Colonization with Carbapenemase producing Enterobacterales (CPE) and associated alteration in microbiota composition in a tertiary care hospital in Egypt
Inas El-Defrawy, Manar Khaled, Amira El-Far, Ahmed El-Shenawy, Dalia Salem, Doaa Gamal, Nevine Fam, Hanan Ali Sayed, Noha N. Hayek, Mohamed A. Elrefaiy, Ahmed El Ray, Noha S. Soliman, May S. Soliman, Amani A. El-Kholy

TL;DR
This study explores how gut microbiota changes in patients colonized with antibiotic-resistant bacteria in an Egyptian hospital, highlighting risks and the need for infection control.
Contribution
The study identifies a link between CPE colonization and gut microbiota dysbiosis, particularly Proteobacteria dominance, in a hospital setting.
Findings
22.85% of patients were colonized with carbapenemase-producing Enterobacterales (CPE).
Proteobacteria were significantly more abundant in the gut microbiota of CPE carriers.
blaNDM and blaOXA-48 were the most common carbapenemase genes detected.
Abstract
The rising incidence of carbapenemase producing Enterobacterales (CPE)s, represents an urgent health threat due to their raised morbidity and mortality consequences. Colonization of CPE represents an important vehicle for hospital acquired infections. Microbiota dysbiosis, especially in critically ill patients, is a risk factor associated with CPE-colonized infections. In this study we aimed to characterize and assess the rate of colonization with CPE, and the possibility of subsequent infection. Also, to investigate the role of microbiota dysbiosis as a potential risk factor for colonization with CPE by comparing the gut niche in colonized patients versus non-colonized patients. Rectal swabs and stools were collected from 70 patients attending Hepatogastroentrology Department at TBRI Hospital on their first 72 h of admission and during their hospital stay. Incidence of infection with…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
