Natural Products in Cancer Prevention and Therapy: Current Challenges and Future Directions
Ruimiao Qian, Jun Ge, Ni Fan, Zheng Sun, Chengcheng Zhao, Yujiao Sun, Yingpeng Li, Yunfei Li, Hui Fu

TL;DR
Natural products offer cancer prevention and treatment by targeting metabolism and the tumor environment, with new delivery methods improving their effectiveness.
Contribution
This review systematically outlines how natural products target metabolism and the tumor microenvironment across all clinical phases of cancer care.
Findings
Natural products modulate glucose, lipid, and glutamine pathways in cancer cells.
They influence tumor-associated macrophages and T lymphocytes in the tumor microenvironment.
Innovative delivery systems like nanocarriers enhance the bioavailability and synergy of natural products with other therapies.
Abstract
Natural products, originating from diverse biological sources, serve as a critical reservoir of bioactive compounds for cancer intervention across prevention, treatment, and supportive care. Their mechanisms extend beyond direct cytotoxicity to include modulation of tumor metabolism—such as glucose, lipid, and glutamine pathways—and the tumor microenvironment (TME), highlighting their multifaceted role in oncology. However, a systematic synthesis of how natural products concurrently target metabolic reprogramming and immune–stromal components across different clinical phases remains lacking. This review delineates the therapeutic applications of natural products—such as flavonoids, alkaloids, and terpenoids—across the clinical continuum, including perioperative support, concurrent chemoradiotherapy, maintenance therapy, and metastasis suppression. We detail their actions in disrupting…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
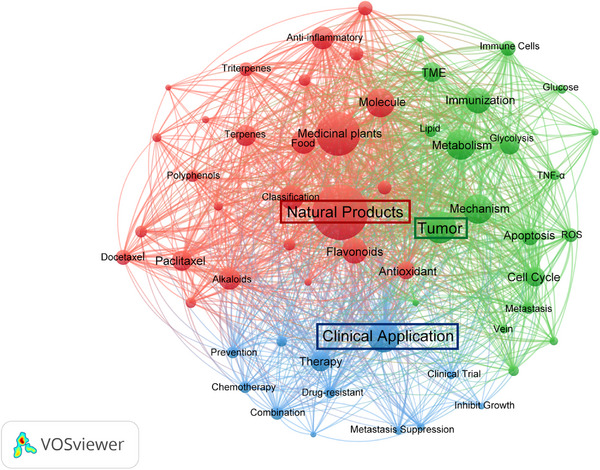 FIGURE 1
FIGURE 1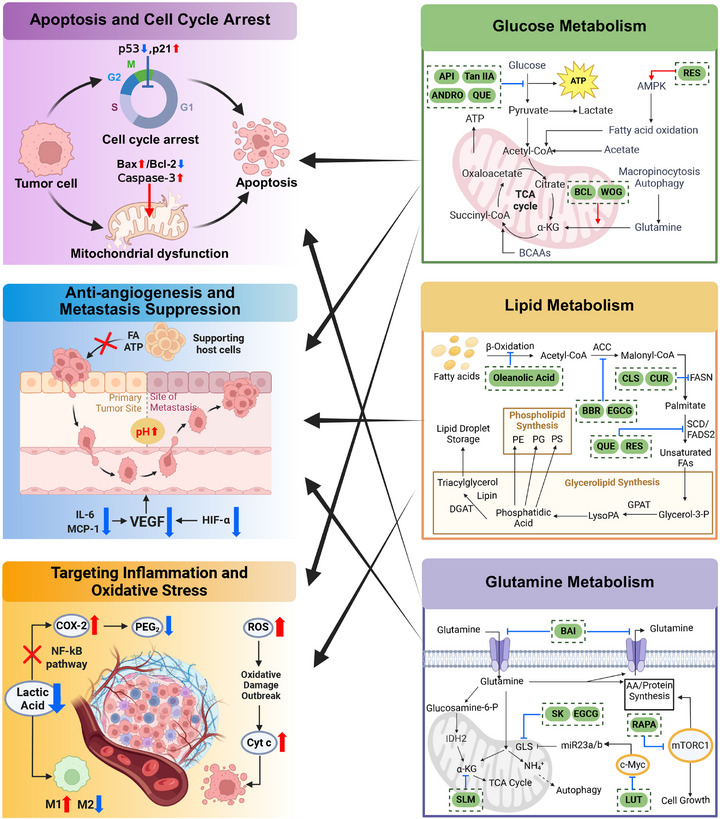 FIGURE 2
FIGURE 2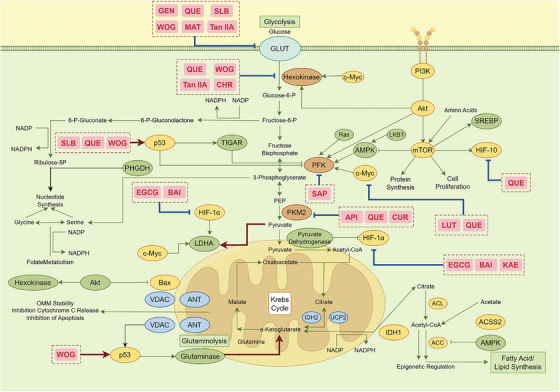 FIGURE 3
FIGURE 3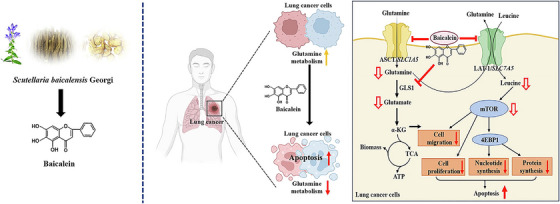 FIGURE 4
FIGURE 4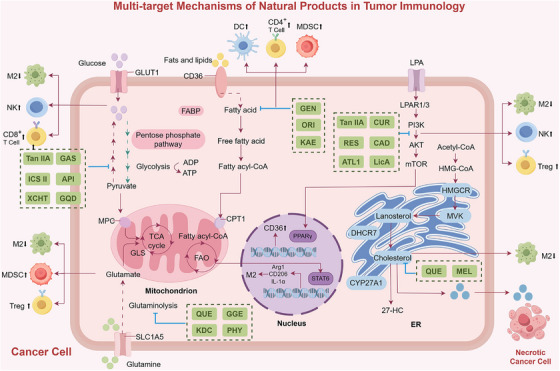 FIGURE 5
FIGURE 5 FIGURE 6
FIGURE 6| Disease stage | Natural products | Cancer type | Function | Dosage and administration | Research phase | Mechanism of action | References |
|---|---|---|---|---|---|---|---|
| Primary prevention | Soy isoflavones | Breast cancer | Reduce the risk of cancer | Oral, 2 servings of soy products daily (containing approximately 50 mg of isoflavone aglycones) | Randomized controlled trial | Competitively binds to estrogen receptors, blocking potent estrogen's procancer signaling; affects levels of risk biomarkers such as IGF‐1, CRP, and leptin | [ |
| Novel purified fraction of almond oil (ricinol, γ‐tocopherol) | LoVo, HT29, and Hep3B cell lines | Chemoprevention of potential cancer | 0.001, 0.01, and 0.1 mg/mL | In vitro study | Alter cell cycle characteristics; tumor cell migration activity↓; suppress metabolic pathways | [ | |
|
| Human endometrial adenocarcinoma cells | Malignancy chemoprevention | 50–1000 µg/mL | In vitro study | Cancer cell metabolic activity↓; DNA synthesis capability↓; induces cell death via apoptosis and/or membrane damage | [ | |
| Resveratrol | Head and neck cancer | As an adjunct to home enteral nutrition, enhances antioxidant defenses, improves cellular health indicators (e.g., phase angle) | Oral, 400 mg daily of liposomal resveratrol, divided into two doses (5 mL each) | Randomized controlled trial | Glutathione peroxidase activity↑; total antioxidant capacity and superoxide dismutase activity↑; increased oxidative stress, malondialdehyde levels↑; induced cytotoxicity | [ | |
| Concurrent treatment phase | SH003 ( | Lung cancer, breast cancer | Enhance chemotherapy efficacy while reducing toxicity |
SH003: oral, 2400–4800 mg/day Docetaxel: intravenous injection, 75 mg/m2 | Phase I clinical trial | Affects the cell cycle and induces apoptosis; inhibits the Akt/mTOR pathway; cancer cells' antiapoptotic capacity↓; immune modulation↑ | [ |
| Quercetin | Breast cancer, prostate cancer | Inhibits lymphocyte tyrosinase activity | 1400 mg/m2, IV infusion every 3 weeks | Phase I clinical trial | Induces apoptosis and has antiproliferative effects | [ | |
| EGCG | Breast, prostate, lung, colorectal cancer | Treatment of COVID‐19 pneumonia | 30 mL daily nebulization (1760–8817 µmol/L) | Phase I/II clinical trial | Downregulates inflammatory mediator expression and signaling by acting on STAT1/3 and NF‐κB factors | [ | |
| Tanshinone IIA (Tan IIA) | Colorectal cancer | Induces ferroptosis in colorectal cancer cells | 20 µM Tan IIA + SLC7A11 plasmid | In vivo study | Suppresses SLC7A11 expression via the PI3K/AKT/mTOR pathway | [ | |
| Postoperative recovery | Rikkunshito | Gastric cancer | Reduces weight loss, improves nutritional indicators, increases food intake | Oral, 7.5 g/day | Retrospective observational study | Auxin secretion↑, appetite↑; Regulates gastrointestinal motility and hormone levels | [ |
| Ginsenoside Rg1/Rh2 | Melanoma (mouse model) | Promotes humoral immunity, enhances T lymphocyte infiltration | Injection, 0.5 or 0.2 mg/kg | In vivo study | Influences Th cell activity, IgA production↑; induces infiltration of CD4+ and CD8a+ T lymphocytes | [ | |
| Icariin | Prostate cancer | Inhibits tumor growth, promotes immunity | Intraperitoneal injection, 10 mg/kg | In vivo study | Amounts of probiotics (such as Lactobacillus and Bifidobacterium)↑; activity of various lymphocyte subsets↑ | [ | |
| Maintenance therapy | Shikonin | CT26 mouse colon cancer cells | Inhibits tumor growth and metastasis, induces cell death | 0.16–20 µg/mL | Preclinical study (in vitro and in vivo) | Induces apoptosis and cell cycle arrest; induces novel cell death forms (e.g., ferroptosis, pyroptosis); inhibits tumor metabolism (Warburg effect) by targeting PKM2 | [ |
| Berberine | Colorectal cancer | Long‐term prevention of adenoma recurrence, reduces risk of colorectal tumorigenesis | Oral, 0.3 g, twice daily |
Randomized controlled trial | Multitargeted regulation of the intestinal microenvironment and inflammatory response. | [ | |
| End‐of‐life care |
Longteng tongluo recipe (LTTL) | Lung cancer | Effectively alleviates lung cancer pain and significantly reduces opioid consumption |
Apply LTTL tincture topically, 0.5 mL per cm2 per application, twice daily for 8 consecutive days |
Randomized controlled trial | Serum miRNA expression levels associated with cancer pain (hsa‐miR‐2110, hsa‐miR‐7d‐3p)↓ | [ |
| Metabolism type | Key target | Representative natural products | Mechanism of action | Cancer type/cell model | Effects | References |
|---|---|---|---|---|---|---|
| Glucose metabolism | PKM2 | Quercetin, apigenin | PKM2 expression via Akt/mTOR or β‐catenin/c‐MYC/PTBP2↓ | Colorectal cancer and breast cancer cells | Glycolysis↓, apoptosis↑ | [ |
| HK2 | Quercetin, xanthohumol | HK2 via Akt/mTOR or EGFR–Akt signaling↓ | HCC, glioblastoma | Glucose phosphorylation and ATP production↓ | [ | |
| PFK1 | Amentoflavone | PFKP expression↓, glycolysis inhibition | HepG2 | Glycolytic flux↓ | [ | |
| GAPDH | Luteolin | Competitive inhibition of GAPDH enzymatic activity | HepG2, HeLa | Glycolytic flux↓, cell cycle arrest | [ | |
| ENO1 | Silibinin | mRNA levels of ENO1 gene↓, disrupting glycolysis | Breast cancer | ATP↓, chemosensitivity↑ | [ | |
| GLUT1/GLUT4 | Apigenin, casticin | Glucose uptake via transporter suppression↓ | Multiple solid tumors | Glucose availability↓, cell proliferation↓ | [ | |
| SGLT1/2 | Trilobatin | Nonselective inhibition of SGLT‐mediated uptake | Human hepatoblastoma | Glucose uptake↓, tumor suppression | [ | |
| PGK1 | Isoflavones | miR‐29a/miR‐1256‐mediated PGK1 inhibition | Prostate cancer | ↓Migration, invasion | [ | |
| LDHA | Catechins | LDHA↓ → ROS↑, apoptosis induction | Gastric cancer | Induce ROS‐mediated cytotoxicity | [ | |
| HIF‐1α | EGCG, baicalein, kaempferol | HIF‐1α expression/activity via AMPK or direct inhibition↓ | HeLa, HepG2 | Angiogenesis↓, glycolysis↓, apoptosis↑ | [ | |
| p53 | Oroxylin A, baicalein | p53 stability↑, expression → GLUTs↓, apoptosis↑ | HepG2 | Tumor growth↓, glycolysis suppression↑ | [ | |
| c‐MYC | Lignans | c‐MYC signaling↓ (PI3K/Akt, Src/Cortactin, EMT‐related) | Breast, lung cancer | Proliferation↓, metastasis↓, glycolysis↓ | [ | |
| NF‐κB | Epicatechin | NF‐κB nuclear translocation↓ | Multiple solid tumors | Glycolysis‐related gene expression↓ | [ | |
| miR‐214‐5p | Cyanidin‐3‐O‐glucoside (C3G) | miRNA‐mediated regulation of GLUT1 and AKT/mTOR/PTEN pathway | Oral squamous carcinoma, others | Metabolic inhibition, cell growth↓ | [ | |
| AMPK, Bax/Bcl‐2 | Multiple flavonoids | AMPK activation, apoptosis modulation | Various cancer types | Mitochondrial dysfunction, viability↓ | [ | |
| Lipid metabolism | FASN | EGCG | KR domain activity of FASN↓, blocks fatty acid synthesis | Various solid tumors | Tumor growth↓ | [ |
| Acetyl‐CoA carboxylase (ACC) | Fisetin | ACC activity↓, malonyl‐CoA production↓ | Prostate cancer | Fatty acid synthesis↓, apoptosis↑ | [ | |
| Stearoyl‐CoA desaturase‐1 (SCD1) | Genistein | SCD1 expression via AMPK activation↓ | Lung cancer | Disrupts membrane fluidity, metastasis↓ | [ | |
| NRF2 | Hesperetin | Activates NRF2 antioxidant pathway, indirectly regulates cholesterol homeostasis | HepG2 | Stress resistance↑, delays tumor progression | [ | |
| HMG‐CoA reductase (HMGCR) | Naringenin | Competitive inhibition of HMGCR catalytic site | Liver cancer | Cholesterol synthesis blocks the mevalonate pathway↓ | [ | |
| PLA2 | Resveratrol | PLA2 activity↓, production of proinflammatory lipid mediators↓ | Breast cancer | Dual antitumor and anti‐inflammatory effects | [ | |
| Glutamine metabolism | Glutaminase (GLS) | Curcumin | Glutaminase activity↓, glutamine utilization↓ | Adrenocortical carcinoma | Deprives cancer cells of nitrogen and carbon sources, inhibits proliferation | [ |
| Glutamine | Pholiota adiposa ethanol extract | Levels of amino acids such as glutamine↓, the compensatory energy supply from branched‐chain amino acids↓ | Liver cancer | Tumor growth↓, tumor cell apoptosis↑, protect liver and kidney function, and regulate immune responses | [ |
- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer Research and Treatments · Cancer, Hypoxia, and Metabolism · Immune cells in cancer
Introduction
1
In addition to conventional cancer treatments such as surgery, radiotherapy, and chemotherapy, clinical management is increasingly exploring multitarget, low‐toxicity adjunctive strategies. Natural products—bioactive compounds derived from plants, microorganisms, and marine organisms—offer significant therapeutic potential in this regard, owing to their chemical diversity and multifaceted biological activities [1].
