---|
| Assimilation | ||||||
| Inulin | + | – | – | – | + | – |
| Trehalose | + | + | – | + | + | + |
| Cellobiose | + | s | – | + | + | s |
| L-Sorbose | d | s, w | – | s | – | – |
| Ribitol | – | s | + | s | – | – |
| D-Mannitol | + | s | – | + | w | s |
| Myo-Inositol | w | + | – | + | + | s |
| Growth tests | ||||||
| Growth in vitamin-free medium | w | – | + | – | – | – |
| Growth at 30 °C | – | w | – | – | – | – |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSlime Mold and Myxomycetes Research · Mycorrhizal Fungi and Plant Interactions · Bryophyte Studies and Records
Introduction
The genus Derxomyces (Bulleribasidiaceae, Tremellales) was established by Wang and Bai (2008), with D. mrakii (Hamamoto & Nakase) F.-Y. Bai & Q.-M. Wang as its type species. Following its classification, Derxomyces has been extensively studied, with a substantial increase in the number of recognised species, based on phylogenetic analyses and phenotypic characteristics (Liu et al. 2012; Li et al. 2020; Jiang et al. 2024). As of now, 39 Derxomyces taxa are recognised, including 12 anamorphic species previously known as Bullera spp. (Hamamoto and Nakase 1996; Sugita et al. 1999; Bai et al. 2001, 2003; Wang et al. 2004).
The genus is morphologically characterised by cream or yellowish colonies, polar budding and a basal phylogenetic placement within Bulleribasidiaceae. Most Derxomyces species produce ballistoconidia, though some may also form hyphae or pseudohyphae (Wang and Bai 2008). Derxomyces species lack fermentative ability, possess Q-10 as a predominant ubiquinone and assimilate various carbon sources, but do not utilie hexadecane (Wang and Bai 2008; Li et al. 2020; Jiang et al. 2024).
Species of Derxomyces are widely distributed across the globe. The majority of these taxa were initially described from East Asia, with a total of 36 species (Wang and Bai 2008; Liu et al. 2012; Li et al. 2020; Jiang et al. 2024). Additionally, two taxa were first reported from Oceania (Hamamoto and Nakase 1996) and one species was described from North America (Bai et al. 2001). In contrast, research in other regions of the world has been limited. In recent years, 22 Derxomyces species have been documented in China, of which 16 were originally described from China, four from Japan, one from Canada and one from New Zealand (Li et al. 2020; Jiang et al. 2024). Despite these contributions, the full extent of Derxomyces species diversity in China remains to be explored. During our recent investigations across various locations in China, several Derxomyces strains were isolated. In this study, we employed molecular phylogenetic data and phenotypic characteristics to assess the species diversity of Derxomyces in China, identifying two previously undescribed species. These species are described and illustrated in detail below.
Materials and methods
Sample collection and yeast isolation
Senescent leaf samples were collected from Wuzhi Mountain in Hainan (18°19'N, 109°9'E) and Baotianman Nature Reserve (32°45'N, 113°30'E) in Hean, respectively. Yeast strains were isolated from the leaf surfaces using the ballistoconidia-fall method described by Toome et al. (2013). The senescent leaves were cut into small pieces (30–50 × 40–50 mm) and affixed to the inner lid of a Petri dish with a thin layer of petroleum jelly. The Petri dish contained yeast extract-malt extract (YM) agar medium (0.3% yeast extract, 0.3% malt extract, 0.5% peptone, 1% glucose and 2% agar), supplemented with 0.01% chloramphenicol to prevent bacterial growth. The plates were incubated at 20 °C and monitored daily for colony formation. Emerging yeast colonies were transferred on to fresh YM agar plates for further purification. Purified strains were suspended in 20% (v/v) glycerol and stored at −80 °C for long-term preservation.
