Bloodstream infection with NDM-1/5 Enterobacter cloacae complex in China: diverse STs, multi-virulence systems and carbapenem resistance
Xinying Wang, Yujing Tian, Qun Zhang, Yan Jin, Chunhong Shao, Zhijun Zhang

TL;DR
This study examines carbapenem-resistant Enterobacter cloacae bloodstream infections in China, highlighting diverse strains and resistance genes.
Contribution
The study identifies ST171 as the dominant clone and highlights the role of IncX3/IncHI2 plasmids in spreading resistance.
Findings
ST171 was the most common sequence type among 13 isolates.
All isolates carried NDM-1 or NDM-5 and showed resistance to multiple antibiotics.
blaNDM-bearing plasmids were transferable to E. coli J53 via conjugation.
Abstract
To elucidate the molecular epidemiology, virulence repertoire and resistance gene characteristics of carbapenem-resistant Enterobacter cloacae complex (CRECC) in bloodstream infections (BSI), thereby providing evidence for precision therapy and infection control. We retrospectively collected 13 non-replicate CRECC-BSI isolates from January 2019 to December 2023 at a tertiary-care hospital in Shandong Province, China. Antimicrobial susceptibility was determined by broth microdilution; Illumina NovaSeq whole-genome sequencing was performed, and genomes were assembled with ABySS and GapCloser. ResFinder, VFDB, CGE and NCBI Pathogen Detection databases were used jointly to analyze resistance genes, virulence factors, plasmid replicons, MLST an SNP-based phylogenetic tree assessed inter-strain relatedness; while filter-mating assays determined the transferability of plasmids. A total of 13…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
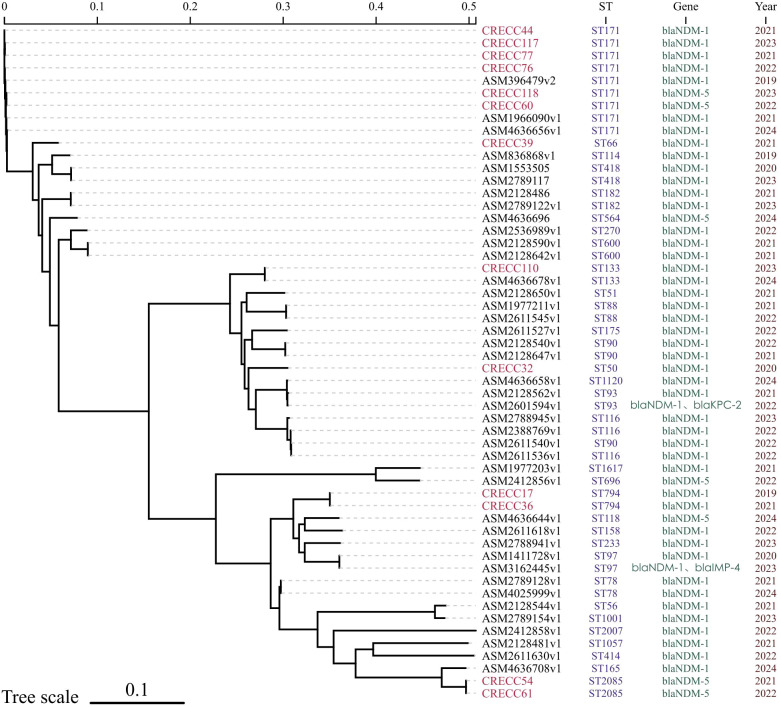 Figure 1
Figure 1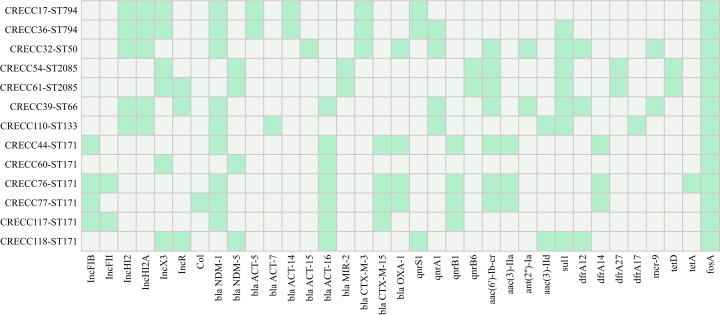 Figure 2
Figure 2| Patient ID | Year | Age (y) | Sex | Ward | Antimicrobial exposure prior to CRECC detection | Antimicrobial therapy after CRECC detection | Length of stay (d) | Source of BSI | Invasive procedures | ICU Admission | Outcome |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CRECC17 | 2019 | 43 | M | ICU | P, LEV, MEM, VA | SCF, TGC | 36 | Pulmonary | UC, MV | Yes | Survived |
| CRECC32 | 2020 | 2 | M | Hematology | IPM, LZD | TGC, AK, IPM | 20 | Unknown | CVC | No | Survived |
| CRECC36 | 2021 | 71 | M | EICU | TZP, MEM, TGC, SCF, CAZ, LZD, AK | TGC, MEM, LZD, LEV, ATM, CZA, CIP | 188 | Pulmonary | UC, CVC, MV | Yes | Survived |
| CRECC39 | 2021 | 67 | F | Hematology | LZD, IPM, MEM | SCF, TGC, ATM | 21 | Pulmonary | None | No | Survived |
| CRECC44 | 2021 | 54 | F | NICU | OX, CIP, TZP | IPM, ATM, FOS | 66 | Unknown | UC, CVC | Yes | Survived |
| CRECC54 | 2021 | 88 | M | ICU | LEV | CAZ, MEM, IPM | 171 | Pulmonary | UC, CVC | Yes | Survived |