Natural products exhibit broad applications throughout the cancer treatment continuum. During the perioperative phase, natural products serve as adjuncts to chemotherapy and radiotherapy. They not only enhance cancer cell sensitivity—for example, apigenin increases pancreatic cancer cells’ sensitivity to gemcitabine—but also mitigate the toxic side effects of conventional drugs such as cisplatin, capecitabine, and fluorouracil [2, 3, 4, 5]. At the same time, certain natural products, such as the active components in rhubarb, can improve patients’ physical condition and reduce inflammatory responses during the perioperative period [6]. In the later stages of treatment and during rehabilitation, natural products play a pivotal role in maintaining therapeutic efficacy, preventing metastasis, and improving overall quality of life [7, 8]. Figure 1 highlights the interconnected roles of natural products in cancer therapy and their clinical applications.
Co‐occurrence network analysis of natural product‐related research using VOSviewer. This network visualization illustrates the central role of natural products in cancer therapy, highlighting their connections to concepts such as clinical applications, tumor metabolism, immune regulation, and mechanisms of action. Key clusters are color coded: red represents natural product sources and classification; green emphasizes cancer biology, related mechanisms, and metabolic processes; and blue denotes tumor clinical treatment and applications.
The primary advantage of natural products lies in their ability to address the limitations of conventional cancer therapies. Drugs like cisplatin and oxaliplatin often induce nephrotoxicity, neurotoxicity, and multidrug resistance. Natural compounds such as the organosulfur compounds, present in garlic, can reverse platinum resistance by downregulating P‐glycoprotein (P‐gp) on tumor cells, thereby enhancing intracellular drug accumulation and therapeutic efficacy [9]. Antimetabolites like 5‐fluorouracil suffer from narrow therapeutic windows, significant toxicity, and extensive first‐pass metabolism [10]. Research suggests that chemically linking 5‐fluorouracil with the natural product curcumin (CUR) to form new derivatives can potentiate their synergistic effects while minimizing adverse reactions [11]. Taxol, derived from the yew tree, exhibits significant efficacy against various malignancies by targeting tumor cell mitosis and inducing apoptosis, and it is now widely used after being synthesized via semi‐synthetic techniques and synthetic biology [12]. Additionally, natural products such as astragaloside IV, matrine, and ginsenosides feature novel structures, multiple targets, diverse pathways, minimal toxicity, and multifaceted biological activities. These properties enable them to exert unique antitumor effects, particularly by modulating tumor metabolism and regulating the tumor immune microenvironment (TIME) to generate antitumor activity [13].
Mechanistically, tumor metabolic reprogramming plays a pivotal role as a core component, underpinning effects such as directly inducing tumor cell apoptosis, inhibiting metastasis, and counteracting oxidative stress. For instance, dietary flavonoids suppress glucose, lipid, and glutamine metabolism in cancer cells, disrupting their biosynthesis and energy supply while inhibiting cell proliferation [14]. Furthermore, natural products profoundly reshape the tumor microenvironment (TME) by modulating the functions of cancer‐associated fibroblast (CAFs), tumor‐associated macrophages (TAMs), T lymphocytes, and other immune cells [15]. This remodeling reverses immunosuppression, thereby activating the body's innate antitumor immune response.
However, translating the vast potential of natural products into effective clinical therapies remains a considerable challenge. Major issues include low oral bioavailability, complex in vivo metabolism, and substantial patient variability, which can significantly influence therapeutic efficacy. To overcome these challenges, cutting‐edge technologies, including nanocarriers and targeted delivery systems, are being extensively explored [16, 17]. Additionally, with the advancement of precision medicine, biomarker‐driven patient stratification strategies are essential for tailoring natural product‐based therapies to individual patients.
This review approaches the topic from the overarching framework of natural products in cancer prevention and therapy, outlining their sources, structural classifications, and functional roles across different clinical stages. At the mechanistic level, particular emphasis is placed on the antitumor effects mediated through metabolic reprogramming and modulation of the TIME, with detailed analysis of their multitarget regulation of glucose, lipid, and glutamine metabolic pathways. In parallel, the review examines the current status of clinical translation, summarizing key challenges such as limited bioavailability, formulation barriers, and patient‐specific variability, while highlighting emerging opportunities including nanodelivery platforms, combination therapeutic strategies, and precision medicine‐oriented applications. Collectively, this work provides an integrated perspective to advance the scientific application and future development of natural products in oncology.
Classification and Clinical Context of Natural Products for Therapeutic Application
2
Natural products hold significant value in clinical cancer treatment due to their diverse taxonomic classification and stage‐specific applications [18]. These compounds are systematically categorized based on origin and chemical structure, encompassing flavonoids, alkaloids, terpenoids, polyphenols, and other classes, each exhibiting unique biological activities [19]. Moreover, they demonstrate multitarget regulatory functions throughout the entire clinical cancer treatment process—from primary prevention, perioperative support, and concurrent chemoradiotherapy to postoperative recovery, maintenance therapy, and end‐of‐life care. This exploration of chemical taxonomy and clinical‐stage applications aids in understanding the application strategies of natural products in precision oncology.
Sources and Classifications of Natural Products
2.1
Natural products can be categorized into food‐derived compounds and medicinal‐derived compounds based on their sources and intended applications. Food‐derived compounds, such as lycopene in tomatoes and anthocyanins in blueberries, originate from the daily diet. Their core value lies in providing nutrition and maintaining physiological homeostasis, emphasizing long‐term, low‐dose disease prevention [20]. These compounds are typically consumed through regular diets or dietary supplements and are generally considered to have relatively high safety profiles. In contrast, medicinal‐source compounds, such as paclitaxel, ginsenosides, and berberine, are derived from medicinal plants with well‐defined pharmacological activities [21, 22, 23]. Their primary development targets disease treatment directly, requiring extraction, purification, and formulation into specific pharmaceuticals to enable high‐dose, phased therapeutic interventions. However, these compounds may carry potential risks, including hepatic, renal, or neurotoxicity [24]. It is important to note that these two categories are not entirely distinct. Many substances, such as CUR, are “food‐medicine dual‐use” compounds, serving both as everyday seasonings and as potential drug candidates, representing a continuum of intervention strategies from dietary prevention to targeted therapy [25].
It is noteworthy that there exists an intrinsic connection between the origin of natural products and their core mechanisms of action, profoundly influencing their positioning within disease intervention strategies. Compounds derived from medicinal sources typically target critical direct targets essential for the survival of rapidly proliferating cells, exhibiting potent cytotoxic effects [26]. For example, Halorotetin A, a novel terpenoid compound isolated from the ascidian Halocynthia rotetzi, significantly inhibits the proliferation of the hepatocellular carcinoma cell line HepG‐2. Its mechanism of action is closely associated with regulating the expression of oncogenes (such as c‐myc and c‐met) and tumor suppressor genes (such as TP53 and KEAP1), demonstrating the ability to directly intervene in tumorigenesis pathways [27]. In contrast, compounds derived from food sources predominantly act on regulatory pathways, indirectly influencing tumor initiation and progression through multitarget mechanisms [28]. For example, the compound isoegomaketone isolated from the plant P. frutescens significantly inhibited cell growth and xenograft tumor formation, potentially by blocking the PI3K/Akt signaling pathway in HCC cells [29]. Asparagus polysaccharides selectively inhibit proliferation in HepG2 and HepG3B cell lines by inducing G/M phase arrest and apoptosis through Bax, Bcl‐2, and caspase‐3 regulation, while exhibiting low cytotoxicity toward normal human hepatocyte 7702 cells [30]. This classification system of mechanisms—ranging from “direct cell killing” to “multidimensional physiological regulation”—establishes a theoretical foundation for the precise application of diverse natural products.
Natural products can also be classified into several important categories based on their chemical structures and biosynthetic pathways. Flavonoids, a diverse group of polyphenolic compounds abundantly present in fruits, vegetables, tea, legumes, and other plant‐based foods, have emerged as significant candidates for cancer prevention and adjuvant therapy [31]. Numerous studies have demonstrated that regular consumption of flavonoid‐rich foods is associated with a reduced incidence of various malignancies, indicating their remarkable potential as dietary chemopreventive agents [32]. All flavonoids share a core structure consisting of two aromatic benzene rings (A and B) connected by a three‐carbon bridge, forming a heterocyclic pyran ring (C), commonly referred to as the C6–C3–C6 skeleton [33]. Their potent biological activities are attributed to their antioxidant properties and their influence on tumor metabolic reprogramming. Representative compounds include quercetin, found in vegetables and fruits, and the widely distributed apigenin, both of which demonstrate multifaceted potential in anticancer research, such as inducing apoptosis, suppressing inflammatory responses, and inhibiting tumor angiogenesis [2, 34].
