Characterization and antibiogram of bacterial isolates from diseased farmed Nile tilapia in Beheira governorate, Egypt
Merna M. A. Hassan, Riad H. Khalil, Mahmoud M. Abotaleb, Mahmoud T. Amer, Hany M. R. Abdel-Latif

TL;DR
This study identifies bacteria causing disease in farmed Nile tilapia in Egypt and examines their resistance to antibiotics.
Contribution
The study provides a detailed characterization of bacterial isolates and their antimicrobial resistance profiles in Nile tilapia farms in Egypt.
Findings
Six bacterial species were identified, with Streptococcus agalactiae being the most prevalent.
Aeromonas veronii showed resistance to all tested antimicrobials, while Enterococcus faecalis was sensitive to ciprofloxacin.
The study highlights the role of antibiotic misuse in promoting antimicrobial resistance in aquaculture.
Abstract
This study aimed to isolate, identify, and describe the bacteria isolated from mortality events occurred among pond-farmed Nile tilapia in different farms at Edku, Beheira province, Egypt, with special emphasis on their antimicrobial resistance profile. Specimens were collected from six private farms that experienced mortality outbreaks that occurred throughout the year between August 2023 and 2024. Clinical and postmortem examinations uncovered characteristic signs of bacterial septicemia. Forty-three bacterial isolates were recovered from the examined moribund and recently dead specimens. Based on the genetic data and evolutionary relationships derived from the 16S ribosomal RNA gene, six different isolates were resolved as Streptococcus agalactiae (46.5%), Vibrio alginolyticus (11.6%), Vibrio campbellii (14%), Vibrio owensii (9.3%), Aeromonas veronii (11.6%), and Enterococcus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1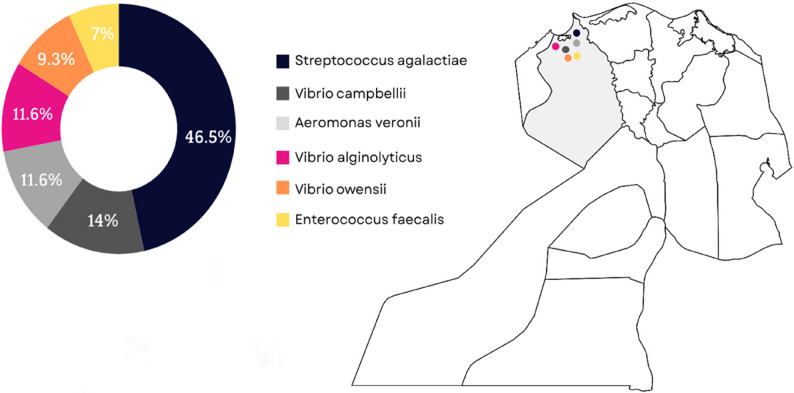 Figure 2
Figure 2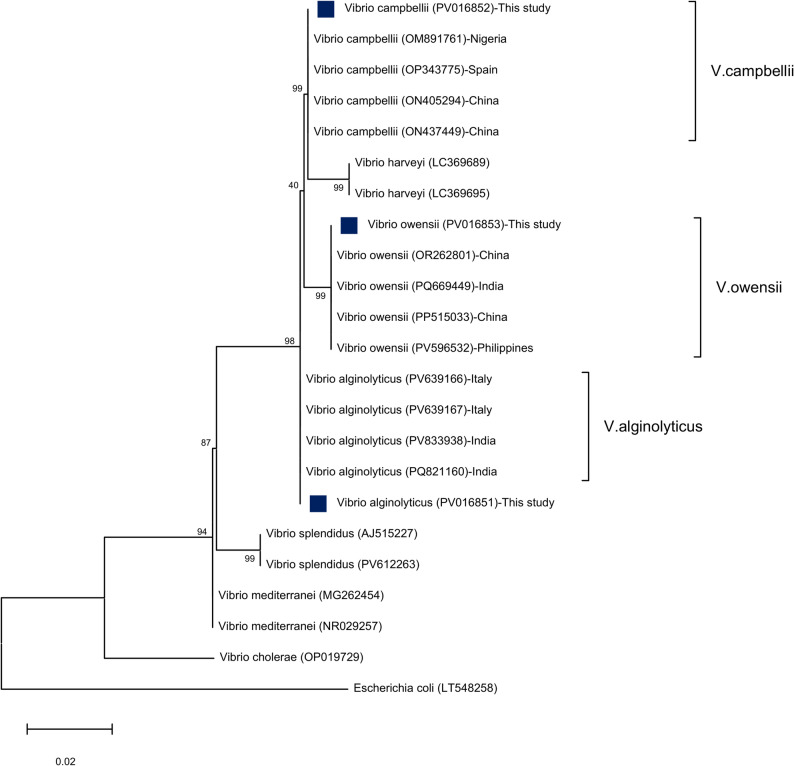 Figure 3
Figure 3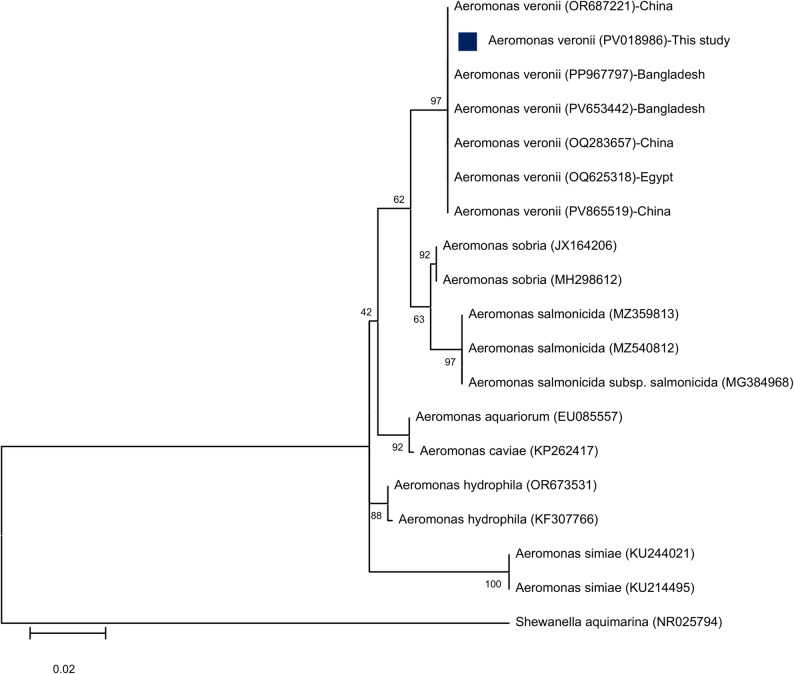 Figure 4
Figure 4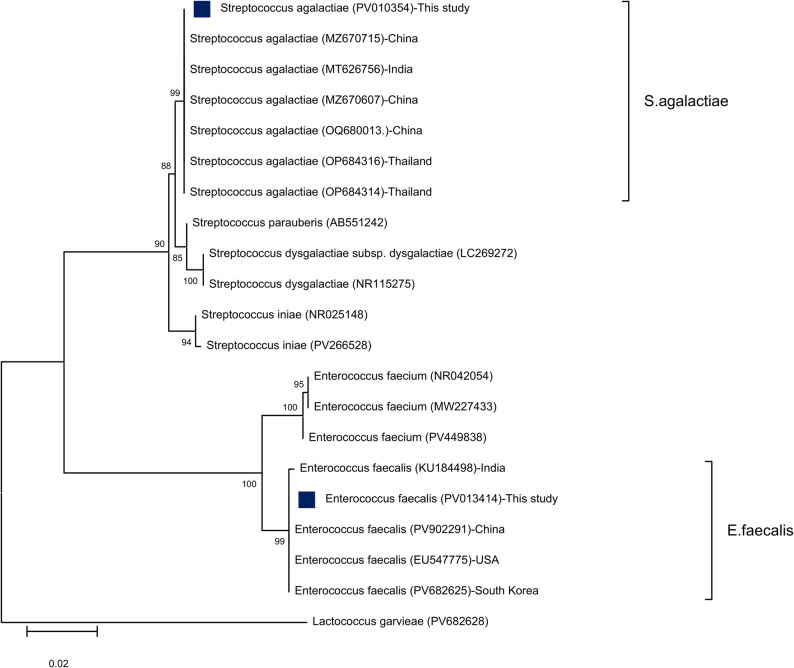 Figure 5
Figure 5- —Alexandria University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Vibrio bacteria research studies · Antibiotic Resistance in Bacteria
Introduction
Egypt is the largest aquaculture producer in Africa with a promise for expansion through transitioning from traditional to intensive farming systems. In Egypt, Nile tilapia represents the key farmed species that places it as one of the top countries in tilapia aquaculture production [1, 2]. Infectious diseases have been a long-standing challenge for the growth and sustainability of fish farming, especially of the bacterial origin resulting in huge economic losses [3–5]. Egyptian tilapia aquaculture has recently been hampered by a series of unusual mass mortalities, with a significant increase during the summer season [6]. Those authors surveyed 113 tilapia farms in several localities in Kafr El Sheikh, Beheira, Sharqia, and Faiyum governorates and declared that source and quality of water were the most significant contributing factors to both emergence and intensity of summer tilapia mortalities.
