Biohealing through biocalcification by urolytic bacteria Bacillus subtilis ATCC 6633 on marble surfaces
Turkan Dal Bicer, Seval Cing Yildirim, Fadime Nulufer Kivilcim, Ahmet Gultek

TL;DR
Ureolytic bacteria can heal marble surfaces by forming calcium carbonate, with live bacteria and calcium chloride promoting stable crystal formation.
Contribution
Demonstrates how bacterial viability and calcium source influence biocalcification for sustainable marble restoration.
Findings
Live cells and calcium chloride promote calcite and aragonite formation, while dead cells and calcium acetate favor vaterite.
CO₂ pre-treatment enhances uniform CaCO₃ nucleation and reduces pore depth on marble surfaces.
Biocalcification partially fills cracks and micropores, offering a sustainable method for marble conservation.
Abstract
This study investigates the biocalcification potential of Bacillus subtilis ATCC 6633, a ureolytic bacterium, for the biohealing of marble surfaces through calcium carbonate (CaCO₃) precipitation. Comparative experiments were conducted using live and dead bacterial cells on CO₂-pre-treated and untreated marble samples, with calcium chloride and calcium acetate employed as calcium sources, to evaluate their effects on crystal polymorphism and surface modification. The results show that bacterial viability and calcium source jointly influence mineral phase formation, with live cells predominantly promoting the formation of stable calcite and aragonite, whereas dead cells and calcium acetate favor the formation of metastable vaterite. Microstructural and mineralogical analyses using scanning electron microscopy (SEM), energy-dispersive X-ray spectroscopy (EDX), X-ray diffraction (XRD), and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 10