| CRECC60 | 2022 | 58 | M | RICU | MEM | MEM, TGC | 25 | Unknown | UC, CVC, MV | Yes | Survived |
| CRECC61 | 2022 | 69 | M | ICU | MEM, CXM, LZD | MEM, LZD, PB, SCF, TGC, FEP | 96 | Unknown | UC, CVC, MV | Yes | Survived |
| CRECC76 | 2022 | 57 | M | RICU | TZP | SCF, TGC | 55 | Catheter-related | UC, CVC, MV | Yes | Died |
| CRECC77 | 2022 | 55 | M | Neurology | TZP, MFX | IPM, AK | 15 | Urinary | UC | No | Survived |
| CRECC110 | 2023 | 63 | M | Hematology | MFX, MEM, SCF, TGC | ATM, IPM | 24 | Pulmonary | None | No | Died |
| CRECC117 | 2023 | 86 | M | ICU | FOS, MEM | VA, MEM, FOS, TZP, TGC, PB, LZD | 187 | Urinary | UC, CVC, MV | Yes | Died |
| CRECC118 | 2023 | 91 | M | CICU | SCF | ATM, FOS, TZP | 19 | Urinary | UC | Yes | Survived |
| Isolation ID | Carbapenemase-encoding genes | ST type | CXM | SCF | CTX | CAZ | FEP | ATM | IPM | MEM | TZP | LEV | CIP | AK | TOB | SXT | PB | TGC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CRECC17 | NDM-1 | 794 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | 4 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | 2 |
| CRECC32 | NDM-1 | 50 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | 4 | ≥16 | ≥16 | ≥128/4 | 0.5 | 0.25 | ≤2 | 8 | >4/76 | ≤0.5 | ≤0.5 |
| CRECC36 | NDM-1 | 794 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | 4 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | ≤2 | <=1 | >4/76 | ≤0.5 | 2 |
| CRECC39 | NDM-1 | 66 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≤1 | ≥16 | ≥16 | ≥128/4 | 1 | 1 | ≤2 | 8 | >4/76 | ≤0.5 | 1 |
| CRECC44 | NDM-1 | 171 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≥64 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | 8 | ≥16 | ≤1/19 | ≤0.5 | 1 |
| CRECC54 | NDM-5 | 2085 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | 16 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | ≤2 | 8 | >4/76 | ≤0.5 | ≤2 |
| CRECC60 | NDM-5 | 171 | ≥64 | ≥64 | ≥64 | ≥64 | 16 | ≤1 | ≥16 | ≥16 | ≥128/4 | 4 | ≥4 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | 2 |
| CRECC61 | NDM-5 | 2085 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | 16 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | ≤2 | 8 | >4/76 | ≤0.5 | ≤2 |
| CRECC76 | NDM-1 | 171 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≥64 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | 4 | ≥16 | ≤1/19 | ≤0.5 | 1 |
| CRECC77 | NDM-1 | 171 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≥64 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | 8 | ≥16 | ≤1/19 | ≤0.5 | 2 |
| CRECC110 | NDM-1 | 133 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≤1 | ≥16 | ≥16 | ≥128/4 | 2 | 1 | ≤2 | 4 | >4/76 | ≤0.5 | ≤2 |
| CRECC117 | NDM-1 | 171 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≥64 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | ≤2 | 1 | ≤1/19 | ≤0.5 | 1 |
