Molecular detection of hemotropic mycoplasmas (hemoplasmas) in humans and dogs living on islands and the seashore mainland of Brazil: a One Health approach
Louise Bach Kmetiuk, Paul Shaw, Ashley Wallington, Jobin Jose Kattoor, Mathew Johnson, Rebecca P. Wilkes, Leandro Meneguelli Biondo, Rogério Giuffrida, Vamilton Alvares Santarém, Aaronson Ramathan Freitas, Ruana Renosto Delai, Claudia Turra Pimpão, Fabiano Borges Figueiredo

TL;DR
This study investigates Mycoplasma infections in humans and dogs on islands and coastal areas of Brazil, finding a higher risk in vulnerable populations.
Contribution
The study is the first to assess hemoplasma infections in oceanic island communities and their dogs in Brazil using qPCR and tNGS.
Findings
Mycoplasma haemocanis was confirmed in 1.0% of humans and 4.5% of dogs.
Candidatus Mycoplasma haematoparvum was detected in 7.9% of dogs.
Human infection was associated with gender and income, while dog infection was linked to gender and forest access.
Abstract
Although Mycoplasma spp. infection has been recently detected in other vulnerable human populations (indigenous and quilombola communities) in Brazil, no study to date has focused on traditional oceanic island communities and their dogs. To address this research gap, we assessed Mycoplasma spp. infection in humans and dogs living on