According to a recent review, it was found that a variety of bacterial infections have been identified as the cause of such frequent mortality events, which were linked to water temperature, high nutrient concentrations, and fish densities [7]. Zaheen et al. [8] also confirmed that the bacterial disease has been resulted from multi-factorial condition influenced by interaction between the bacteria, its host, and aquatic environment. Several other studies revealed that the prevalence of bacterial fish diseases is triggered by numerous stressors such as the water temperature over 28 °C, unionized ammonia exceed 0.2 mg/L, pH > 8.5, high stocking density and bad managemental practices [9]. Similarly, El-Son et al. [10] elucidated that poor water quality as decreased DO levels, elevated levels of unionized ammonia and nitrite as well as increased heavy metal contents were identified as stress triggers influencing fish health and increased their vulnerability to bacterial infections.
Bacterial fish pathogens have multiple virulence mechanisms that play a critical role in their pathogenesis [11]. Abdallah et al. [12] reported a mass mortality case of Streptococcus agalactiae infecting cultured O. niloticus in Assiut, Egypt. Huicab-Pech et al. [13] clarified that tilapia is susceptible to multiple pathogenic bacteria under stress conditions. Motile Aeromonas septicemia (MAS), Streptococcosis, and Vibriosis which are among bacterial diseases causing summer mortalities in tilapia farms in Egypt. A disease outbreak has also been reported by Aeromonas spp., Vibrio alginolyticus, and Enterococcus faecalis infection in Nile tilapia and African catfish in Manzala, Egypt [14, 15]. Those authors declared that inferior water quality such as high levels of ammonia, nitrite, and nitrate are the prominent cause of high mortalities. Abu-Elala et al. [16] identified three other bacterial pathogens including E. faecalis, S. agalactiae, and Lactococcus garvieae as the most significant pathogens that severely impact tilapia aquaculture. A study conducted by Elgohary et al. [17], found that E. faecalis and Aerococcus viridans have emerged as important fish diseases affecting Tilapia fish farms in El Sharkia and El Fayoum provinces. Likewise, Osman et al. [18] illustrated that Streptococcus sp., E. faecalis and L. garvieae were considered as new candidate bacterial pathogens in tilapia aquaculture.
Antibiotics are commonly used to manage and cure bacterial infections in fish [19]. Nevertheless, misusing these medications significantly drives the evolution of antibiotic-resistant bacteria, which can then pass their resistance genes to other bacterial types [20]. This ultimately reduces the effectiveness of these drugs for both aquatic life and human health [21, 22]. Therefore, the purpose of study to describe the clinical picture, and antimicrobial profile of the most common bacterial pathogens isolated from pond-reared Nile tilapia with health disorders in some Egyptian farms within Beheira governorate, Egypt.