| CRECC118 | NDM-5 | 171 | ≥64 | ≥64 | ≥64 | ≥64 | ≥32 | ≤1 | ≥16 | ≥16 | ≥128/4 | ≥8 | ≥4 | ≤2 | 8 | >4/76 | ≤0.5 | ≤2 |
| CRECC44TC | NDM-1 | – | ≥64 | ≥64 | ≥64 | ≥64 | 2 | ≤1 | 2 | ≥16 | ≥128/4 | ≤0.12 | ≤0.25 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | ≤0.5 |
| CRECC54TC | NDM-5 | – | ≥64 | ≥64 | ≥64 | ≥64 | 8 | ≤1 | ≥16 | ≥16 | ≥128/4 | ≤0.12 | ≤0.25 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | ≤0.5 |
| CRECC61TC | NDM-5 | – | ≥64 | ≥64 | ≥64 | ≥64 | 16 | ≤1 | ≥16 | ≥16 | ≥128/4 | ≤0.12 | ≤0.25 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | ≤0.5 |
| CRECC76TC | NDM-1 | – | ≥64 | ≥64 | ≥64 | ≥64 | 4 | ≤1 | 8 | ≥16 | ≥128/4 | ≤0.12 | ≤0.25 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | ≤0.5 |
| CRECC77TC | NDM-1 | – | ≥64 | ≥64 | ≥64 | ≥64 | 2 | ≤1 | 8 | ≥16 | ≥128/4 | ≤0.12 | ≤0.25 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | ≤0.5 |
| J53 | – | – | 4 | ≤8 | ≤1 | ≤0.12 | ≤0.12 | ≤1 | ≤0.25 | ≤0.25 | ≤4/4 | ≤0.12 | ≤0.25 | ≤2 | ≤1 | ≤1/19 | ≤0.5 | ≤0.5 |
| VF classs | Relate genes | CRECC17 | CRECC32 | CRECC36 | CRECC39 | CRECC44 | CRECC54 | CRECC60 | CRECC61 | CRECC76 | CRECC77 | CRECC110 | CRECC117 | CRECC118 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ST794 | ST50 | ST794 | ST66 | ST171 | ST2085 | ST171 | ST2085 | ST171 | ST171 | ST133 | ST171 | ST171 | ||
| Adherence | ||||||||||||||
| Curli fibers |
| |||||||||||||
| P fimbriae |
| |||||||||||||
| Type I fimbriae |
| |||||||||||||
| Autotransporter |
| |||||||||||||
| Invasion | ||||||||||||||
| Invasion of brain endothelial cells (Ibes) |
| |||||||||||||
| Flagella(Burkholderia) |
| |||||||||||||
| Iron uptake | ||||||||||||||
| Aerobactin siderophore |
| |||||||||||||
| Heme uptake |
| |||||||||||||
| Iron/manganese transport |
| |||||||||||||
| Salmochelin siderophore |
| |||||||||||||
| Regulation |
| |||||||||||||
| Secretion system | ||||||||||||||
| SCI-I T6SS |
| |||||||||||||
| T2SS |
| |||||||||||||
| Toxin |
| |||||||||||||
| Antiphagocytosis | ||||||||||||||
| Capsular polysaccharide(Vibrio) |
| |||||||||||||
| Biofilm formation |
| |||||||||||||
| Immune evasion |
| |||||||||||||
| Serum resistance |
| |||||||||||||
| Stress adaptation |
| |||||||||||||
| Endotoxin |
| |||||||||||||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antimicrobial Resistance in Staphylococcus · Antibiotic Use and Resistance
Introduction
1
The Enterobacter cloacae complex (ECC) is a ubiquitous group of opportunistic pathogens that thrive in hospital environments, water and soil, and readily colonize the human gastrointestinal, respiratory and urinary tracts (Wendel et al., 2022). ECC comprises several clinically relevant species, most notably Enterobacter hormaechei, Enterobacter cloacae, Enterobacter asburiae and Enterobacter roggenkampii among others (Wu et al., 2025), and can cause a spectrum of healthcare-associated infections including bloodstream, urinary tract, wound, peritonitis and hospital-acquired pneumonia (Emeraud et al., 2024). Bloodstream infection (BSI) is the most severe manifestation, associated with high mortality, prolonged hospitalization and substantial healthcare costs (Gupta et al., 2024; Zhou et al., 2024).
