Cultured cells activate IRE1 during attachment and flattening after routine passaging
Paige Dillon, Lincoln Hollingshead, Julie Hollien

TL;DR
This study shows that cultured cells activate an ER stress response during routine passaging, even without added stressors.
Contribution
The study reveals that Xbp1 splicing occurs during cell passaging, indicating an unfolded protein response during reattachment.
Findings
Xbp1 mRNA is spliced in MC3T3-E1 cells after routine passaging without canonical ER stressors.
Xbp1 splicing is reduced when cells are plated on non-adherent culture dishes.
The splicing is independent of the dissociation buffer used.
Abstract
During endoplasmic reticulum (ER) stress, the ER membrane protein IRE1 initiates the regulated splicing of Xbp1 mRNA, leading to the production of a potent transcription factor that helps cells restore proteostasis. We report that Xbp1 is also spliced following the routine passaging of mouse MC3T3-E1 cells, without the addition of canonical ER stressors. This splicing was independent of the type of dissociation buffer used to release cells from the surface, but was reduced when cells were plated on non-adherent culture dishes. These findings suggest that certain cultured mammalian cells induce an unfolded protein response during reattachment and spreading after passaging.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
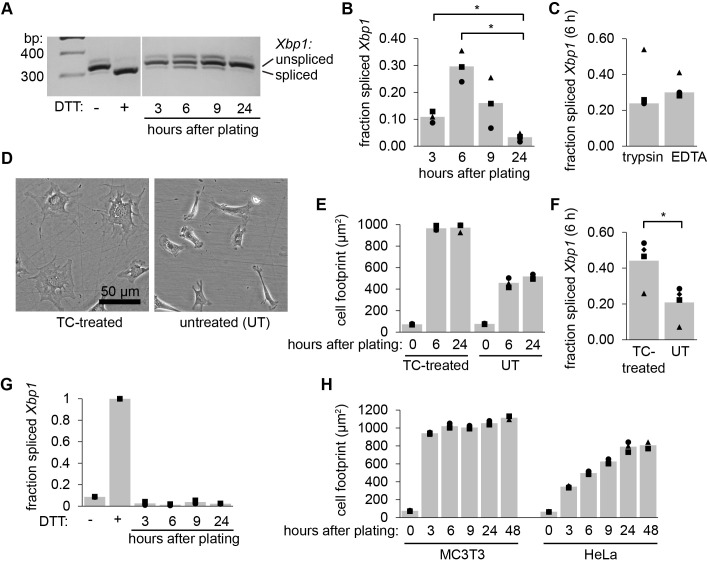 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEndoplasmic Reticulum Stress and Disease · Cellular transport and secretion · Fungal and yeast genetics research
Description
IRE1 is an endoplasmic reticulum (ER) transmembrane resident protein and key mediator of the unfolded protein response (Walter and Ron, 2011; Wiseman et al., 2022). When activated by protein misfolding or lipid stress in the ER, the cytosolic-facing nuclease domain of IRE1 cleaves the mRNA encoding the transcription factor XBP1, initiating a noncanonical splicing event that leads to the downstream upregulation of many genes involved in protein folding, processing, and degradation (Yoshida et al., 2001; Calfon et al., 2002). We noticed an increase in Xbp1 splicing following routine passaging of MC3T3-E1 cells, which are cultured fibroblasts derived from mouse calvaria, often used for microscopy due to their broad and flat morphology. This observation led us to investigate how passaging affects IRE1 activation in these cells.
We released MC3T3-E1 cells from their culture flasks using trypsin-EDTA, diluted in fresh media, replated cells in new flasks, and collected RNA after 3, 6, 9, and 24 hours. As a control, we treated cells with dithiotreitol (DTT, 2 mM, 6 h), a strong reducing agent and inducer of protein misfolding in the ER. We then purified total RNA, synthesized cDNA, used PCR to amplify Xbp1 with primers surrounding the 26 nucleotide intron, and analyzed the products by agarose gel electrophoresis. We observed substantial Xbp1 splicing, which peaked at 6 hours post-passaging and returned to baseline levels by 24 hours (A, B). This splicing did not appear to be a consequence of the proteolysis of plasma membrane proteins by trypsin, as we observed similar levels of splicing when we used a non-enzymatic, EDTA-only dissociation reagent (C).