Materials and methods
Sampling
Sixty adult diseased O. niloticus of different body weights (80 ± 20 g) and lengths were collected from six private farms located in different localities at Edku, Beheira province, Egypt throughout a year between August 2023 and 2024. Fish were stocked at a density of 20,000 to 25.000 fish per hectare. The main water supply of these farms is agricultural drainage from Edku. We observed poor biosecurity and managemental practices within the surveyed tilapia farms throughout the production cycle. Freshly dead samples only were taken and then were packed in an icebox while the moribund and alive fish were kept in well-aerated plastic bags. Clinical, necropsy, postmortem (PM) and bacteriological examinations were conducted using the methods outlined by Austin and Austin [23]. Fish samples showing septicemic signs were examined. The necropsy of the naturally infected tilapia fish was performed under completely sterile conditions.
Bacterial isolation and identification
Bacterial isolation
Before fish opening, the skin of each fish sample was wiped with 70% ethyl alcohol for surface disinfection. The liver, kidney, and spleen of the diseased tilapia were sampled for microbiological isolation by direct culturing onto Tryptic Soy Broth (TSB; Difco™) for pre-enrichment, incubated overnight at 30 °C. Sterile loopfuls were inoculated onto Tryptic Soya Agar (TSA; Difco™), Brain Heart Infusion Agar (BHIA; Difco™), and Thiosulphate Citrate Bile Salts Sucrose agar (TCBS; HiMedia™) followed by incubation at 30 °C for 24–48 h. The dominant bacterial colonies were selected and sub-cultured. After incubation, bacterial colonies were picked and streaked onto TSA supplemented with 5% sheep blood to examine the hemolytic activity. Consequently, the pure stock isolates were preserved at − 80 °C with 20% (v/v) glycerol for further biochemical and molecular examinations.
Morphological and biochemical characterizations
Gram staining, culture characteristics, hemolytic activity, motility, oxidase test, catalase test, and salt-tolerant test (NaCl 2%, 4%, 6%, 8%, and 10%) were performed to characterize the isolated bacteria. The results were recorded and interpreted based on the identification manual of bacteria from fish [24]. Successively, the VITEK examination was performed to evaluate the phenotypic characteristics of recovered isolates [25]. Pure bacterial colonies were streaked over TSA and incubated at 30 °C for 24 h to be identified by VITEK 2 compact analyzer (BioMérieux, France) following the manufacturer’s instructions.
Molecular identification and characterization
Molecular examination was carried out on six different randomly selected bacterial isolates. In brief, the bacterial DNA was extracted from 24 h growing bacterial isolates in TSB using QIAamp DNA mini kit, Catalogue No. 51,304, USA following the manufacturer’s protocol. Partial fragments of 16S rRNA gene were amplified using Aeromonas 16S rRNA (953 bp), Vibrio 16S rRNA (663 bp), S. agalactiae 16S rRNA (405 bp), and universal 16S bacterial primers (1485 bp) for S. agalactiae and other bacterial species (Table 1).
Table 1. Oligonucleotide primers that were used for bacterial identification in this studyBacterial speciesTarget genesProduct size (bp)Primer sequencesReferencesV. alginolyticus**Vibrio 16S rRNA663CGGTGAAATGCGTAGAGAT[26]V. campbelliiTTACTAGCGATTCCGAGTTCV. owensiiA. veroniiAeromonas 16S rRNA953CTACTTTTGCCGGCGAGCGG[27]TGATTCCCGAAGGCACTCCCE. faecalisBacterial 16S rRNA1485AGAGTTTGATCMTGGCTCAG[28]TACGGYTACCTTGTTACGACTTS. agalactiaeBacterial 16S rRNAS. agalactiae16S rRNA405CGCTGAGGTTTGGTGTTTACA[29]CACTCCTACCAACGTTCTTC
Preparation of PCR reaction mixture was performed using 2X Dream Taq Green master mix kit and the reactions were executed in a final volume of 25 µL in a DNA thermal cycler (T3 Thermal cycler, Biometra). The amplification conditions were detailed in our recently published research [1]. The PCR products were analyzed by electrophoresis in 1.5% agarose gel, stained and photographed by gel documentation system (Alpha Innotech). The PCR products were purified using QIA quick PCR Purification kit (QIAGEN, USA) according to the manufacturer’s protocol.
Virulence genes detection
The presence of key virulence genes in the genome of the six identified bacterial isolates was detected by PCR. V. alginolyticus was analyzed for thermostable direct hemolysin (tdh), and thermostable direct hemolysin-related hemolysin (trh). V. campbellii and V. owensii were examined for thermolabile hemolysin (tlh). A. veronii was tested for the presence of the genes encoding cytotoxic enterotoxins (act) and aerolysin (aerA). E. faecalis was tested for gelatinase (gelE), and cytolysin (cylA). S. agalactiae was screened for β-hemolysincytolysin (cylE), and hyaluronidase (hyl) to determine the relationship between the existence of these genes and the virulence of retrieved bacteria in cultured O. niloticus. Amplicon genomic DNA was extracted using a method analogous to the protocol established for bacterial identification. Primers used and PCR conditions are summarized in Table S2 (Supplementary Material).
Sequencing and phylogenetic analysis
The resulting purified DNA was subjected to sequencing using an Applied Biosystems 3130 automated DNA Sequencer (ABI 3130, USA). This was accomplished via a cycle sequencing reaction using the BigDye Terminator v3.1 ready reaction kit (Perkin-Elmer/Applied Biosystems, Cat. No. 4336817). The obtained 16S rRNA gene sequences from the bacterial isolates were then cross-referenced and confirmed. This identification process involved comparing the sequences’ homology against existing entries in the GenBank database utilizing the Basic Local Alignment Search Tool (BLAST analysis). The obtained sequential data was aligned by CLUSTAL W in MEGA 12.0 software. The aligned sequences were analyzed to suggest possible evolutionary relationships and to construct the phylogenetic tree via the maximum likelihood methods with 1000 bootstrapping test [30].