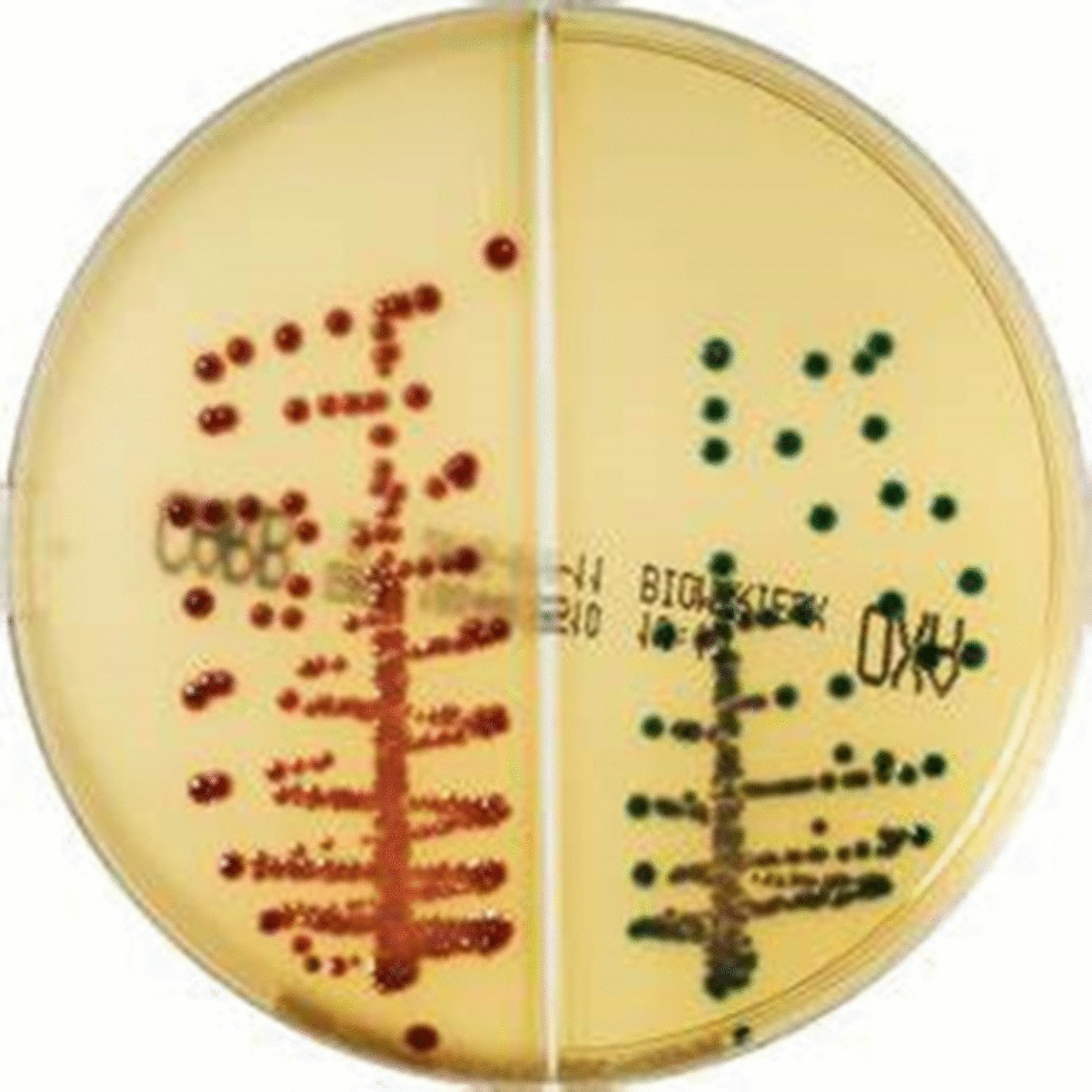 Figure 1
Figure 1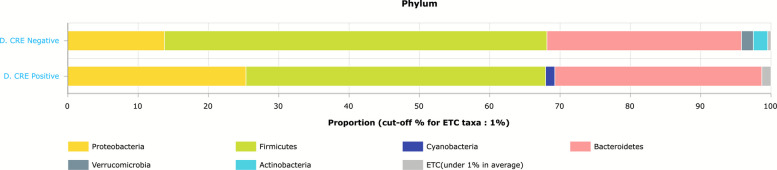 Figure 2
Figure 2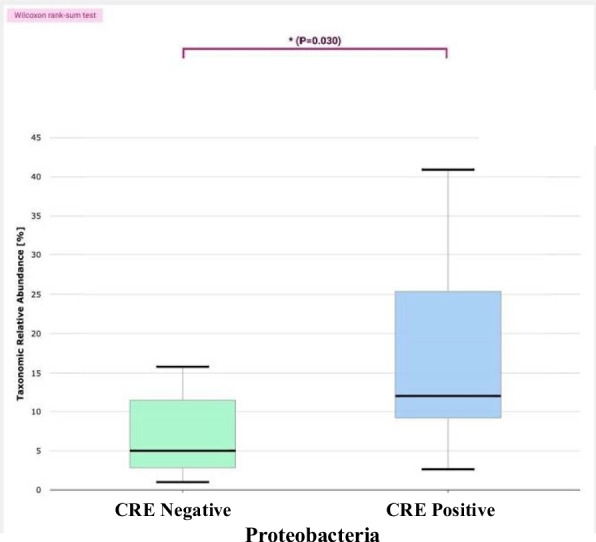 Figure 3
Figure 3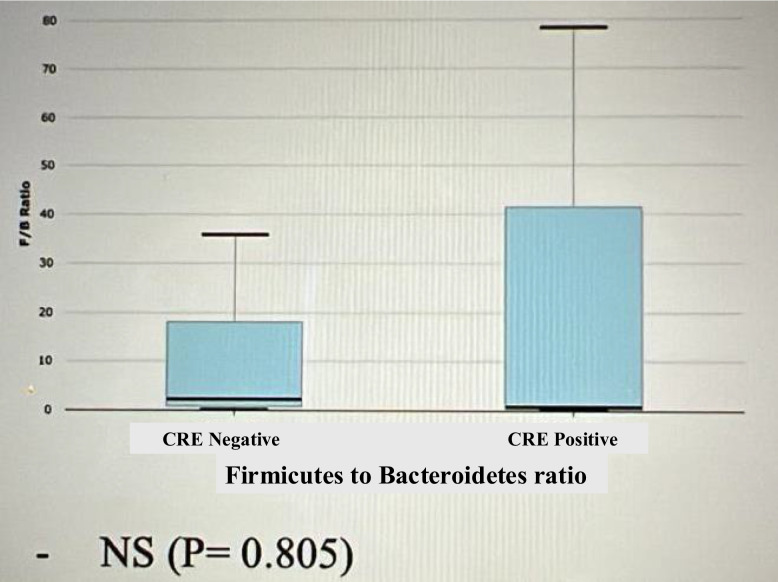 Figure 4
Figure 4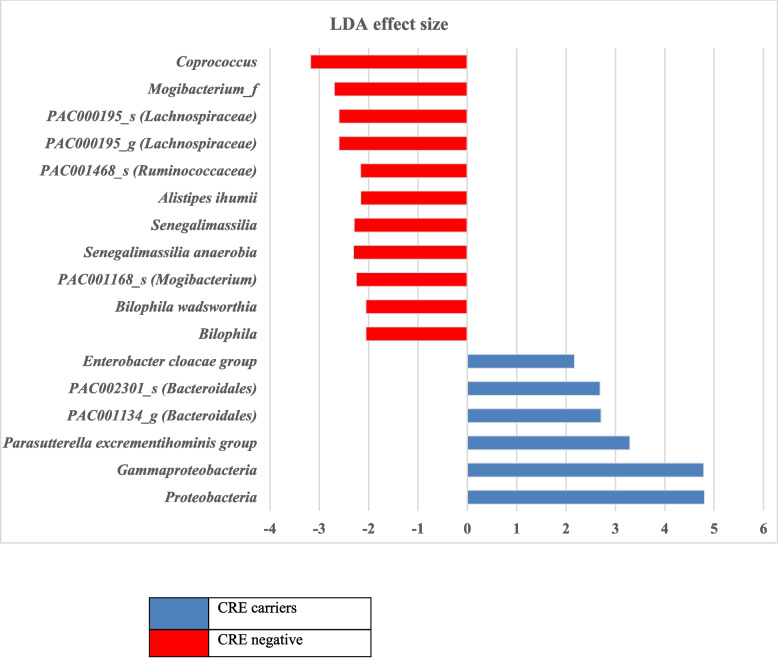 Figure 5
Figure 5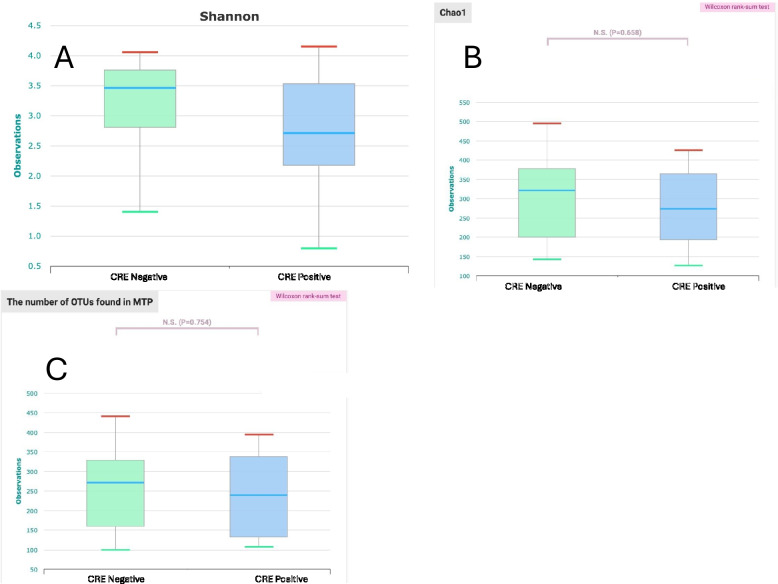 Figure 6
Figure 6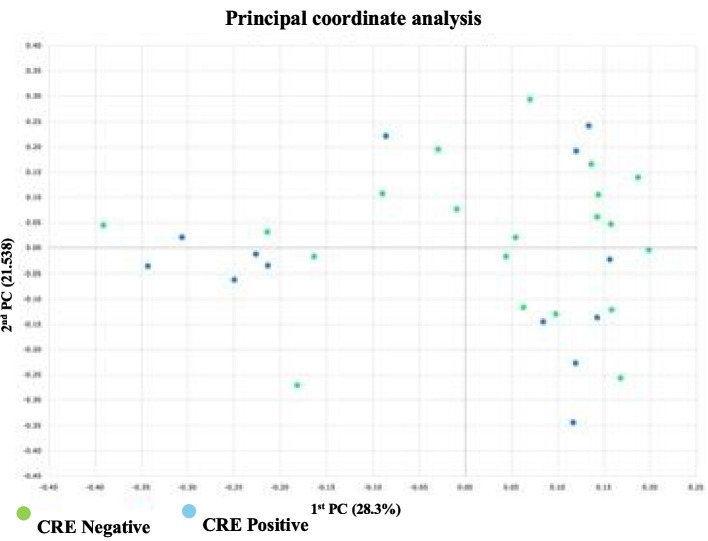 Figure 7
Figure 7- —Theodor Bilharz Research Institute
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Clostridium difficile and Clostridium perfringens research · Nosocomial Infections in ICU
Background
Carbapenems are considered as the last therapeutic options for treatment of multidrug resistant (MDR) Gram-negative bacterial infections [1]. The rising incidence of carbapenem-resistant Enterobacterales (CRE), usually in MDR pathogens, represents an urgent health threat due to their morbidity and mortality consequences that could exceed 50% in critically ill patients. Thus, the World Health Organization (WHO) reported CRE as critical priority pathogens highlighting the urgency for active surveillance and prompt containment strategies [2]. Carbapenemase production is the main mechanism of resistance in CRE. Other mechanisms of resistance include overproduction of AmpC or extended-spectrum β-lactamase (ESBL) in combination with impermeability of outer membrane porin and active efflux mechanisms [3]. While CRE are known for their therapeutic challenges, carbapenemase producing Enterobacterales (CPE)s, defined with a detectable carbapenemase gene, pose a significant threat for both treatment and infection