The principal mechanisms of ECC resistance to β-lactam antibiotics are chromosomal AmpC enzyme derepression, and production of extended-spectrum β-lactamases (ESBLs) and carbapenemases (Annavajhala et al., 2019; Yang et al., 2022). These genes can be horizontally transferred via plasmids and integrons, accelerating the spread of resistant clones (Moraz et al., 2021; Fadare and Okoh, 2021). Consequently, the relentless rise of multidrug-resistant (MDR) ECC poses a severe global health threat. Carbapenem-resistant Enterobacterales (CRE) were classified among the pathogens of critical priority by the World Health Organization in 2024 (Antimicrobial Resistance Collaborators, 2022; Miller and Arias, 2024) Nation-wide surveillance underscores this trend: CHINET data show that the prevalence of carbapenem-resistant Enterobacter cloacae complex (CRECC) in China rose from 0.7% in 2007 to ~8% in 2024, making it the third most common nosocomial CRE after Klebsiella pneumoniae and Escherichia coli (Chen et al., 2021; Davin-Regli et al., 2019; China Antimicrobial Surveillance Network, 2024).
Plasmid-mediated New Delhi metallo-β-lactamase (NDM) production is now the dominant resistance mechanism (Bush and Bradford, 2019). NDM enzymes hydrolyze penicillins, cephalosporins and carbapenems, exhibit only weak activity against aztreonam, and remain unaffected by classical β-lactamase inhibitors such as avibactam but are inhibited by EDTA (Nagulapalli Venkata et al., 2021). Consequently, NDM-producing ECC isolates are frequently susceptible only to tigecycline and polymyxins, with occasional aztreonam sensitivity. Epidemiological studies show that healthcare-associated outbreaks of NDM-producing ECC are driven by a restricted set of MLST clones-most notably ST78, ST171 and ST182 (Gomez-Simmonds et al., 2018)-that acquire blaNDM on IncX3, IncHI2 or IncFII plasmids (Mavroidi et al., 2023). Once formed, these high-risk ST–plasmid pairings spread rapidly: the ST171-IncX3 complex has driven repeated hospital outbreaks across India, Japan and the United States (Wang et al., 2018), while ST182, tightly associated with IncFII, triggered the largest European ECC epidemic on record in Greece (Gartzonika et al., 2023). Collectively, these findings underscore that the intersection of a susceptible clone with a promiscuous, NDM-carrying replicon acts as a molecular trans-regional escalation switch for ECC, providing clear targets for genomic surveillance.
