Chemical inhibition of SUMOylation activates the FSHD locus
Alice Nordlinger, Loéva Morin, Alexandra Andrieux, Jean Philippe Trani, Pierre Perrin, Nathalie Eudes, Anne Bigot, Anne Dejean, Frédérique Magdinier

TL;DR
A drug that blocks SUMOylation can activate a harmful gene in a muscle disease, offering a new treatment approach.
Contribution
SUMOylation inhibition is shown to activate DUX4 in FSHD without affecting DNA methylation or SMCHD1 activity.
Findings
TAK-981 promotes transcriptional reprogramming of the 4q35 locus.
SUMOylation inhibition activates DUX4 expression independently of DNA methylation.
This identifies SUMOylation as a novel regulatory mechanism for DUX4.
Abstract
Facioscapulohumeral muscular dystrophy (FSHD) is a progressive and debilitating muscle disease for which no cure currently exists. In the majority of cases, FSHD is associated with the contraction of the D4Z4 macrosatellite repeat array at the 4q35 locus, leading to the inappropriate activation of DUX4, normally expressed during early embryogenesis. In FSHD, the genetic contraction is accompanied by hypomethylation of the D4Z4 array. Although a connection between DNA hypomethylation and DUX4 expression has been suggested, the precise mechanisms that regulate DUX4 transcription remain incompletely defined. The post-translational modification by SUMO was shown previously to repress the expression of Dux, the DUX4 homolog, in mouse embryonic stem cells. Based on these findings, we explored here the contribution of SUMOylation in the regulation of DUX4 in human muscle cells. We demonstrate…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
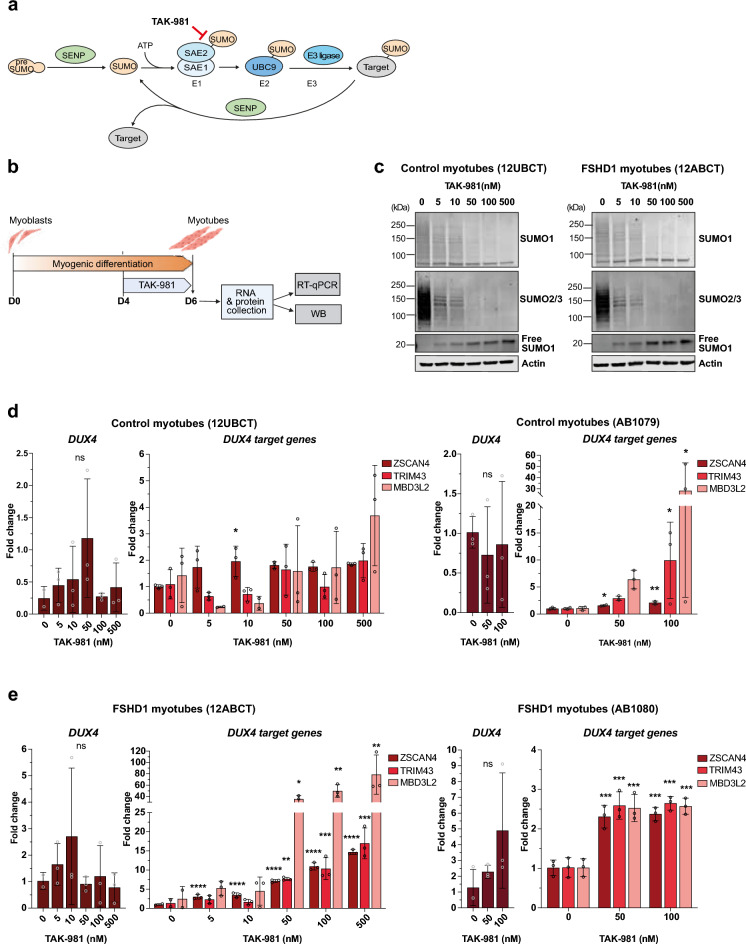 Figure 1
Figure 1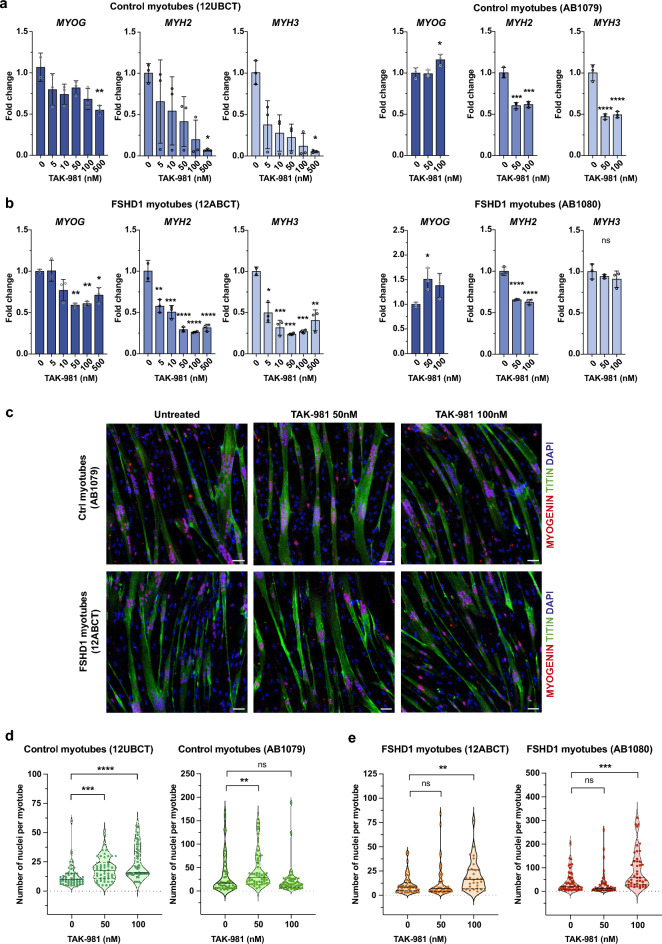 Figure 2