control, as carbapenemase genes are usually carried on transferrable plasmids leading to hospital outbreaks [4]. Carbapenemase production usually results in clinically relevant levels of carbapenem resistance, however it may only result in reduced susceptibility that does not reach the susceptibility breakpoints. That makes the detection of CPEs more challenging and of paramount importance [1, 2, 4].
According to Ambler classification of beta lactamases, carbapenemases are located into the three classes (A, B, and D). Among these, KPC (class A), NDM, VIM and IMP (class B; metallo-beta-lactamases (MBL)s), and OXA-48-like (class D) are the most clinically significant carbapenemases in Enterobacterales [2]. KPC and OXA-48 both utilize serine residues in their activity [4].
Colonization of CPE represents an important precondition for subsequent hospital acquired infections whether among healthcare workers or patients**.** There are multiple risk factors associated with CPE colonized infections, such as prior antimicrobial exposures, prolong hospitalization, comorbidities, the presence of invasive medical devices, as well as microbiota dysbiosis especially in critically ill patients [2, 5, 6]. The abundant phyla in the gut microbiome normally are Firmicutes (60–75%), Bacteroidetes (30–40%), Actinobacteria, and Proteobacteria. This normal microbiota composition protects against the enteric pathogens, including MDRs, and reduce the colonization pressure. However, microbiota disrupting exposures such as prior antibiotics intake compromises colonization resistance, thus facilitating CRE persistence and overgrowth [7]. The 16S rRNA profiling has been used for identification of gut microbiota yielding sufficient insight about their composition and function [5].