Antimicrobial susceptibility testing (AST)
Before testing, the Quality Control (QC) strains were used for ensuring the accuracy, reliability, and reproducibility of AST as suggested by CLSI (Clinical and Laboratory Standards Institute) [31]. The antibiogram was executed for the identified bacterial isolates to nine types of commercial antimicrobial discs, namely ampicillin (AMP) 10 µg, penicillin (P) 10 µg, amoxycillin/clavulanic acid (AMC) 20/10 µg, oxytetracycline (O) 30 µg, erythromycin (E) 15 µg, kanamycin (K) 30 µg, novobiocin (NV) 30 µg, ciprofloxacin (CIP) 5 µg, and trimethoprim/sulfamethoxazole (COT) 25 µg placed on Mueller-Hinton agar (Oxoid™) using the disc diffusion method. The findings were measured as inhibition zone (clear zone around the antibiotic disc) and interpreted based on susceptible (S), intermediate (I), and resistance (R) as previously described by CLSI [31].
Results
Clinical signs and necropsy findings
In this study, the diseased Nile tilapia generally exhibited lethargy, sluggish movement, anorexia, and certain cases showed erratic swimming. Most infected tilapia fish showed septicemic clinical signs. As shown in Fig. 1, the diseased fish showed several external signs including erythema and hemorrhagic patches over the abdomen, corneal opacity associated with hemorrhage at the caudal peduncle, unilateral exophthalmia and opaqueness on the eye associated with severe opercular hemorrhages, detached scales, abdominal dropsy with protruded vent, exophthalmia, dermal hemorrhages. The gross lesions appear as congested spleen, mottled liver and congested kidneys, and liver with accumulation of serous ascitic blood-tinged fluid in the abdominal cavity.
Fig. 1. Clinical signs and gross lesions of diseased Nile tilapia. Diseased fish showed several external signs including erythema (A; black arrow) and hemorrhagic patches over the abdomen (B; yellow arrow), corneal opacity (green arrow) associated with hemorrhage at the caudal peduncle (blue arrow), unilateral exophthalmia and opaqueness on the eye (yellow arrow) associated with severe opercular hemorrhages (black arrow) (C), detached scales (D; yellow arrow), abdominal dropsy, exophthalmia (green arrow), dermal hemorrhages (blue arrow) (E). The gross lesions appear as congested spleen (F; orange arrow), mottled liver and congested kidneys (G; white arrows), and congested liver (H; yellow arrow)
Phenotypic identification and biochemicals characterization
A total of 43 (n) bacterial isolates were retrieved and identified using conventional and biochemical tests, as shown in Table 2. The laboratory bacteriological examination detected three genera: Streptococcus spp., Vibrio spp., Aeromonas spp., and Enterococcus spp. Initially, Vibrio isolates produced characteristic yellow, green, or yellow-greenish colonies on their selective culture media (TCBS). On sheep blood agar, V. alginolyticus produced β-hemolytic colonies, while V. campbellii, V. owensii, and A. veronii showed no hemolysis. Moreover, E. faecalis showed α-hemolytic activity whereas S. agalactiae typically formed gray-white colonies with narrow zone of beta hemolysis.
Table 2. Microbiological and biochemical characteristics of the retrieved bacterial isolatesBacterial isolatesV. alginolyticus (n = 5)V. campbellii(n = 6)V. owensii (n = 4)A. veronii (n = 5)E. faecalis (n = 3)Str. agalactiae (n = 20)Gram stain-ve-ve-ve-ve+ve+veShapecurved rodscurved rodscurved rodsshort rodscoccicocciMotilityMotileMotileMotileMotileNon-motileNon-motileSwarming activity++++--Oxidase++++--Catalase++++--Hemolysis on blood agarβγγγαβTCBS mediumlarge convex, yellow-colored coloniesRound green coloniescircular yellowish-green coloniesNDNDNDGrowth at % NaCl 0%++++++ 2%++++++ 4%++++++ 6%++++++ 8%++++++ 10%------+ ve Positive, -ve Negative, ND Not Done, α Alpha hemolysis, β Beta hemolysis, γ Gamma hemolysis
The Gram stain grouped the isolates into (23) Gram-positive and (20) Gram-negative collected from moribund tilapia. The Vibrio spp. was Gram-negative, curved or comma-shaped rods, while A. veronii were Gram-negative, short, rod-shaped bacteria. On the other hand, E. faecalis were defined as Gram-positive cocci arranged in clusters, whereas S. agalactiae were Gram-positive cocci that organize into marked chains. All retrieved bacteria could survive in varying salt concentrations, up to 8% NaCl. For oxidase and catalase tests, Vibrio and Aeromonas spp. demonstrated positive results, while Enterococcus and Streptococcus spp. showed negative reactions.
Based on the results of VITEK examination, purified isolates were tested for different test parameters as detailed in Supplementary Material (Tables S2-S4). It was reported that some isolates belonged to either Vibrio and Aeromonads or Enterococcus spp. The tested V. alginolyticus were identified based upon positive reaction to glucose, maltose, and mannitol utilization tests. The isolates were typically negative for O/129 resistance, H_2_S production, citrate utilization, and urea hydrolysis. These results confirm the identity of V. alginolyticus, which is consistent with the expected phenotypic characteristics. VITEK identification only validated Aeromonas isolates at the genus level with 50% probability. Retrieved Aeromonas spp. produced positive reactions to glucose, maltose, and mannose utilizations, O/129 resistance, and β-galactosidase production. However, it tested negative for citrate utilization, urease, and H_2_S production. In addition, E. faecalis could utilize various sugars including galactose, mannose, maltose, ribose, and N-acetylglucosamine, while also detecting negative for raffinose and urease tests. The significant findings facilitate the differentiation of these isolates from other bacterial species.
Molecular characterization of isolated bacteria based on 16S rRNA sequence
Six isolates of three different Vibrio spp., one isolate of A. veronii, one isolate of S. agalactiae, and one isolate of E. faecalis were positive for 16S rRNA gene primers amplified the expected conserved region at 663, 953, 405, and 1485 bp, respectively.