the mainland seashore and oceanic islands of southern Brazil. Humans from three oceanic islands and two coastal mainland municipalities of southern Brazil were sampled, and Mycoplasma spp. infection was determined using quantitative polymerase chain reaction (qPCR) (cycle threshold; Ct ≤ 34.4). Dog samples were collected and tested using the Canine Hemotropic Mycoplasma panel (Idexx Reference Laboratory, Sacramento, CA, USA). To ensure accurate results, samples were also subjected to targeted next-generation sequencing (tNGS), and results were used to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
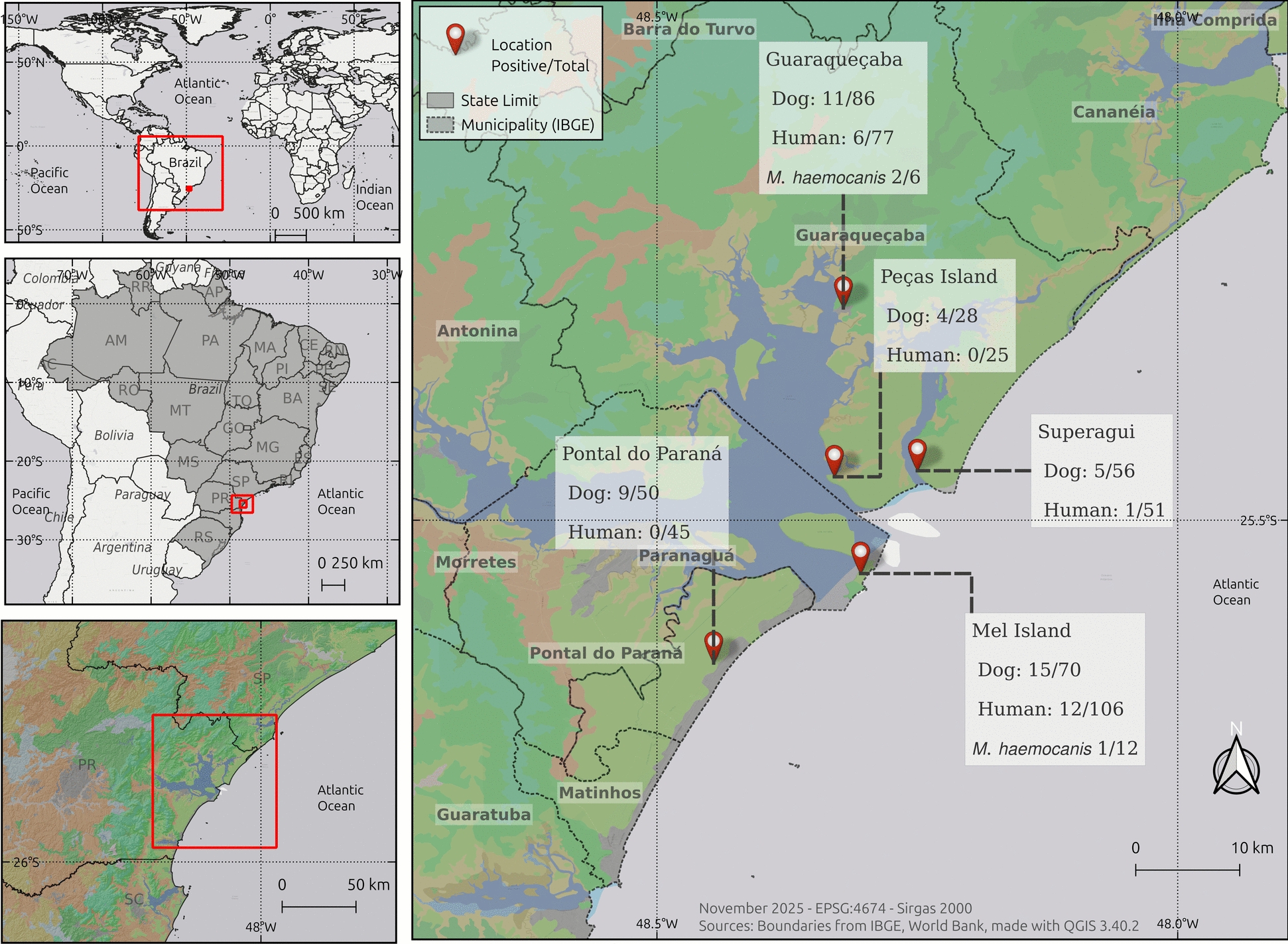 Figure 1
Figure 1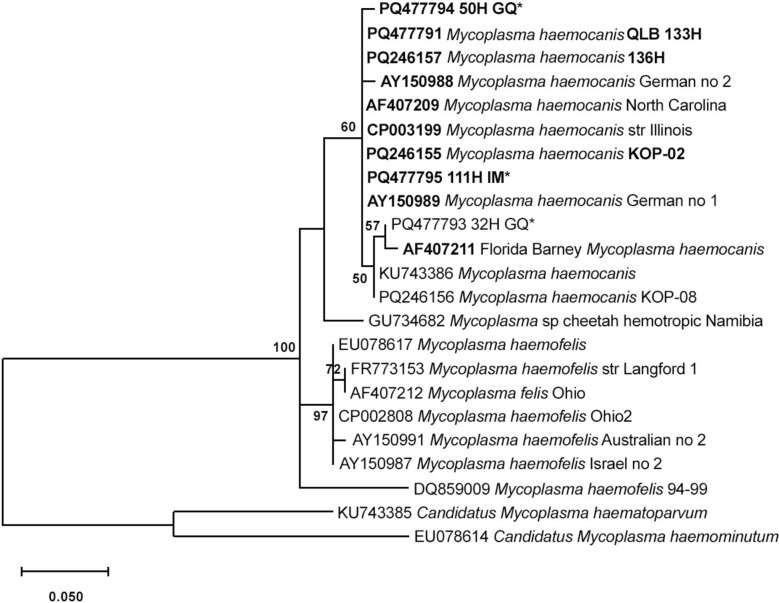 Figure 2
Figure 2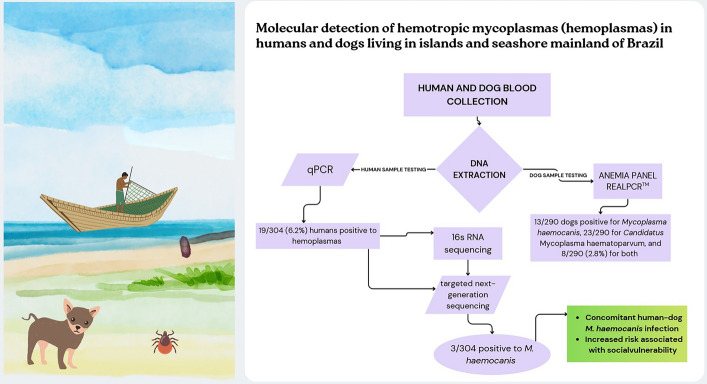 Figure 3
Figure 3- —https://doi.org/10.13039/501100015997Brazilian Biodivesity Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Bartonella species infections research · Toxoplasma gondii Research Studies
Background
Several species of hemotropic mycoplasma (hemoplasma) have been described in wildlife, domestic animals, and human beings, but the prevalence and transmission routes have not yet been determined [1]. Clinical signs of infection in humans and animals vary, but they usually include pyrexia, hemolytic anemia, lymphadenomegaly, and hepatosplenomegaly [2].