In this study we aimed to characterize and assess the rate of community and hospital acquired colonization with CPE, and the possibility of subsequent infection. Also, to investigate the role of microbiota dysbiosis as a potential risk factor for colonization with CPE by comparing the gut niche in colonized patients versus non-colonized patients.
Methods
Patients and specimens included
Rectal swabs were collected from 70 patients attending Hepatogastroentrology Department at TBRI Hospital on their first 72 h of admission.
History taken from participants included education background, comorbidities (diabetes mellitus, hypertension, chronic liver disease and previous history of inflammatory bowel disease), the usage of medical devices, previous hospitalization and previous administration of antibiotics.
CPE cases were defined as adult patients with CPE intestinal colonization identified by rectal swab screening culture within 72 h of admission to the TBRI hospital. The patients who were found negative at admission were monitored by rectal swabs and stool specimens during their hospital stay by repeating the specimens on weekly basis until turned positive or being discharged from the hospital. DNA extraction from fecal specimens obtained from patients was performed for later 16S rRNA sequencing.
As reported in previous studies [8, 9], community and hospital colonization were categorized according to the timing of positive rectal swabs, so colonization detected later in patients whose admission screening was negative for CPE was classified as hospital colonization.
Incidence of infection with CPE was followed up in the same patients with relevant clinical specimens, mainly urine culture and sensitivity.
Patients were excluded from the study participation if they were under 18 years old, had a history of travelling outside Egypt in the past three months, suffered from acute bacterial or viral or active inflammatory bowel disease within 14 days before sample collection, fecal incontinence device or colostomy at the time of sample collection. These exclusion criteria were applied to reduce potential confounding effects of colonic inflammation and ensure that samples will be representative of recently passed stool. Patients participated in the study only for once.
Microbiological cultural methods
Rectal swabs were screened for CPE by culture on CHROM ID CARBA SMART agar (BioMérieux, France). This is a ready-made biseptate chromogenic agar plate (CARB and OXA sides) of two different types of media with mixture of antibiotics which enables the selective growth of CPEs mainly of KPC and metallo-beta-carbapenemase-type on the CARB medium side, and OXA-48-type on the other OXA side.
The cultured plates were then incubated aerobically at 37 °C to be inspected for growth after 18–24 h.
Colonies on CHROM ID CARBA SMART were inspected for their color and interpreted provisionally as manufacturer’s instructions; E. coli (pink to burgundy color), Klebsiella, Enterobacter, Serratia and Citrobacter (bluish-green to bluish-grey or purple color).
Each type of colonies was further subcultured on MacConkey’s agar and blood agar for further species confirmation and antibiotic sensitivity testing (AST).
Species confirmation and antibiotic sensitivity testing (AST)
Identification to the species level and AST of all CPE isolates were performed by Vitek-2 compact system, where sensitivity was performed and interpreted according to CLSI, 2024 breakpoints to the two carbapenems: meropenem (MEM) (sensitive (S) ≤ 1 μg/ml) and imipenem (IPM) (S ≤ 1 μg/ml), as well as to ceftazidime (CAZ) (S ≤ 4 μg/ml), cefepime (FEP) (S ≤ 2 μg/ml), piperacillin (PRL) (S ≤ 8 μg/ml), piperacillin/tazobactam (TZP) (S ≤ 8/4 μg/ml), ticarcillin/clavulanic acid (TCC) (S ≤ 16/2 μg/ml), aztreonam (ATM) (S ≤ 4 μg/ml) as well as ciprofloxacin (CIP) (S ≤ 0.25 μg/ml), minocycline (MIN) (S ≤ 4 μg/ml), amikacin (AK) (S ≤ 4 μg/ml), tobramycin (TOB) (S ≤ 2 μg/ml), gentamycin (CN) (S ≤ 2 μg/ml) and trimethoprim/sulfamethoxazole (SXT) (S ≤ 2/38 μg/ml) [10]. Epidemiological cut-off points “ECOFFs” released by the European Committee on Antimicrobial Susceptibility Testing (EUCAST) were also referred to in cases of isolates recovered on chromogenic agar media and showing sensitivity to carbapenems, where the screening cut-off breakpoint of meropenem was much reduced (R > 0.125 μg/ml), [11, 12].
Polymerase chain reaction (PCR) for carbapenemase genes
DNA extraction and detection of carbapenemase genes were performed to all recovered Enterobacterales by conventional PCR using previously described primers for detection of blaNDM, blaOXA-48, blaVIM, blaKPC and blaIMP by Poirel et al. in 2011 [13]. PCR conditions included initial denaturation at 94 °C for 10 min, followed by 34 cycles of amplification (94 °C for 30 s, 52 °C for 40 s and 72 °C for 50 s) and lastly a final extension step at 72 °C for 5 min. Detection of amplified PCR products was performed on 2% agarose gel electrophoresis.
Sequencing for 16Sr RNA
Stool specimens were kept at −70 °C till DNA extraction.