Phenotypic characterisation
Morphological, physiological and biochemical characteristics were examined following standardised methods established by Kurtzman et al. (2011). Colony morphology was observed on YM agar after 7 days of incubation at 20 °C. Cell morphology was examined in YM broth after 3 days of incubation at 20 °C using a LEICA DM2500 microscope (LEICA, Wetzlar, Germany) with LAS V4.13 software. The ballistoconidium-forming activity of all new species was assessed using the inverted-plate method (do Carmo-Sousa and Phaff 1962) on corn meal agar (CMA; 2.5% corn meal infusion and 2% agar) at 20 °C. After 3 to 14 days, discharged spores were collected on a glass slide and examined microscopically. Potential sexual reproduction was investigated for individual strains and strain pairs on CMA, potato dextrose agar (PDA; 20% potato infusion, 2% glucose and 2% agar) and V8 agar (10% V8 juice and 2% agar) at 17 °C for up to two months, with observations made at two-week intervals (Li et al. 2020; Jiang et al. 2024). Glucose fermentation was tested in liquid medium using Durham fermentation tubes. Carbon and nitrogen assimilations were assessed in liquid media, with nitrogen assimilation tests performed using starved inoculum (Kurtzman et al. 2011). Growth at different temperatures (15, 20, 25, 30, 35 and 37 °C) was evaluated on YM agar plates. All new taxonomic descriptions and proposed names were deposited in the MycoBank database (http://www.mycobank.org; 10 November 2025).
DNA extraction, PCR amplification and sequencing
Genomic DNA was extracted from actively growing yeast cells cultured on YM agar using the Ezup Column Yeast Genomic DNA Purification Kit, following the manufacturer’s protocol (Sangon Biotech Co., Shanghai, China). Two nuclear loci were sequenced: the ITS region and the D1/D2 domain of the LSU rRNA gene, using primer pairs ITS1/ITS4 (White et al. 1990) and NL1/NL4 (Kurtzman and Robnett 1998), respectively. PCR amplification was carried out in a 25 µl reaction volume consisting of 1 µl of DNA template (20 ng/µl), 1 µl of each primer (10 µM), 12.5 µl of Taq 2X PCR Master Mix with blue dye (containing 0.05 u/µl Taq DNA polymerase, 4 mM MgCl_2_, 0.4 mM of each dNTP and reaction buffer; Sangon Biotech Co., Shanghai, China) and 9.5 µl of ddH_2_O. The PCR protocol included: an initial denaturation at 98 °C for 2 min, followed by 35 cycles of denaturation at 98 °C for 10 s, annealing at 55 °C for 10 s, elongation at 72 °C for 15 s and a final elongation at 72 °C for 5 min. PCR products were analysed by electrophoresis in 1% agarose gels. Sanger sequencing was performed by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China). The identity and accuracy of each sequence were confirmed by comparison with sequences in the GenBank database. Sequence assembly was conducted using BioEdit v.7.1.3.0 (Hall 1999). All newly-generated sequences were deposited in the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/).
Phylogenetic analyses
The sequences generated in this study, along with those obtained from previous work and deposited in the GenBank database, were used in our phylogenetic analyses (Table 1). Sequences of the two gene fragments (ITS and LSU) were aligned separately using MAFFT v.7.110 (Katoh and Standley 2013), followed by manual adjustments to remove ambiguous regions using BioEdit v.7.1.3.0 (Hall 1999). The aligned datasets were concatenated using PhyloSuite v.1.2.3 (Zhang et al. 2020).
Phylogenetic analysis, based on single LSU or ITS sequences, was performed using evolutionary distance data calculated from Kimura’s two-parameter model with the Neighbour-Joining (NJ) algorithm in MEGA v.7 (Kimura 1980; Kumar et al. 2016; Lachance 2022). Bootstrap analysis was conducted with 1,000 random re-samplings (Felsenstein 1985). Maximum Likelihood (ML) and Bayesian Inference (BI) analyses were performed on the concatenated LSU and ITS sequences using RAxML v.8.2.3 with 1,000 rapid bootstrap replicates (Stamatakis 2014) and MrBayes v.3.2.7a with 5,000,000 generations (Ronquist et al. 2012), respectively. The optimal nucleotide substitution model was determined using MrModelTest v.2.3 (Posada and Crandall 1998), with the GTR + I + G model selected for both the ML and BI analyses. Branches with bootstrap support (BS) values ≥ 50 and Bayesian posterior probabilities (BPPs) ≥ 0.95 were considered significantly supported in all phylogenetic trees constructed in this study.