Alkaloids are naturally occurring organic compounds containing nitrogen atoms. Their use dates back to ancient Greece, where poppies were employed for pain relief. In modern times, alkaloids’ potential in cancer prevention and treatment has been extensively studied, with their anticancer effects often linked to interference with cell division [35]. Vinblastine, extracted from Catharanthus roseus, and its derivative vincristine inhibit microtubule protein polymerization, thereby blocking cancer cell mitosis [36]. Similarly, two derivatives of camptothecin, topotecan and irinotecan, have been widely used in cancer treatment [37].
Terpenoids play a crucial role in chemical communication between plants and their environment. Plants can produce a wide variety of terpenoids, making them the largest class of plant‐derived natural products. Composed of isoprene units, terpenoids exhibit significant antitumor activity and are considered promising agents for antitumor drug development [38]. Paclitaxel, isolated from the Pacific yew tree (Taxus brevifolia), was the first plant‐derived chemotherapeutic agent widely used in oncology and remains one of the most common drugs in this field. It exerts potent cytotoxic effects by stabilizing microtubules and preventing their depolymerization [21].
Natural polyphenols, a broad category of polyhydroxyphenolic compounds, are among the most widely distributed and abundant natural compounds in nature. Flavonoids, phenolic acids, lignans, and stilbenes are the four main structural types of natural polyphenols. In recent years, polyphenols have garnered extensive attention due to their broad therapeutic effects, including anti‐inflammatory, antioxidant, and antitumor activities [39]. Representative compounds include resveratrol from grape skin and epigallocatechin gallate (EGCG) from green tea. These compounds primarily inhibit tumor initiation and development by regulating key signaling pathways, such as NF‐κB and Wnt [40, 41].
Other categories of natural products, such as quinones (e.g., juglone), saponins (e.g., ginsenosides), polysaccharides (e.g., Ganoderma lucidum polysaccharides), and organosulfur compounds (e.g., allicin), also exhibit unique anticancer activities, further enriching the source for anticancer drug research and development [9, 22, 42, 43].
The Functions of Natural Products Across Various Clinical Phases
2.2
Throughout the entire course of cancer treatment, natural products demonstrate unique value at different clinical stages due to their multitargeted regulation and relatively low toxicity. They play significant roles in primary prevention, concurrent treatment, postoperative recovery, maintenance therapy, and end‐of‐life care, offering adjunctive treatment strategies distinct from traditional chemotherapy (Table 1) [44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57].
Perioperative Period
2.2.1
During the perioperative period, the use of traditional chemotherapy drugs is often limited due to their significant toxic side effects, making it difficult to employ them safely for adjuvant interventions either preoperatively or shortly after surgery [58]. Furthermore, surgical trauma triggers systemic inflammatory responses and hypercoagulable states, which may create favorable conditions for residual or circulating tumor cells, thus increasing the risk of postoperative recurrence and metastasis [59]. In contrast, natural products hold unique value in this regard.
A recent clinical study in gastric cancer patients demonstrated that initiating oral administration of the Japanese Kampo medicine Rikkunshito immediately after minimally invasive gastrectomy (on postoperative Day 1) effectively mitigated weight loss, improved nutritional indicators (such as PNI and GNRI), and increased postoperative food intake [52]. The mechanism of action is closely linked to the multitargeted properties of natural products: Rikkunshito stimulates appetite by promoting ghrelin secretion and creates a more recovery‐friendly physiological environment by regulating gastrointestinal motility and hormone levels [60]. These findings confirm that natural products serve as a safe perioperative adjunct when combined with postoperative nutritional support regimens. They directly address surgery‐induced anorexia and malnutrition without significant side effects, offering a novel strategy beyond chemotherapy drugs to improve both short‐term outcomes and long‐term prognosis.
Concurrent Chemoradiotherapy
2.2.2
During the combined chemotherapy and radiotherapy phase, conventional treatments face numerous challenges, including multidrug resistance, adverse reactions limiting dosage, and severe immunosuppression, all of which constrain improvements in therapeutic efficacy [61].
Natural products demonstrate multifaceted synergistic advantages at this stage. They serve as radiosensitizers and chemotherapeutic sensitizers, enhancing efficacy while reducing toxicity. For example, a multicenter randomized clinical trial in non‐small cell lung cancer (NSCLC) demonstrated that the compound traditional Chinese medicine formulation Shengbai Oral Solution, when combined with platinum‐based chemotherapy, significantly reduced the incidence of chemotherapy‐induced neutropenia and decreased the use of granulocyte colony‐stimulating factor. Concurrently, this treatment markedly alleviated the burden of core symptoms reported by patients, such as fatigue and pain [62]. Modified peanut skin decoction (PSD) demonstrated efficacy in preventing and treating chemotherapy‐induced bone marrow suppression in patients with advanced squamous cell lung cancer. PSD significantly delayed the onset of Grade III–IV bone marrow suppression, shortened its duration, and improved patients’ quality of life [63]. These provide direct clinical evidence for natural products as chemotherapy adjuvants, enabling synergistic anticancer strategies that enhance efficacy while reducing toxicity.
The steroid alkaloid paimonin (PMI) from Fritillariae Thunbergii Bulbus synergizes with Oxa to significantly enhance cell death and inhibit gastric cancer cell proliferation. This combination activates the mitochondrial apoptosis pathway by upregulating Bax, inducing CYCS release, and cleaving caspase‐9, caspase‐3, and poly(ADP‐ribose) polymerase (PARP). Concurrently, it inhibits the RAS/PI3K/AKT survival pathway by suppressing Ras‐GTP and phosphorylated AKT [64]. Tanshinone IIA synergistically potentiates the cytotoxic effects of Olaparib in both wild‐type and BRCA‐deficient triple‐negative breast cancer cells. It induces apoptosis by increasing double‐strand breaks in these cells and subsequently disrupting ataxia–telangiectasia mutated stability, presenting a potential combination therapy approach with PARP inhibitors [65].
Postoperative and Recovery Phase
2.2.3
Postoperative patients often face challenges such as persistent immunosuppression, cancer‐related fatigue, and intestinal dysfunction, while conventional chemotherapy is strictly limited during this phase. Natural products demonstrate unique advantages during this period: Astragalus has been shown to possess anti‐inflammatory and antifibrotic properties. It inhibits the aggregation and activation of monocytes/macrophages while reducing TGF‐β1 production in the peritoneal cavity [66]. Ginsenoside Rg1 directly influences Th cell activity and Th1/Th2 system development, selectively enhancing glycocalyx transcription product expression, increasing IgA antibody production, and promoting humoral immunity [67]. In a melanoma mouse model, ginsenoside Rh2 induces substantial infiltration of CD4^+^ and CD8^+^ T lymphocytes into tumor tissues, enhancing immune responses [68]. Gegen Qinlian Tablets (GQT), a classical Chinese herbal formula, demonstrates synergistic effects with immune checkpoint inhibitor (ICI) therapy in patients with advanced NSCLC. GQT significantly reduces the incidence of immune‐related adverse events and delays their onset. Regarding antitumor efficacy, GQT also exhibits higher objective response rates and disease control rates [69].
Furthermore, growing research indicates that various natural compounds can improve immune function by altering tight junction structures, microbial metabolites, and gut microbiota composition [70]. For instance, diosgenin can improve gut microbiota composition. After diosgenin administration, the abundance of Sutterella and Lactobacillus species within the Proteobacteria phylum, as well as Bacteroides species within the Bacteroidetes phylum, significantly increased. Tumor tissues exhibited clear IFN‐γ production and CD4/CD8 T cell infiltration [71]. Treatment with icariin significantly increased probiotic bacteria such as Lactobacillus and Bifidobacterium, whose ability to promote antitumor immunity and enhance the activity of various lymphocyte subsets has been demonstrated [72]. Additionally, metabolites like short‐chain fatty acids and indole derivatives increased, exhibiting proven immunomodulatory functions and promoting host metabolic regulation via colonic Gpr41/Gpr43 and AhR signaling pathways [73]. These natural compounds can be integrated with nutritional support and exercise rehabilitation to construct a precise immunonutrition strategy, promoting functional recovery and reducing recurrence risk.
Maintenance and Metastasis Prevention
2.2.4
During the maintenance therapy phase, traditional chemotherapy struggles to achieve long‐term intervention due to its toxic side effects. In contrast, natural products, characterized by low toxicity and suitability for prolonged use, emerge as an ideal choice for maintenance therapy. Research indicates that genistein, the primary active component in soy isoflavones, inhibits tumor metastasis through a multitarget mechanism: on one hand, it downregulates the hypoxia‐inducible factor‐1α (HIF‐1α)/vascular endothelial growth factor (VEGF) signaling pathway to suppress tumor angiogenesis [74]; on the other hand, it modulates the Wnt/β‐catenin pathway to influence the epithelial–mesenchymal transition (EMT) process, thereby reducing tumor cell invasiveness [75]. Concurrently, resveratrol activates the deacetylase SIRT1, promoting metabolic reprogramming in tumor stem cells and inducing their transition toward a differentiated state [76]. Furthermore, it inhibits the activation of the NF‐κB signaling pathway, reducing the release of proinflammatory factors in the TME and altering the microenvironment that favors tumor stem cell maintenance [77]. These natural products with well‐defined molecular mechanisms can achieve sustained “micro‐control” of tumors through standardized dietary supplements, offering novel strategies for long‐term disease management.
Palliative and Adjuvant Therapy
2.2.5
In end‐stage cancer treatment, traditional chemotherapy often becomes unsuitable due to excessive toxicity burdens. Natural products frequently replace chemotherapeutic agents by leveraging their multitarget regulatory properties. Triterpenoids in medicinal Ganoderma lucidum exert comprehensive therapeutic effects through dual mechanisms: centrally, by activating μ‐opioid receptors and modulating the GABA ergic system for analgesic effects; and peripherally, by inhibiting COX‐2 and TNF‐α expression to provide anti‐inflammatory effects. They also improve cachexia‐related anorexia by stimulating ghrelin receptors [78]. The primary active component in Indian frankincense extract, boswellic acid, alleviates anxiety and depression by regulating hypothalamic–pituitary–adrenal axis function, reducing cortisol levels, and enhancing 5‐HT1A receptor activity in the prefrontal cortex [79]. These natural products, with well‐defined molecular mechanisms, not only delay disease progression but also significantly improve patients’ quality of life through multitarget regulation. They provide scientific evidence aligned with holistic medical principles for palliative care, achieving an optimal balance between disease control and quality of life.
Mechanistic Insights: Targeting Tumor Hallmarks With Natural Products
3
Natural products exert their anticancer activity by targeting key hallmark features of tumors. These compounds can modulate multiple core biological processes, including apoptosis and cell cycle arrest, angiogenesis and metastasis, and inflammation and oxidative stress [80, 81, 82]. Metabolic alterations in tumor cells play a pivotal role as a central mechanism influencing tumor growth. Natural products can precisely intervene in tumor cell metabolic reprogramming to exert anticancer activity [83].
Apoptosis and Cell Cycle Arrest
3.1
Apoptosis is a regulated form of cell death triggered in response to developmental cues or cellular stress. Disruption of apoptotic regulatory mechanisms leads to abnormal proliferation and survival [80]. This selective cell suicide plays a crucial role in numerous physiological and pathological processes, including development, immunity, and disease, where the elimination of damaged or redundant cells helps ensure organismal health [84]. Key apoptotic regulators include Bcl‐2 family proteins (Bcl‐2, Bax) and caspase family proteins (caspase‐3) [85, 86].