Figure 2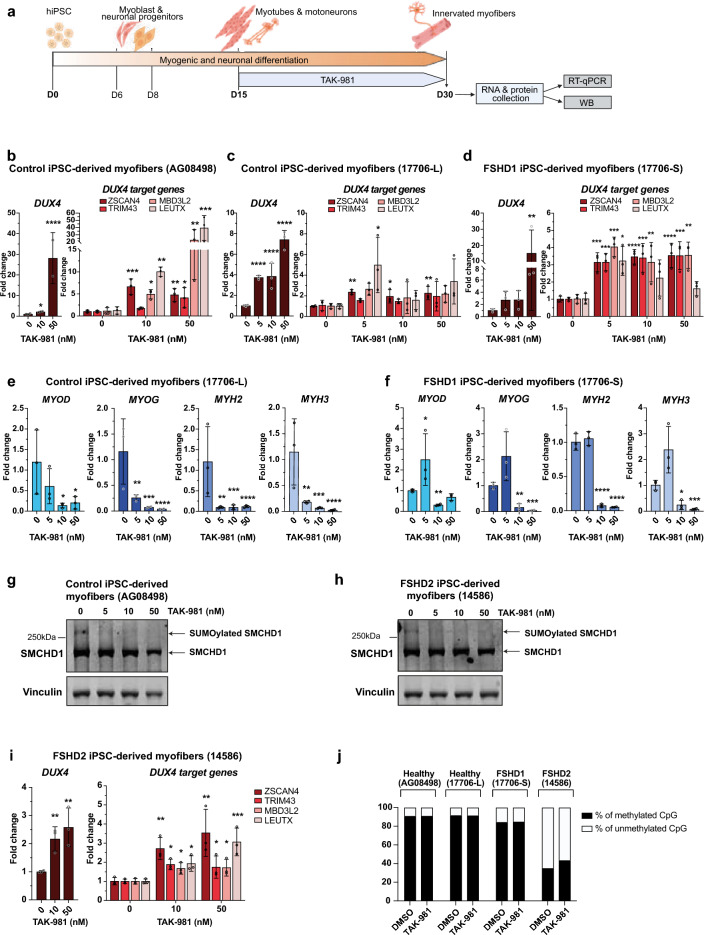 Figure 3
Figure 3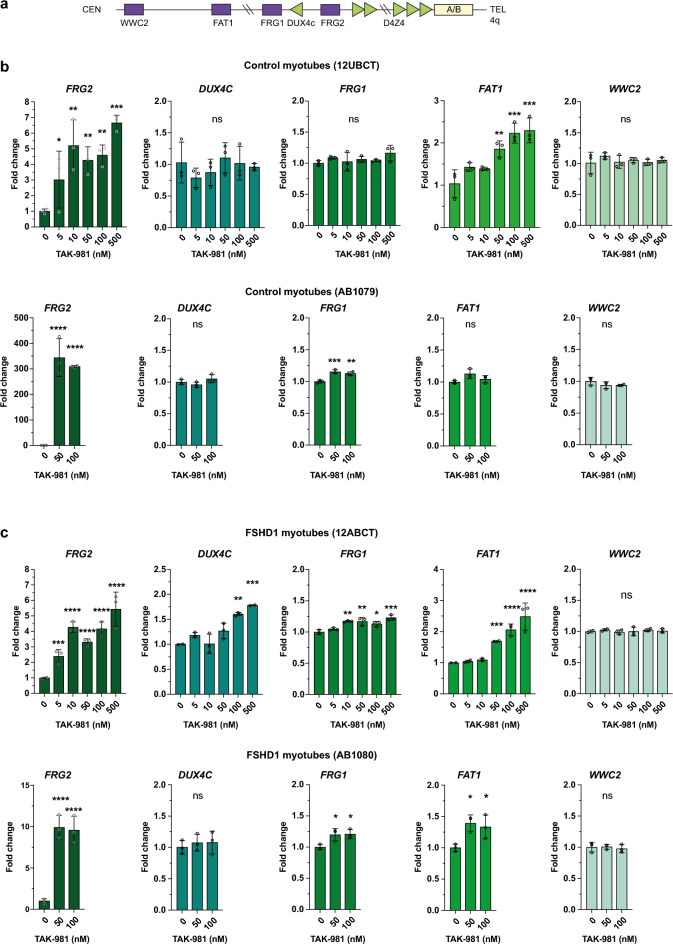 Figure 4
Figure 4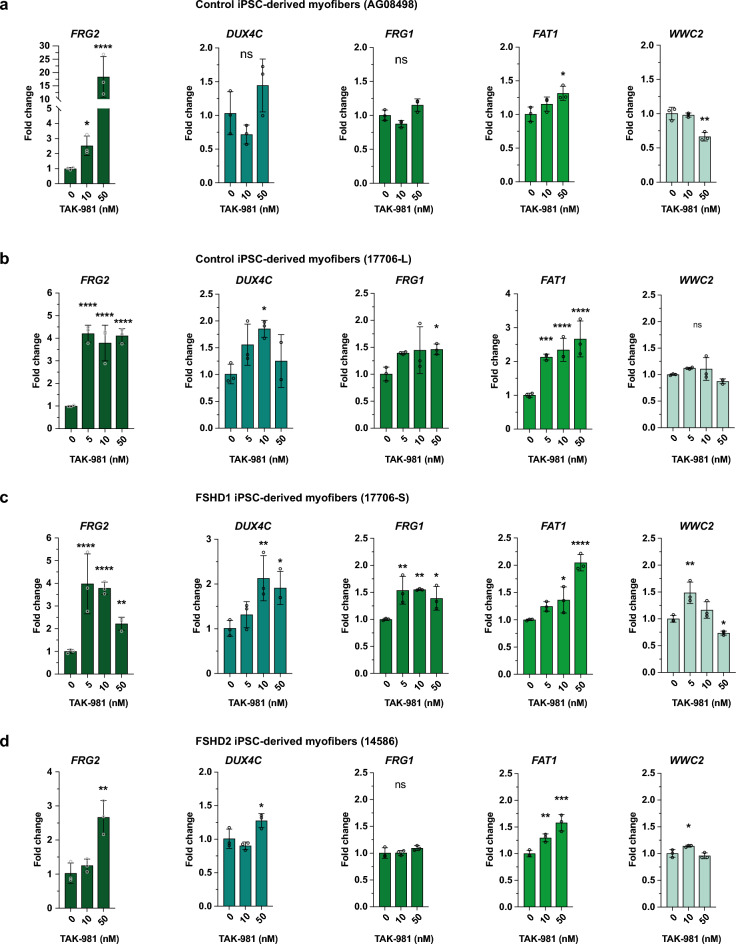 Figure 5
Figure 5- —https://doi.org/10.13039/501100001665Agence Nationale de la Recherche
- —https://doi.org/10.13039/100016669Sjöbergstiftelsen
- —https://doi.org/10.13039/501100000781European Research Council
- —https://doi.org/10.13039/501100002915Fondation pour la Recherche Médicale
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMuscle Physiology and Disorders · Ubiquitin and proteasome pathways · Sirtuins and Resveratrol in Medicine
Introduction
SUMOylation is a reversible post-translational modification that regulates protein functions through covalent attachment of small ubiquitin-like modifier (SUMO) proteins to their protein substrates in eukaryotic cells ^1,2^. SUMO proteins exist as three main paralogues in mammals, SUMO1, SUMO2 and SUMO3, with SUMO2 and 3 sharing high similarities. SUMOylation is a dynamic energy-dependent process involving unique E1 and E2 enzymes and a series of E3 ligases. The reversible aspect of SUMOylation is ensured by action of specific SUMO proteases called SENPs (Fig. 1a). The SUMO pathway, which predominantly targets transcription factors and chromatin-associated proteins ^3–6^, is a key regulator of nuclear processes and cell identity. In particular, SUMOylation was reported to regulate the expression of Dux in mESCs, with loss of SUMOylation leading to derepression of Dux and transition to a totipotent-like state ^7–10^. In mESCs, SUMOylation-dependent silencing of Dux was shown to involve Prc1.6 and Kap/Setdb1 recruitment onto the Dux locus^7^, as well as proper Dppa2/Dppa4^8,9^ and histone