Figure 10 Figure 11
Figure 11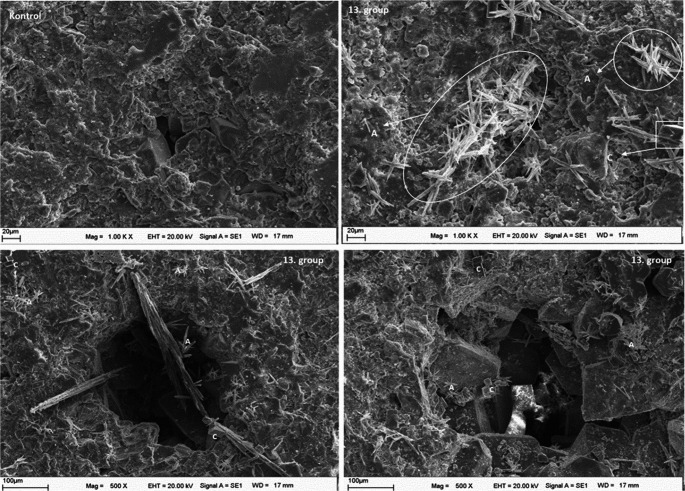 Figure 12
Figure 12 Figure 13
Figure 13 Figure 14
Figure 14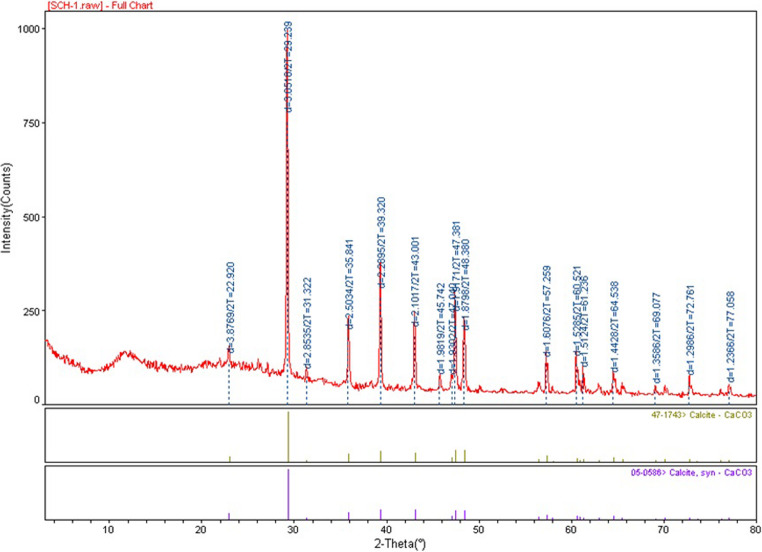 Figure 15
Figure 15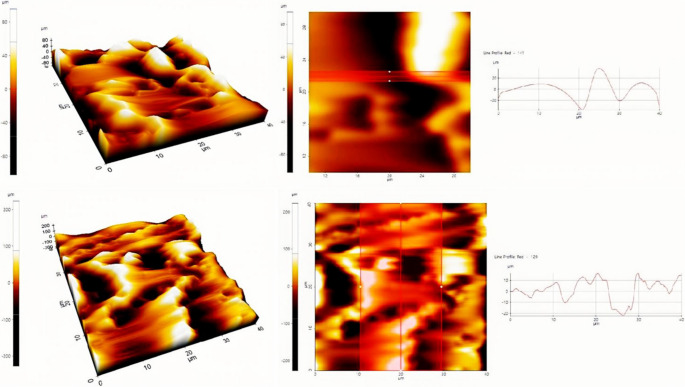 Figure 16
Figure 16 Figure 1
Figure 1 Figure 2
Figure 2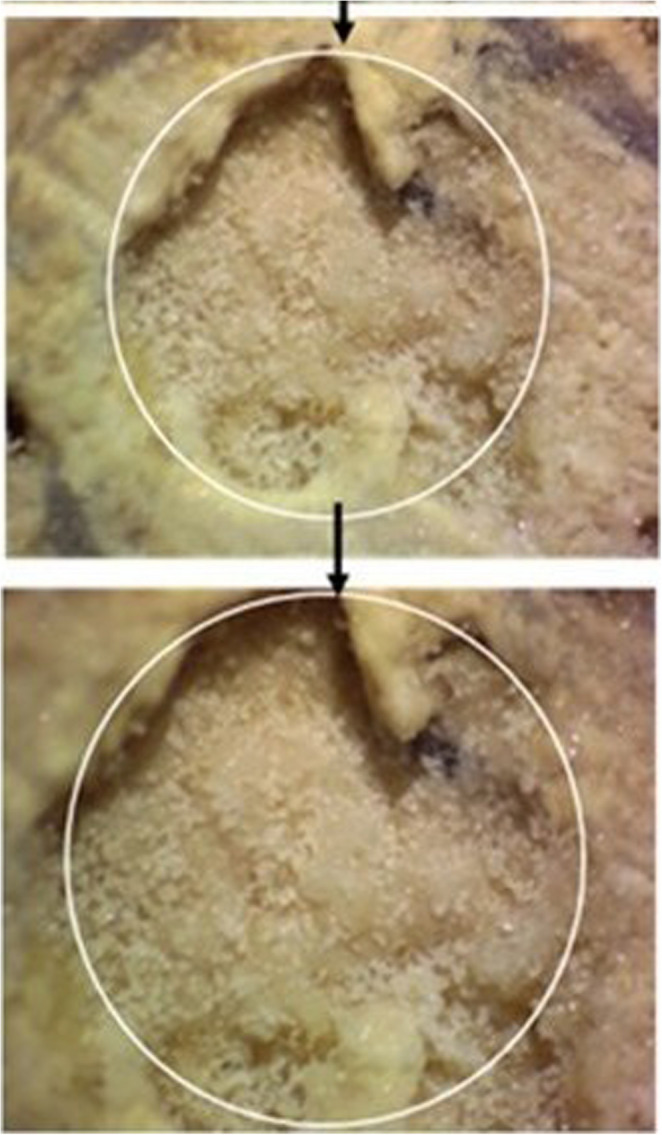 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5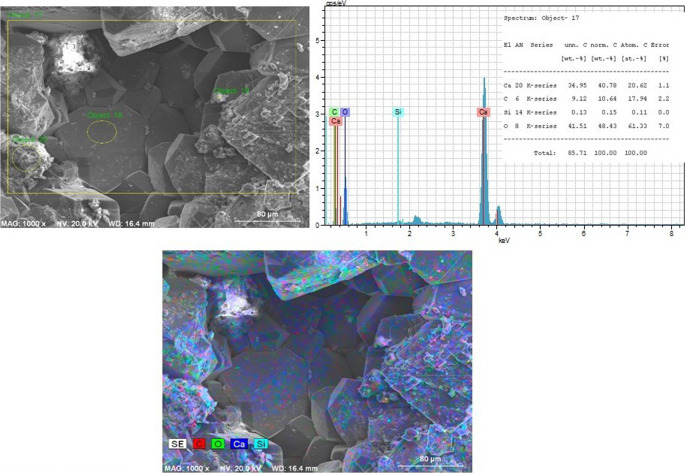 Figure 6
Figure 6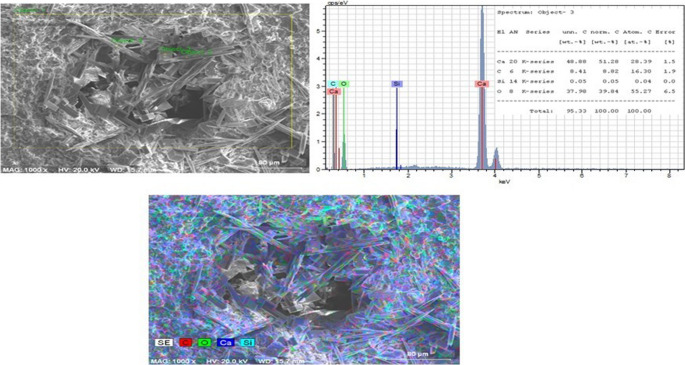 Figure 7
Figure 7 Figure 8
Figure 8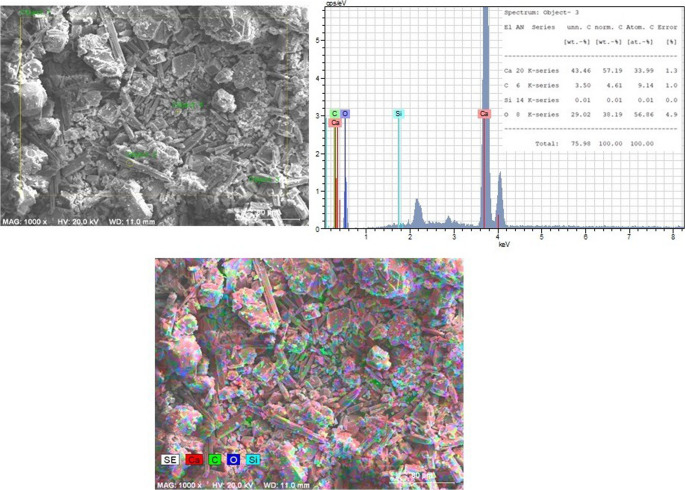 Figure 9
Figure 9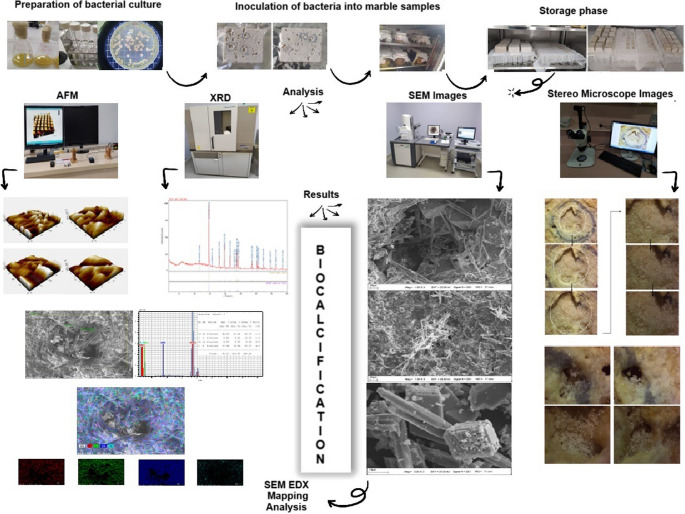 Figure 17
Figure 17- —Inonu University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Applications in Construction Materials · Building materials and conservation · Calcium Carbonate Crystallization and Inhibition
Introduction
Biohealing represents an emerging strategy in material science and conservation, leveraging the metabolic activity of microorganisms to repair and reinforce structural materials. Among these, biocalcification—a process driven by urea-hydrolyzing bacteria such as Bacillus subtilis has gained attention for its ability to induce calcium carbonate (CaCO₃) precipitation on mineral surfaces (Rajasekar et al. 2024; Sreekala et al. 2024). This bacterium metabolizes urea into carbonate ions, promoting calcite crystal formation, particularly on marble. By harnessing such microbial pathways, biohealing not only mitigates deterioration caused by environmental stressors but also enhances the durability and sustainability of marble-based architectural heritage. This interdisciplinary approach highlights the synergy between microbiology and materials engineering, offering a novel and eco-friendly method for preserving cultural assets.