Results
Phylogeny
In this study, 56 yeast strains were isolated from 22 leaf samples collected in Hainan and Henan Provinces, China. Based on ITS and LSU sequence analyses, the isolates were assigned to 14 previously described species — Bulleribasidium foliicola, Cystofilobasidium macerans, Derxomyces melastomatis, Derxomyces mrakii, Derxomyces nakasei, Derxomyces schimicola, Derxomyces taiwanicus, Derxomyces wuzhishanensis, Derxomyces yunnanensis, Hannaella phyllophila, Sporobolomyces bannaensis, Sporobolomyces roseus, Symmetrospora marina, Symmetrospora symmetrica — and two Derxomyces taxa that have not yet been formally described and, therefore, likely represent novel species.
Twelve strains preliminarily identified as Derxomyces were subjected to further analyses. The concatenated ITS and LSU dataset consisted of 122 sequences, including 61 ITS sequences and 61 LSU sequences, with 24 sequences newly generated in this study. These sequences represent 61 strains from 49 taxa, with Dioszegia aurantiaca L.B. Connell, Redman, R.J. Rodr. & Á. Fonseca and Dioszegia crocea (Buhagiar) M. Takash., T. Deák & Nakase serving as the outgroup species.
The phylogenetic trees inferred from both ML and BI analyses showed identical topologies. Therefore, only the tree generated from the ML analysis is presented, with BS ≥ 50% and BPPs ≥ 0.95 indicated on the branches (Fig. 1). The 12 isolates from China were clustered into nine distinct lineages within the Derxomyces clade, consistent with the single LSU or ITS dataset phylogeny (Suppl. materials 1, 2). Seven of these lineages corresponded to previously described species: D. melastomatis Q.M. Wang, F.Y. Bai & A.H. Li, D. mrakii (Hamamoto & Nakase) F.Y. Bai & Q.M. Wang, D. nakasei F.Y. Bai, Q.M. Wang & M. Takash. ex F.Y. Bai & Q.M., D. schimicola (Sugita, Gibas, M. Takash. & Nakase) F.Y. Bai & Q.M. Wang, D. taiwanicus Q.M. Wang, F.Y. Bai & A.H. Li, D. wuzhishanensis F.Y. Bai & Q.M. Wang and D. yunnanensis F.Y. Bai & Q.M. Wang. The remaining two lineages were genetically distinct from all known Derxomyces species, representing two new species within the Derxomyces clade.
Maximum Likelihood (ML) phylogram of Derxomyces inferred from the combined ITS and LSU sequences. The tree is rooted with Dioszegia aurantiaca and Dioszegia crocea as the outgroup. Bootstrap values ≥ 50 and Bayesian posterior probabilities ≥ 0.95 are indicated on the branches. Sequences generated in this study are shown in bold and ex-type strains are indicated with T after the strain number.
Two strains, NYNU 24817 and NYNU 2487, were isolated from the leaves of Alseodaphne rugosa and Polyalthia suberosa, respectively. Despite being isolated from different plant species, these strains shared identical D1/D2 and ITS sequences. These two strains clustered with D. hubeiensis with low support in the tree obtained from the ITS dataset (Suppl. material 2), but they formed a distinct branch in the trees of the LSU and combined ITS and LSU datasets (Fig. 1, Suppl. material 1). They differed from D. hubeiensis and other Derxomyces species by more than 13 nucleotide (nt) substitutions (2.2%) in the D1/D2 domain and 51 nt mismatches (8.7%) in the ITS region. These findings indicate that strains NYNU 24817 and NYNU 2487 represent a new Derxomyces species, for which the name Derxomyces alseodaphnes sp. nov. is proposed.
Three strains, NYNU 2211270, NYNU 2210175 and NYNU 2211264, were isolated from the same host plant, Corydalis balansae and exhibited similar sequences with three nucleotide differences in the D1/D2 domain and four in the ITS region, indicating conspecificity. These three strains, along with four other known species — D. komagatae, D. pseudoschimicola, D. melastomatis and D. schimicola — clustered together in a highly supported branch (Fig. 1, Suppl. materials 1, 2). They differed from the other species in this cluster by three to nine nt substitutions (0.5–1.5%) in the D1/D2 domain and 35–42 nt mismatches (6.2–7.8%) in the ITS region. The results suggest that these three strains represent a new Derxomyces species, for which the name Derxomyces henanensis sp. nov. is proposed.