The BCL‐2 protein family modulates the activation of intrinsic apoptotic pathways in response to cellular stressors such as DNA damage, γ‐irradiation, oncogene activation, and growth factor withdrawal [87]. Certain natural products can modulate tumor cell apoptosis by influencing BCL‐2 protein levels [88]. Leaf extracts of Vitex doniana, particularly its dichloromethane extract, significantly downregulate the expression of the antiapoptotic gene Bcl‐2 in cervical carcinoma HeLa cells. This pivotal action relieves Bcl‐2's inhibition of the mitochondrial apoptosis pathway, thereby promoting the initiation of intrinsic apoptosis programmes in cancer cells and ultimately effectively suppressing tumour cell proliferation [89]. In DEN‐induced mouse models of hepatocellular carcinoma, leaf extracts from Haloxylon scoparium significantly downregulated the expression of the prosurvival protein Bcl‐2 while simultaneously upregulating levels of the proapoptotic protein BAX. Molecular docking studies confirmed that the flavonoid glycosides abundant in these extracts—such as quercetin and isorhamnetin glycosides—exhibit high‐affinity binding to targets including BCL‐2 [90]. Celastrol has been demonstrated to significantly promote apoptosis in MDA‐MB‐231 cells. Its mechanism of action involves directly targeting and binding to HSDL2, inhibiting its expression, which subsequently downregulates the level of the Bcl‐2 and upregulates the expression of Bax. Ultimately, this process induces apoptosis by suppressing the HSDL2/mitogen‐activated protein kinase (MAPK)/ERK signaling pathway [91].
As a member of the protease family, caspase is a key executor of cell death, participating in both the initiation and execution phases of apoptosis. Numerous pathological processes have been found to correlate with altered caspase activity or variations in the gene expression levels of these enzymes across different cancer types [92]. Among them, caspase‐3 plays a particularly crucial role. As a key protease in the clear apoptosis pathway, its absence or downregulation is associated with carcinogenic effects, indicating its potential as a biomarker for cancer prevention and treatment [93]. Research indicates that Moringa extract significantly inhibits EL4 lymphoma cell growth at a concentration of 100 µg/mL [94]. Tobacco leaf extract effectively suppresses the prosurvival EGFR/PI3K/AKT signaling pathway while activating the TP53‐mediated apoptotic cascade. This mechanism ultimately leads to caspase‐3 cleavage, thereby inducing cell apoptosis [95].
Meanwhile, mutations in cell cycle regulatory genes play a significant role in tumorigenesis. Under normal circumstances, DNA damage triggers cell cycle arrest at checkpoints, providing time for repair to minimize mutations and prevent tumor formation [96]. Zedoary turmeric oil exerts anti‐HCC effects primarily by modulating the EGFR/p53/Bcl‐2 signaling axis, inhibiting cell proliferation and migration, inducing apoptosis, and causing G1 phase cell cycle arrest [97]. The important anticancer natural product—paclitaxel—promotes the assembly of tubulin into microtubules and prevents their depolymerization, thereby further facilitating the formation of stable microtubules. It effectively blocks the G2/M phase of the cell cycle, prevents mitosis, and ultimately inhibits cancer cell proliferation or promotes apoptosis [98]. Podophyllotoxin, an aryltetralin‐type lignan isolated from Podophyllum plants, is widely used in the treatment of various cancers. Similar to colchicine, podophyllotoxin binds to the colchicine site at the α‐ and β‐tubulin interface, inhibits tubulin assembly into microtubules, blocks the cell cycle at the G2/M phase, and ultimately leads to mitotic arrest [99].
Anti‐angiogenesis and Metastasis Suppression
3.2
Angiogenesis refers to the process by which tumor cells obtain increased nutrient and oxygen supply through the formation of new blood vessels, which also facilitates their metastasis to other sites via the circulatory system. Therefore, interventions targeting tumor angiogenesis have become a key strategy in cancer treatment [81].
Natural products exert their antiangiogenic effects primarily by inhibiting VEGF and NF‐κB expression, as well as suppressing vascular endothelial cell (EC) growth. The flavonoid natural product naringenin possesses certain antiangiogenic properties. Research indicates that naringenin first reduces interactions among angiogenic factors by inhibiting the secretion of inflammatory cytokines such as IL‐6 and MCP‐1. It then directly suppresses the tyrosine kinase activity of the VEGFR, thereby inhibiting the phosphorylation of related cytokines like FAK and Akt in downstream signaling cascades. and downregulates estrogen receptor alpha to inhibit VEGF production, thereby mediating its antiangiogenic effects [100]. Additional studies indicate that treatment of prostate cancer cells with Tripterygium wilfordii homospermeolide significantly reduces VEGF and COX‐2 expression, inhibiting angiogenesis while enhancing autophagy signaling intensity in PC tissues [101]. Furthermore, dihydroartemisinin(DHA) exhibits potent antiangiogenic activity by blocking NF‐κB p65 nuclear translocation and specifically downregulating VEGF‐2 expression in ECs, thereby demonstrating antiangiogenic properties. Consequently, DHA represents an ideal candidate for use as an angiogenesis inhibitor in tumor therapy [102].
Cancer metastasis is a major cause of cancer mortality, and it begins with the degradation of extracellular matrix (ECM) proteins, enabling tumor cells to enter capillaries or lymphatic vessels and infiltrate various tissues throughout the body [103]. Proteases that promote ECM degradation, such as matrix metalloproteinases (MMPs), are considered enzymes that facilitate metastasis [104]. Compounds from flavonoids, isoflavonoids, cannabinoids, chalcones, naphthoquinones, terpenoids, alkaloids, steroids, and saponins have been identified to inhibit tumor invasion and metastasis. The antimetastatic mechanisms of many natural compounds are found to be associated with MMP inhibition [105]. Studies indicate that evodiamine significantly inhibits the invasion of two nasopharyngeal carcinoma cell lines (HONE1 and CNE1) while only mildly affecting cell proliferation. Evodiamine treatment markedly reduces MMP‐2 mRNA and protein levels, potentially through reduced translocation of nuclear factor‐κB p65, but does not affect MMP‐9 expression levels [106]. According to extensive literature, the anticancer effects of baicalin may be related to inhibiting cell metastasis and invasion [107, 108]. Dose‐dependent anti‐invasive activity of baicalin was observed in OC2 human oral cancer cells and MDA‐MB‐231 human breast cancer cells, accompanied by downregulation of MMP‐2, MMP‐9, and uPA [108, 109].
Targeting Inflammation and Oxidative Stress
3.3
Inflammation is one of the primary determinants of cancer, and inflammatory responses are also key characteristics of cancer. Inflammatory cells—including macrophages, dendritic cells (DCs), and neutrophils—are vital components of the TME. Cancer cells can also release large quantities of cytokines and chemokines, which summon immune cells and further exacerbate inflammation, demonstrating the crucial link between inflammation and cancer [110]. Research indicates that chronic inflammation increases cancer risk [111]. For instance, chronic bronchitis elevates lung cancer risk; chronic pancreatitis can induce pancreatic cancer; and Helicobacter pylori load is a major determinant of gastric cancer [112, 113, 114]. Natural products play a vital role in anti‐inflammatory and antioxidant stress mechanisms, holding significant potential for cancer treatment by modulating inflammatory responses.
Puerarin, extracted from kudzu root, has demonstrated anti‐inflammatory effects across various disease models. Puerarin elevated IL‐2 and superoxide dismutase activity in plasma of U14 cervical carcinoma mice. At specific doses, it eliminated excess free radicals [115]. Reports indicate puerarin mitigated reactive oxygen species (ROS)‐induced tissue damage, subsequently enhancing antitumor efficacy [116]. Luteolin has been found to possess multiple therapeutic effects. It promotes tumor cell apoptosis by inhibiting NF‐κB activation through TNF‐α suppression and enhancing natural killer (NK) cell activity [117]. EGCG is renowned for its diverse pharmacological activities. As a catechin compound, EGCG simultaneously inhibits COX‐2 by suppressing NF‐κB activation in HeLa and SiHa cells while regulating ROS levels, suggesting its antitumor effects may operate through antioxidant pathways [118]. In line with this mechanism, extracts from the traditional medicinal plant Rhus punjabensis Stewart and its isolated major triterpenoid compounds (such as lupeol) significantly inhibit the NF‐κB inflammatory signaling pathway, effectively scavenge free radicals (as confirmed by the DPPH assay), and induce apoptosis in cancer cells, demonstrating remarkable anticancer potential [119].
Metabolic Reprogramming as a Key Step in Natural Product‐Based Cancer Therapy
3.4
Metabolic reprogramming serves as the core driver of natural product‐targeted tumor therapy mechanisms. The defining characteristics of cancer cells lie not only in their uncontrolled proliferation but also in their unique energy metabolism patterns [120]. The classic “Warburg effect” reveals that even under oxygen‐rich conditions, cancer cells tend to undergo high‐rate glycolysis. This phenomenon signifies that metabolic reprogramming has become one of the core hallmarks of cancer, enabling tumor cells to sustain rapid proliferation, resist cell death, and adapt to microenvironmental stress [121]. Consequently, targeting these abnormal metabolic pathways has emerged as a highly promising anticancer strategy. Structurally diverse, multitargeted natural products can exert their anticancer effects by precisely intervening in key metabolic processes of cancer cells [83]. Figure 2 integrates the multitarget mechanisms of natural products’ anticancer effects.
Multitarget antitumor mechanisms of natural products: from metabolic reprogramming to tumor hallmarks. Abbreviations: API: apigenin, Tan IIA: tanshinone IIA, ANDRO: andrographolide, QUE: quercetin, RES: resveratrol, BCL: baicalin, WOG: wogonin, CLS: celastrol, CUR: curcumin, BBR: berberine, EGCG: epigallocatechin‐3‐gallate, BAI: baicalein, SK: shikonin, RAPA: rapamycin, SLM: silymarin, LUT: luteolin.
Regarding glucose metabolism, cancer cells heavily rely on aerobic glycolysis, and natural products can effectively inhibit key nodes in this process. For instance, berberine has been demonstrated to downregulate glucose transporter 1 (GLUT1) expression and inhibit hexokinase 2 (HK2) activity [122]. Concurrently, flavonoids like quercetin interfere with pyruvate kinase M2 (PKM2) function, effectively suppressing glycolytic flux [123]. Ferulic acid reduces GLUT1 expression and glucose uptake in cancer cells, while silymarin competitively inhibits GLUT4, offering pathways to restrict nutrient supply and inhibit tumor growth [124, 125]. Regarding lipid metabolism, cancer cells enhance lipid synthesis to meet the rapid demands of cell membrane construction. Among dietary flavonoids, EGCG from green tea exhibits significant fatty acid synthase (FASN) inhibitory activity, primarily through its galloyl group specifically binding to the ketone reductase domain of FASN. Despite EGCG's multifaceted effects on tumor cells, its FASN‐targeting properties make it a highly promising lead compound for chemoprevention and therapy [126]. Additionally, natural products play crucial roles in glutamine metabolism. Glutaminase 1 (GLS1), a key enzyme in glutamine degradation, can be downregulated by compounds like CUR and quercetin. This deprives cancer cells of essential nitrogen and carbon sources, thereby inhibiting their proliferation [127, 128]. From alkaloids to polyphenols, diverse classes of natural products effectively reverse cancer cell metabolic reprogramming through multitargeted, multilevel mechanisms.
The Role of Flavonoids in Cancer Metabolic Reprogramming
4
Flavonoids, as important natural products, exert significant anticancer effects by targeting tumor metabolic reprogramming processes. These compounds can systematically regulate core pathways such as glucose metabolism, lipid metabolism, and glutamine metabolism, thereby influencing energy supply and biosynthesis in tumor cells [129]. As multitarget metabolic modulators, they hold significant application potential in cancer therapy.
Structural Types and Sources
4.1
As natural compounds, flavonoids are representative compounds that exert antitumor effects by regulating abnormal metabolic networks. By targeting key metabolic pathways—including glycolysis, lipid synthesis, glutamine utilization, and nucleotide metabolism—flavonoids disrupt both the energy supply and biosynthetic demands essential for tumor survival and progression [129]. Uniquely, flavonoids can simultaneously influence cancer cell energy metabolism, redox homeostasis, and intracellular signaling cascades, underscoring their versatility as metabolic regulators in cancer therapy [130].
Structurally, over 4000 flavonoid compounds have been identified in plants. All flavonoids share a core backbone composed of two aromatic benzene rings (A and B) connected by a three‐carbon bridge forming a heterocyclic pyran ring (C), commonly described as the C6–C3–C6 skeleton [33]. Variations in hydroxylation, methoxylation, glycosylation, and prenylation patterns give rise to multiple subclasses, including flavonols, flavones, isoflavones, flavanones, flavanols (catechins), chalcones, anthocyanins, and flavanonols [131]. This structural diversity is closely linked to their biological activities and influences their capacity to target various aspects of tumor metabolism.