In China, nationwide molecular epidemiological and virulence data on CRECC-BSI remain scarce. Using whole-genome sequencing and comprehensive in-silico virulence/plasmid profiling, this retrospective, single-center study (n = 13 isolates) systematically delineates the resistance, virulence and transmission patterns of CRECC bloodstream isolates from a tertiary-care hospital in Shandong Province, providing an evidence base for targeted therapy and infection-control strategies.
Materials and methods
2
Collection and identification of bacterial isolates
2.1
Thirteen non-replicate CRECC isolates first recovered from blood cultures between January 2019 and December 2023 at our hospital were collected. Carbapenem resistance was defined as a MIC ≥ 4 mg/L for imipenem or meropenem (Senneville et al., 2024). Species identification was initially performed using Autof ms1000 (Autobio, China). To achieve species-level resolution within the Enterobacter cloacae complex, genome-to-genome distances were calculated against all relevant type strains using the isDDH web server (https://ggdc.dsmz.de/ggdc.php) with GGDC formula 2. A digital DNA-DNA hybridization (dDDH) value of ≥ 70% was considered indicative of the same species.
Antimicrobial susceptibility testing
2.2
Antimicrobial susceptibility of all isolates was tested using the VITEK 2 system(bioMérieux, Marcy-l’Étoile, France) against the following agents: cefuroxime (CXM), cefoperazone-sulbactam (SCF), cefotaxime (CTX), ceftazidime (CAZ), cefepime (FEP), aztreonam (ATM), imipenem (IPM), meropenem (MEM), piperacillin-tazobactam (TZP), levofloxacin (LEV), ciprofloxacin (CIP), amikacin (AK), tobramycin (TOB), sulfamethoxazole-trimethoprim (SXT), tigecycline (TGC) and polymyxin B (PB). Antimicrobial susceptibility testing results were interpreted according to the EUCAST (European Committee on Antimicrobial Susceptibility Testing, 2024) criteria for tigecycline and polymyxin B, and the CLSI M100-S34 (Clinical and Laboratory Standards Institute, 2024) criteria for the remaining antibiotics. Escherichia coli ATCC 25922 served as the quality-control strain.
Whole-genome sequencing and bioinformatics analysis
2.3
Fresh CRECC colonies from blood agar were selected and high-quality genomic DNA extracted using the OMEGA Bacterial DNA Kit (Omega Bio-tek, Inc., Norcross, GA, USA) following the manufacturer’s instructions. DNA was sent to Shanghai Baizelong Biotechnology Co., Ltd. for library preparation and sequencing on the Illumina NovaSeq platform. Reads were quality-trimmed and assembled with ABySS using a multi-k-mer strategy; gaps were closed and single-nucleotide errors corrected with GapCloser. Resistance genes were identified using ResFinder (http://www.genomicepidemiology.org/services/). Plasmid replicons were extracted via BAcWGSTdb, and virulence factors were screened with VFAnalyst (http://www.mgc.ac.cn/VFs/, accessed).
Multilocus sequence typing and phylogenetic analysis
2.4
Seven housekeeping loci (dnaA, fusA, gyrB, leuS, pyrG, rplB and rpoB) were extracted (Miyoshi-Akiyama et al., 2013) and queried against the Enterobacter cloacae PubMLST database to assign sequence types (STs). For phylogenomic analyses, 41 publicly available CRECC blood isolates from mainland China were downloaded from NCBI Pathogen Detection (http://www.ncbi.nlm.nih.gov/pathogens/); core-genome SNPs were called with CSI Phylogeny (CGE, DTU) - chosen for its robust performance within the Enterobacteriaceae MLST framework and previously validated reliability on ECC genomes and a phylogenetic tree was constructed. The final phylogenetic tree was visualized and annotated using ChiPlot (Xie et al., 2023).
Conjugation assay
2.5
Donor CRECC and sodium-azide-resistant E. coli J53 were mixed 1:1 (500 µl each), filter-mated for 4 h, and plated on Mueller–Hinton agar containing IPM (4 mg/L) plus sodium azide (50 mg/L). Transconjugants were retained only when they met all three criteria: (1) IPM or MEM MIC ≥ 2 mg/L; (2) tolerance to sodium azide; and (3) sequencing-positive for blaNDM.