Previous reports have shown that humans can be infected with Candidatus Mycoplasma hominis, Mycoplasma ovis, Mycoplasma haemofelis, and Mycoplasma suis from sheep, cats, and pigs, respectively [3–6]. Infection is generally associated with occupational exposure, contact with animals, and natural areas, and veterinarians, travelers, farm workers, immunocompromised individuals, and hunters are mainly affected [3, 4, 7, 8]. In addition, Mycoplasma spp. has been detected in socially vulnerable individuals in close contact with positive dogs and potential vectors in natural areas [9, 10].
Island communities experience geographical isolation and limited access to health assistance, and they have a low responsiveness to public health challenges [11]. Appropriate sanitation, drinking water supply, and solid waste management practices are lacking within human and dog populations in island and seashore mainland areas within Brazil. Such populations also have contact with wildlife species in overlapped environmentally protected areas, and this may favor exposure to several zoonotic pathogens [12–17].
Island and seashore populations in southern Brazil are socially isolated from other populations, and they live closely with their dogs and other animals near natural areas that are home to wild animals potentially carrying vectors. However, no study to date has focused on human–dog infection with Mycoplasma spp. infection under such conditions. Therefore, we aimed to molecularly detect hemoplasmas in such populations.
Methods
Study design and area
A cross-sectional molecular survey was conducted. Sampling of humans and dogs living on three oceanic islands (Superagui Island, Mel Island, and Peças Island) and two seashore mainland municipalities of Paraná State (Guaraqueçaba and Pontal do Paraná), southern Brazil, was conducted from July 2019 to February 2020. The study areas were located within the most extended area of the Atlantic Forest biome of Brazil, overlapping different conservation units and protected areas [18]. Peças Island and Superagui Island comprise the Superagui National Park, which includes reefs, mangroves, and beaches and is home to endangered species [19]. Mel Island comprises two conservation units, an Ecological Station and a State Park, covering 81% and 12% of the total area, respectively [20]. Pontal do Paraná and Guaraqueçaba are two mainland cities that operate as access routes to these islands. Guaraqueçaba is a semi-isolated municipality located within three conservation units, and local access is difficult [19].
Blood collection
After providing voluntary signed consent, human blood samples were collected by cephalic puncture conducted by certified nurses of the City Secretary of Health of each location. To obtain dog blood samples, the owner signed consent and completed epidemiological questionnaire, and samples were collected through jugular puncture by certified veterinarians. The blood samples were placed in ethylenediaminetetraacetic acid (EDTA) anticoagulant tubes, aliquoted, and stored at −80 °C until processing.
Laboratory analysis
DNA was extracted from each human and dog blood sample using a commercially available kit (DNeasy Blood and Tissue Kit, QIAGEN, Hilden, Germany). Human samples were screened using a universal hemoplasma qPCR assay (SYBR™ Green PCR Master Mix, Applied Biosystems, Waltham, MA, USA) to detect hemoplasma 16S rDNA gene (∼ 200 bp), as already described [21]. Briefly, forward primer SYBR for 5′-AGC AAT RCC ATG TGA ACG ATG AA-3′ and the two reverse primers 5′-TGG CAC ATA GTT TGC TGT CAC TT-3′ and SYBR_Rev2 5′-GCT GGC ACA TAG TTA GCT GTC ACT-3′ in a concentration of 0.3 mM each were used for the reaction. The reaction mixture comprised 12.5 μL of 2 × SYBR green PCR master mix, 0.75 μL of each primer, 5 μL of DNA template, and 5 μL of RNAse-free water to entire volume of 25 μL. A previously established temperature was employed [22]. Two stored samples from dogs naturally infected with Mycoplasma haemocanis (GenBank accession no. CP003199) and two swine samples with Mycoplasma suis (GenBank accession no. ADWK01000001) were included as positive controls, and ultrapure water was used as the negative control. Positive samples were retested in triplicate and considered positive when the cycle threshold (Ct) was ≤ 34.4 [21]. The qPCR-positive samples were subjected to conventional PCR for nearly the entire 16S rRNA gene, using a commercial PCR mix (GoTaq Colorless Master Mix; Promega Corporation, Madison, WI, USA). The conventional PCR (cPCR) assay was conducted using a commercial mix (GoTaq Colorless Master Mix; Promega Corporation, Madison, WI, USA) following the manufacturer’s protocol. The cPCR proceeded using the forward primer 16S_HAEMOforw: 5′-GGC CCA TAT TCC T(AG)C GGG AAG-3′ and the reverse primer 16S_HAEMOrev: 5′-AC(AG) GGA TTA CTA GTG ATT CCA-3′, to amplify a DNA fragment of ∼966 bp. A total of 12.5 μL Master Mix, 1 μL of each primer (10 mM), 5 μL of nuclease-free water, and 5 μL of DNA sample were used. The thermoprofile was conducted using enzyme activation at 95 °C for 5 min, followed by 40 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 1 min, and extension at 72 °C for 1 min, followed by one cycle of final extension at 72 °C for 5 min. A DNA sample of Mycoplasma haemocanis (GenBank accession no. CP003199), not previously detected in humans, was used as a positive PCR control, and nuclease-free water was used as the negative control. Amplicons with an expected DNA band size of approximately 1500 bp were purified using a PCR Purification Kit (QIAquick, QIAGEN). All positive samples were sequenced using the WideSeq method at the Purdue University Genomics Core Facility [23]. To ensure accurate results, samples were subjected to targeted next-generation sequencing (tNGS) as previously described [24] at the Animal Disease Diagnostic Laboratory (ADDL), which is the official diagnostic laboratory for the State of Indiana, USA.