Marble and limestone have long been used in historical monuments due to their durability and aesthetic value. However, these structures are exposed to environmental stressors and pollutants that gradually degrade the stone. In the absence of restoration, such deterioration can lead to substantial structural damage (Liu et al. 2011). Marble, primarily composed of calcium carbonate (CaCO₃), has low solubility in water but readily reacts with acids generated by acid rain to form soluble salts. This reaction causes material loss, pore formation, surface abrasion, and discoloration (Naidu et al. 2016). Numerous historical buildings have suffered such damage and remain at risk of further deterioration if not properly restored. Although several restoration methods have been developed, traditional techniques still exhibit critical limitations (Liu et al. 2011).
Acrylic and epoxy resins, commonly used for surface coating, can produce incompatible and potentially harmful films. Furthermore, the widespread use of organic solvents contributes to environmental pollution (Down et al. 1996) and poses health risks, while also diminishing the aesthetic quality of stone surfaces. The high cost and limited longevity of these materials have encouraged the search for sustainable alternatives (Luo et al. 2024). In this context, ureolytic bacteria-mediated CaCO₃ precipitation has emerged as a promising bio-based repair mechanism. Studies have demonstrated that bacterial facilitation enhances CaCO₃ formation, effectively filling cracks and pores, reducing water permeability, and extending structural lifespan (Sumathi et al. 2020; Zhong et al. 2022; Luhar et al. 2022). The goal of such investigations is to employ microbial self-healing processes to restore the physical integrity of structures (Seifan et al. 2016).
Microbially induced carbonate precipitation (MICP) refers to the mineralization process driven by microbial metabolic activity (Ahmad et al. 2024). During MICP, urease-producing bacteria hydrolyze urea to generate carbonate ions (CO₃²⁻), which react with calcium ions (Ca²⁺) in the environment to form CaCO₃ precipitates (De Muynck et al. 2010; Achal and Pan 2014). The process is influenced by various environmental and biochemical parameters, including pH, temperature, ion concentration, the carbonate-to-calcium ratio, and the presence of additives (Liendo et al. 2022). Among the polymorphs of CaCO₃—calcite, aragonite, and vaterite—calcite is the most thermodynamically stable form.
In recent years, increasing attention has been paid to the role of carbon dioxide (CO₂) in enhancing biomineralization efficiency (Isar 2020; Polat 2019). When applied as a surface pre-treatment, CO₂ can modify marble surface chemistry and influence local carbonate equilibria, which in turn promotes more uniform CaCO₃ crystal formation. However, excessive CO₂ concentrations or prolonged exposure may drive the system toward high supersaturation, leading to the formation of larger but less adherent crystals (Feng et al. 2007). In addition to gas-related parameters, the calcium source plays a critical role in controlling the morphology and density of precipitated crystals (Anbu et al. 2016).