Results
3
Clinical characteristics
3.1
From January 2019 to December 2023, 13 patients with CRECC-BSI were enrolled. Eleven (84.6%) were male and two (15.4%) female; the median age was 63 years (range 2-91). Nine patients (69.2%) had been admitted to the ICU, 11 (84.6%) had undergone invasive procedures such as central-venous catheterization, mechanical ventilation or indwelling urinary catheterization, and eight (61.5%) had received carbapenems (defined as ≥ 48 h intravenous therapy before the first positive blood culture) prior to the positive blood culture. The presumed sources of infection were pulmonary (n = 5), urinary tract (n = 3) and unknown (n = 5) (Table 1).
Species identification and phylogeny analysis
3.2
MALDI-TOF MS accurately assigned all 13 isolates to the Enterobacter cloacae complex (100% concordance at the complex level).However, at the species level, only 4/13 designations were confirmed by the genome-based digital DNA-DNA hybridization (dDDH) analysis (30.8% concordance), demonstrating the limited resolution of MALDI-TOF within this complex (Supplementary Table S1) MLST revealed five sequence types: ST171 predominated (6/13, 46.2%), followed by ST794 and ST2085 (2/13 each, 15.4%), and ST50, ST66 and ST133 (1 isolate each). When combined with 41 bloodstream CRECC sequences downloaded from NCBI, ST171 remained the most common type nationwide (9/54) and all carried blaNDM-1 (Figure 1). cgMLST uncovered the genetic divergence among strains of the ST171 clone : pairwise core-genome SNP distances ranged from 4 to 63 (median 11) (Supplementary Table S2).
Phylogenetictreeof 54 CRECC strains. Red color denotes isolates obtained in our hospital.
Resistance genes, phenotypes and conjugation analysis
3.3
Whole-genome analysis revealed that all isolates carried blaNDM: blaNDM-1 in 9 isolates (69.2%) and blaNDM-5 in 4 (30.8%). Additional resistance genes were detected at the following frequencies: blaACT-16 (53.8%), blaCTX-M-15 (30.8%), blaCTX-M-3 (23.1%), blaOXA-1 (30.8%), aac(6′)-Ib-cr (53.8%), aac(3)-II (23.1%), qnrB1/qnrA1 (each 30.8%), sul1 (53.8%) and mcr-9 (15.4%) (Figure 2). Antimicrobial susceptibility testing showed that all 13 isolates were resistant to cephalosporins, carbapenems and β-lactam/β-lactamase-inhibitor combinations; resistance rates to LEV, CIP, TO, SXT and ATM were 84.6%, 92.3%, 61.5%, 53.8% and 46.2%, respectively. AK, TGC and PB retained 100% susceptibility (Table 2). In filter-mating assays using the 13 CRECC isolates as donors and E. coli J53 as recipient, five (38.5%) successfully transferred blaNDM, yielding transconjugants CRECC44, 54, 61, 76 and 77. Post-conjugation, recipient MICs rose from ≤ 0.25 mg/L to 2–≥ 16 mg/L for IPM and ≥ 16 mg/L for MEM, and sequencing confirmed the presence of the same blaNDM subtype as the donor (Table 2).
Heat-map displaying plasmid replicon profiles (left panel) and resistance-gene content (right panel) of 13 CRECC bloodstream isolates. X-axis represents plasmid replicons and resistance genes, Y-axis shows the strain sequence numbers, Green fill indicates presence of the corresponding gene; Grey fill indicates absence.
Plasmid replicons and virulence profiles
3.4
IncX3 was the most prevalent plasmid replicon (6/13, 46.2%), followed by IncHI2/HI2A (5/13, 38.5%) and IncR (4/13, 30.8%), with three isolates co-harboring IncX3+IncR (Supplementary Table S1). Among the five successful transconjugants, a single replicon was consistently retained: IncFII for blaNDM-1 (ST171) and IncX3 for blaNDM-5 (ST2085). For virulence systems, all strains carried adherence, iron/manganese transport, biofilm formation and T6SS modules; isolates CRECC76, 77, 110 and 117 harbored the complete set of screened virulence genes (Table 3).