According to the culture characters, biochemical, and molecular profile, bacterial isolates were identified as S. agalactiae (46.5%), V. alginolyticus (11.6%), V. campbellii (14%), V. owensii (9.3%), A. veronii (11.6%), and E. faecalis (7%) (Fig. 2). The presence of virulence-associated genes in the identified bacteria isolated from diseased tilapia samples is shown in Figures S1-S3 (Supplementary Material). V. alginolyticus strain harbored tdh and trh genes whereas, tlh gene was detected in V. campbellii and V. owensii isolates. A. veronii strain was positive for presence of act and aerA genes. E. faecalis strain was negative for cylA gene, while it possessed the gelE gene. In addition, cylE gene was identified in S. agalactiae strain, while it tested negative for hyl gene.
Fig. 2. Egypt map, grey zone is Edku, Beheira province from which diseased Nile tilapia were sampled with the prevalence percentage of isolated bacteria
Results of sequencing and phylogenetic analysis
The obtained nucleotide sequences showed identity with the following strains: V. alginolyticus (strain VA1), V. campbellii (strain VC2), V. owensii (strain VO3), A. veronii (strain AV2), E. faecalis (strain EF2), and S. agalactiae (strain SA1) deposited in GenBank under accession numbers; PV016851, PV016852, PV016853, PV018986, PV013414, and PV010354, respectively. The BLAST result of Vibrios, the 16S rRNA sequence of V. alginolyticus shared 100% identity with V. alginolyticus PQ821160 from India, PV833938 from India, PV639166 from Italy, and PV639167 from Italy. The retrieved sequence of V. campbellii showed 100% similarity with V. campbellii OM891761 from Nigeria and ON437449 from China. The similarity was 99.55% with V. campbellii OP343775 from Spain and 99.85% with V. campbellii ON405294 from China. Furthermore, V. owensii sequence showed 100% identity with V. owensii PQ669449 from India, OR262801 from China, PP515033 from China, and PV596532 from Philippines.
A maximum-likelihood based phylogenetic tree revealed that three representative Vibrio spp. combined with V. harveyi, V. splendidus, V. mediterranei, and V. cholera showing evolutionary relatedness with 99% bootstrapping values of V. campbellii and V. owensii in addition to 98% bootstrap value of V. alginolyticus (Fig. 3).
Fig. 3. Phylogenetic tree based on 16S rRNA partial sequences showing relationships of three isolated Vibrio strains; V. alginolyticus, V. campbellii, and V. owensii with other described closely related Vibrio species and out-group E. coli retrieved from GenBank database. Phylogenetic analyses were conducted with MEGA12.0, using the maximum likelihood method with the Jukes-Cantor model allowing sites to be evolutionary invariable (I). Percentage bootstrap values (1000 replicates) are shown at each branch point. The scale bar 0.02 represents substitutions per nucleotide position. The blue squared dots represent the studied isolates
Blast analysis of A. veronii sequence shared 100% identity with A. veronii OR687221 from China, PP967797 from Bangladesh, PV653442 from Bangladesh, OQ283657 from China, OQ625318 from Egypt, and PV865519 from China. Another phylogenetic tree was drawn using maximum-likelihood method showing that the A. veronii isolates were separated from other Aeromonas spp. including A. sobria, A. caviae, A. hydrophila, A. aquariorum, A. simiae, and A. salmonicida with a bootstrap value of 97% as described in (Fig. 4).
Fig. 4. The Phylogenetic tree showing relationship of 16S rRNA partial sequence of A. veronii and the 16S rRNA gene of other reported closely related Aeromonad species and with Shewanella aquimarina was selected as out-group from GenBank. Phylogenetic analyses were conducted with MEGA12.0, using the maximum likelihood method with the Hasegawa-Kishino-Yano model with Gamma distribution (G). Percentage bootstrap values (1000 replicates) are shown at each branch point. The scale bar 0.02 represents substitutions per nucleotide position. The blue squared dot represents the studied isolate
Similarly, the blast result of sequenced E. faecalis shared 100% identity with E. faecalis KU184498 from India. The similarity was 99.86% with E. faecalis PV902291 from China, EU547775 from USA, and PV682625 from South Korea. In addition, sequenced S. agalactiae revealed 100% identity with S. agalactiae MZ670715 from China, MT626756 from India, MZ670607 from China, OQ680013 from China, OP684316 from Thailand, and OP684314 from Thailand. A phylogenetic tree based on maximum likelihood showed two main lineages. The first clade included S. agalactiae isolate of this study accompanied by S. parauberis, S. dysgalactiae, and S. iniae with a 99% bootstrap value. The second subclade contained the isolated E. faecalis strain with other similar species, exhibiting a 99% bootstrap value (Fig. 5).
Fig. 5. Phylogenetic tree based on 16S rRNA partial sequences showing relationships of S. agalactaie and E. faecalis with other described closely related species and out-group Lactococcus garvieae retrieved from GenBank database. The Maximum likelihood method was used with the Kimura 2-parameter model with Invariant sites (I). The support level in percentage, after 1000 repetitions, is indicated next to each branch. The scale bar 0.02 represents substitutions per nucleotide position. The blue squared dots represent the studied isolates
Distribution and diversity of the bacterial isolates
According to Table 3, forty-three (43) bacterial isolates were retrieved from the liver, kidney, and spleen of naturally infected fish. From which, S. agalactiae (20) was the most prevalent bacterial species followed by V. campbellii (6), A. veronii (5), V. alginolyticus (5), V. owensii (4), and E. faecalis (3). The distribution of these bacteria in different tissues of infected fish was equally in liver and kidney (15) followed by spleen (13).
Table 3. Details of fish sampling from beheira province, EgyptDate of collectionCulture systemNo. of isolatesTissue samplesLiverKidneySpleenAugust 2023F1-CP9153October 2023F2-EP5311October 2023F3-EP6312October 2023F3-EP4211October 2023F4-CP5212November 2023F5-EP3210August 2024F6-EP11254Total643151513F Farm, EP Earthen Pond, CP Cement Pond
AST results
Most bacterial isolates showed resistance to most tested antibiotics as summarized in Table 4. Three Vibrionaceae isolates revealed complete resistance against ampicillin, penicillin, amoxycillin/clavulanic acid, erythromycin, kanamycin, novobiocin, and trimethoprim/sulfamethoxazole. Conversely, V. alginolyticus exhibited intermediate sensitivity to ciprofloxacin, whereas V. campbellii was moderately susceptible to oxytetracycline. The unexpected finding revealed that A. veronii was completely resistant to all antimicrobial agents. In contrast, E. faecalis was susceptible to ciprofloxacin, while this isolate was intermediate-sensitive to oxytetracycline, erythromycin, and kanamycin. Intermediate antibiotic susceptibility of S. agalactiae was only shown to amoxycillin/clavulanic acid, oxytetracycline, and ciprofloxacin.