H1 SUMOylation^10^.Fig. 1. Chemical inhibition of SUMOylation induces a coordinated DUX4 program in FSHD1 patient-derived myotubes. (a) Representation of the reversible SUMOylation process. Before the first conjugation, a nascent SUMO precursor is proteolytically processed by a SUMO protease (SENP). Mature SUMO is then conjugated to a SAE1-SAE2 complex (E1) in an ATP-dependent manner. Next, activated SUMO is transferred to the E2 conjugating enzyme UBC9, before being conjugated to a target protein by the E3 ligase. SUMO conjugates can be cleaved from the target protein by a SENP, ensuring a dynamic and reversible SUMOylation process. TAK-981 inhibits SUMOylation by blocking activated SUMO at the catalytic site of SAE2. (b) Schematic representation of the experimental design for human hTERT-immortalized myoblasts^29^. At confluence, control or FSHD1 immortalized myoblasts were differentiated for 6 days in the presence of 2% horse serum and treated with DMSO or TAK-981 at different concentrations from day 4 to 6 of differentiation. At day 6, RNA and proteins were collected for gene expression analysis by RT-qPCR and western blot experiments. (c) Immunoblots for SUMO1 and SUMO2/3 in myotubes from control (12UBCT) and FSHD1 (12ABCT) cells. Actin was used as a loading control. (d) DUX4-fl and DUX4 target gene expression analyses in two cell lines of control myotubes (12UBCT and AB1079). Fold-changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001. (e) DUX4-fl and DUX4 target gene expression analyses in myotubes obtained from two FSHD1 patients (12ABCT and AB1080). Fold-changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3 independent experiments. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001.