Discussion
4
BSI is a life-threatening condition caused by the invasion of pathogenic microorganisms into the circulation and remains one of the most severe complications in clinical practice. Its high mortality, coupled with the global spread of multidrug-resistant organisms, poses formidable challenges to public health (Temkin et al., 2024). Recent diagnostic and therapeutic advances have lagged behind the rapid emergence of resistant pathogens, underscoring the need for improved prevention and management strategies. CRECC-BSI is closely linked to prior CRECC colonization or infection, extensive antibiotic exposure (especially carbapenems) and invasive procedures that disrupt physiological barriers (Lumbreras-Iglesias et al., 2023). In this retrospective cohort of 13 patients, 84.6% were male, 84.6% underwent invasive interventions, and 62.9% were admitted to the ICU; these figures that align with the Italian multicenter CR-GNB BSI report (Falcone et al., 2023). Pulmonary (38.5%) and urinary-tract (23.1%) sources predominated, indicating that airway- and catheter-related infections are the main portals of entry in our hospital. Notably, 61.5% of patients had received carbapenems for ≥48 h before blood cultures became positive, a selective pressure that favors resistant clones.
In our results, all 13 isolates carried blaNDM (NDM-1 69.2%, NDM-5 30.8%); their geographical distribution differs markedly from the KPC predominance in North America and the VIM/OXA-48 prevalence in Europe, but aligns with the ongoing NDM surge reported in southern, north-eastern and north-western China (Chen et al., 2021; Han et al., 2023; Cai et al., 2024; Shi et al., 2017; Tian et al., 2020), underscoring pronounced regional divergence of CRECC resistance. In addition to carbapenemases, each isolate carried an AmpC β-lactamase of distinct subtype (ACT-16; n = 7, ACT-14; n = 2, MIR-2; n = 2, ACT-15; n = 1, ACT-5; n = 1, ACT-7; n = 1), while 50% harbored ESBLs (CTX-M-15 30.8%, CTX-M-3 23.1%), consistent with a previous report by Cai et al (Cai et al., 2024). Detection of two mcr-9-positive isolates further restricts polymyxin options and complicates therapeutic choices.
MLST revealed that ST171 accounted for 46.2% of isolates; in-hospital deaths were associated with this lineage, consistent with US and Japanese reports that designate ST171 as a hyper-virulent, highly transmissible clone (Pereira et al., 2019; Harada et al., 2017; Ghazawi et al., 2024). Although three isolate pairs differed by ≤5 SNPs and may represent short-term transmission links (Snitkin et al., 2012), the majority showed >20 SNP differences, indicating absence of a single-strain nosocomial outbreak. ST93, frequently reported in China, was not detected (Zhao et al., 2020). NCBI data reveal that Chinese bloodstream CRECC isolates exhibit diverse, sporadic STs, mirroring the polyclonal pattern observed here (Gomez-Simmonds et al., 2016). Based on the results of the conjugation assays, five donor strains harboring blaNDM successfully transferred their resistance plasmids to recipient E. coli J53, yielding transconjugants with ≥8-fold MIC increases for IPM and MEM compared with the recipient. Despite donors harboring multiple replicons, every transconjugant carried only a single plasmid replicon. IncFII for blaNDM-1 (ST171) or IncX3 for blaNDM-5 (ST2085). These results confirm the superior conjugative fitness of both replicons. They also align with global surveillance data identifying IncFII and IncX3 as the primary vehicles for blaNDM-1 and blaNDM-5 dissemination, respectively (Carattoli, 2013; Mouftah et al., 2019). These data provide direct experimental evidence that plasmid-mediated carbapenem resistance can be functionally transferred within our hospital environment.