Calcification can occur through different nucleation pathways, depending on bacterial viability and surface properties, as microbial cell surfaces and extracellular components may serve as nucleation sites for carbonate formation (Keren-Paz et al. 2022; Zhu and Dittrich 2016). In contrast, dead bacterial cells and cellular remnants can promote carbonate precipitation through surface-mediated (passive) nucleation mechanisms, as mineral formation occurs on pre-existing cellular templates in the absence of metabolic activity, although the role of dead cells in biomineralization remains less explored (Robles-Fernández et al. 2022).
The present study investigates the potential of Bacillus subtilis ATCC 6633 to repair marble cracks via MICP. B. subtilis is a Gram-positive, spore-forming bacterium known for its resilience and environmental adaptability. Owing to its well-characterized genome, it serves as a model organism for studies on bacterial physiology, biofilm formation, quorum sensing, and antibiotic resistance (Su et al. 2020). In this study, both live and dead B. subtilis cells were evaluated to determine their respective roles in calcium carbonate precipitation, and the influence of CO₂ pre-treatment on biomineralization efficiency was systematically examined. Controlling CO₂ concentration is thus essential to optimize the microstructure and adhesion of the biogenic calcite layer formed on marble surfaces.
Materials and methods
Chemical materials and marble samples
The chemical reagents used in this study included urea (CO(NH₂)₂), yeast extract, D-glucose monohydrate (C₆H₁₂O₆·H₂O), calcium acetate ((CH₃COO)₂Ca), calcium chloride dihydrate (CaCl₂·2 H₂O), sodium hydroxide (NaOH), physiological saline solution (0.9% NaCl, w/v), and ethyl alcohol (C₂H₅OH).
Marble samples were obtained from a private marble processing company. Square marble specimens measuring 2 × 2 cm and 4 × 4 cm were prepared by cutting from larger blocks. For each experimental group, a total of 50 marble samples were used. Prior to experimental treatments, all marble specimens were stored in clean, dry containers under ambient laboratory conditions (Fig. 1).Fig. 1. Representative macroscopic images of untreated marble samples before experimental treatment and bacterial inoculation
Microorganism and culture conditions
In this study, Bacillus subtilis ATCC 6633, obtained from the American Type Culture Collection (ATCC), was used. The strain was routinely cultured on brain heart infusion agar (BHIA) at 37 °C. Short-term storage of the cultures was carried out at 4 °C until further use.
Preparation of marble samples
Square marble samples measuring 4 × 4 cm were immersed in 70% (v/v) ethanol for 30 min to ensure surface sterilization. During immersion, the samples were rotated at 15-min intervals to allow uniform contact of all surfaces with the alcohol solution. Following sterilization, the marble samples were removed from the solution and dried in an oven at 40 °C for 6 h. During drying, the samples were periodically rotated to ensure complete and uniform drying of all surfaces. After drying, both surfaces of each marble sample were exposed to ultraviolet (UV) radiation for 30 min. The same sterilization and preparation procedures were applied to the 2 × 2 cm marble samples. Following UV treatment, the samples were transferred to sterile containers and stored aseptically until use in subsequent experiments. For surface pre-conditioning, half of the prepared marble samples were placed in a controlled CO₂ chamber maintained at 75% relative humidity and 5% CO₂ for 30 days.