Taxonomy
Derxomyces
alseodaphnes
Taxon classificationFungiTremellalesBulleribasidiaceae
C.Y. Chai & F.L. Hui sp. nov.
F905EF9B-9804-58F5-AB70-E5AEB8980C1D
861306
Etymology.
The specific epithet alseodaphnes refers to Alseodaphne, the plant genus from which the type strain was isolated.
Morphology of Derxomyces alseodaphnes (NYNU 24817). A. Colony on YM agar after 7 days at 20 °C; B. Budding cells in YM broth after 3 days at 20 °C.
Typus.
China • Hainan Prov.: Wuzhishan City, Wuzhi Mountain, on the phylloplane of Alseodaphne rugosa, August 2024, S.L. Lv, NYNU 24817 (holotype GDMCC 2.534^T^ preserved as a metabolically inactive state, culture ex-type JCM 38177).
Description.
On YM agar after 7 days at 20 °C, the streak culture is pale-yellow, butyrous and smooth, with an entire margin. After 3 days in YM broth at 20 °C, cells are ovoid and ellipsoidal, 2.1–4.0 × 2.6–4.9 μm and single, budding is polar. After 1 month at 20 °C, a ring and sediment are present. In Dalmau plate culture on CMA, pseudohyphae and hyphae are not formed. Sexual structures are not observed on PDA, CMA or V8 agar. Ballistoconidia are not produced. Glucose fermentation is absent. Glucose, inulin, sucrose, raffinose, melibiose, galactose, trehalose, maltose, melezitose, methyl-α-D-glucoside, cellobiose, salicin (weak), L-sorbose (delayed), L-rhamnose, D-xylose, L-arabinose, D-arabinose, 5-keto-D-gluconate, D-ribose (delayed), erythritol (delayed), galactitol (delayed), D-mannitol, D-glucitol (delayed), myo-inositol (weak), DL-lactate, succinate (weak), citrate (weak and delayed), D-gluconate, D-glucosamine, N-acetyl-D-glucosamine, 2-keto-D-gluconate, D-glucuronate and glucono-1,5-lactone (weak) are assimilated as sole carbon sources. Lactose, methanol, ethanol, glycerol and ribitol are not assimilated. Ethylamine and L-lysine are assimilated as sole nitrogen sources. Nitrate, nitrite and cadaverine are not assimilated. Growth is observed on YM agar at 25 °C, but not at 30 °C. Growth on 50% (w/w) glucose-yeast extract agar is negative. Growth in vitamin-free medium is positive. Starch-like substances are not produced. Urease activity is positive. Diazonium Blue B reaction is positive.
Additional strain examined.
China • Hainan Prov.: Wuzhishan City, Wuzhi Mountain, on the phylloplane of Polyalthia suberosa, August 2024, S.L. Lv, NYNU 2487.
GenBank accession numbers.
Holotype GDMCC 2.534^T^ (ITS: PQ568969, D1/D2: PQ568968); additional strain NYNU 2487 (ITS: PV404185, D1/D2: PV404184).
Note.
Physiologically, D. alseodaphnes can be distinguished from the closely-related species, D. hubeiensis, in its inability to assimilate ribitol and grow at 30 °C, while being able to assimilate inulin and to grow in vitamin-free medium (Table 2).
Derxomyces
henanensis
Taxon classificationFungiTremellalesBulleribasidiaceae
C.Y. Chai & F.L. Hui sp. nov.
C2EFEFFE-E0BA-5097-914C-D8FB6900E03D
861307
Etymology.
The specific epithet henanensis refers to the geographic origin of the type strain, Baotianman Nature Reserve, Nanyang City, Henan Province.
Morphology of Derxomyces henanensis (NYNU 2211270). A. Colony on YM agar after 3 days at 25 °C; B. Budding cells in YM broth after 3 days at 25 °C; C. Ballistoconidia on corn meal agar after 15 days at 20 °C.
Typus.
China • Henan Prov.: Nanyang City, Baotianman Nature Reserve, on the phylloplane of Corydalis balansae, August 2022, J.Z. Li, NYNU 2211270 (holotype GDMCC 2.336^T^ preserved as a metabolically inactive state, culture ex-type PYCC 9941).
Description.