Different flavonoid subclasses exert distinct effects on cancer‐related metabolic pathways. For instance, flavanols such as EGCG modulate receptor tyrosine kinases, lipid rafts, fatty acid metabolism, and epigenetic regulators, and also target proteasomes, telomerase, and cancer stem cells [132]. Isoprenylated flavonoids and chalcones influence carcinogen metabolism by inhibiting phase I metabolizing enzymes and activating phase II detoxification systems, while also exerting anti‐inflammatory and antiproliferative effects [133]. Flavonols such as kaempferol demonstrate antiproliferative activity in lung and endothelial cancer cells through MAPK signaling modulation [134]. Polymethoxyflavones derived from citrus peels have been shown to inhibit ovarian cancer cell growth by downregulating Akt, HIF‐1α, NF‐κB, and VEGF, thereby impairing angiogenesis [135].
Anthocyanins also play key roles in metabolic regulation. These pigments suppress cancer cell migration and reverse EMT by inactivating PI3K/Akt signaling in hepatocellular carcinoma cells [136]. Certain anthocyanin derivatives, such as geranylgeranyl anthocyanins, induce G2/M cell cycle arrest, mitochondrial dysfunction, and ROS‐mediated autophagy, while inhibiting p‐PI3K and p‐Akt in a dose‐dependent manner [137].
Together, the structural diversity and functional specificity of flavonoids enable them to act on multiple metabolic and signaling axes in cancer cells. These properties make flavonoids a promising class of natural compounds for innovative anticancer strategies. Their ability to selectively target tumor metabolism while maintaining low systemic toxicity offers considerable therapeutic potential for both prevention and adjuvant cancer treatment [138].
Mechanism of Action: Glucose, Lipid, and Glutamine Metabolism
4.2
Flavonoids play a crucial role in inhibiting tumor growth by interfering with the metabolic reprogramming of cancer cells. Their regulatory effects are primarily manifested in three core pathways: glucose metabolism, lipid metabolism, and glutamine metabolism [129].
Inhibition of Glucose Metabolism
4.2.1
The aerobic glycolysis characteristic of cancer cells is a hallmark of their metabolic reprogramming. This metabolic shift enables cancer cells to rapidly generate ATP while acquiring abundant biosynthetic precursors to meet their proliferative demands [139]. Figure 3 demonstrates that flavonoids effectively reverse this metabolic abnormality through multitargeted action.
Central regulatory network of flavonoids targeting glucose‐driven metabolic reprogramming in cancer cells. Abbreviations: GEN: genistein, QUE: quercetin, SLB: silybin, MAT: matrine, WOG: wogonin, Tan IIA: tanshinone IIA, chrysin: CHR, EGCG: epigallocatechin‐3‐gallate, BAI: baicalein, SAP: sapogenin, API: apigenin, CUR: curcumin, LUT: luteolin, KAE: kaempferol.
At the molecular mechanism level, apigenin significantly downregulates the expression and activity of PKM2 by inhibiting the Akt/mTOR signaling pathway. As a key regulatory enzyme in glycolysis, reduced PKM2 activity directly leads to decreased glycolytic flux, triggering ATP depletion and NADH/NAD^+^ imbalance. This downregulates antiapoptotic proteins such as Mcl‐1, thereby promoting mitochondrial apoptosis [140]. Concurrently, ferulic acid reduces glucose transporter GLUT1 expression, curtailing glucose uptake at its source [124]. Notably, catechin inhibits lactate dehydrogenase A (LDHA), blocking pyruvate conversion to lactate. This not only elevates intracellular pH but also triggers substantial ROS accumulation, ultimately inducing apoptosis [141].
Furthermore, flavonoids exert global regulation of glycolysis by modulating key transcription factors. For instance, quercetin and EGCG activate the AMPK pathway to inhibit the stability and transcriptional activity of HIF‐1α [142, 143]. This simultaneously impairs glycolysis while inducing cell cycle arrest and senescence. Meanwhile, flavonoids like Oroxylin A stabilize the p53 protein, enhancing its suppression of glycolysis‐related genes [144]. Collectively, these actions substantially diminish cancer cells' glycolytic capacity, reshaping their energy metabolism patterns.
Regulate Lipid Metabolism
4.2.2
Lipid metabolism reprogramming is another key feature enabling cancer cells to sustain rapid proliferation. By enhancing the synthesis of fatty acids, cholesterol, and phospholipids, cancer cells provide the material foundation for cell membrane biogenesis, signaling, and energy storage [145]. Flavonoids exhibit multifaceted regulatory roles in this process.
Regarding fatty acid metabolism, EGCG specifically binds to the ketone acyl reductase domain of FASN via its unique galloyl group, effectively inhibiting the enzyme's catalytic activity. As the key enzyme for de novo fatty acid synthesis, FASN inhibition directly impedes fatty acid production [126]. Studies indicate that flavonoids such as quercetin, kaempferol, and luteolin exhibit similar FASN inhibitory activity, suggesting this may represent a common mechanism of action for flavonoids [146, 147, 148]. As the key rate‐limiting enzyme in de novo fatty acid synthesis, inhibition of FASN directly blocks fatty acid supply, leading to impaired membrane lipid synthesis and disruption of lipid rafts. This induces endoplasmic reticulum stress and activates the mitochondrial apoptosis pathway [149].
In cholesterol metabolism regulation, flavonoids modulate the transcription of cholesterol synthesis‐related genes by influencing the activity and expression of SREBPs. Quercetin and EGCG interfere with SREBP cleavage and nuclear translocation, thereby suppressing the expression of key enzymes like 3‐hydroxy‐3‐methylglutaryl‐coenzyme A reductase (HMGCR)[150]. Additionally, these compounds activate liver X receptor, promoting cholesterol reverse transport and further maintaining cholesterol homeostasis.
At the phospholipid metabolism level, resveratrol reduces arachidonic acid release and subsequent inflammatory mediator production by inhibiting PLA2 activity [151]. This effect blocks prosurvival signaling pathways and reduces levels of antiapoptotic factors such as PGE_2_, thereby sensitizing cells to death signals like TNF‐α [152]. Simultaneously, quercetin interferes with phosphatidylcholine synthesis by suppressing choline kinase activity, directly affecting cell membrane biosynthesis and associated signaling pathways [153]. This blocks cell membrane biosynthesis and disrupts lipid second messenger balance, leading to cell cycle arrest in the S phase [154].
Disrupt Glutamine Metabolism
4.2.3
Glutamine serves as a crucial carbon and nitrogen source for cancer cells, playing a vital role in sustaining their proliferation [155]. Flavonoids effectively disrupt the metabolic balance of cancer cells by interfering with different stages of glutamine metabolism.
Silymarin specifically binds to isocitrate dehydrogenase 1 (IDH1), inhibiting its catalytic activity. IDH1 catalyzes the oxidative decarboxylation of isocitrate to form α‐ketoglutarate (α‐KG), forming a critical link between glutamine metabolism and the tricarboxylic acid cycle. Inhibiting IDH1 activity alters the NADP^+^/NADPH ratio, elevates oxidative stress levels, and ultimately suppresses tumor cell proliferation [156].
Notably, baicalin achieves dual inhibition of glutamine metabolism by suppressing the mTOR signaling pathway and downregulating glutamine transporter expression. In lung cancer models, baicalin treatment significantly reduced intracellular glutamine utilization and inhibited tumor growth (Figure 4) [157]. This multitargeted action deprives tumor cells of their synthetic precursors and energy sources, inducing autophagic cell death by activating the AMPK/p53 axis.
Baicalein derived from Scutellaria baicalensis inhibits glutamine uptake and mTOR signaling, leading to metabolic disruption and apoptosis in lung cancer cells. Reproduced with permission [157]. Published by Elsevier Inc.
In addition to targeting enzymatic regulators, flavonoids also interfere with glutamine uptake and downstream catabolism. Flavokawain A, a chalcone derived from Piper methysticum (kava), significantly reduces intracellular levels of glutamine, glutamate, and proline in prostate cancer cells. This metabolic suppression leads to a reduction in glutathione (GSH) synthesis, triggering oxidative stress and inducing apoptosis via ROS accumulation [158].
Metabolic Reprogramming of Other Categories of Natural Products and Their Anticancer Activity
5
Natural products play a crucial role in modulating the metabolic reprogramming of cancer cells. These compounds regulate key processes such as tumor cell energy metabolism, biosynthesis, and redox balance, contributing to their anticancer activity [159]. A comprehensive overview of how various natural products influence tumor metabolism is summarized in Table 2 [127, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188], highlighting the diverse mechanisms through which they exert their therapeutic effects. In addition to flavonoids, a wide range of structurally diverse natural products also target metabolic pathways to interfere with cancer cell reprogramming, offering unique mechanisms of action for cancer therapy [83].
Alkaloids
5.1
Alkaloid compounds exhibit significant activity in regulating tumor metabolism [189]. Berberine, as a representative of isoquinoline alkaloids, effectively disrupts glucose metabolism and lipid synthesis in tumor cells by inhibiting mitochondrial complex I function and activating the AMPK signaling pathway, thereby inducing metabolic stress, and this energy and synthetic impairment ultimately triggers oxidative stress and mitochondrial dysfunction, leading to cancer cell apoptosis [190]. Research confirms that physapubescin modulates glutamine metabolism by inhibiting kidney‐type glutaminase activity, leading to reduced intracellular α‐KG levels and exerting antitumor effects [191].
Terpenoids
5.2
Terpenoids exert multitarget regulatory effects in metabolic reprogramming. DHA significantly reduces the viability of JF‐305 cells, arrests the cell cycle at the G2/M phase, and induces apoptosis by diminishing mitochondrial membrane potential and ROS accumulation. Concurrently, it suppresses tumor cell glucose uptake, lactate production, and ATP generation, while also downregulating the Akt/mTOR signaling pathway and the expression of GLUT1 [192]. Triptolide, a primary active compound isolated from Tripterygium wilfordii Hook. F., exhibits significant anticancer potential by promoting cell death and autophagy in lung and prostate cancer cells through the activation of CaMKKβ–AMPK signaling and modulation of the AMPK/mTORC2 axis [193].
Polyphenols
5.3
Other polyphenolic compounds demonstrate unique advantages in regulating tumor metabolic reprogramming. CUR not only inhibits FASN and ATP citrate lyase, but also reduces GLUT1 and HK2 expression by modulating c‐Myc [194, 195]. These effects synergistically impair cancer cells’ lipid synthesis and glycolytic capacity, leading to membrane structural damage and energy depletion, which in turn trigger apoptosis and cell cycle arrest. Green tea polyphenol EGCG inhibits glucose transporter and lactate dehydrogenase activity, while ellagic acid interferes with the expression of key enzymes in glutamine metabolism [196, 197]. Furthermore, studies indicate that water extracts of Chamerion angustifolium L. and its primary polyphenolic component oenothein B can inhibit colon cancer cells by targeting mitochondrial energy metabolism. Both agents reduce Caco‐2 cell viability in a dose‐dependent manner and significantly suppress oxidative phosphorylation function in mitochondrial complexes I and II [198]. These polyphenolic compounds effectively disrupt tumor cell metabolic networks through multitarget synergistic actions.
Sulfur Compounds
5.4
Sulfur‐containing natural products play a significant role in metabolic regulation due to their unique mechanisms of action. Allicin specifically inhibits HK2 and PKM2 activity while influencing lipid synthesis by activating the AMPK pathway [199]. Sulforaphane not only enhances antioxidant capacity through the Nrf2 pathway but also promotes mitochondrial metabolism by suppressing PDK activity. By inhibiting PDK to promote mitochondrial oxidative phosphorylation, sulforaphane may induce metabolic imbalance while enhancing oxidation, ultimately eliminating cancer cells through activation of apoptotic pathways or ferroptosis [200]. Furthermore, glucosinolate metabolites in broccoli inhibit histone deacetylases, affecting metabolic gene expression; while sulfur compounds in onions disrupt GSH metabolism, amplifying oxidative stress [201, 202]. These sulfur‐containing compounds offer unique targets for modulating tumor metabolism.
Synergistic Metabolic Regulation Strategy
5.5
The combined use of different categories of natural products can produce synergistic metabolic regulatory effects. Traditional Chinese medicine formulas, as typical representatives of multicomponent natural products, demonstrate unique advantages in synergistically regulating tumor metabolism. For example, Huangqi Fuling Decoction (HQFLD) synergistically targets the RAS–MEK–ERK signaling pathway through its multiple active components (such as astragaloside IV and astragaloside V) to inhibit gastric cancer cell proliferation and metastasis. It also downregulates key metabolism‐related genes including VCAM1, ICAM1, PTGS2, thereby disrupting tumor energy metabolism and cell adhesion processes [203]. This multitargeted intervention collectively impairs the tumor's energy supply, invasive capacity, and inflammatory microenvironment, ultimately enhancing tumor growth suppression through metabolic collapse and enhanced apoptotic signaling.