Table 4. The AST of the retrieved bacterial isolates by disc diffusion assayThe tested antimicrobialsClassConc. (µg)Bacterial isolatesVA1VC2VO3AV2EF2SA1Ampicillin (AMP)Beta lactam100(R)0(R)0(R)0(R)10(R)0(R)Penicillin (P)Beta lactam100(R)0(R)0(R)0(R)8(R)0(R)Amoxycillin/Clavulanic acid (AMC)Beta lactam20/109(R)10(R)0(R)0(R)13(R)15(I)Oxytetracycline (O)Tetracycline308(R)21(I)0(R)0(R)20(I)25(I)Erythromycin (E)Macrolides150(R)0(R)0(R)0(R)14(I)0(R)Kanamycin (K)Aminoglycosides300(R)0(R)0(R)0(R)16(I)0(R)Novobiocin (NV)Aminocoumarin309(R)0(R)0(R)0(R)15(R)0(R)Ciprofloxacin (CIP)Fluoroquinolones518(I)0(R)0(R)0(R)23(S)20(I)Trimethoprim/Sulfamethoxazole (COT)Diaminopyrimidine + Sulfonamides250(R)0(R)0(R)0(R)0(R)0(R)Zone of inhibition measured = mmR Resistant, I Intermediate, S Sensitive(VA1): V. alginolyticus; (VC2): V. campbellii; (VO3): V. owensii; (AV2): A. veronii; (EF2): E. faecalis; (SA1): Str. agalactiae
Discussion
The present study depicted the bacterial pathogens encountered in heavy mortality events among pond-farmed Nile tilapia at Beheira Province, Egypt. The presumptive identification based on the clinical examination of the diseased fish revealed clinical signs like those reported in septicemic bacterial infections. These findings were like those found in previous reports in Nile tilapia [32, 33].
The PCR-based molecular analysis plays an important part in identifying new organisms or confirming morphological and biochemical features of fish-associated pathogens [34]. The 16S rRNA sequence testing confirmed the suspected bacterial isolates collected from moribund tilapia. Blast results and phylogenetic analysis based mainly on the 16S rRNA gene sequences of is important for diagnosis of the retrieved bacterial pathogens [10]. The 16S rRNA gene sequences of recovered isolates were almost identical to those of their related species on GenBank database, with a similarity greater than 99%. This gene facilitates classifying those bacteria into different genera, including Vibrionaceae, aeromonads, enterococci, and Streptococcus sp [35–38]. In our study, V. alginolyticus (PV016851), V. campbellii (PV016852), V. owensii (PV016853), A. veronii (PV018986), E. faecalis (PV013414), and S. agalactiae (PV010354) were identified and isolated from diseased tilapia.
A. veronii was primarily misidentified as A. sobria using VITEK2 with 50% probability, while the sequencing confirmed its identity. In this essence, Abdelsalam et al. [39] confirmed that the Vitek^®^ system faces certain limitations, including the significant manual effort required, a prolonged turnaround time, and an inability to identify several key bacterial pathogens relevant to the aquaculture industry. Moreover, Haung et al. [40] clarified that A. dhakensis was primarily misidentified as A. hydrophila via the VITEK-2 system, while its identity was confirmed by sequencing. Herein, the 16S rRNA partial gene sequence of recovered isolates were identical to those of their related species on GenBank database, with a similarity 100%, then has been accepted and published on GenBank under accession number PV018986. El Latif et al. [41] elucidated that 16S rRNA partial gene sequencing and phylogenetic analysis succeeded to verify the identity of A. veronii isolates.
S. agalactiae was the most predominant isolate among others during summer 2023 and 2024. This bacterium has emerged as a growing concern in aquaculture worldwide affecting a variety of fish species particularly those raised in warm water [42]. It causes streptococcosis, which can lead to high morbidity and mortality rates among the infected fish [43]. S. agalactiae has been frequently encountered in unusual mass mortality events that are reported among farmed tilapia during summer. These findings support the idea that summer outbreaks clarified by Abu-Elala et al. [16] in Kafr Elsheikh province and by Abdallah et al. [12] in Assiut province. Another survey confirmed that S. agalactiae was the third most prevalent pathogen following Aeromonas and Vibrio species causing massive mortalities of cultured tilapia during the summer months at Beheira, Fayoum, Ismailia, Kafr El Sheikh, and Sharqia provinces [14]. Nervous signs or erratic swimming, skin darkness, “pop eye” with hemorrhages and corneal opacity were the most common signs associated with S. agalactiae infection [44]. The cylE gene is highlighted as a significant virulence factor, contributing to the development of characteristic signs and lesions [45]. Furthermore, the detection of this gene within S. agalactiae isolates in Nile tilapia confirms its involvement in the pathogenesis of these streptococcal infections [46].
Vibrio sp. ranked the second most prevalent pathogen with a prevalence rate of 34.9% during autumn season. Vibriosis is primarily associated with brackish and marine aquaculture, however, the ability of Vibrio strains to infect farmed tilapia in freshwater and low-salinity environments has also been recorded [47, 48]. Our results resemble the previous study that investigated mortality outbreak in cultured tilapia caused by V. mimicus and V. cholerae during autumn season at Kafr El-Sheikh province [49]. Further study reported a case of co-infection between V. alginolyticus, Aeromonas sp., and E. faecalis in poly-cultured Nile tilapia and African catfish during early autumn at Port Said governorate [50]. V. alginolyticus and V. vulnificus were also identified in cultured O. niloticus in Al Fayoum governorate [51]. A study was conducted by Elgohary et al. [19] confirmed that the highest prevalence of V. vulnificus in autumn season is within tilapia farms.