The mouse Dux and human DUX4 transcription factors are involved in the zygotic genome activation (ZGA) during the 2C-4C stage in mouse and 4C-8C stage in human embryos, respectively^11,12^. In human cells, DUX4 is encoded by the D4Z4 macrosatellite element that is located in the subtelomeric region of the 4q arm (4q35 locus)^13^. Each D4Z4 element is approximately 3.3 kilobases in size and forms large tandemly repeated array structures at the subtelomeric 4q35 and 10q26 loci^14,15^. Smaller size or partial D4Z4-like sequences are scattered in heterochromatin regions throughout the genome, in particular at the p arm of acrocentric chromosomes. At 4q35 and 10q26 loci, the number of repeat units can vary between individuals, typically ranging from 11 to over 100^16^. These arrays are highly GC-rich and present with heterochromatin features such as DNA methylation, leading to epigenetic repression ^17–19^.
The 4q35 locus is linked to type 1 Facioscapulohumeral Dystrophy (FSHD1, OMIM #158900), an autosomal dominant neuromuscular disease that is characterized by a progressive weakness and wasting of specific skeletal muscles of the face, shoulder girdle, and upper arms^20,21^. In individuals with FSHD1, the D4Z4 repeat array on chromosome 4q35 is contracted to contain 1–10 repeat units, compared to the normal size range of 11–100 repeats in the unaffected population. This contraction is associated with D4Z4 DNA hypomethylation and chromatin opening enabling aberrant expression of the DUX4 gene from the most distal D4Z4 repeat. Importantly, disease manifestation requires the presence of a permissive 4qA haplotype distal to the last D4Z4 unit, which provides a functional polyadenylation signal necessary for stable DUX4 transcript production^22^. FSHD2 (5% of patients, OMIM #158901) is clinically similar to FSHD1 but arises from a digenic mechanism in the absence of D4Z4 array contraction. In approximately 80% of these FSHD2 patients, the disease is associated with pathogenic variants of SMCHD1 and involves D4Z4 hypomethylation^23,24^, allowing DUX4 expression from a normally sized repeat array^25^. In human somatic cells, experimental overexpression of DUX4 activates the expression of more than 400 genes. This cascade of transcriptional activation has been associated with immune response, muscle inflammation and atrophy or inhibition of muscle regeneration^26^.
Given the role of SUMOylation in the regulation of Dux in mESCs, we decided to evaluate whether the SUMO pathway may similarly regulate the expression of DUX4 in human muscle cells and, in particular, in the context of FSHD. To this aim, we exploited two different cellular models in which we decreased the global cellular levels of SUMOylation using a selective chemical inhibitor to investigate the role of SUMOylation in the regulation of DUX4 and the 4q35 region, as well as the possible involvement of SMCHD1 post-translational modifications^27^. We show that TAK‑981 (subasumstat), a first-in-class SUMOylation inhibitor^28^, promotes transcriptional reprogramming at the 4q35 locus and induces DUX4 expression, independently of changes in DNA methylation or SMCHD1 ATPase activity.
Materials and methods
Ethics declaration and cellular models
CDK4/hTERT immortalized myoblasts (12UBCT, 12ABCT) were described in^29^ and obtained from the BBRI/Wellstone Institute. The 12ABCT cells were obtained from a FSHD1 patient carrying 5 D4Z4 units. The hTERT/CDK4-immortalized AB1080 and AB1079 lines were provided by the MyoLine platform of the Institute of Myology (Paris, France). Prior to hTERT/CDK4 immortalization, primary myoblasts were isolated from muscle samples anonymously provided by MyoBank, a tissue bank affiliated with EuroBioBank and authorized by the Ministry of Higher Education, Research and Innovation (AC-2019-3502). The AB1080 cell line was derived from paravertebral muscles of a FSHD1 patient carrying 6.3 D4Z4 units. The AB1079 cell line was obtained from the quadriceps of a 38 years old male. These cell lines were cultured in proliferative conditions in a Growth Medium composed of 4:1 volumes of Dulbecco’s modified Eagle medium (DMEM, Gibco)/Medium 199 (Gibco), supplemented with 15% fetal bovine serum (Euroclone), 0.02 m HEPES buffer (Invitrogen), 1.4 mg L^−1^ vitamin B12 (Sigma); 0.03 mg L^−1^ ZnSO4 (Fisher Scientific), 0.055 mg L^−1^ dexamethasone (Sigma), 2.5 µg L^−1^ hepatocyte growth factor (Chemicon International) and 10 µg L^−1^ basic fibroblast growth factor (BioPioneer) in the presence of 1% Penicillin/streptomycin. Cells were tested for the presence of the PAS sequence and are all permissive for DUX4 expression.