Another compound LTTL, regulates serum microRNAs (e.g., miR‐2110, miR‐7d‐3p) associated with glucose metabolism pathways (such as glycolysis/gluconeogenesis, fructose, and mannose metabolism). This modulates metabolic reprogramming processes related to lung cancer pain, demonstrating the compound's multitarget synergistic effects within the metabolism–immunity–pain regulatory network. miR‐7d‐3p to influence metabolic reprogramming processes associated with lung cancer pain, demonstrating the multitarget synergistic effects of the compound within the metabolic–immune–pain regulatory network [57].
These studies demonstrate that traditional Chinese medicine formulas, through the synergistic effects of multiple natural products, can simultaneously intervene in several key aspects of tumor metabolism—such as energy metabolism, oxidative stress, and signaling pathway activation—thereby more effectively addressing the heterogeneity and plasticity of tumor metabolism.
Regulation of the TME by Natural Products
6
The TME is a complex ecosystem composed of tumor cells, immune cells, stromal cells, vascular networks, and ECM [204]. Through intricate interactions, these components collectively influence cancer progression, immune evasion, and therapeutic resistance [205]. Recent studies have revealed that natural products can effectively modulate various components within the TME via multitarget mechanisms, thereby inhibiting tumor progression [206].
Cancer‐Associated Fibroblasts
6.1
CAFs are key effector cells in the tumor stroma that actively participate in ECM remodeling by secreting multiple cytokines and growth factors, thereby promoting tumor growth and metastasis [207]. Studies indicate that Huaier can significantly reduce the expression levels of α‐smooth muscle actin and fibronectin in CAFs by inhibiting the TGF‐β/Smad signaling pathway, thereby suppressing the activation process of CAFs [208]. Tripterygium glycosides can inhibit the TGF‐β signaling pathway in CAFs, reversing their activated state to a quiescent state. This reduces ECM secretion and disrupts their role in supporting tumor growth [209]. Additionally, the traditional Chinese medicine compound Qingre Huayu Jianpi decoction has been demonstrated in colorectal cancer studies to effectively inhibit the activation and migration of CAFs, reduce the deposition of ECM components such as collagen, and suppress the growth of tumor organoids by regulating the Wnt/β‐catenin pathway. This highlights the comprehensive advantages of natural multicomponent drugs in synergistically regulating both tumor cells and CAFs [210].
Endothelial Cells
6.2
Tumor‐associated ECs support tumor angiogenesis through unique metabolic reprogramming, a process critical for tumor growth and metastasis [211]. CUR effectively inhibits the phosphorylation of VEGFR2, blocking the activation of downstream angiogenic signaling pathways [212]. Berberine B induces EC apoptosis, specifically disrupting the structural integrity of the tumor vascular network [213]. Notably, the natural product deoxypodophyllotoxin isolated from Anthriscus sylvestris promotes cytoskeletal remodeling in human umbilical vein ECs by activating the AMPK signaling pathway, thereby inhibiting angiogenesis and blocking tumor growth and metastasis [214]. Recent studies indicate that betulin, a triterpenoid compound derived from white birch bark, reduces mRNA stability by inhibiting m6A modification of PAX2, thereby decreasing PAX2 expression. This process dissociates PAX2 from the VEGF‐A promoter, ultimately blocking the VEGF‐A/VEGFR2 signaling pathway. This process inhibits EC migration and tubulogenesis in vitro, reduces tumor microvascular density in vivo, and thereby effectively suppresses tumor growth and metastasis [215]. These natural products stimulate ECs, collectively exerting an inhibitory effect on tumor angiogenesis.
ECM Components
6.3
During tumor progression, the ECM undergoes a series of dynamic changes that profoundly influence the physical properties and biochemical characteristics of the TME [216]. Flavonoids including lignans counteract these effects by inhibiting MMP expression through modulation of Notch signaling and specific microRNAs (e.g., miR‐21) [217]. Compounds like hypericin further suppress MMP‐3, MMP‐13, and ADAMTS5 expression in response to TNF‐α, thereby preserving ECM integrity and restricting tumor cell access to metabolic resources [218]. Collectively, these actions maintain the structural and functional integrity of the ECM, creating favorable conditions for suppressing tumor invasion and metastasis.
Diverse Tumor‐Infiltrating Immune Cells
6.4
Natural products exert extensive regulatory effects on immune cells within the TME [219]. Icariin promotes the maturation process of DCs, enhancing their antigen‐presenting function [220]. Arctigenin activates the Toll‐like receptor signaling pathway in macrophages, enhancing their phagocytic and cytotoxic capabilities [221]. The traditional Chinese medicine compound Jianpi‐Huayu Decoction has been shown to enhance NK cell cytotoxicity by suppressing TREM1/DAPI2 signaling in TAMs, thereby alleviating immunosuppression in the TME and synergizing with PD‐1 inhibitors in hepatocellular carcinoma [222]. Tetrandrine, derived from Stephania tetrandra S. Moore, is a bisbenzylisoquinoline alkaloid with the ability to activate the STING/TBK1/IRF3 pathway, promoting CCL5 and CXCL10 production. This enhances the infiltration of macrophages, DCs, and CD8 T cells in the TME, significantly inhibiting the growth of NSCLC [223]. Collectively, these immunomodulatory effects amplify the body's antitumor immune response, offering novel insights for tumor immunotherapy.
The Regulatory Effects of Natural Products on the Immune System
7
Natural products modulate tumor immune responses through multiple pathways and targets, not only directly enhancing immune cell function but also effectively reversing immune suppression in the TME, thereby strengthening the body's antitumor immune capacity [224] (Figure 5).
Multitarget mechanisms of natural products in tumor immunology. Abbreviations: Tan IIA: tanshinone IIA, GAS: gastrodin, ICS II: icariside II, API: apigenin, XCHT: Xiao Chai Hu Tang, QUE: quercetin, GGE: Glycyrrhiza glabra extract, KDC: Llex tarajois (Kudingcha), PHY: physapubescin I, GEN: genistein, ORI: oridonin, KAE: kaempferol, CUR: curcumin, RES: resveratrol, CAD: cardamonin, ATL1: atractylenolide I, LicA: licochalcone A, MEL: melitidin.
Tumor‐Associated Macrophages
7.1
Ginsenoside Rb1 promotes macrophage polarization toward the M1 phenotype with antitumor functions by regulating the NF‐κB signaling pathway, while simultaneously enhancing phagocytic activity [225]. Soy isoflavones, conversely, inhibit the polarization of macrophages toward the tumor‐promoting M2 phenotype, reducing the secretion of immunosuppressive factors [226]. Notably, resveratrol influences the metabolic reprogramming of macrophages by modulating the AMPK/mTOR signaling pathway [227]. In the context of STAT pathway‐mediated M2 polarization driving an aggressive tumor phenotype, Garcinone E blocks the M2 polarization process by inhibiting STAT6 phosphorylation, while Mongolian Iris Extract specifically targets the IL‐10/STAT3/PD‐L1 axis to reverse M2‐mediated immune evasion [228, 229]. Collectively, these actions reverse macrophage‐mediated immunosuppression, creating favorable conditions for improving the TME.
Dendritic Cells
7.2
Berberine promotes DC maturation and antigen presentation by activating the AMPK signaling pathway [230]. Research indicates that quercetin improves the immune function of DCs by influencing glucose metabolism [231]. These studies hold significant reference value for the development of whole‐cell nanovaccines targeting DCs. The GGT nanovaccine carrier, based on the self‐assembly of chitin oligosaccharides and Ganoderma lucidum polysaccharides, activates systemic antitumor immunity across multiple tumor models by efficiently delivering antigens to DCs and promoting their maturation [232]. Collectively, these effects amplify the pivotal role of DCs in antitumor immune responses.
NK Cells
7.3
Apigenin enhances NK cells’ ability to recognize and kill tumor cells by regulating the expression of surface activation receptors [233]. EGCG promotes NK cell production of perforin and granzyme, thereby amplifying their cytotoxic effects [234]. Furthermore, allicin improves mitochondrial function, thereby enhancing NK cell metabolic activity and sustained killing capacity [235]. The natural alkaloid piperlongumine significantly enhances tumor cell sensitivity to NK cell‐mediated killing by inducing ROS accumulation within tumor cells, disrupting protein homeostasis, and enhancing NK cell binding to tumor cells. This provides new evidence for utilizing natural products to synergistically enhance NK cell‐based immunotherapy [236].
T Lymphocytes (CD4+ and CD8+ T Cells)
7.4
Trichosanthin can alleviate immunosuppression by modulating the TGF‐β signaling pathway and inhibiting the function of regulatory T cells [237]. CUR promotes the proliferation and effector functions of CD8^+^ T cells, enhancing cytotoxic T lymphocyte responses [238]. Artesunate targets Ido1 to elevate tryptophan levels, thereby inhibiting NFATc1‐mediated PD1 transcription and activating CD8 T cells [239]. The structurally unique arabinogalactan‐polysaccharide fraction NLBPE1, isolated and purified from Lycium barbarum, induces proinflammatory cytokine release by activating the TLR4–IKK–NF‐κB signaling axis in DCs. This efficiently initiates antigen‐specific CD8^+^ T cell responses, demonstrating potent tumor suppression and preventive effects across multiple melanoma models [240]. Collectively, these actions enhance T cell‐mediated specific antitumor immune responses.
Myeloid‐Derived Suppressor Cells
7.5
Myeloid‐derived suppressor cells (MDSCs) promote immune evasion by inhibiting T cell activity in the TME. Their function is closely associated with lipid uptake and accumulation, a mechanism that enhances their immunosuppressive phenotype [241]. Studies indicate that soy isoflavones reduce lipid production in the TME, potentially weakening the function of MDSCs and reactivating T cell responses [226]. The traditional Chinese medicine formulation HBK, composed of 17 herbs and foods, modulates tumor immunity by promoting MDSC reduction through inhibition of the TLR3/NF‐κB signaling pathway [242]. Among bioactive plant‐derived compounds, sanguinarine has broad therapeutic applications in lung cancer. It inhibits M2 macrophage polarization through the Wnt/β‐catenin pathway, thereby blocking angiogenesis in lung cancer. Additionally, it induces the differentiation of MDSCs into macrophages and DCs via the NF‐κB pathway [243].
Delivery Systems and Formulation Innovation
8
Innovation in natural product delivery systems is key to enhancing their clinical efficacy. Intelligent delivery systems responsive to tumor metabolism and microenvironments, novel nanocarriers for oral and mucosal administration, and combined delivery systems integrating natural products with chemotherapy or immunotherapy collectively demonstrate the pivotal role of delivery technology innovation in advancing the clinical translation of natural products [244, 245, 246].
Natural Product‐Targeted Delivery Systems: Tumor Metabolism and Microenvironmental Adaptation
8.1
The therapeutic efficacy of natural products is often limited by their poor bioavailability and nonspecific distribution. To address these challenges, researchers are developing advanced delivery systems targeting tumor‐specific metabolic pathways and microenvironmental characteristics. These systems leverage the unique properties of tumor cells and their surrounding environment to achieve targeted delivery [247, 248].
Regarding tumor metabolic adaptation, natural product delivery systems can be engineered to target key metabolic pathways. For example, a nanocarrier loaded with mangiferin and CUR can effectively regulate tumor metabolism and induce apoptosis by synergistically inhibiting the PI3K/Akt/mTOR pathway in ovarian cancer. This nanomedicine also significantly enhances drug bioavailability and therapeutic efficacy [249]. Collagen‐modified nanoliposomes loaded with ginkgo biloba leaf and green tea extracts can induce apoptosis and arrest tumor cell cycles at the G0/G1 phase, demonstrating significant efficacy against hepatocellular carcinoma and colorectal cancer [250]. MPEG‐b‐PLA diblock copolymer micelles encapsulate the natural anticancer compound podophyllotoxin and its source, juniper extract. This carrier not only significantly enhances the drug's water solubility and stability but also selectively induces apoptosis in skin squamous cell carcinoma cells by inducing cell cycle arrest, activating caspase‐3, and generating ROS [251].