Bacterial inoculation
Prior to bacterial calcification experiments, Bacillus subtilis was inoculated into brain heart infusion broth (BHIB) and incubated for 24 h at 37 °C. Subsequently, 0.1 mL of this culture was transferred into 20 mL of fresh BHIB and incubated for an additional 24 h at 37 °C with shaking at 110 rpm. Viable cell counts were determined using the standard plate count method. Serial dilutions of the bacterial culture were prepared in sterile physiological saline solution (0.9% NaCl, w/v). Aliquots of 100 µL from appropriate dilutions were spread evenly onto brain heart infusion agar (BHIA) plates using a sterile L-shaped spreader. The plates were incubated at 37 °C for 24 h, after which colony-forming units (CFU) were counted, and the viable cell concentration of the bacterial culture was calculated.
Preparation of media used in bacterial calcification and sample application
Preparation of the inoculation medium
The inoculation medium consisted of yeast extract (25 g/L) and urea (25 g/L). For experiments involving 4 × 4 cm marble samples, 18 sterile glass jars with a maximum volume of 300 mL were prepared. Similarly, for experiments using 2 × 2 cm marble samples, 18 sterile glass jars with a maximum volume of 100 mL were used. Volumes of 120 mL and 15 mL of inoculation medium were added to the large and small jars, respectively. All media were sterilized by autoclaving at 121 °C for 20 min.
Preparation of the precipitation media
Two different precipitation media were prepared to investigate calcium carbonate formation. The first medium contained calcium acetate [Ca(CH₃COO)₂] (6 g/L), glucose (2 g/L), yeast extract (2 g/L), and urea (12 g/L). The second medium consisted of calcium chloride dihydrate (CaCl₂·2 H₂O) (60 g/L) and urea (24 g/L). The prepared media were distributed into sterile glass jars of appropriate volumes and sterilized by autoclaving.
Inoculation and experimental groups
The Bacillus subtilis liquid culture was inoculated into the inoculation and precipitation media according to the experimental design summarized in Table 1. Experimental groups were categorized as follows: bacteria-free groups (Groups 1, 2, 8, and 9), live-bacteria groups (Groups 3, 4, 10, and 11), and dead-bacteria groups (Groups 5, 6, 12, and 13). Bacteria-free groups served as negative controls, whereas dead-bacteria groups were included to evaluate passive, non-metabolic contributions to calcium carbonate precipitation. Prior to inoculation, bacterial suspensions were adjusted to a final concentration of 10⁷ CFU/mL. Cultivation was performed in either small or large glass jars, depending on the marble sample size.Table 1. Composition of media and experimental grouping used in biocalcification assaysCO_2_ConditionInoculation MediumBacteria ApplicationPrecipitation MediaGroup #CO_2_ pre-treatedYEU^a^NoneCaAcetatGYE^b^ +U^c^1CaCl_2_ + U2LiveCaAcetatGYE + U3CaCl_2_ + U4DeadCaAcetatGYE + U5CaCl_2_ + U6**—None—7 (Control)CO_2_ untreatedYEUNoneCaAcetatGYE + U8CaCl_2_ + U9LiveCaAcetatGYE + U10CaCl_2_ + U11DeadCaAcetatGYE + U12CaCl_2_ + U13—None—14 (Control)^a^YEU: Yeast extract-urea medium^b^CaAcetatGYE: Medium containing Ca(C_2_H_3_O_2_)2, glucose and yeast extract^c^ U: UreaNone: Bacteria-free“—**“: No inoculation or precipitation medium applied
Bacterial coating applications on marbles
For bacterial coating, the inoculation media in large and small jars were inoculated with 100 µL and 10 µL of bacterial suspension, respectively. Inoculation was applied to Groups 3–6 and 10–13, as outlined in Table 1. The jars were covered with sterile sponge plugs to allow gas exchange and incubated at 37 °C with shaking at 90 rpm for 24 h.
Following this incubation period, cultures designated for dead-bacteria treatments were removed from the incubator and autoclaved at 121 °C for 20 min, with jar lids securely closed to prevent liquid loss. Groups without bacterial inoculation were maintained in the dark at room temperature for 24 h under identical conditions.