On YM agar after 7 days at 20 °C, the streak culture is pale-yellow, butyrous and smooth, with an entire margin. After 3 days in YM broth at 20 °C, cells are ovoid and ellipsoidal, 3.2–3.8 × 6.3–8.5 μm and single, budding is polar. After 1 month at 20 °C, a ring and sediment are present. In Dalmau plate culture on CMA, pseudohyphae and hyphae are not formed. Ballistoconidia are produced on corn meal agar and are ovoid, 2.2–3.5 × 3.5–5.5 μm. Sexual structures are not observed on PDA, CMA or V8 agar. Glucose fermentation is absent. Glucose, sucrose (weak and delayed), raffinose (delayed), melibiose (weak and delayed), galactose (delayed), maltose (weak and delayed), melezitose (weak and delayed), methyl-α-D-glucoside (delayed), salicin (delayed), L-rhamnose (delayed), D-xylose, L-arabinose, D-arabinose, 5-keto-D-gluconate, D-ribose, ribitol, galactitol, D-glucitol (delayed), DL-lactate (delayed), succinate, citrate (weak and delayed), D-glucosamine (weak), N-acetyl-D-glucosamine (delayed), D-glucuronate (weak and delayed) and glucono-1,5-lactone (weak) are assimilated as sole carbon sources. Inulin, lactose, trehalose, cellobiose, L-sorbose, methanol, ethanol, glycerol, erythritol, D-mannitol, myo-inositol, D-gluconate and 2-keto-D-gluconate are not assimilated. L-Lysine is assimilated as the sole nitrogen source. Nitrate, nitrite, ethylamine and cadaverine are not assimilated. Growth is observed on YM agar at 25 °C, but not at 30 °C. Growth on 50% (w/w) glucose-yeast extract agar is negative. Growth in vitamin-free medium is positive. Starch-like substances are not produced. Urease activity is positive. Diazonium Blue B reaction is positive.
Additional strain examined.
China • Nanyang City, Baotianman Nature Reserve, on the phylloplane of Corydalis balansae, August 202, J.Z. Li, NYNU 2211270 and NYNU 2211264.
GenBank accession numbers.
Holotype GDMCC 2.336^T^ (ITS: OP954741, D1/D2: OP954742); additional strains NYNU 2210175 (ITS: OP954655, D1/D2: OP954654) and NYNU 2211264 (ITS: PV404187, D1/D2: PV404186).
Note.
Physiologically, D. henanensis can be distinguished from the closely-related species D. pseudoschimicola, D. melastomatis and D. komagatae in its inability to assimilate trehalose, cellobiose, D-mannitol and myo-Inositol, while being able to grow in vitamin-free medium (Table 2).
Discussion
The genus Derxomyces exhibits considerable taxonomic richness in China, making it one of the most well-studied genera within the family Bulleribasidiaceae in the country. To date, a total of 40 species have been recorded in China, including the previously reported 37 species and two new species described in this study: D. alseodaphnes and D. henanensis. Additionally, D. schimicola, a species new to China, was also discovered during this research. This study, therefore, adds two new species, six previously known species and one newly-recorded species in China, thus enriching the species diversity of the genus in the country. Moreover, sequence data from public databases suggest the presence of several unpublished strains, including XZL-2021-1412, XZL-2021-0372, XZL-2021-0872 and XZL-2021-0871 from China. These findings imply the potential existence of two additional Derxomyces species in China, warranting further investigation.
Species of Derxomyces are ballistoconidium-forming yeasts, as inferred from their original classification within Bullera (Wang and Bai 2008). Most species in this genus typically produce ballistoconidia, which are identified as an opaque mirror image of the culture formed by the discharged spores on the lid of an inverted Petri dish (do Carmo-Sousa and Phaff 1962; Kurtzman et al. 2011). However, the production of ballistoconidia is influenced by cultivation conditions and varies between clones (Nakase and Takashima 1993; Nakase 2000). In this study, D. henanensis sp. nov. produces ovoid ballistoconidia, whereas D. alseodaphnes sp. nov. does not. This phenomenon is exceptionally rare within the genus Derxomyces, with only D. pseudoboekhoutii previously reported as lacking the ability to produce ballistoconidia (Li et al. 2020).