All hiPSC clones were generated from reprogramming of primary fibroblasts as described in^30^. The C5 hiPSC clone (AG08498) was derived from human fibroblasts purchased from the Coriell Institute^31^. The 17706 hiPSC clones, 17706-L (long) and 17706-S (short, 2 D4Z4 units) were derived from a mosaic patient carrying one normal 4q35 allele and one shorter allele with 2 RU (FSHD1; female; age 56 at sampling). The 14,586 hiPSC clone was derived from a patient carrying two normal 4q35 alleles but one SMCHD1 mutation (c.573A > C; p.Q193P; FSHD2; male; age 67 at sampling). hiPSC colonies were grown and expanded in mTeSR1 medium (Stemcell Technologies) on BD Matrigel (BD Biosciences, 354,277) coated dishes as described^30,31^. Primary fibroblasts were provided by the Center for biological Resources (Department of Medical Genetics, La Timone Children’s hospital) with the DC-2024–6282 (sample collection) and N°IE-2013-710 (ethical approval, Assistance Publique des hôpitaux de Marseille) accreditation numbers. Informed consent was obtained for all patients participating in the study or legal guardian for minor participants.
All other materials and methods are described in the supplemental information.
Results
SUMOylation inhibition activates the DUX4 program in human myotubes
Given the repressive role of SUMOylation on Dux expression in mESCs, we explored the consequences of loss of SUMOylation in human muscle cells, in particular in the context of FSHD, which is characterized by aberrant expression of DUX4 in skeletal muscle. To this end, we treated myotubes derived from hTERT/CDK4-immortalized control and FSHD myoblasts with the TAK-981 SUMOylation inhibitor (henceforth called SUMOi) (Fig. 1a) and analyzed the expression of the pathogenic long DUX4 transcript (DUX4-fl) and some of its target genes by RT-qPCR. TAK-981 covalently binds to the SUMO-E1 Activating Enzyme complex (SAE1) and prevents the activation and transfer of SUMO proteins to SUMO E2 UBC9 and downstream protein targets^28^ (Fig. 1a). Immortalized myotubes were treated for 48 h (from day 4 to day 6 post-differentiation) with increasing doses of TAK-981 (0–500 nM, final concentration) and harvested for molecular analyses (Fig. 1b).
By western blotting, we observed a progressive and dose-dependent accumulation of free SUMO1 coupled with a decrease in SUMO1- and SUMO2/3-conjugated proteins in control and FSHD myotubes (Figs. 1c, S1a, b), attesting the efficacy of TAK-981 at tested concentrations. We next showed that SUMOylation inhibition is associated with a trend towards increased DUX4-fl expression, mainly in FSHD myotubes (Figs. 1d, e, S1c), with possible differences associated with the muscle-of-origin for available cell lines. Concomitantly, we observed a slight dose-dependent increase in the expression of some of the selected DUX4 target genes (ZSCAN4, TRIM43, MBD3L2) in control cells (12UBCT^29^, AB1079). Increase in DUX4 target gene expression is more pronounced and statistically significant in FSHD cells (12ABCT^29^, AB1080), in particular for high doses of TAK-981 (Fig. 1e). In addition, we noted a moderate but significant impact of SUMOi on the expression of the MyoG myogenic marker in both control and FSHD cells, with a maximal effect at higher doses of TAK-981 (Fig. 2a, b). We also reported a dose-dependent decreased expression in MyH2 and MyH3. Immunostaining of myofibers at Day6 with anti-Titin or anti-Myogenin antibodies showed no significant change in the proportion of myotubes in treated control or FSHD cells compared to mock-treated cells (Fig. 2c). Notably, SUMOi-treated cells exhibited a significant increase in the fusion index, indicating that inhibition of SUMOylation does not impair but rather enhances muscle differentiation in both control and FSHD cells (Fig. 2d, e).Fig. 2. Inhibition of SUMOylation impairs myogenic gene expression in control and FSHD1-patient derived myotubes. (a) MYOG, MYH2 and MYH3 expression analyses in control myotubes (12UBCT and AB1079). Fold- changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p *< 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001. (b) MYOG, MYH2 and MYH3 expression analyses in myotubes obtained from FSHD1 patients (12ABCT and AB1080). Fold-changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001. (c) Immunostaining of differentiated control (AB1079) and FSHD1 (12ABCT) myotubes collected at day 6 after TAK-981 treatment at indicated concentrations (50 and 100 mM from day 4 to 6). Cells were stained with DAPI, anti-Titin and anti-Myogenin antibodies. Scale bar = 50 µm. (d) Quantification of nuclei per myotube for each immortalized myoblast line at day 6 after TAK-981 treatment at indicated concentrations. Each violin plot represents the distribution of individual nuclei count values. Black horizontal lines mark the median and dashed lines indicate quartiles. Data were compared using Kruskal–Wallis tests with Dunn’s correction for multiple comparisons. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001.