In terms of microenvironment adaptation, intelligent delivery systems can respond to specific physicochemical conditions within the TME to achieve precise drug release. By engineering nanoparticles to modulate their surface chemistry and functional groups for binding to receptors overexpressed on senescent cells, resveratrol‐loaded nanoparticles enhance targeting and uptake efficiency toward immune‐senescent cells within the TME [252]. The highly CAF‐specific nanomedicine system (Dex‐GP‐DOCA, DPD) developed by Huo's team responds to fibroblast activation protein‐alpha overexpressed on CAF surfaces, enabling precise release of the flavonoid compound quercetin. This targeted delivery achieves multiple effects including ECM degradation, vascular normalization, reversal of hypoxia resistance, and blockade of Wnt16 paracrine signaling [253]. Collectively, these cases demonstrate that microenvironment‐responsive smart delivery mechanisms can effectively intervene in tumor metabolism and matrix processes, enabling multifaceted synergistic regulation of the TME.
Nanocarriers for Oral and Mucosal Delivery of Natural Products
8.2
Natural products primarily originate from food and medicinal substances. Their oral administration method aligns closely with traditional medication habits, offering patients convenience and high compliance. Moreover, after absorption through the gastrointestinal mucosa, oral drugs can activate mucosal immune responses and leverage the mucosal system's efficient transport mechanisms to enhance systemic absorption. This approach avoids the trauma of injections while capitalizing on the unique advantages of the oral route [254]. However, the poor solubility and instability of natural products in the gastrointestinal tract present significant challenges. Nanocarrier systems offer a highly promising solution for enhancing oral bioavailability and enabling mucosal drug delivery.
Multiple nanotechnology approaches have been developed for oral delivery, including polymeric micelles (NPs) loaded with naringenin (NAR) designed for oral cancer treatment. Both free oral NAR and NAR‐NPs reversed lipid peroxidation and antioxidant status in DMBA‐treated animal buccal mucosal tissues. Compared with free NAR, NAR‐NPs exhibited higher antilipoperoxidation, antiproliferative, and antioxidant activities. NAR nanoparticles demonstrated significant chemopreventive potential by limiting or reducing abnormal cell growth in buccal mucosa through disruption of DMBA‐induced metabolic activation [245]. Inulin (IN) is a polymer with colon‐specific targeting capability, while hyaluronic acid (HA) targets CD44 on the surface of colon cancer cells. Existing research has developed an oral‐colon targeted delivery system (5‐Flu/MET@MSNs/Ce6@HIL) with HA and IN as key components, effectively inhibiting tumor growth [255].
Mucosal delivery systems offer an alternative pathway for local and systemic effects, enabling rapid absorption of natural products through the oral mucosa while bypassing hepatic metabolism [256]. Chemical constituents in blackberries have demonstrated preventive effects against multiple malignancies. A mucosal adhesion gel composed of Noveon AA1 and Carbopol 971 polymers has been developed and studied for site‐specific intraoral delivery of blackberry anthocyanins. Gel pH and storage temperature significantly impact blackberry anthocyanins chemical stability and mucosal permeability. The mucosal adhesion gel exhibits optimal stability for blackberry anthocyanins. Following incubation with human oral mucosal explants, anthocyanins rapidly diffuse into the human oral mucosa [257].
Co‐delivery Systems for Combination Therapy: Natural Products + Chemotherapy or Immunotherapy
8.3
Combination therapy strategies integrating natural products with conventional treatments represent a novel research focus. This approach enhances therapeutic efficacy while reducing side effects, producing synergistic effects [246].
In chemotherapy combination approaches, multiple drugs have been studied and developed. Quercetin competitively blocks various multidrug resistance efflux transporters, such as P‐gp, MRP‐1, and BCPR. Quercetin nanocrystals stabilized by glycyrrhizic acid (QT‐NCs/GL) successfully achieved liver‐targeted delivery, demonstrating significantly higher liver distribution compared with conventional nanocrystals (QT‐NCs/P188) in vivo. This provides an effective targeted delivery strategy for treating liver cancer using the combination of quercetin and glycyrrhizic acid [258]. A nanoscale system based on HA self‐assembly for targeted delivery of CUR (CUR@CS‐NP@HA) achieves active targeting of triple‐negative breast cancer cells expressing high levels of CD44. The nanoparticles exhibit pH‐responsive drug release properties, accelerating drug delivery within the acidic TME. Concurrently, CUR@CS‐NP@HA significantly enhances drug accumulation in tumor tissues, prolongs circulation time, improves bioavailability, and effectively inhibits tumor growth and metastasis [259]. The cabazitaxel liposomes modified with ginsenoside Rk1 (Rk1/CTX‐Lip) also serve as a typical example. This system utilizes Rk1 to replace cholesterol in constructing liposomes, which not only enhances drug targeting and stability but also achieves synergistic antitumor effects through the immunomodulatory action of Rk1 combined with cabazitaxel, while significantly reducing chemotherapy‐induced immunosuppression and infection risks [260].
In immunotherapy combinations, natural products modulate the TME to enhance immune responses. Nanoparticles codelivering anti‐PD‐1 antibodies and EGCG demonstrate improved T‐cell activation and tumor infiltration [261]. Micelles containing natural product immunomodulators (e.g., ginsenosides) with ICIs promote DC maturation and antigen presentation [262]. These approaches leverage the immunomodulatory properties of natural products to overcome resistance to conventional immunotherapies.
Clinical Translation and Precision Strategies
9
Natural products hold immense potential for clinical translation and precision applications in oncology [263]. We propose four approaches to achieve precise clinical translation of natural products: patient stratification based on molecular phenotypes and the immune microenvironment; promoting multimodal combinations of natural products with conventional therapies; personalized nutritional interventions guided by biomarkers; and concurrently advancing evidence accumulation and standardization in clinical translation. These strategies provide an implementation pathway for the scientific and rational application of natural products in precision oncology.
Patient Stratification for Natural Product‐Based Therapy
9.1
Although the biological activity of natural products has been well established, their therapeutic efficacy varies significantly across patient populations due to substantial differences in tumor biology, host metabolism, and genetic makeup. This variability underscores the necessity of stratification based on tumor‐specific signaling pathway abnormalities to identify individuals most likely to benefit from specific natural product interventions [264].
Natural product monomers can be selected based on specific signaling pathway abnormalities in a patient's tumor. For example, quercetin and CUR demonstrate significant therapeutic potential in tumors with abnormally activated Notch signaling pathways. Quercetin blocks downstream target gene expression by inhibiting proteolytic activation of Notch receptor proteins, while CUR effectively reverses chemotherapy resistance mediated by the Notch pathway [265, 266]. In tumors with overactivated PI3K/AKT/mTOR pathways, artemisinin and EGCG demonstrate multitarget regulatory advantages. Artemisinin simultaneously regulates the PI3K/AKT signaling axis and mTOR pathway, while EGCG synergistically induces programmed cell death through multiple nodes including p53 and Akt [267, 268]. A similar layered strategy applies to tumors with NF‐κB pathway overactivation, where ginsenoside Rg3 blocks signaling by inhibiting IκBα phosphorylation and p65 nuclear translocation [269].
The composition and functional state of the TIME can determine responses to natural product monomers. Certain natural compounds can reprogram the immune microenvironment, such as shifting TAMs from a protumor M2 phenotype to an antitumor M1 phenotype [219]. Baicalin has been demonstrated to promote TAM polarization toward M1, thereby alleviating the immunosuppressive environment [270]. For patients with immunologically “cold” tumors (characterized by insufficient T‐cell infiltration), natural product monomers like camptothecin and psoralen can induce immunogenic cell death (ICD), potentially transforming tumors into “hot” tumors and enhancing their sensitivity to immunotherapy. This enables stratification based on immune signatures such as the M1/M2 macrophage ratio or PD‐L1 expression [271].
Natural product monomers help overcome resistance to conventional therapies, providing a theoretical basis for stratification based on patient‐specific resistance mechanisms. For tumors exhibiting multidrug resistance due to overexpression of ABC transporters such as ABCG2, berberine inhibits these transporters and increases the accumulation of chemotherapeutic drugs within cancer cells [272]. In resistance scenarios linked to excessive inflammatory signaling, oxymatrine can reverse EMT‐associated resistance by inhibiting the NF‐κB pathway [273]. Furthermore, the natural compound acevaltrate can induce ferroptosis—a novel form of cell death—by dual‐targeting iron metabolism proteins (PCBP1/2) and the ferroptosis inhibitor GPX4, effectively eliminating drug‐resistant colorectal cancer cells [274].
In summary, these stratified strategies—ranging from molecular mechanisms to the immune microenvironment and specific resistance markers—enable the establishment of precision medicine models based on natural products. This approach allows the selection of natural products not only based on their general biochemical activity but also precisely matched to the individual tumor's biological characteristics, thereby unlocking immense potential for precision clinical oncology.
Integrating Natural Products Into Multimodal Clinical Regimens
9.2
The scientific integration of natural products into multimodal clinical treatment regimens represents a key strategy for enhancing cancer therapy outcomes. In current clinical practice, treatment strategies combining natural products with conventional therapies are widely adopted [246].
Regarding synergistic use with first‐line chemotherapeutic agents, multiple studies confirm that natural products can enhance the efficacy of chemotherapy drugs while reducing their toxic side effects. When CUR is combined with cisplatin, it not only increases tumor cell sensitivity to cisplatin by inhibiting the NF‐κB signaling pathway but also significantly mitigates cisplatin‐induced nephrotoxicity [275]. When combined with paclitaxel, baicalin reverses tumor cell multidrug resistance by regulating P‐gp function, thereby increasing paclitaxel accumulation in tumor tissues [276]. However, this combination strategy faces challenges such as unpredictable pharmacokinetic interactions and unclear optimal dosing sequences.
Combination therapy with ICIs shows promising prospects. Research indicates that resveratrol enhances PD‐1 inhibitor‐mediated antitumor immune responses by modulating gut microbiota composition, promoting DC maturation, and activating T cells [277]. Ginsenoside Rg3 improves the TIME by downregulating PD‐L1 expression in tumor cells, thereby increasing immunotherapy response rates [278]. These mechanisms provide a robust theoretical foundation for synergistic applications of natural products with immunotherapy.
Synergistic effects with metabolic inhibitors and antiangiogenic agents have also been extensively studied. Combining quercetin with bevacizumab not only enhances antiangiogenic effects but also delays resistance development by inhibiting the HIF‐1α/VEGF signaling pathway [279]. The combination of metformin and apigenin synergistically modulates the HGF/MET signaling pathway, inhibits MET‐mediated invasion, and influences downstream pathways such as PI3K/AKT, offering an innovative approach to enhance the efficacy and outcomes of cancer treatment [280].
Regarding application timing, natural products demonstrate unique value across different treatment phases. Preoperative administration of Ganoderma lucidum polysaccharides improves immune function, creating more favorable conditions for surgery [281]. Postoperative use of astragaloside IV promotes wound healing and reduces surgical stress responses [282]. During intervals between radiotherapy and chemotherapy, silymarin accelerates normal tissue repair and reduces cumulative toxicity [283]. These sequential application strategies provide comprehensive supportive care throughout the treatment journey for cancer patients.
However, integrating natural products into multimodal treatment regimens still faces challenges such as determining standardized dosages, assessing drug interactions, and optimizing treatment sequencing. Further clinical research is needed to establish appropriate combination therapies and fully leverage the potential of natural products in comprehensive cancer treatment.
Biomarker‐Driven Precision Nutrition and Supplementation
9.3
Natural products are playing an increasingly vital role in precision cancer therapy. Their interactions with biomarkers not only elucidate drug mechanisms of action but also drive paradigm shifts in biomarker discovery. Traditionally, biomarker identification relied primarily on comparative analysis between tumor and normal tissues [284]. However, natural small molecules, acting as “chemical probes,” offer a functionally guided approach to biomarker discovery.
By modulating pathways associated with tumor hallmarks, natural products directly alter the expression levels of specific molecular markers. This approach not only validates drug targets but also uncovers potential biomarkers. For instance, EGCG effectively inhibits tumor invasion and metastasis by suppressing MMP‐9 and upregulating E‐cadherin [285]. Resveratrol and quercetin reverse the Warburg effect by downregulating glycolytic key molecules such as lactate dehydrogenase (LDHA) and GLUT1. These changes not only demonstrate the metabolic regulatory function of natural products but also establish LDHA and GLUT1 as functional biomarkers for assessing tumor metabolic status [286]. Similarly, baicalin suppresses the STAT3 signaling pathway to reduce PD‐L1 expression, thereby modulating the TIME while suggesting that p‐STAT3 and PD‐L1 can serve as monitoring indicators for immunomodulatory therapy efficacy [287].