After preparation of the inoculation media, marble samples were fully immersed to ensure contact of all surfaces with the culture medium. The jars were again covered with sterile sponge plugs, and the marble samples were incubated statically at 28 °C for 2 days to allow bacterial attachment and surface interaction.
Subsequently, the marble samples were aseptically transferred to the corresponding precipitation media. The samples were incubated statically at 28 °C for an additional 4 days, allowing calcium carbonate precipitation to occur on the marble surfaces. After completion of the precipitation stage, the marble samples were incubated at 30 °C for 2 days and then stored at 24 °C under air-conditioned laboratory conditions for 3 months to allow crystal maturation and stabilization. Marble samples subjected to CO₂ pre-treatment were processed using the same post-precipitation incubation protocol. All control groups were maintained under identical conditions throughout the experiment.
Instrumental analysis
X-ray diffraction (XRD)
The crystal structure of the identification powdered marble samples were analyzed by XRD. Rigaku Rad B-Dmax II powder X-ray diffraction meter was used. Changed the value of 2θ by 20° to 80° with 0.04° step size using Cu Kα radiation. 5 mg samples were on a coverslip with silica grease.
Scanning electron microscopy (SEM) combined with energy dispersive x-ray analysis (EDX) (SEM-EDX)
The surface morphology of the marble samples was displayed using SEM (LEO Evo-40 VPX microscope). Chemical composition analysis was performed with EDX, using Rönteck xflash detector analyzer associated with Scanning electron microscope (SEM, Leo-Evo 40xVP).
Atomic force microscopy (AFM)
Atomic force microscopy (AFM; XE-100E, Park Systems Corp., Suwon, Korea) was employed to quantitatively evaluate surface topography and pore/crack geometry of the marble samples. The maximum pore (or crack) depth was defined as the vertical height difference between the bottom of surface pores or cracks and the adjacent intact marble surface. This parameter was determined from AFM height images using line profile analysis across representative pores and cracks. Measurements were performed at four different regions per sample, and results are reported as mean ± standard deviation. Although AFM line profile analysis involves measuring vertical height differences, the measurements in this study were performed specifically across individual pores and cracks to determine their depth, rather than using peak-to-valley values to describe overall surface roughness.
Pore size was defined as the lateral pore dimension measured from stereomicroscopic and SEM images, and pores were classified as micropores or macropores based on their measured size ranges.
Results
Macroscopic and stereomicroscopic imaging results
Following the completion of a three-month incubation period, macroscopic examination of the surfaces of the 4 × 4 cm marble samples revealed the presence of white, fine granular deposits localized within the inner regions of the macropores in samples from the CO₂-pre-treated group containing live bacteria (Group 4). These deposits appeared as discrete accumulations within pore structures rather than as continuous surface layers.
Stereomicroscopic observations further confirmed the presence of calcium carbonate deposits in Group 4 samples. In contrast, no CaCO₃ crystal formation was detected on the marble surfaces of the CO₂-pre-treated control group (Group 7), the CO₂-untreated control group (Group 14), or the bacteria-free groups (Groups 1, 2, 8, and 9). Similarly, marble samples that were not subjected to CO₂ pre-treatment did not exhibit detectable CaCO₃ deposition, regardless of whether live or dead bacteria were present.
As shown in Fig. 2b and c, the macropores of marble samples treated with live bacteria in calcium chloride medium exhibited partial closure, whereas the micropores were largely filled as a result of CaCO₃ precipitation. Microscopic observations revealed that the newly formed mineral structures were clearly distinguishable from the original marble matrix and were associated with pores exhibiting pore sizes in the range of approximately 3–5 mm. Calcium carbonate deposition on the marble surfaces was more pronounced in the calcium chloride–containing medium than in the calcium acetate medium. In addition, the morphology of the precipitated CaCO₃ differed between the two media: in the CaCl₂·2 H₂O medium, granular white crystalline deposits were observed (Fig. 3), whereas in the Ca(CH₃COO)₂-containing medium, the deposits appeared as fine, powdery layers.Fig. 2. CaCO₃ crystals formed on CO₂-pre-treated marble surfaces in CaCl₂+U medium (10× magnification): (a) control group (group 7); (b) live-bacteria-treated group (group 4); (c) dead-bacteria-treated group (group 6)Fig. 3. Stereo microscopic images of CaCO₃ crystals formed by live bacteria on CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (group 4) at 10x and 15x magnification
Figure 2 CaCO₃ crystals formed on CO₂-pre-treated marble surfaces in CaCl₂+U medium (10× magnification): (a) control group (Group 7); (b) live-bacteria-treated group (Group 4); (c) dead-bacteria-treated group (Group 6).