V. campbellii and V. owensii were often related to acute hepatopancreatic necrosis disease affecting early phases of penaeid shrimp, leading to a decrease in their survival rates [52, 53]. However, a study investigated V. campbellii for the first time as a newly emerging pathogen affecting Egyptian farmed gilthead seabream [54]. The present work represents the first study to identify V. campbellii and V. owensii in cultured tilapia farms. This may be attributed to Vibrio species are characterized by high genome plasticity due to recombination, frequent mutation, and lateral gene transfer allowing Vibrio to adapt rapidly to environmental alterations [55]. Several studies highlighted the relationship between water quality parameters and epidemiology of Vibrio strains. Winfield [56] established a strong correlation between the incidence of Vibrio outbreaks and temperature fluctuations, particularly, in spring and fall seasons. Moreover, poor water quality measures and bad managemental practices exacerbated the Vibrio infection in tilapia, especially cultured in earthen ponds [21]. Anorexia, lethargy, red spots, dark skin, corneal opacity, exophthalmia, congestion in hematopoietic tissues as well as distended intestines were the typical Vibrio signs in infected fish [57]. Their ability to cause disease is related to a variety of virulence factors, especially hemolysins [58]. The presence of trh, tlh, and tdh genes in Vibrio spp. refers to their pathogenicity that might correspond with the potential of infections and diseases [59, 60].
A. veronii was the third most widespread bacterial pathogens isolated from farmed tilapia during autumn season. Disease and mortality caused by motile aeromonads in freshwater fish have been mainly linked with A. hydrophila whereas other species were probably ignored. Nonetheless, it has been attributed to various virulent Aeromonas sp., including A. veronii, A. caviae, A. sobria, A. jandaei and A. hydrophila [61–64]. A. veronii is ubiquitous in the aquatic environment that are often termed as fish pathogen causing epizootic ulcerative syndrome and hemorrhagic septicemia [65]. Egyptian fish farms have experienced frequent outbreaks of A. veronii infection during the summer season among farmed tilapia within different localities [41, 66, 67]. In accordance with the present findings, previous study has demonstrated that A. veronii can multiply faster and cause serious infections under adverse conditions during both summer and autumn seasons [68]. Ulcerations over the body surface, hemorrhage, and congestion in internal organs affecting the liver, kidney, and spleen were specific signs of Aeromonad infection [69]. Extracellular hydrolytic enzymes such as hemolysins, proteases, aerolysin, phospholipases, and cytotoxic enterotoxins might contribute to their disease progression in fish [2, 70].
E. faecalis has also been identified to cause hemorrhagic septicemia in Nile tilapia, with high mortality rates during late autumn. Repeated outbreaks in aquaculture worldwide, including Egypt, have been associated with E. faecalis [71–74]. Tilapia were highly susceptible to E. faecalis infection owing to sewage pollution in Egyptian fish farms or poultry manure which used as a natural fertilizer in ponds [75, 76]. Our findings agreed with those surveyed the natural outbreaks of E. faecalis infection in O. niloticus with the highest infection rate in both autumn and summer seasons at El Fayoum and the highest prevalence was recorded in the winter season at El Sharkia province [17]. It has been characterized to cause septicemic signs in tilapia fish such as skin hemorrhage, ulcers, dark discoloration, exophthalmia, eye opacity, abdominal ascites, pale or hemorrhagic liver, enlarged spleen, hemorrhagic kidney, and brain [77]. One of the important virulence determinants of E. faecalis is gelatinase toxin that hydrolyzes gelatin, hemoglobin, and collagen which is essential for the bacteria to cause disease [78, 79].
Stress is considered the major factor of the fish susceptibility to diseases [80]. The identified bacterial strains could be natural inhabitants of aquatic environment such as Vibrio and Aeromonas sp. or introduced as new pathogens for tilapia aquaculture such as Enterococcus and Streptococcus sp [18, 74, 81, 82]. Contaminated agriculture drainage water and untreated poultry manure used as organic fertilizer have participated in adding more pathogens to fish farms [83]. Most fish diseases caused by bacteria stem from secondary invaders targeting hosts that are already immune-compromised or subjected to environmental stress [84]. Poor water parameters, including a surplus of organic material or ammonia, elevated heat, and reduced oxygen levels, provide an optimal setting for these pathogens to thrive [85]. Adverse management practices such as handling or crowding, lack of biosecurity, intensive culture in the affected farms are potent risk factors impairing the defense mechanisms of tilapia fish and increasing the eruption and severity of those infections [39].
Regarding the antibiogram, the isolated strains showed a high degree of resistance to antibiotics most frequently used and prescribed in Egypt. All isolates exhibited complete resistance to penicillin, ampicillin, novobiocin, and trimethoprim/sulfamethoxazole. A remarkable resistance against kanamycin, erythromycin, and amoxycillin/clavulanic acid was detected in Vibrio sp. These results match those observed in Ayoub et al. [86], who explained that high antimicrobial resistance against ampicillin, amoxicillin, and erythromycin expressed by Vibrio, Aeromonas, and Pseudomonas sp. which recovered from diseased Nile tilapia. In the study, moderate antibiotic susceptibility was also found in V. alginolyticus to ciprofloxacin and V. campbellii to oxytetracycline. Moreover, A. veronii was found to be highly resistant to all tested antibiotics. Elgendy et al. [21] identified A. veronii in Egypt with antimicrobial resistance to ampicillin, amoxicillin, gentamicin, and neomycin. However, those authors clarified that all isolates were sensitive to ciprofloxacin, one isolate was sensitive to oxytetracycline, and one isolate showed moderate susceptibility to sulfamethoxazole/trimethoprim. The present findings seem to be consistent with dos Santos et al. [87], who investigated A. veronii in Brazil showing resistance to three antimicrobials: enrofloxacin, oxytetracycline, and amoxicillin, while the susceptibility only to florfenicol.