Sequences obtained for this project were submitted to GenBank, and these sequences were highlighted in the tree with their accession numbers. Additional sequences were obtained from GenBank (including Candidatus Mycoplasma spp., which were considered an outgroup) for comparison. Alignment was performed using MUSCLE in MEGA 12 [25]. The best DNA model for the tree was determined using Model Selection in MEGA 12. The phylogeny was inferred using the maximum likelihood method and the Hasegawa–Kishino–Yano model of nucleotide substitutions. The tree with the highest likelihood is shown, and the percentage of replicate trees in which the associated taxa clustered together (1000 replicates) is shown next to the branches. Clades ≥ 70 were considered significant. These analyses were conducted using MEGA 12.
To detect M. haemocanis and Candidatus M. haematoparvum, dog blood samples were tested at IDEXX Reference Laboratory, Sacramento, California, USA, using the Anemia Panel RealPCR™ Panel–Canine with Anemia Panel Lab 4Dx Plus Test–Canine (2907) (proprietary information, Idexx Reference Laboratory, Sacramento, CA, USA) The Anemia Panel RealPCR™ Panel-Canine provides positive or negative results for Mycoplasma haemocanis, Candidatus M. haematoparvum, and can detect other pathogens including Anaplasma spp., Babesia spp., Hepatozoon spp., Leptospira spp., Rickettsia spp., and Ehrlichia spp. [26]. Interferences for PCR test included poor sample quality, an aged sample, or recent antimicrobial therapy. The Anemia Panel Lab 4Dx provides positive or negative results for Dirofilaria immitis, Ehrlichia canis, Ehrlichia ewingii, Anaplasma phagocytophilum, Anaplasma platys, and Borrelia burgdorferi [26].
Statistical analysis
The overall and species-specific prevalence of Mycoplasma spp. was estimated with 95% confidence intervals (CI 95%). Univariate analyses were performed using Pearson’s chi-squared test or Fisher’s exact test to assess associations between a positive result for a dog and potential risk factors, including location (Guaraqueçaba, Peças Island, Mel Island, Pontal do Paraná, and Superagui), sex (female or male), mixed-breed status (yes or no), domicile (yes or no), access to the beach (yes or no), forest entry (yes or no), presence of ectoparasites (yes or no), use of flea and tick control (yes or no), and age category (puppy/juvenile: 0–3 years; young adult: 4–6 years; mature adult: 7–10 years; senior: ≥ 11 years).
The potential risk factors for human Mycoplasma spp. infection included locality (continental coast, island), age group in years (18–34, 35–43, 44–56, 56–93), gender (female, male), education level (higher education or more, completed secondary school, completed primary school, illiterate or not completed primary school), income (less than 1 minimum wage, 1–3 minimum wages, more than 3 minimum wages), native status (yes or no), number of dogs in household (no dogs, 1–2 dogs, 3–16 dogs), and tick bite history (yes or no).
Variables with P < 0.2 on univariate analyses for dogs and humans were considered eligible for inclusion in a multivariate logistic regression model. A backward stepwise selection procedure was applied to identify the model with the optimal explanatory capacity, on the basis of minimization of the Akaike information criterion (AIC).
Dogs or humans for which there were missing or incomplete data were excluded from the logistic final model, as the backward stepwise selection procedure required complete datasets to reliably estimate coefficients and perform model comparisons. Odds ratios (ORs) and their corresponding 95% confidence intervals were calculated for both univariate and multivariate models.