Stool DNA extraction of 13 CPE carriers and 20 non-carriers was performed using QIAamp Fast DNA stool mini kits (Qiagen) in accordance with the manufacturer’s instructions. Primers 341 F and 805R that target bacterial 16S rRNA gene were used for bacterial PCR amplification. The 16S rRNA gene (V4 region) was addressed utilizing universal primers that target a broad range of bacteria (F: 50-GTGCCAGCMGCCGCGGTAA-30, R: 50-GGACTACHVGGGTWTCTAAT −30), incorporating a unique 6-bp barcode for each sample associated with the paired primer [14]. The amplified products were purified and sequenced with Illumina Miseq Sequencing System (Illumina). By using the EzBioCloud 16S rRNA database, raw sequencing data was quality filtered and subjected to rarefaction analysis to evaluate sequencing depth, followed by further taxonomic classification and annotation of operation taxonomic units [15].
The microbiota composition was categorized at the phylum, genus, and species taxonomical levels, and was represented as the average relative abundance (RA). The assessment of Alpha and Beta microbial diversities was conducted using the Shannon index for Alpha diversity and the Bray–Curtis dissimilarity index for Beta diversity. Linear discriminant analysis effect size (LEfSe) was employed to identify the distinctive taxonomic biomarkers between the CRE/CPE positive (+ ve) and CRE/CPE negative (-ve) groups. Ultimately, group comparisons were made between CRE/CPE positive and negative groups in terms of microbial taxonomical composition as well as alpha and beta diversities.
Statistical analysis
Data were entered and managed using IBM SPSS version 25 and JAMOVI software (version 2.3.28). The association between categorical variables was primarily examined using Chi-square test of independence for nominal data. When the expected count in any cell was less than five, Fisher's exact test was used to determine the exact P-value. The strength of association was measured using the Odds Ratio (OR) with a 95% Confidence Interval (CI). For reliability, a Haldane correction was applied to contingency tables where any cell count was zero. Variables that showed no detection in any CPE positive patients were excluded from the analysis. To facilitate the calculation of interpretable positive odds ratios, certain variables were re-categorized or mathematically transformed. Statistical significance was defined as a P value w below P < 0.05).
Further analyses were conducted using the EzBiocloud software (EzBiome, Inc, USA). The Wilcoxon rank-sum test was employed to assess significant differences in microbial abundance and alpha diversity across group comparisons. Additionally, PERMANOVA (per-mutational multivariate analysis of variance) was utilized to evaluate the differences in beta diversity measures among the study groups.
Results
Between March 2022 and September 2022, 70 patients with equal sex distribution were included in the study. The median age of the participants was 56.6 years (min 19 – max 77). Sixteen patients (10 females and 6 males) were identified as CPE carriers (22.85%), and 54 (77.14%) were non carriers by culture results on chromogenic agar media. Thirteen were positive at admission and three acquired CPE during their hospital stay as detected in their follow up rectal swab cultures (3/16; 18.75%). Prior hospitalization was found in five of the CPE negative patients who were administered to the ICU. Unfortunately, history of previous administration of antibiotics was not conclusive as nearly all patients were exposed to several antimicrobial agents, due to the irrational use of antibiotics available over the counter, other risk factors were not significant (Table 1).Table 1. Risk factors for CPE colonizationRisk factorCPETotalOdds ratioConfidence intervalP valuePositive****NegativeAge1.0150.243- 4.2471 < 403 (4.3%)10 (14.3%)13 (18.6%) > 4013 (18.6%)44 (62.9%)57 (81.4%)Sex1.9330.615- 6.0740.255 Female10 (14.3%)25 (35.7%)35 (50%) Male6 (8.6%)29 (41.4%)35 (50%)Education3.8370.456—32.2840.272 non-educated15 (21.4%)43 (61.4%)58 (82.9%) educated1 (1.4%)11 (15.7%)12 (17.1%)Occupation5.210.282–96.30.339 working0 (0%)7 (10.0%)7 (10.0%) not-working16 (22.9%)47 (67.1%)63 (90.0%)Diabetes mellitus9.710.543–1730.055 negative16 (22.9%)42 (60.0%)58 (82.9%) positive0 (0.0%)12 (17.1%)12 (17.1%)Hypertension1.3220.426 −4.1000.628 positive7 (10.0%)20 (28.6%)27 (38.6%) negative9 (12.9%)34 (48.6%)43 (61.4%)Chronic liver disease1.1540.321–4.1451 negative12 (17.1%)39 (55.7%)51 (72.9%) positive4 (5.7%)15 (21.4%)19 (27.1%)Chronic renal disease10.50.409 −2720.229 positive1 (1.4%)0 (0.0%)1 (1.4%) negative15 (21.4%)54 (77.1%)69 (98.6%)History of chemotherapy2.940.150–57.60.567 negative16 (22.9%)50 (71.4%)66(94.3%) positive0 (0.0%)4 (5.7%)4 (5.7%)History of antibiotic use1.7330.147–20.4560.547 positive1 (1.4%)2 (2.9%)3 (4.3%) negative15(21.4%)52 (74.3%)67(95.7%)History of ICU3.670.192–69.90.582 positive0 (0.0%)5 (7.1%)5 (7.1%) negative16 (22.9%)49 (70.0%)65 (92.9%)