SUMOylation inhibition enhances DUX4 expression in hiPSC-derived muscle cells
Next, we investigated the impact of SUMOi on DUX4-fl induction during muscle tissue differentiation. To this aim, we treated control and FSHD muscle cells derived from induced pluripotent cells (hiPSCs)^30^ over a period of 15 days post-differentiation with daily change in cell culture medium supplemented with TAK-981 or DMSO alone (Fig. 3a). Different concentrations of TAK-981 were used and cells were collected at the end of the differentiation procedure (day 30, D30) for RNA and protein extraction. Efficiency of SUMOylation inhibition was verified by western blotting (Fig. S2a-c).Fig. 3. Loss of SUMOylation triggers *DUX4 *activation in FSHD1 iPSC-derived myotubes. (a) Experimental design for hiPSC-derived myofibers. HiPSCs derived from a healthy donor or FSHD1 patient^30^ were differentiated for 30 days as described^53,54^. TAK-981 was added every day from day 15 to 30. Cell culture medium supplemented in DMSO or TAK-981 was changed every 24 h. At day 30, RNA and proteins were collected. (b–d) DUX4-fl and DUX4 target gene expression analyses in control AG08498^30^ (b), control 17,706-L^30^ (c) or FSHD1 17,706-S^30^ (d) hiPSC-derived myofibers. Healthy iPSC (17,706-L) and FSHD1 iPSC (17,706-S) are isogenic clones obtained from a mosaic patient (17,706). Fold-changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001 (e–f) MYOD, MYOG, MYH2 and MYH3 expression analyses in the control 17706-L (e) and FSHD1 17706-S (f) hiPSC-derived myofibers. Fold-changes were calculated as above. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001. (g-h) Immunoblots for the endogenous SMCHD1 protein in control AG08498 (g) and FSHD2 14,586 (h) hiPSC-derived myofibers treated with TAK-981 at indicated concentrations. Vinculin is used as a loading control. Arrows indicate SMCHD1 and its SUMOylated form. (i) DUX4-fl and DUX4 target gene expression analyses in the FSHD2 hiPSC-derived myofibers (14586). Fold-changes were calculated as above. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001. (j) Percentage of methylated (black) and unmethylated (white) (y-axis) CpG at the DR1 region located in the proximal part of the D4Z4 repeat. Methylation was determined for each CpG within the sequence of interest (31 CpGs). On the x-axis, histogram bars represent the mean percentage of methylated (black) or unmethylated (white) CpG of all CpG in the region of interest after sodium bisulfite sequencing analysis in control and FSHD hiPSC-derived myofibers after TAK-981 or DMSO treatment. This global methylation level was determined after sequencing of > 45,000 amplicons per condition.
In control cells, we observed a robust increase in DUX4-fl expression at the highest dose of TAK-981 (Fig. 3b), associated with increased expression of selected DUX4 target genes, MBD3L2 and LEUTX in particular. For FSHD1, we first compared the effect of SUMOi on the transcriptional activation of DUX4-fl and DUX4 target gene in hiPSC-derived muscle tissue for a mosaic patient for which healthy (Fig. 3c) and diseased (Fig. 3d) isogenic clones were isolated^30^. In both contexts, we observed a dose-dependent increase in DUX4-fl expression together with a significant increase in the expression of DUX4 target genes that is more consistent in FSHD cells (17706-S) and observed at low doses of TAK-981 (Fig. 3c, d). Similarly, expression of the MyoD (a DUX4 target) and MyoG myogenic factor and expression of Myosin genes (MyH2, MyH3) were decreased in both control and FSHD cells, at low doses of TAK-981 (Fig. 3e,f).
Impact of loss of SUMOylation on SMCHD1 and SMCHD1-deficient cells
The SMCHD1 protein contains six principal lysine residues (K784, K1373, K1496, K1958, K1976, K2002) modified by SUMO2/3^27^. In hiPSC-derived myofibers, SUMOylated SMCHD1 was detected as an upper band by western blotting (Fig. 3g, h). This upper band is no longer detected after TAK-981 treatment in either controls or FSHD cells^30^, showing that SMCHD1 SUMOylation is decreased upon TAK981 treatment.
We next assessed whether SMCHD1 loss of function in SMCHD1-deficient cells together with loss of SUMOylation might exacerbate DUX4-fl induction. We treated FSHD2 hiPSC-derived muscle cells from D15 to D30 of differentiation with SUMOi using hiPSC from a FSHD2 patient carrying a pathogenic SMCHD1 variant that abrogates the protein ATPase activity^30^ (Fig. 3i). DUX4 and its target genes exhibited expression profiles similar to those found in FSHD1 patient-derived myofibers (Fig. 3d,i). In addition, the presence of a loss of function variant in SMCHD1 does not seem to affect SUMOylation of the protein in basal conditions in FSHD2 hiPSC-derived myofibers (Fig. 3h). We concluded that SUMOylation depletion promotes DUX4-fl expression in both control and FSHD cells, regardless of SMCHD1 catalytic activity.
SUMOylation inhibition does not impact D4Z4 methylation profile
As the D4Z4 macrosatellite is differentially methylated at the DNA level between non-affected individuals and FSHD patients, we next investigated whether DUX4 induction upon TAK-981 resulted from D4Z4 hypomethylation. To this aim, hiPSC-derived muscle of two controls (healthy donor and healthy clone isolated from the mosaic patient), one FSHD1 patient (diseased clone from the mosaic patient) and one FSHD2 patient were exposed to the SUMOylation inhibitor from D15 to D30 post-differentiation as described (Fig. 3a). Genomic DNA was extracted at the end of the treatment and D4Z4 methylation level was analyzed by sodium bisulfite sequencing at the DR1 region which is hypomethylated in FSHD patients compared to non-affected individuals^23,24^. In all conditions, we compared cells treated at a final concentration of 50 nM TAK-981 to cells treated with DMSO only. Inhibition of SUMOylation does not impact the level of methylation at DR1 in all tested hiPSC derived-myofibers (Fig. 3j), suggesting that TAK-981-induced activation of DUX4 expression does not involve the hypomethylation of the repeat.