The antimicrobial sensitivity profile of S. agalactiae exhibited variable resistance to antibiotics. It expressed total resistance to erythromycin, and kanamycin, while the bacterial isolate was intermediately susceptible to ciprofloxacin, oxytetracycline, and amoxycillin/clavulanic acid. These findings agree with Abu-Elala et al. [16], who illustrated that Streptococcus, Enterococcus, and Lactococcus sp. displayed significant resistance to ampicillin, amoxicillin, oxytetracycline, tetracycline, and neomycin. However, these bacterial isolates showed high susceptibility to ciprofloxacin, gentamycin, and trimethoprim/sulfamethoxazole. Osman et al. [18] also identified Streptococcus spp. with abundant antibiotic resistance to ampicillin, penicillin, erythromycin, chloramphenicol, rifampicin, vancomycin, clindamycin, ofloxacin, and tetracycline. The antimicrobial sensitivity patterns in this study were only detected in E. faecalis strain to ciprofloxacin, whereas it displayed moderate susceptibility to oxytetracycline, erythromycin, and kanamycin. Hence, ciprofloxacin is effective against this bacterium and can be a drug of choice to treat affected fish. This also accords with Osman et al. [88], who demonstrated that all E. faecalis isolates were susceptible for penicillin, gentamicin, nitrofurantoin, and streptomycin. As well, most of the isolates were susceptible to ciprofloxacin, ampicillin, chloramphenicol, and vancomycin. In our study, E. faecalis was resistant only to amoxycillin/clavulanic acid. Arumugam et al. [89] confirmed that this isolate showed resistance to amoxiclav, oxytetracycline, ampicillin, kanamycin, erythromycin, gentamicin, nitrofurantoin, streptomycin, penicillin-G, and sulphafurazole and was intermediately resistant to ciprofloxacin, chloramphenicol, norfloxacin, clindamycin, and vancomycin.
The uncontrolled and indiscriminate application of antibiotics in fish farming is a major contributor to the emergence of drug-resistant bacteria [90, 91]. In Egypt, some antimicrobials usually prescribed without veterinarian to be use in aquaculture [92]. Some of those agents are used prophylactically in fish farming either to promote growth or to prevent the risk of disease incidences [93]. The long-term or misuse of these chemicals not only leaves residues but leads to the emergence of resistant bacteria [94]. The worrying findings of antimicrobial susceptibility analysis emphasize restricting the antibiotics used in aquaculture by developing drug dosage guides for antimicrobial treatment of diseased fish. Thus, it is urgently necessary to outlaw the preventative application of antibiotics in raising fish [95, 96].
Conclusion
In short, V. alginolyticus, V. campbellii, V. owensii, A. veronii, E. faecalis, and S. agalactiae were identified from heavy mortalities of pond-farmed Nile tilapia. These bacteria were isolated from fish with hemorrhagic septicemia and mortality cases among cultured O. niloticus in Beheira province. The findings highlighted that S. agalactiae was the most prevalent one during high temperatures of the summer season. V. campbellii and V. owensii have emerged as new candidate bacterial strains for tilapia aquaculture. The most concerning aspect is the existence of high resistance among the strains to most of the tested antimicrobial discs particularly, novobiocin, trimethoprim/sulfamethoxazole, penicillin, and ampicillin. Therefore, biosecurity plans and improving farming practices could be a better choice to reduce the occurrence of such infections in the affected farms. Controlling the haphazard use of antimicrobials is also crucial. Further research and monitoring should be implemented to develop effective treatment and vaccination for controlling bacterial infections as well as enhancing the sustainability and health of tilapia farming.
Supplementary Information
Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hassan MMA, Khalil RH, Abotaleb MM, Amer MT, Abdel-Latif HMR. Bacterial pathogens and their antimicrobial resistance in farmed nile tilapia experiencing summer mortality in Kafr El-Sheikh, Egypt. In: Microorganisms. 2025;13. 10.3390/microorganisms 13112448 PMC 1265439641304134 · doi ↗ · pubmed ↗
- 2Zaheen Z, War AF, Ali S, Yatoo AM, Ali MN, Ahmad SB, Rehman MU, Paray BA. Chap. 7 - Common bacterial infections affecting freshwater fish fauna and impact of pollution and water quality characteristics on bacterial pathogenicity. In: Bacterial Fish Diseases. edn. Edited by Dar GH, Bhat RA, Qadri H, Al-Ghamdy KM, Hakeem KR: Academic Press. 2022:133–154.
- 3Wang B, Mao C, Feng J, Li Y, Hu J, Jiang B, et al. A first report of Aeromonas veronii infection of the sea Bass, lateolabrax maculatus in China. Front Veterinary Sci. 2021;7–2020. 10.3389/fvets.2020.600587.10.3389/fvets.2020.600587 PMC 785597333553279 · doi ↗ · pubmed ↗
- 4Aboyadak I, Ali NG. Enrofloxacin, effective treatment of Pseudomonas aeruginosa and Enterococcus faecalis infection in Oreochromis niloticus. In: Microorganisms. 2024;12. 10.3390/microorganisms 12050901 PMC 1112403538792731 · doi ↗ · pubmed ↗
- 5dos Santos SB, Alarcon MF, Ballaben AS, Harakava R, Galetti R, Guimarães MC, Natori MM, Takahashi LS, Ildefonso R, Rozas-Serri M. First report of Aeromonas veronii as an emerging bacterial pathogen of farmed nile tilapia (Oreochromis niloticus) in Brazil. In: Pathogens 12; 2023.10.3390/pathogens 12081020 PMC 1045980537623980 · doi ↗ · pubmed ↗
- 6Osman KM, Ali MN, Radwan I, El Hofy F, Abed AH, Orabi A, et al. Dispersion of the Vancomycin resistance genes Van A and Van C of Enterococcus isolated from nile tilapia on retail sale: a public health hazard. Front Microbiol. 2016;7–2016. 10.3389/fmicb.2016.01354.10.3389/fmicb.2016.01354 PMC 499947927617012 · doi ↗ · pubmed ↗
- 7Yuan X, Lv Z, Zhang Z, Han Y, Liu Z, Zhang H. A review of antibiotics, antibiotic resistant Bacteria, and resistance genes in aquaculture: Occurrence, Contamination, and transmission. In: Toxics 11; 2023.10.3390/toxics 11050420 PMC 1022322737235235 · doi ↗ · pubmed ↗