CPE isolates included E. coli as the most commonly detected species (9/16; 56.25%), followed by K. pneumoniae (7/16; 43.75%) as identified by their colony color in accordance with the manufacturer’s documents of CHROM ID CARBA SMART agar (Fig. 1). Confirmation of species identification and antibiotic sensitivity testing was performed by Vitek-2 system.Fig. 1CHROM ID CARBA SMART agar showing E. coli pink colonies on CARB side and Klebsiella pneumoniae blue colonies on OXA side
Antibiotic sensitivity testing by Vitek-2 compact system, following CLSI, 2024 breakpoints, showed that 11 CPE isolates were resistant to both carbapenems (imipenem and meropenem). Whereas two isolates were sensitive to both carbapenems (Imipenem; MIC = 0.5 μg/ml, meropenem; MIC = 0.5 & 1 μg/ml). three isolates were found intermediate to imipenem (MIC = 2 μg/ml), one of them was also intermediate to meropenem (MIC = 2 μg/ml) and the other two were sensitive to meropenem (MIC ≤ 0.25 & 1 μg/ml), however they were considered resistant by ECOFFs. (Table 2). Regarding β-lactam antibiotics; all 16 CPE isolates were resistant to ceftazidime, cefepime, and β-lactam/β-lactamase inhibitor combinations, including piperacillin/tazobactam, except one blaOXA-48 positive E. coli isolate that was sensitive to ceftazidime and cefepime (MIC = 1 μg/ml for both), whereas 12.5% (2/16) were sensitive to aztreonam. Best sensitivity was found to aminoglycosides primarily to amikacin (10/16; 62.5%), followed by gentamicin (8/16; 50%), then tobramycin (5/16; 31.25%). Sensitivity to ciprofloxacin was 12.5% (2/16), and to minocycline 18.75% (3/16). All isolates were resistant to trimethoprim/sulfamethoxazole.Table 2. Antibiotic sensitivity testing results to carbapenems of 16 CPE isolates recovered on chromogenic agar media in relation to their carbapenemase genesIsolateSpeciesCarbapenemase producing genes detected by PCRSensitivity to carbapenems (MIC(μg/ml); Interpretation(S/I/R)Imipenem**Meropenem6RE. colibla*NDM, blaOXA-48, blaVIM ≥ 16; R8; R8RK. pneumoniablaNDM, blaOXA-48 ≥ 16; R ≥ 16; R13RE. coli**blaNDM ≥ 16; R ≥ 16; R26RE. coli**blaNDM ≥ 16; R ≥ 16; R27RE. coli**bla_OXA-48_0.5; S1; S31RK. pneumonia**blaNDM ≥ 16; R ≥ 16; R42RE. coli**blaNDM, blaOXA-48 ≥ 16; R ≥ 16; R47RE. coli**blaNDM, blaOXA-48 ≥ 16; R ≥ 16; R52RE. coli**blaNDM, blaOXA-48 ≥ 16; R ≥ 16; R60RE. coli**bla_OXA-48_0.5; S0.5; S61RK. pneumonia**bla_OXA-48_2; I ≤ 0.25; S69RK. pneumonia**blaNDM, blaOXA-48, bla_VIM_2; I2; I71RK. pneumonia**blaNDM ≥ 16; R ≥ 16; R72RE. coli**bla_OXA-48_2; I1; S73RK. pneumonia**blaNDM ≥ 16; R8; R74RK. pneumonia**blaNDM ≥ 16; R ≥ 16; R^*^MIC Minimum inhibitory concentration, S Sensitive, I Intermediate, R Resistant
Carbapenemase genes were detected in all tested isolates confirming they were all CPEs. According to PCR results four genotypes of carbapenemases genes existed (Table 3); where blaNDM was found alone in six isolates (four K. pneumoniae and two E. coli), and with blaOXA-48 in four isolates (three E. coli and one K. pneumoniae). Two isolates (one E. coli and one K. pneumoniae) had the three genes (blaNDM, blaOXA-48 and blaVIM), blaOXA-48 existed alone in four isolates (three E. coli and one K. pneumoniae). So blaNDM was the predominant gene in (12/16) (75%) of all CPE isolates; mainly in K. pneumoniae isolates (n = 6/7) (85.7%), followed by blaOXA-48 in 62.5% (10/16), predominantly in E. coli isolates (n = 7/9) (77.8%). Two isolates had blaVIM together with blaNDM and blaOXA-48, whereas blaKPC and blaIMP were not detected in all tested isolates.Table 3. Types, number and distribution of resistance genes among isolated species from 16 CPE colonized patientsType of Resistance genesNumber of IsolatesNumber of isolates in each speciesK. pneumoniaeE. coli1- bla_NDM_6422- bla_OXA-48_4133- blaNDM + bla_OXA-48_4134- blaNDM + blaOXA-48 + bla_VIM_211Total1679
Urinary tract infection was confirmed in two out of the 16 CPE carriers (12.5% (2/16)), one at admission and the other was detected with the follow up clinical specimens. Isolates were of the same species and molecular characterization as the colonizing CPE ones; one was E. coli and the other was K. pnuemoniae and both were positive for blaNDM only.