Because Xbp1 splicing was delayed following passaging and was not dependent on proteolytic dissociation methods, we wondered whether IRE1 activation was caused by reattachment of cells to the flasks. To test this, we compared cells plated onto standard TC-treated plates vs. untreated plates that do not promote attachment. Both Xbp1 splicing (D) and the area of the cell footprint (E, F) were diminished by about 50% on the untreated plates. We conclude that MC3T3-E1 cells activate IRE1 while reattaching and spreading out following passaging.
To test whether other tissue culture cells also splice Xbp1 after passaging, we carried out similar experiments in HeLa cells. HeLa cells did not splice Xbp1 after passaging (G) and did not spread out as rapidly or to the same extent as MC3T3-E1 cells (H), again suggesting that the rapid attachment and spreading of MC3T3-E1 cells is the key event that leads to Xbp1 splicing following passaging.
We speculate that Xbp1 splicing may be caused by the increase in surface area and membrane tension as cells transition from a sphere-like morphology during dissociation to a flattened morphology when attaching to the substrate. The unfolded protein response can be activated by a variety of types of mechanical stress (Townson and Progida, 2025; Chen et al., 2025), and IRE1 is activated by saturated fatty acids and lipid bilayer stress resulting in increased membrane packing and thickness (Halbleib et al., 2017; Promlek et al., 2011; Thibault et al., 2012; Volmer et al., 2013). It remains unclear how changes in cell shape and attachment would propagate to the ER, but this could happen through ER-plasma membrane contact sites, through alterations in the cytoskeleton, or through disruptions to Ca ^2+^ homeostasis mediated by mechanically-gated ion channels. In any case, the unfolded protein response coordinates many aspects of lipid synthesis and metabolism (Moncan et al., 2021), which may help to re-establish homeostasis at the plasma membrane following the disruptions inherent to passaging cells. Our observations may therefore inform studies on the role of IRE1 and the ER in responding to mechanical stress, and may be generally useful to researchers studying ER stress in cultured cells.
Methods
Cell culture
We cultured MC3T3-E1 cells in MEMα with nucleosides, L-glutamine, and no ascorbic acid (Life Technologies) supplemented with 10% FBS at 37 C and 5% CO 2 . We cultured HeLa cells in DMEM supplemented with 10% FBS at 37 C and 5% CO 2 . Both cell lines were purchased from American Type Culture Collection (ATCC).
Xbp1 splicing assay
We washed subconfluent MC3T3-E1 and HeLa cells with D-PBS (GenClone), treated with 0.05% trypsin-EDTA (Gibco) for 4 minutes, then diluted to a final cell count of 2-3×10 ^5^ in MEMα and plated in 25 cm ^2^ flasks. For nonenzymatic dissociation, we treated cells with Versene (Gibco) for 8 minutes before diluting in MEMα. For experiments comparing *Xbp1 * splicing on treated and untreated plates, we used 10 cm, non-treated, sterilized VWR Tissue Culture Dishes (Avantar) along with treated 10 cm tissue culture dishes (Gen Clone). To collect RNA, we gently washed with PBS, then added 500 μL of RNA lysis buffer directly to the flasks or plates and purified total RNA using Quick RNA Miniprep kits (Zymo Research). Most cells remained attached to the surface during washing, and RNA yields were unchanged between 3 and 6 h after trypsinization, increasing by ~26% after 9 h and ~70% after 24 h, as expected based on their ~24-hour doubling time. Average RNA yield from untreated plates was ~82% of that from tissue-culture treated plates, suggesting that some cells were lost due to lack of attachment but not enough to explain the 2-fold difference in Xbp1 splicing. We synthesized cDNA using Moloney murine leukemia virus reverse transcriptase (New England Biolabs) and T18 as a primer. We amplified cDNA with primers surrounding the regulated Xbp1 splice site (mouse: AGAAGAGAACCACAAACTCCAG and GGGTCCAACTTGTCCAGAATGC or human: AGCTCAGACTGCCAGAGATCG and AATCCATGGGGAGATGTTCTG) and ran the PCR products on 2% agarose gels. We quantified the intensities of the spliced and unspliced Xbp1 bands using ImageJ.