Loss of SUMOylation modulates expression of genes at the 4q35 locus
We then evaluated whether TAK-981-induced hypoSUMOylation only affects DUX4 expression or also the expression of other genes at the 4q35 locus. We analyzed expression of four different genes within a 7 Mb range of the D4Z4 region (WWC2) and located in different Topologically Associated Domains (TADs)^32^. We included genes previously implicated in FSHD such as DUX4c^33^, FRG2 ^33–35^, FRG1^33^ and FAT1^36^ (Fig. 4a). The expression of the most distal gene WWC2 is not impacted by TAK-981 treatment compared to mock-treated controls or FSHD immortalized myotubes (Fig. 4b, c). In contrast, we observed a robust and statistically significant upregulation of FRG2 in all cell types at low doses of TAK-981 and of FAT1 in both control and FSHD1 cells, whereas a more heterogenous effect on FRG1 expression as well as on DUX4c expression was observed depending on samples and doses of SUMOi (Fig. 4b-c).Fig. 4SUMOylation inhibition affects expression of other genes of the DUX4 locus in myotubes. (a) Schematic representation of human chromosome region 4q35. (b-c) FRG2, DUX4c, FRG1, FAT1 and WWC2 expression analyses in control myotubes 12UBCT and AB1079 (b) and in FSHD1 patient-derived myotubes 12ABCT and AB1080 (c). Fold-changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001.
We observed the same trend in hiPSC-derived muscle tissue when cells were treated during differentiation, in particular for FRG2 and FAT1, both in controls and FSHD cells (Fig. 5a–d). These observations overall suggest that SUMOylation inhibition contributes to the relaxation of the 4q35 locus.Fig. 5. Expressions of genes in DUX4 locus are affected in hiPSC-derived myofibers. (a-d) FRG2, DUX4c, FRG1, FAT1 and WWC2 expression analyses in control AG08498 (a), control 17,706-L (b), FSHD1 17,706-S (c) and FSHD2 14,586 (d) hiPSC-derived myofibers. Fold-changes were calculated relative to three housekeeping genes and normalized to the untreated (0) condition for each gene. Error bars represent mean ± SD. n = 3. Significance: one-way ANOVA testing. ns: not significant. * *p * < 0.05, ** *p * < 0.01, *** *p * < 0.001 and **** *p * < 0.0001.
Discussion
Post-translational modification by SUMO acts to stabilize cell identity in a variety of contexts^7,37–39^. In particular, hypoSUMOylation enhances pluripotency reprogramming in vitro* and *in vivo and suppression of SUMOylation promotes the spontaneous conversion of ESCs to a 2-cell-embryo-like (2C-like) state^7,8,39^. This transition involves two distinct mechanisms. First, hypoSUMOylation activates transcription of genomic loci regulated by the H3K9 histone methyltransferases Setdb1 and Suv39H1 by inducing a global decrease in H3K9 trimethylation^7,40–42^. Second, hypoSUMOylation leads to Dux derepression by interfering with the recruitment of Prc1.6 and Kap/Setdb1 at the Dux promoter^7^, together with impairing Dppa2/Dppa4 and histone H1 repressing activities^1,9,10^.
Mouse Dux and human DUX4 both encode a transcription factor that plays a key role in triggering zygotic genome activation (ZGA) in the early stages of embryogenesis^11,12^. In human somatic cells, DUX4 is normally silenced through epigenetic repression of the D4Z4 macrosatellite repeat array on chromosome 4q35, involving DNA methylation and repressive histone marks such as H3K9me3 and H3K27me3^22,43^. In FSHD, ectopic expression of the long DUX4 transcript (DUX4-fl) has been proposed as the main trigger leading to disease-associated muscle dysfunction^22^. Activation of DUX4-fl in a subset of muscle fibers (1/1000 nuclei) is subsequent to the reduction in the number of D4Z4 macrosatellite at the 4q35 locus in FSHD1^22^ or to the presence of a variant in SMCHD1 in FSHD2^25^, both leading to hypomethylation of D4Z4 and chromatin relaxation of the 4q35 locus. However, the mechanisms underlying burst of DUX4 transcriptional activation in muscle remain unknown.
A large number of transcription factors and chromatin regulators are SUMOylated, and SUMOylation is generally associated with transcriptional repression ^3–6,27^. SUMO facilitates the recruitment of SUMO interacting motif (SIM)-containing proteins^44,45^ such as histone deacetylases or the Setdb1 histone methyltransferase. Interestingly, the SMCHD1 protein contains six main lysine residues (K784, K1373, K1496, K1958, K1976, K2002) that are modified by SUMOylation and 15 minor acceptor lysines containing the consensus sequence required for SUMOylation^27^. Together, these findings point towards a role for SUMOylation in the regulation of the 4q35 locus and potentially of aberrant DUX4 expression in the context of FSHD. However, nothing is currently known on how SUMOylation might regulate SMCHD1 function. To date, no changes in global SUMOylation or in the levels of SUMOylation pathway components have been detected in FSHD cells^46,47^.