Using the EzBioCloud 16S rRNA database for taxonomic assignment comparative MTP analyzer of stool microbiota between 20 CPE negative stool specimens and 13 CPE positive revealed the following:
The relative abundance of the phyla in CPE carriers comprised 43% Firmicutes, 29% Bacteroidetes and 25% Proteobacteria (Fig. 2). In CPE non-carriers, the relative abundance of the phyla comprised 54% Firmicutes, 28% Bacteroidetes and 14% Proteobacteria. Thus, gut microbiota from CPE carriers showed a significant increased proportion of Proteobacteria (p = 0.03), (Fig. 3). In addition, reduction in Fermicutes/Bacteriodetes (F/B) ratio was noticed, though not statistically significant (p = 0.796) (Fig. 4). Also, Linear discriminant analysis (LDA) with linear discriminant effect size (LEfSe) revealed that several bacterial genera were noticeably altered in the gut microbiota of CPE carriers compared with those in non-carriers. Among CPE carriers, the top genera were Enterobacter cloacae, Bacteroidales, Prasutterella excrementihominis, Gammaproteobacteria and Proteobacteria, whereas other genera as Coprococcus, Mogibacterium, Bilophilia showed lower 2 LDA scores (Fig. 5).Fig. 2. EzBioCloud analysis showing relative abundance of composition of different phyla existing in CRE negative versus CRE positive isolatesFig. 3Boxplot showing significant increased proportion of Proteobacteria in gut microbiota in CRE carriers compared to CRE non-carriers (Wilcoxon rank-sum test, p = 0.03). The boxes illustrate interquartile ranges (IQR), with the horizontal line indicating the median and whiskers extending to the most extreme valuesFig. 4Microbiota taxonomical analysis showing non-significance decrease in Fermicutes/Bacteriodetes (F/B) ratio in CRE positive relative to CRE negative isolates (p = 0.796)Fig. 5. Linear discriminant analysis (LDA) scores obtained from linear discriminant effect size (LEfSe) analysis of fecal microbiome. An LDA effect size of more than 2 was used as a threshold for the LEfSe analysis showing the top genera in CRE carriers; Enterobacter cloacae, Bacteroidales, Prasutterella excrementihominis, Gammaproteobacteria, Proteobacteria. Whereas other genera as Coprococcus, Mogibacterium, Bilophilia showed lower 2 LDA scores in CRE negative patients
The alpha diversity indices (Shannon, Chao1, and OTUs) were non-significantly decreased inCPE carriers compared to the CPE non-carriers (p = 0.210, 0.658, 0.754 respectively) (Fig. 6). However, the beta diversity analysis (Bray–Curtis distances) demonstrated that the two groups were not significantly apart (p = 0.178) (Fig. 7).Fig. 6. Gut Microbial Alpha diversity indices among CRE positive and negative groups showing non-significant decrease in CRE carriers analyzed by Wilcoxon rank-sum test: A) Shannon, B) Chao1, C) number of OTUsFig. 7Beta diversity analysis of Gut microbiota using Bray–Curtis dissimilarity index. among CRE positive and negative groups, where the 2 groups are not significantly apart (p = 0.178). A principal coordinate analysis (PCoA) plot illustrating the degree of similarity in microbiota profiles between the two groups, with each coordinate indicating the percentage of variance. The differences between study groups were assessed by PERMANOVA test (per-mutational multivariate analysis of variance)
Discussion
Intestinal colonization with CPE poses a significant source of community dissemination and hospital acquired infection. Therefore, it is mandatory to promptly identify and isolate CPE carriers to reduce transmission and improve patients’ outcomes [12].
The current work aimed to assess the rate of colonization of CPE, the possibility of subsequent infection, and the associated alteration in the gut microbiota.
Our results showed that CPE carriage was found in 22.85%, which is comparable with previous studies in Egypt, where it represented 24% & 28% [12, 16]. Lower rates of colonization were obtained from Kuwait that ranges between 8 to 12% [17], 17% in Nigeria [18], and 9.6% in Kenya (9.6%) [19]. Colonization prevalence of CPE is reported highest in Asia and Africa, whereas lower, but still significant, rates were detected in the Americas and Europe. This geographical variations in colonization prevalence confirms the need to identify contributing regional factors in the community, including poor water and sanitation [20].
In this study colonization that occurred after 72 h from admission was considered hospital acquired and it represented 19% of total carriers that was going with what has been reported recently about the three days median duration suggested from admission to colonization with CPE. However, a limitation in the current work is that it represents a single center study that may reflect the environment of our hospital, located in an endemic area with poor sanitary resources. Thus, though most of the CPE colonization was found to be community acquired, that may represent the special nature of the patients attending our hospital who were mainly from the neighborhood. It is also suggested that variation in colonization rates occur within different departments and could be associated with pharmacological therapies, patient groups and clinical settings [21].
No significant differences were found between CPE positive and negative groups regarding risk factors, as the presence of comorbidities, invasive procedures, previous administration of antibiotics and previous ICU admission (all P > 0.05). This was in accordance with previous single center study that could not identify any clinical risk factors which were significantly associated with acquisition of gut pathogens [9]. However, that may be attributed to the endemic location of our hospital and the small sample size included in the current study.