Model calibration was assessed using the Hosmer–Lemeshow goodness-of-fit test [27], while the discriminatory performance was evaluated by the area under the receiver operating characteristic curve (AUC) calculated by DeLong method. Collinearity among predictors was assessed by calculating the variance inflation factor (VIF). All statistical analyses were conducted in the blorr R package and using R internal functions [28, 29]. For all analyses, P < 0.05 was considered statistically significant.
Results
A total of 19/304 (6.25%; 95% CI: 4.04–9.55) humans were molecularly positive for hemoplasma infection by quantitative polymerase chain reaction (qPCR) (cycle threshold, Ct ≤ 34.4) (Fig. 1). The 16S ribosomal RNA gene sequencing confirmed Mycoplasma heamocanis infection in the 3 out of 304 (1.00%; 95% CI: 0.37–2.86), and M. haemocanis infection was also confirmed by tNGS in these three positive samples, two from Guaraqueçaba City and one from Mel Island (Fig. 2). Both RNase P and 16S gene sequences obtained from humans were submitted to GenBank (accession no. PQ477793, PQ477794, and PQ477795).Fig. 1. Positive samples to Mycoplasma spp. in seashore and islands of southern BrazilFig. 2Phylogenetic tree of amplicon sequences obtained by tNGS of 16S rRNA gene sequences from positive human individuals
The overall prevalence of Mycoplasma spp. in dogs was 44/290 (15.2%; 95% CI: 11.50–19.75). Among the hemoplasma species, M. haematoparvum was the most frequently detected, with a prevalence of 23/290 (7.9%; 95% CI: 5.34–11.62), followed by M. haemocanis at 13/290 (4.5%; 95% CI: 2.64–7.52), and coinfection with both species was identified in 8/290 (2.8%; 95% CI: 1.40–5.35) dogs.
On univariate analysis, the variables with P-values < 0.2 considered for inclusion in the logistic regression model were sex, domicile, forest entry, presence of ectoparasites, and age categories. However, owing to missing data for the age variable, 27 dogs were excluded from the multivariate analysis. The final model was obtained after following the two-step backward stepwise procedure, retaining sex, forest entry, and age category as significant predictors. The results of the univariate and multivariate analyses are presented in Table 1.Table 1. Associated risk factors for molecular Mycoplasma spp. detection in dogs of southern Brazil, Paraná (n = 290), by univariate and multivariate (logistic regression) statistical analysesVariableSerologyStatistical analysisPositiveNegativen (%)n (%)UnivariateMultivariate44 (15.2)246 (84.2)OR (95% CI)pOR (95% CI)pLocation0.345 Guaraqueçaba11 (25.0)75 (30.5)Ref Peças Island4 (9.09)24 (9.76)1.16 (0.29–3.82) Mel Island15 (34.1)55 (22.4)1.85 (0.79–4.46)Pontal do Paraná9 (20.5)41 (16.7)1.50 (0.55–3.96) Superagui5 (11.4)51 (20.7)0.68 (0.20–2.02)Sex0.048* Female15 (34.1)127 (51.6)RefRef Male29 (65.9)119 (48.4)2.05 (1.06–4.12)2.56 (1.19- 5.79)0.019Mixed-breed status0.477 No11 (25.0)78 (31.7)Ref Yes33 (75.0)168 (68.3)1.38 (0.68–3.01)Domicile0.050 No23 (52.3)87 (35.4)Ref Yes21 (47.7)159 (64.6)0.50 (0.26–0.96)Access to the beach0.514 No15 (34.1)100 (40.7)Ref Yes29 (65.9)146 (59.3)1.32 (0.68–2.65)Forest entry < 0.001 No10 (22.7)151 (61.4)RefRef Yes34 (77.3)95 (38.6)5.32 (2.59–11.9)5.96 (2.64–14.94) < 0.001*Presence of ectoparasites0.183 No8 (18.2)72 (29.3)Ref Yes36 (81.8)174 (70.7)1.83 (0.85–4.46)Use of flea and tick control0.605 No14 (32.6)67 (27.3)Ref Yes29 (67.4)178 (72.7)0.78 (0.39–1.61)Age group^a^0.016Puppy/juvenile: 0–3 years1 (2.70)37 (16.4)RefYoung adult: 4–6 years5 (13.5)32 (14.2)5.13 (0.74–141)3.60 (0.50–73.31)0.268Mature adult: 7–10 years16 (43.2)112 (49.6)4.65 (0.90–115)3.41(0.61–64.28)0.254Senior: ≥ 11 years15 (40.5)45 (19.9)10.8 (2.01–269)8.07 (1.40–153.99)0.055^a^ Missing data (27 dogs). Bold values indicate that the p-value lower than 0.05
The logistic regression model for dogs demonstrated good discriminative ability, with an area under the curve (AUC) of 0.777 (95% CI = 0.705–0.849), indicating that the model performed significantly better than chance in distinguishing between outcome categories. The Hosmer–Lemeshow goodness-of-fit test indicated no evidence of poor model fit (χ^2^ = 2.46, df = 8, P = 0.963). No significant VIF was detected for variables retained in final model (1.020 for sex, 1.0260 for forest entry, and 1.009 for age category).