Our results shown every isolate harbored the type VI secretion system (T6SS), a key virulence module that directly contributes to bacterial pathogenicity and host interaction, thereby posing a major clinical threat (Ghazawi et al., 2024; Theriault et al., 2021). Additionally, the adherence system was detected at varying levels; by promoting cell adhesion and interacting with the host immune response, it amplifies inflammatory reactions. The diverse virulence repertoire of CRECC strains thus complicates both the treatment and control of bloodstream infections (Azam et al., 2023). The 13 CRECC isolates harbored multiple plasmid replicon types, with IncHI2, IncHI2A and IncX3 being the most prevalent — mirroring reports from North America and Europe (Wu et al., 2025). IncX3 was detected most frequently, in line with a recent nationwide survey in China; its small size and high stability facilitate rapid inter-strain transfer of resistance genes (Wang et al., 2018). These findings underscore the need for early rectal screening of carbapenem-resistant organisms in high-risk patients, minimization of invasive procedures, prompt device replacement with regular microbiological surveillance, and enhancement of host immunity to curtail the spread of CRECC.
Conclusion
5
Our institutional CRECC-BSI isolates exhibited diverse sequence types, a complex plasmid repertoire and multiple virulence genes. ST171 was the dominant clone, NDM-1 the principal carbapenemase, and IncX3/IncHI2 plasmids the key vehicles for resistance-gene dissemination. Enhanced surveillance of this high-risk lineage and interventions against plasmid-mediated horizontal transfer are essential to curb further spread of CRECC in health-care settings.
Limitations
6
This study has several limitations. First, its retrospective, single-center design may introduce selection bias and limit the generalizability of our findings. Second, the relatively small sample size may not fully represent the national molecular epidemiology of CRECC. Future work should enlarge the isolate collection and integrate multicenter prospective data to obtain a comprehensive picture of CRECC trends and resistance mechanisms.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Annavajhala M. K. Gomez-Simmonds A. Uhlemann A. C. (2019). Multidrug-resistant Enterobacter cloacae complex emerging as a global, diversifying threat. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.00044, PMID: 30766518 PMC 6365427 · doi ↗ · pubmed ↗
- 2Antimicrobial Resistance Collaborators (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399, 629–655. doi: 10.1016/S 0140-6736(21)02724-0, PMID: 35065702 PMC 8841637 · doi ↗ · pubmed ↗
- 3Azam M. W. Zarrilli R. Khan A. U. (2023). Updates on the virulence factors produced by multidrug-resistant Enterobacterales and strategies to control their infections. Microorganisms 11, 1901. doi: 10.3390/microorganisms 11081901, PMID: 37630461 PMC 10456890 · doi ↗ · pubmed ↗
- 4Bush K. Bradford P. A. (2019). Interplay between β-lactamases and new β-lactamase inhibitors. Nat. Rev. Microbiol. 17, 295–306. doi: 10.1038/s 41579-019-0159-8, PMID: 30837684 · doi ↗ · pubmed ↗
- 5Cai S. Quan J. Wang Z. Hu H. Han X. Jiang Y. . (2024). High prevalence of carbapenem-resistant Enterobacter cloacae complex in a tertiary hospital over a decade. Microbiol. Spectr. 12, e 0078024. doi: 10.1128/spectrum.00780-24, PMID: 39475294 PMC 11619405 · doi ↗ · pubmed ↗
- 6Carattoli A. (2013). Plasmids and the spread of resistance. Int. J. Med. Microbiol. 303, 298–304. doi: 10.1016/j.ijmm.2013.02.001, PMID: 23499304 · doi ↗ · pubmed ↗
- 7Chen J. Tian S. Nian H. Wang R. Li F. Jiang N. . (2021). Carbapenem-resistant Enterobacter cloacae complex in a tertiary hospital in Northeast China 2010-2019. BMC Infect. diseases. 21, 611. doi: 10.1186/s 12879-021-06250-0, PMID: 34174823 PMC 8235818 · doi ↗ · pubmed ↗
- 8China Antimicrobial Surveillance Network . Available online at: https://www.chinets.com (Accessed October 10, 2025).