Species of the genus Derxomyces are widely distributed in nature and are primarily regarded as epiphytic yeasts associated with plant leaves in tropical and subtropical regions (Wang and Bai 2008; Boekhout et al. 2011; Liu et al. 2012; Li et al. 2020; Jiang et al. 2024). In this study, the five strains representing two novel Derxomyces species share the common ecological niche of plant leaves, consistent with most previously described species in the genus. Derxomyces alseodaphnes was isolated from two different plant species, Alseodaphne rugosa and Polyalthia suberosa, collected on Wuzhi Mountain in Hainan Province. Derxomyces henanensis was repeatedly recovered from the same host plant, Corydalis balansae, collected on Baotianman Mountain in Henan Province. The new record species, D. schimicola, was exclusively isolated from Euonymus alatus collected on Baotianman Mountain in Henan Province. These findings highlight that Derxomyces species are commonly associated with tropical and subtropical plant leaves. Therefore, future studies on Derxomyces species diversity should give full consideration to the ecological significance of leaf habitats.
As a result of this study, the genus Derxomyces now comprises 41 recognised species. Previous research has shown that certain Derxomyces species play a significant role in food and medical fields. These species are known to produce carotenoids and astaxanthin and have been shown to regulate intestinal immune homeostasis in antibiotic-treated mice with diarrhoea (Kong et al. 2021). As a result, interest in these fungi extends beyond taxonomy to their ecological roles and potential applications in agriculture, industry and medicine, underscoring their economic significance.
Supplementary Material
XML Treatment for Derxomyces alseodaphnes
XML Treatment for Derxomyces henanensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bai FY, Takashima M, Nakase T (2001) Phylogenetic analysis of strains originally assigned to Bullera variabilis: Descriptions of Bullera pseudohuiaensis sp. nov., Bullera komagatae sp. nov. and Bullera pseudoschimicola sp. nov. International Journal of Systematic and Evolutionary Microbiology 51(6): 2177–2187. 10.1099/00207713-51-6-217711760961 · doi ↗ · pubmed ↗
- 2Bai FY, Takashima M, Zhao JH, Jia JH, Nakase T (2003) Bullera anomala sp. nov. and Bullera pseudovariabilis sp. nov., two new ballistoconidium-forming yeast species from Yunnan, China. Antonie van Leeuwenhoek 83(3): 257–263. 10.1023/a:102338231186512776921 · doi ↗ · pubmed ↗
- 3Boekhout T, Bai FY, Nakase T (2011) Bullera Derx (1930). In: Kurtzman CP, Fell JW, Boekhout T (Eds) The Yeasts – a Taxonomic Study, 5th edn, vol. 3. Amsterdam, Elsevier, 1623–1659. 10.1016/B 978-0-444-52149-1.00137-3 · doi ↗
- 4do Carmo-Sousa L, Phaff HJ (1962) An improved method for the detection of spore discharge in the Sporobolomycetaceae. Journal of Bacteriology 83(2): 434–435.10.1128/jb.83.2.434-435.1962 PMC 27775116561932 · doi ↗ · pubmed ↗
- 5Felsenstein J (1985) Confidence limits on phylogenies: An approach using the bootstrap. Evolution; International Journal of Organic Evolution 39(4): 783–791. 10.1111/j.1558-5646.1985.tb 00420.x 28561359 · doi ↗ · pubmed ↗
- 6Hamamoto M, Nakase T (1996) Ballistosporous yeasts found on the surface of plant materials collected in New Zealand. The genera Bensingtonia and Bullera with descriptions of five new species. Antonie van Leeuwenhoek 69(3): 279–291. 10.1007/BF 003996178740911 · doi ↗ · pubmed ↗
- 7Jiang YL, Bao WJ, Liu F, Wang GS, Yurkov AM, Ma Q, Hu ZD, Chen XH, Zhao WN, Li AH, Wang QM (2024) Proposal of one new family, seven new genera and seventy new basidiomycetous yeast species mostly isolated from Tibet and Yunnan provinces, China. Studies in Mycology 109: 57–153. 10.3114/sim.2024.109.02PMC 1166342839717653 · doi ↗ · pubmed ↗
- 8Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Molecular Biology and Evolution 30(4): 772–780. 10.1093/molbev/mst 010PMC 360331823329690 · doi ↗ · pubmed ↗