The logistic regression coefficients and model parameters were gathered and are presented in Supplementary Tables S1 and S2 along with the receiver operating characteristic (ROC) curve in Supplementary Figs. S1 and S2.
The human variables with P < 0.2 included in the logistic model were gender and income range. However, owing to missing data for the income range, one human was excluded from the multivariate analysis. All predictors (gender and income range) were retained in the final model after conducting the backward selection procedure. The results of the univariate and multivariate analyses are presented in Table 2.Table 2. Associated risk factors for Mycoplasma spp. positivity in humans from seashore mainland and islands of southern Brazil (n = 290), by univariate and multivariate (logistic regression) statistical analysesVariableSerologyStatistical analysisPositiveNegativen (%)n (%)UnivariateMultivariate19 (6.2)285 (93.7)OR (95% CI)pOR (95% CI)pLocation0.587 Seashore mainland6 (31.6)116 (40.7)Ref Islands13 (68.4)169 (59.3)1.47 (0.56–4.35)Age group (years)0.217 18–347 (36.8)63 (22.1)Ref 35–433 (15.8)70 (24.6)0.40 (0.08–1.54) 44–567 (36.8)79 (27.7)0.80 (0.26–2.50) 56–932 (10.5)73 (25.6)0.26 (0.03–1.16)Gender0.027* Female7 (36.8)185 (64.9)RefRef Male12 (63.2)100 (35.1)3.13 (1.21–8.79)2.89 (1.11–8.08)0.033Education level0.482 Higher education or more4 (21.1)40 (14.0)Ref Complete secondary school8 (42.1)131 (46.0)0.60 (0.18–2.44) Complete primary school5 (26.3)53 (18.6)0.94 (0.23–4.18) Illiterate or incomplete primary school2 (10.5)61 (21.4)0.34 (0.04–1.95)Income^a^0.015* Less than 1 minimum wage8 (42.1)44 (15.5)Ref 1–3 minimum wages10 (52.6)227 (79.9)0.24 (0.09–0.68)0.27 (0.10–0.74)0.009 More than 3 minimum wages1 (5.26)13 (4.58)0.47 (0.02–3.05)0.46 (0.02–2.99)0.492Native status^a^0.602 No11 (57.9)189 (66.5)Ref Yes8 (42.1)95 (33.5)1.45 (0.54–3.75)Dog owner0.523 No dogs2 (10.5)29 (10.2)Ref 1–2 dogs15 (78.9)192 (67.4)1.07 (0.28–7.65) 3–16 dogs2 (10.5)64 (22.5)0.46 (0.05–4.57)Tick bite history^a^1.0 No13 (72.2)200 (70.2)Ref Yes5 (27.8)85 (29.8)0.92 (0.28–2.56)^a^ Missing data (one human); Ref. = reference category. Bold values indicate that the p-value lower than 0.05
The Hosmer–Lemeshow goodness-of-fit test yielded χ^2^ = 0.334 (df = 1, P = 0.563). Owing to the small number of events, only two predictive groups were formed, which limited the test’s power. However, the nonsignificant result suggested no evidence of miscalibration. The model showed good discrimination, with an AUC of 0.718 (95% CI: 0.607–0.830), and negligible multicollinearity, as indicated by the low variance inflation factors for gender and income range (1.001 each).
Discussion
To the authors’ knowledge, this is the first study of hemoplasma detection that concomitantly accessed humans and dogs living on oceanic islands and seashore mainland areas within natural areas and with close wildlife contact.
Previous studies conducted by our research group with this same population have shown that human and dog populations living in seashore areas are exposed to other vector borne diseases, including Brazilian spotted fever, Q fever, and Chagas disease [12, 16, 30]. In addition, the social vulnerability of this population also confirmed exposure to other zoonotic diseases such as Toxoplasma gondii, Toxocara spp., and Brucella spp. [14, 15, 17].