Cell footprint calculations
We imaged cells using an Amscope 4k HD408L camera and a cell-culture inverted microscope (Olympus CKX53, 10x, NA = 0.25). We collected measurements of the cell footprint, defined as the surface area occupied by a cell, by tracing 50 cells per condition. All experiments were repeated in triplicate and quantified using ImageJ.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Calfon M Zeng H Urano F Till JH Hubbard SR Harding HP Clark SG Ron D 200213 IRE 1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 m RNA.Nature 41568670028-0836929610.1038/415092 a 11780124 · doi ↗ · pubmed ↗
- 2Chen Z Chen P Li J Landao-Bassonga E Papadimitriou J Gao J Liu D Tai A Ma J Lloyd D Kennedy BF Zheng MH 2025625 External strain on the plasma membrane is relayed to the endoplasmic reticulum by membrane contact sites and alters cellular energetics.Sci Adv 1126 eads 6132 eads 613210.1126/sciadv.ads 613240561024 PMC 12190009 · doi ↗ · pubmed ↗
- 3Halbleib K Pesek K Covino R Hofbauer HF Wunnicke D Hänelt I Hummer G Ernst R 201776 Activation of the Unfolded Protein Response by Lipid Bilayer Stress.Mol Cell 6741097-2765673684.e 810.1016/j.molcel.2017.06.01228689662 · doi ↗ · pubmed ↗
- 4Moncan M Mnich K Blomme A Almanza A Samali A Gorman AM 202114 Regulation of lipid metabolism by the unfolded protein response.J Cell Mol Med 2531582-18381359137010.1111/jcmm.1625533398919 PMC 7875919 · doi ↗ · pubmed ↗
- 5Promlek Thanyarat Ishiwata-Kimata Yuki Shido Masahiro Sakuramoto Mitsuru Kohno Kenji Kimata Yukio 2011915 Membrane aberrancy and unfolded proteins activate the endoplasmic reticulum stress sensor Ire 1 in different ways Molecular Biology of the Cell 22181059-15243520353210.1091/mbc.e 11-04-029521775630 PMC 3172275 · doi ↗ · pubmed ↗
- 6Thibault Guillaume Shui Guanghou Kim Woong Mc Alister Graeme C. Ismail Nurzian Gygi Steven P. Wenk Markus R. Ng Davis T.W. 2012101 The Membrane Stress Response Buffers Lethal Effects of Lipid Disequilibrium by Reprogramming the Protein Homeostasis Network Molecular Cell 4811097-2765162710.1016/j.molcel.2012.08.01623000174 PMC 3496426 · doi ↗ · pubmed ↗
- 7Townson Jonathan Progida Cinzia 2025215 The emerging roles of the endoplasmic reticulum in mechanosensing and mechanotransduction Journal of Cell Science 13840021-953310.1242/jcs.26350339976266 · doi ↗ · pubmed ↗
- 8Volmer Romain van der Ploeg Kattria Ron David 201334 Membrane lipid saturation activates endoplasmic reticulum unfolded protein response transducers through their transmembrane domains Proceedings of the National Academy of Sciences 110120027-84244628463310.1073/pnas.1217611110 PMC 360697523487760 · doi ↗ · pubmed ↗