More importantly, natural products play a unique “reverse‐engineering” role in biomarker discovery. Unlike traditional “pathological differential” approaches, this mechanism‐driven method starts with natural small‐molecule interventions and identifies key functional molecules by analyzing their induced systemic responses. In colorectal cancer research, the natural compound acevaltrate was revealed to potently induce ferroptosis in cancer cells by dual‐targeting PCBP1/2 and GPX4 proteins. Based on this mechanism, the expression levels of PCBP1/2 and GPX4 can serve as potential biomarkers for predicting acevaltrate treatment sensitivity [274]. Furthermore, studies indicate that Tripterygium glycosides and camptothecin can significantly regulate the expression profiles of characteristic miRNAs in osteosarcoma cells. These miRNA populations, specifically modulated by natural products, are emerging as novel molecular markers for evaluating the anticancer effects of dual‐purpose medicinal and edible compounds. This approach reveals functional sensitive sites undetectable by conventional pathological differential analysis, thereby identifying therapeutic response biomarkers with higher clinical guidance value [288].
In summary, the interaction between natural products and biomarkers is propelling the advancement of precision cancer therapy. Clinically, by monitoring the dynamic changes in biomarkers following natural product interventions, physicians can not only objectively evaluate the comprehensive regulatory effects of drugs on tumor metabolism, apoptosis, and immunity but also identify patient populations sensitive to specific natural products based on “reverse‐guided” biomarker discovery, thereby achieving true personalized treatment. This mutually reinforcing and inspiring model between natural products and biomarkers provides more precise scientific rationale for the clinical translation of natural products [289].
Clinical Trials, Real‐World Evidence, and Translational Challenges
9.4
These findings highlight the emerging role of natural products—not only as therapeutic agents but also as multidimensional regulatory tools in biomarker‐guided precision nutrition and oncology. By modulating tumor‐specific characteristics, natural product‐based interventions enable personalized customization, enhancing therapeutic efficacy while minimizing adverse reactions and drug resistance [290].
At the epigenetic regulation level, natural products exhibit remarkable potential. Genistein from soy isoflavones reverses the silencing of tumor suppressor genes by inhibiting DNA methyltransferase activity; resveratrol modulates the expression of multiple tumor‐associated genes by activating the histone deacetylase SIRT1; EGCG in tea polyphenols influences the malignant phenotype of tumor cells by regulating the microRNA expression profile [76, 288, 291].
From multiple perspectives, natural products demonstrate significant advantages at cellular, microenvironmental, and clinical levels. At the cellular level, terpenoids like Tripterygium lactones effectively inhibit tumor proliferation by inducing DNA damage and cell cycle arrest; alkaloids such as vincristine block tumor cell mitosis by interfering with tubulin polymerization; while polysaccharides like Ganoderma lucidum polysaccharides enhance the body's antitumor immune response by activating immune‐related signaling pathways [36, 281].
At the TME level, saponins like ginsenoside Rg3 reduce tumor invasion and metastasis by inhibiting MMP‐2/9 activity; polyphenols improve the immunosuppressive microenvironment by modulating TAM polarization; wedelolactone is a naturally occurring coumarin that enhances IFN‐γ signaling by inhibiting the phosphatase activity of T‐cell protein tyrosine phosphatase, thereby promoting apoptosis in tumor cells via a STAT1‐dependent mechanism [227, 292, 293].
Clinically, natural products’ multitargeted properties, low toxicity, and ability to modulate multiple biomarkers (e.g., PD‐L1, CTLA‐4, VEGF) make them ideal candidates for personalized cancer therapy. For instance, alkaloids like homoharringtonine demonstrate unique advantages in leukemia treatment by inducing cell differentiation; flavonoids like baicalin demonstrate significant efficacy in treating precancerous lesions by suppressing inflammatory factor expression [108, 294].
Natural products have emerged as a key focus in innovative drug development for cancer treatment. Among those successfully applied clinically, paclitaxel is widely used for breast and ovarian cancer; camptothecin and its derivatives, such as irinotecan, target colorectal cancer and leukemia; while vincristine and vinblastine serve as core agents in treatment regimens for leukemia, lymphoma, and various solid tumors [21, 36, 37]. In recent years, chlorogenic acid for injection has demonstrated promising results in clinical studies for treating recurrent Grade IV glioblastoma. Early research data indicate that it significantly prolongs median survival in patients [295]. Among promising drug candidates, the small molecule 3,4‐diisobutyryl derivative of auxarthrol A—isolated from endophytic fungi of Euphorbiaceae plants and chemically modified—directly targets the LIC1 protein. It inhibits NSCLC tumor growth by inducing autophagic cell death and enhancing tumor sensitivity to anti‐PD‐1 immunotherapy [296]. Furthermore, multiple studies indicate that various natural plant bioactive compounds can reverse tumor multidrug resistance by downregulating P‐gp expression on tumor cell membranes, offering a potential strategy to overcome clinical chemotherapy resistance [9, 258, 276].
Meanwhile, real‐world studies further support the value of natural products in cancer prevention and long‐term maintenance therapy. For instance, a prospective cohort study involving 820 Greek women demonstrated a significant inverse association between flavonoid intake and breast cancer risk: a 13% reduction in risk for every 0.5 mg increase in daily intake. Specific dietary flavonoids may exert preventive effects against breast cancer, providing crucial epidemiological evidence for the cancer‐preventive potential of natural products [297]. Another study involving patients with postoperative tumor‐free metastatic colorectal cancer demonstrated that adding Qu‐yu‐jie‐du decoction (QYJD) to standard care for 2 years significantly improved disease‐free survival and showed a trend toward overall survival benefit. The study also found QYJD treatment reduced systemic inflammatory markers and lactate dehydrogenase levels with good safety [298]. This provides clinical‐grade evidence for natural products as low‐toxicity, effective maintenance strategies in preventing tumor recurrence and metastasis. Natural products demonstrate significant adjunctive value in postcancer surgery and long‐term care. Danggui Buxue Decoction significantly reduces the incidence and severity of chemotherapy‐induced thrombocytopenia in patients with malignant solid tumors, minimizing treatment interruptions [299]. The Japanese Kampo medicine Yokukansan prevents delirium following major cancer surgery in elderly patients by regulating plasma phospholipid metabolism, with its mechanism of alleviating cerebral oxidative stress validated in animal models [300].
Despite these significant achievements, the clinical translation of natural products still faces numerous critical challenges that extend far beyond target identification and drug properties to include key aspects such as quality control and standardization. Take the study on HBK compound herbal preparations as an example: This research employed UHPLC–ESI–Q‐TOF–MS/MS technology to establish a chemical fingerprint profile of the compound formulation, precisely identifying seven phytochemical markers including rutin and astragaloside IV, along with three nutritional components such as β‐glucan. Simultaneously adhering to Chinese Pharmacopoeia standards, it rigorously monitored safety indicators including heavy metals, pesticide residues, and microbial limits, fully demonstrating the practical application of natural product standardization. These systematic quality control practices provide critical technical support for the clinical translation of natural products [242].
With continuous advancements in chemical biology, nanotechnology, and synthetic biology, multiple challenges in the clinical translation of natural products are gradually being overcome [301, 302]. Future research should focus on establishing more comprehensive quality control standards, strengthening the evidence base of evidence‐based medicine, and promoting the deep integration of natural products with precision medicine. This will ultimately provide cancer patients with safer and more effective personalized treatment options (Figure 6).
Benefits and limitations of natural products in tumor prevention and treatment at cellular, tissue, and clinical levels.
Conclusion
10
Natural products demonstrate multidimensional value in cancer prevention and treatment, with their core advantage lying in influencing the entire process of tumorigenesis and progression through multitarget regulatory mechanisms. These compounds not only directly intervene in cancer cell metabolic reprogramming, induce apoptosis, and inhibit metastasis, but also reshape the TME and modulate immune cell function to achieve synergistic tumor suppression. From prevention to treatment, from adjuvant radiotherapy and chemotherapy to improving quality of life, natural products offer unique complementary strategies for tumor therapy.
However, the journey from laboratory to clinical application presents significant challenges. Issues such as low bioavailability, complex quality control, and substantial interindividual variability in efficacy remain major obstacles. Fully realizing their potential necessitates multifaceted strategies. These include leveraging nanotechnology and novel delivery systems to enhance bioavailability, employing biomarker‐driven approaches for patient stratification, and integrating rigorous clinical trials with real‐world evidence. The future of natural products in oncology lies in their integration into multimodal and precision treatment regimens, transforming them into evidence‐based, effective components of precision cancer medicine. This ultimately offers safer, more personalized therapeutic options for patients worldwide.
In summary, natural products not only exert multifaceted preventive and therapeutic effects against cancer but also demonstrate highly promising adjunctive benefits when combined with metabolic targeting and immune response strategies. Moving forward, interdisciplinary collaboration among food science, oncology, and systems biology will be pivotal in transforming these natural compounds into effective tools for precision cancer therapy.
Author Contributions
Ruimiao Qian, Hui Fu, and Jun Ge framed the synthesis. Ni Fan and Zheng Sun collected literature and sifted through articles. Ruimiao Qian, Chengcheng Zhao, and Yunfei Li wrote the manuscript and drew diagrams. Hui Fu, Yujiao Sun, and Yingpeng Li proposed the project and revised the manuscript. All authors have contributed to the article and approved the submitted version.
Ethics Statement
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Y. Zhu , Z. J. Ouyang , H. J. Du , et al., “New Opportunities and Challenges of Natural Products Research: When Target Identification Meets Single‐cell Multiomics,” Acta Pharmaceutica Sinica B 12, no. 11 (2022): 4011–4039.36386472 10.1016/j.apsb.2022.08.022PMC 9643300 · doi ↗ · pubmed ↗
- 2S. H. Lee , J. K. Ryu , K. Y. Lee , et al., “Enhanced Anti‐tumor Effect of Combination Therapy With Gemcitabine and Apigenin in Pancreatic Cancer,” Cancer Letters 259, no. 1 (2008): 39–49.17967505 10.1016/j.canlet.2007.09.015 · doi ↗ · pubmed ↗
- 3E. A. Mohamad , M. R. El‐Garhy , and M. M. Rageh , “Improving Cisplatin Chemotherapy in Vivo by Niosomal Propolis and Chrysin as Nanoadjuvants,” Medical Oncology 42, no. 12 (2025): 539.41193918 10.1007/s 12032-025-03105-5 · doi ↗ · pubmed ↗
- 4H. Shulan , N. Peng , L. Likun , et al., “Effectiveness of Yiqi Chupi Powder for Alleviating Cancer‐related Fatigue in Patients Following Colorectal Cancer Surgery: A Randomized Controlled Trial,” Journal of Traditional Chinese Medicine 45, no. 5 (2025): 1119–1126.41015810 10.19852/j.cnki.jtcm.2025.05.017PMC 12454259 · doi ↗ · pubmed ↗
- 5F. Yang , Z. Li , H. Zhang , et al., “Natural Product Erianin: Mitigating FOLFOX Toxicity and Enhancing Against Colorectal Cancer,” Front Chem 13 (2025): 1650197.40919243 10.3389/fchem.2025.1650197 PMC 12411860 · doi ↗ · pubmed ↗
- 6J. Cai , Z. R. Xuan , Y. P. Wei , H. B. Yang , and H. Wang , “[Effects of perioperative administration of Rhubarb on acute inflammatory response in patients With gastric cancer],” Zhong Xi Yi Jie He Xue Bao = Journal of Chinese Integrative Medicine 3, no. 3 (2005): 195–198.15885167 10.3736/jcim 20050309 · doi ↗ · pubmed ↗
- 7Y. Zheng , N. Feng , C. L. Li , and Z. Q. Li , “Natural Products Target Programmed Cell Death Signaling Mechanisms to Treat Colorectal Cancer,” Frontiers In Pharmacology 16 (2025): 26.10.3389/fphar.2025.1565332 PMC 1205879140342991 · doi ↗ · pubmed ↗
- 8C. Yu , C. Jia , G. Chen , et al., “Yiai Fuzheng Decoction Inhibits Triple‐negative Breast Cancer by Remodeling the Immune Microenvironment,” Frontiers In Immunology 16 (2025): 1615631.41098716 10.3389/fimmu.2025.1615631 PMC 12518410 · doi ↗ · pubmed ↗