The disease prevalence in humans in this study (19/304, 6.2%) within seashore individuals was higher than that recorded of 23/644 (3.6%) indigenous individuals from ten communities in southern and southeastern Brazil [10] and 2/208 (5.8%) quilombola individuals from southern Brazil [9]. Another previous study conducted by our research group in rural and anthropized areas of southern Brazil showed no molecular positivity in 25 hunters, despite their close contact with 94/159 (59.1%) positive hunting dogs and 38/65 (58.5%) positive wild boars, along with tick exposure [22]. In addition, low human positivity was reported in 1/100 (1%) human from a rural settlement of southern Brazil [31]. Finally, the positivity determined in the current study was also higher than the 9/193 (4.6%) veterinarians, veterinary technicians, and partners of veterinary professionals who were in close contact with animals and vectors in the USA [8].
Some species of hemoplasma have been described in human case reports, including Mycoplasma suis, Mycoplasma ovis, Mycoplasma haemofelis, Candidatus M. haematoparvum, and recently Candidatus Mycoplasma haemohominis [4, 5, 7]. Human hemoplasma infections have been related to close contact with animals, immunocompromised conditions, and occupational exposure. In the present study, M. haemocanis was confirmed as the hemoplasma species involved in human infection of three individuals. Our research group previously diagnosed M. haemocanis in indigenous and quilombola individuals in Brazil [9, 10], and together with the present results, this reinforces the hypothesis that M. haemocanis occurs more often in individuals who are overexposed to and have close contact with infected dogs and who live under socially vulnerable conditions. Although Candidatus M. haematoparvum was not diagnosed in humans herein, this hemoplasma species was previously reported in a veterinarian (along with Anaplasma platys and Bartonella henselae) who presented with neurological signs such as seizure and headaches [7].
The positivity herein of 44/290 (15.2%) in dogs was similar to that of 19/100 (19.0%) dogs from quilombola communities in southern Brazil [9], 91/416 (21.9%) dogs from indigenous communities in southern and southeastern Brazil [10], and 78/437 (17.8%) household dogs from southeastern Brazil [32]. The results were lower than that of 94/159 (59.1%) hunting dogs [22] and 59/132 (44.7%) dogs from settlements, both from southern Brazil [22]. Although tick transmission has been suggested as the main transmission route [1], no statistical association was observed herein between Mycoplasma spp. positivity and the presence of vectors at the time of sampling. Hemoplasma transmission may occur in the absence of vectors by social contact or in transplacental form [33, 34], which may be exacerbated in island communities owing to the high number of unneutered dogs. The dogs in this study were infected by Candidatus M. haematoparvum (23/290; 7.9%) and M. heamocanis (13/290; 4.5%), the major hemoplasma species infecting dogs worldwide [6, 35–37]. In this study, male dogs were more likely to be positive for hemoplasmas, which has also been reported in dogs from indigenous communities in Brazil [10], hunting dogs in Italy [38], dogs from rural and urban areas in Chile [39], urban dogs in Iraq province [40], and dogs in northern Tanzania [41]. As already suggested, the higher exposure to hemoplasmas in male dogs herein may be the consequence of gender behavior due to aggressive territorial interactions and blood contact [42]. Access to forest areas was also found to be associated with hemoplasma infection in dogs in this study. The outdoor lifestyle and contact with wildlife can contribute to exposure to routes of possible hemoplasma transmission via ticks or through direct contact during fighting or wounds acquired during hunting [38, 43]. Similar to dogs, male humans were more likely to be positive to hemoplasmas in this study, and this could also be related to the gender behavior of hunting and associated access to forest areas. Mycoplasma spp. has been reported in both male and female patients [4, 5, 44]. Although female positivity has been reported in veterinarians, researchers, wildlife managers, and tourists, male positivity has been reported in other situations, such as in immunocompromised men, farmworkers, veterinarians, and insurance workers [4, 5, 8, 44]. Further populational studies should be conducted to better understand the association between hemoplasma infection and gender exposure, as well as with family income. In addition, further studies should compare Ct values and the results of full sequencing of positive samples between humans and dogs to determine whether the haplotypes are indeed the same.