Carbapenemases resistance genes were detected in all tested isolates confirming it was the main mechanism of resistance. However, according to CLSI, 2024 breakpoints, five isolates were not resistant, where blaOXA-48 was detected in these isolates alone in four of them, and with blaVIM and blaNDM in one isolate (Table 2). This could be explained by the diminished carbapenemase activity of OXA-48 enzymes compared to KPC and MBLs. For this reason, detection of OXA-48-producing Enterobacterales based on susceptibility phenotype is more challenging as it may not meet the current CLSI criteria for intermediate or resistance sensitivity results [4, 10]. However, the carbapenems intermediate susceptibility results expressed in the isolate harboring the three resistance genes; blaOXA-48, blaNDM, and blaVIM may indicate the lack of expression or the presence of uncommon enzyme derivatives (especially of NDM). Unfortunately sequencing this gene was not accessible. Resistance to other classes of antibiotics found in our study confirms the MDR nature of nearly all CPE isolates [21, 22].
Although blaOXA-48 represented the main carbapenemase producing gene in previous work [12], in the current study blaNDM was the leading gene in 75% of the isolates, which confirms the persistent nature described of NDM carbapenemases and their tendency to be endemic in Egypt [23, 24]. Enterobacterales producing OXA-48 are endemic in other neighboring countries as Tunisia, Morocco and [25]. Rectal swabs surveillance specimens from ICU patients in Bahrain, Kuwait, Saudi Arabia, Oman, and the United Arab Emirates in 2019 revealed blaOXA-48 as the most prevalent carbapenemase gene in 15% of all specimens screened and up to 51% of those in Saudi Arabia [25, 26]. Different regions in the world and sometimes difference in same countries show different predominance in CPE determinants [12]. The rise in carbapenem resistance was reported globally by the European Center for Disease Prevention and Control (ECDC) and the World Health Organization (WHO) where the prevalence of carbapenem resistance in K. pneumoniae isolates raised by 20% in 2023 compared to 2017,whereas the national average of resistance to carbapenems in E. coli remained at 1.6% [27]. The overall risk of CPE infection in colonized patients was reported as 16.5% and 22.5% in other studies [28, 29]. This is comparable with our study where infection associated with CPE colonization was detected in 12.5%, thus supporting the strong association between CPE colonization and development of clinical infections previously reported [1].
Normal constituent of Microbiota in the Egyptian population is still unrevealed and needs more advanced future studies to correlate types of dysbiosis due to CPE carriage. Proteobacteria represent one of the most abundant phyla in the human gut microbiota and its dominance is associated with increased inflammation. It was reported that CPE carriers have higher number of Proteobacteria and lower prevalence of Firmicutes and Bacteroidetes than non-carriers and healthy controls [30].
Our results confirmed the significant predominance of Proteobacteria in CPE carriers with p < 0.03. However, the decrease in Bacteroidetes phyla was not significant. The Enterobacterales family are normally present in small fraction of total bacteria in healthy individuals, whereas it can abnormally proliferate in the gut of individuals affected by intestinal or extra-intestinal diseases such as obesity, inflammatory bowel disease or metabolic disorders [31]. It has been reported that Proteobacteria abundance is implicated in the genesis of endotoxemia and in the development of metabolic disorders [32].
Similarly, LDA with LEfSe showed alteration in several bacterial genera in the gut microbiota of CPE carriers, though not statistically significant, compared with those in non-carriers. Although as reported by Baek et al., 2023 [5], the alpha diversity was decreased in CPE carriers compared to the CPE non-carriers, however Shannon index was statistically non-significantly (p = 0.210), also the beta diversity analysis (Bray–Curtis distances) demonstrated that the two groups were not significantly apart (p = 0.178). Further studies of larger sample sizes may reveal more details in that context.
Our study has limitations that included being a single center study, the small sample size that hindered the performance of multivariate analysis for risk factors as they were not statistically significant.
Conclusion
Increased laboratory capacity to improve and shorten duration of detection is on top priorities to combat challenge of antimicrobial resistance. Detection of CPE colonization, efficient antimicrobial stewardships, strict infection control measures and active surveillance programs are imperative to trace and prevent their acquisition and spread. The use of differential chromogenic agar in screening in endemic regions, as ours, is highly recommended. Alternatively, the application of ECOFFs is encouraged to screen to detect low enzymatic activity of some carbapenemases as OXA-48-like. Our results confirm the pervasiveness of carbapenem resistance in our region especially NDM enzyme. Alteration in microbiota and the abundance of Proteobacteria in CPE carriers may indicate a predisposition to inflammatory states. Further multi-centered studies of larger sample size are needed to investigate the effect of microbiota alteration on the health of patients colonized by CPE.
Supplementary Information
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al Fadhli AH, Jamal WY, Rotimi VO. Prevalence of carbapenem‐resistant Enterobacteriaceae and emergence of high rectal colonization rates of bla OXA‐181‐positive isolates in patients admitted to two major hospital intensive care units in Kuwait. Plos One. 2020;2020. 10.1371/journal.pone.0241971.10.1371/journal.pone.0241971 PMC 767151433201906 · doi ↗ · pubmed ↗
- 2European Centre for Disease Prevention and Control and World Health Organization. Antimicrobial Resistance Surveillance in Europe 2023–2021 Data. Available online at: https://www.ecdc.europa.eu/en/publications-data/antimicrobial-resistance-surveillance-europe-2023-2021-data. 2023