Recent development of a highly specific small molecule inhibitor of SUMOylation, TAK-981, proved to be a powerful tool for studying various facets of SUMOylation biology^28^. Notably, TAK-981 is able to induce a strong spontaneous type I interferon response in myeloid cells^48^, mirroring genetic inactivation of SUMOylation^49^. In line with this finding, TAK-981 was shown to activate antitumor immune response in pre-clinical models^48,50^ and in the clinics^51^. Here, we show that short or prolonged treatment of muscle cells in vitro with TAK-981 activates the DUX4 expression program in control cells and exacerbates DUX4 activation in cells from FSHD patients. Using cells from a patient affected with FSHD2 and carrying a variant in SMCHD1 altering the functional activity of the ATPase domain and showing a marked decrease in D4Z4 methylation^30^, we show that SMCHD1 loss of function and associated D4Z4 hypomethylation does not lead to increased TAK-981-induced DUX4 activation beyond levels observed in TAK-981-treated FSHD1 cells. These data suggest that TAK-981-induced activation of DUX4 expression is uncoupled from the D4Z4 methylation state.
In addition, we show that treatment with SUMOi increases expression of other genes located at the 4q35 locus such as the FRG2 non-coding RNA and FAT1, that might also contribute to the FSHD phenotype ^33–36^. This suggests that SUMOylation inhibition contributes to a more global chromatin relaxation in the vicinity of the D4Z4 array that particularly affects genes located within the same TAD and that show variable long-distance interactions depending on the cell context^32^.
Notably, recent research by an independent group investigated the link between SUMOylation and DUX4 regulation in FSHD, using ML-792, an alternative inhibitor of the SUMO-conjugation pathway, in cultured cells^52^. As observed in our cell models, the authors demonstrated that SMCHD1 is SUMOylated in both myoblasts and myotubes. Consistent with our observations, treatment with ML-792 induces DUX4 expression and a variable decrease in myogenic gene expression. Their analysis of SMCHD1 mutants lacking SUMOylation sites revealed no significant impact on SMCHD1 protein stability, nuclear localization, dimerization, or chromatin binding at the D4Z4 locus. Similarly, our results on FSHD2-patient derived cells with a loss-of-function of SMCHD1 did not uncover a definitive role for SUMOylated SMCHD1 in regulating the 4q35 region, indicating that further functional studies are needed^52^.
Overall, using different control or FSHD muscle cells, we provide here a proof of concept of the effect of SUMOylation in the regulation of DUX4 expression and epigenetic regulation of the 4q35 region. Our data suggest that manipulating SUMOylation, potentially through SENP deSUMOylase inhibition, may help silence inappropriate de-repression of DUX4, opening new avenues for investigating the contribution of SUMOylation in FSHD. Further studies will be essential to define the therapeutic relevance of such approaches.
Supplementary Information
Supplementary Information 1. Supplementary Information 2. Supplementary Information 3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Boulanger, M., Chakraborty, M., Tempe, D., Piechaczyk, M. & Bossis, G. SUMO and Transcriptional Regulation: The Lessons of Large-Scale Proteomic, Modifomic and Genomic Studies. Molecules 26, 10.3390/molecules 26040828 (2021).10.3390/molecules 26040828 PMC 791533533562565 · doi ↗ · pubmed ↗
- 2Cossec, J. C. et al. SUMO Safeguards Somatic and Pluripotent Cell Identities by Enforcing Distinct Chromatin States. Cell Stem Cell 23, 742–757 e 748, 10.1016/j.stem.2018.10.001 (2018).10.1016/j.stem.2018.10.00130401455 · doi ↗ · pubmed ↗
- 3Sheban, D. et al. SUM Oylation of linker histone H 1 drives chromatin condensation and restriction of embryonic cell fate identity. Mol Cell 82, 106–122 e 109, 10.1016/j.molcel.2021.11.011 (2022).10.1016/j.molcel.2021.11.011PMC 1182287234875212 · doi ↗ · pubmed ↗
- 4Gabriels, J. et al. Nucleotide sequence of the partially deleted D 4Z 4 locus in a patient with FSHD identifies a putative gene within each 3.3 kb element. Gene 236, 25–32 (1999).10.1016/s 0378-1119(99)00267-x 10433963 · doi ↗ · pubmed ↗
- 5Salsi, V., Magdinier, F. & Tupler, R. Does DNA Methylation Matter in FSHD? Genes (Basel)11, 10.3390/genes 11030258 (2020).10.3390/genes 11030258 PMC 714082332121044 · doi ↗ · pubmed ↗
- 6Nikolic, A. et al. Interpretation of the Epigenetic Signature of Facioscapulohumeral Muscular Dystrophy in Light of Genotype-Phenotype Studies. Int J Mol Sci 21, 10.3390/ijms 21072635 (2020).10.3390/ijms 21072635 PMC 717824832290091 · doi ↗ · pubmed ↗
- 7Salsi, V. et al. Nucleolar FRG 2 lnc RN As inhibit r RNA transcription and cytoplasmic translation, linking FSHD to dysregulation of muscle-specific protein synthesis. Nucleic Acids Res 53, 10.1093/nar/gkaf 643 (2025).10.1093/nar/gkaf 643PMC 1224277140637237 · doi ↗ · pubmed ↗
- 8Jagannathan, S., Ogata, Y., Gafken, P. R., Tapscott, S. J. & Bradley, R. K. Quantitative proteomics reveals key roles for post-transcriptional gene regulation in the molecular pathology of facioscapulohumeral muscular dystrophy. Elife 8, 10.7554/e Life.41740 (2019).10.7554/e Life.41740 PMC 634939930644821 · doi ↗ · pubmed ↗