The samples in this study were obtained and processed as follows: first, various professionals (nurses and veterinarians) obtained samples within the same workday. Human samples were separately processed, aliquoted, and stored (mostly owing to biosafety and handling regulations) away from dog samples. Second, DNA from human blood samples was extracted at the Oswaldo Cruz Foundation (Fiocruz), Brazil, and tested at the Clinical and Molecular Pathology Laboratory (CMPL) and the Animal Disease Diagnostic Laboratory (ADDL) in West Lafayette, IN, USA. Dog whole blood samples were shipped in dry ice from Brazil to USA for DNA extraction and molecular diagnostic testing at the Idexx Laboratories, Sacramento, CA. Third, each laboratory independently performed a different molecular approach method to detect and identify Mycoplasma species: qPCR at the CMPL and Wideseq sequencing tNGS at the ADDL, while both approaches were used at Purdue, and a vector-borne qPCR diagnostic panel was conducted at Idexx Laboratories. Thus, human–dog cross-contamination during sampling, sample handling, aliquoting, storage, shipment, DNA extraction, and molecular detection of nonviable DNA would have been very unlikely, if not impossible.
Finally, the subclinical or nonspecific clinical signs of hemoplasmosis in dogs and the lack of veterinarian assistance may contribute to the underdiagnosis of Mycoplasma spp. and its spillover in susceptible human populations. Therefore, to prevent the zoonotic transmission of diseases, veterinary preventive measures should be largely implemented, particularly in vulnerable populations, by employing anti-tick treatments, vaccines, and deworming. Although veterinary assistance in indigenous communities was established in 2025 by the Brazilian Ministry of Health as a consequence of our One Health research study [45], other vulnerable populations remain unassisted, including quilombola, inmates, the homeless, and traditional island and seashore communities.
Conclusions
The concomitant human–dog Mycoplasma haemocanis infection detected in communities living on oceanic islands and in remote seashore areas in Brazil, together with our previous reports of indigenous and quilombola communities, may suggest that socially vulnerable populations are at an increased risk of exposure to the disease. Future studies should be conducted to fully establish human–dog Mycoplasma spp. infection rates in other vulnerable communities worldwide, and transmission routes and the clinical presentations of M. haemocanis in humans should be further investigated and established.
Supplementary Information
Additional file1 (DOCX 50 kb)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hattori N, Kuroda M, Katano H, Takuma T, Ito T, Arai N, et al. Candidatus Mycoplasma haemohominis in Human, Japan. Emerg Infect Dis J CDC. 2020;26:11–19. 10.3201/eid 2601.19098310.3201/eid 2601.190983 PMC 692490631855136 · doi ↗ · pubmed ↗
- 2Santos AP dos, Santos RP dos, Biondo AW, Dora JM, Goldani LZ, Oliveira ST de, et al. Hemoplasma Infection in HIV-positive Patient, Brazil. Emerg Infect Dis J CDC. 2008;14:1922–4. 10.3201/eid 1412.08096410.3201/eid 1412.080964 PMC 263464919046522 · doi ↗ · pubmed ↗
- 3Mohd-Zahir NA, Mohd-Kamal FN, Sipin Q, Ammar-Qusyairi MH, Moktar MA, Low SY, et al. Detection of Mycoplasma haemocanis and Candidatus Mycoplasma haematoparvum in shelter and pet dogs in Malaysia. Trop Biomed. 2024;41:399–403. 10.47665/tb.41.S.00310.47665/tb.41.S.00339876495 · doi ↗ · pubmed ↗
- 4Coutinho - COORDENAÇÃO DE ELABORAÇÃO E REVISÃO DE PLANO DE MA.pdf. https://www.gov.br/icmbio/pt-br/assuntos/biodiversidade/unidade-de-conservacao/unidades-de-biomas/marinho/lista-de-ucs/parna-do-superagui/arquivos/plano_de_manejo_parna_do_superagui.pdf. Accessed 12 Jun 2025.
- 5Parque Estadual da Ilha do Mel (PEIM) [Internet]. Instituto Água e Terra. https://www.iat.pr.gov.br/Pagina/Parque-Estadual-da-Ilha-do-Mel-PEIM. Accessed 11 Jun 2025.
- 6Willi B, Meli ML, Lüthy R, Honegger H, Wengi N, Hoelzle LE, American Society for Microbiology, et al. Development and application of a universal hemoplasma screening assay based on the SYBR green PCR principle. J Clin Microbiol. 2009;47:4049–54. 10.1128/jcm.01478-09.10.1128/JCM.01478-09PMC 278668019828748 · doi ↗ · pubmed ↗
- 7Wide Seq [Internet]. https://genomics.rcac.purdue.edu/cgi-bin/index.cgi?service=Wide Seq&only=O. Accessed 22 Sept 2025.
- 8Real PCR Tests | IDEXX Reference Laboratories—IDEXX US [Internet]. https://www.idexx.com/en/veterinary/reference-laboratories/idexx-realpcr-tests/. Accessed 25 Nov 2025.