Microscopic examination showed that marble samples treated with live bacteria in calcium chloride medium developed less complex surface structures than those observed in CO₂-pre-treated samples. Stereomicroscopic observations revealed no detectable crystal formation in samples containing dead bacteria or in samples that were not subjected to CO₂ pre-treatment. In contrast, SEM analysis revealed the presence of CaCO₃ crystal structures in both live- and dead-bacteria-treated samples, although crystal abundance and surface coverage were markedly lower in the CO₂-untreated groups. Furthermore, surface coverage and macropore filling were less extensive in the CO₂-untreated samples than in the CO₂-pre-treated samples.
Microscopic examination showed that the surface structures formed on marble samples treated with live bacteria in a calcium chloride medium were less complex than those observed in comparable samples subjected to CO₂ pre-treatment. Stereomicroscopic observations revealed no detectable crystal formation in samples containing dead bacteria or in samples that were not subjected to CO₂ pre-treatment. In contrast, SEM analysis revealed the presence of CaCO₃ crystal structures in both live- and dead-bacteria-treated samples, although crystal abundance and surface coverage were markedly lower in the CO₂-untreated groups. Furthermore, surface coverage and macropore filling were less extensive in the CO₂-untreated samples than in the CO₂-pre-treated samples.
Effects of Live/Dead Cells, culture Media, and CO2 Pre-treatment on biocalcification efficiency
Comparative evaluation of the marble samples demonstrated that biocalcification efficiency was strongly influenced by bacterial viability (live versus dead cells), the type of culture medium, and CO₂ pre-treatment. These factors governed not only the extent of CaCO₃ deposition but also the resulting crystal polymorphs (calcite, aragonite, and vaterite) and the spatial distribution of precipitates within marble pores. SEM observations revealed distinct CaCO₃ crystal structures within the micropores of treated marble samples, which were distinguishable from the original marble matrix based on their morphology and size. In contrast, the marble surfaces of the control group retained their original amorphous appearance, with no evidence of crystal formation (Fig. 4).Fig. 4SEM images of CO₂-pre-treated marble surfaces from the control group and CaCO₃ crystals formed by live bacteria on CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (group 4)
These findings were further supported by SEM imaging, which provided clear evidence of bacterial calcification. SEM images revealed calcite and aragonite crystals with sizes ranging from approximately 20 to 100 μm on the marble surfaces and within micropores. The biogenic CaCO₃ deposits partially filled pore spaces and were distinguishable from the amorphous surface structures observed in the control group. Among all experimental groups, Group 4 exhibited the most extensive biocalcification, characterized by the largest and densest crystal formations (Fig. 5).Fig. 5SEM images of CaCO₃ crystals formed by live bacteria on CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (group 4). C, calcite; A, aragonite
Beyond morphological observations, SEM-EDX and elemental mapping analyses provided chemical support for bacterial calcification. Figures 6 and 7 present SEM-EDX spectra and elemental mapping analyses of marble surfaces from the CO₂-pre-treated control group and Group 4. EDX analysis showed that the carbon content of CO₂-pre-treated marble samples was higher than that of untreated control samples. Furthermore, SEM-EDX results indicated that the newly formed structures associated with bacterial calcification exhibited elevated Ca, C, and O signals compared to the control group, consistent with the presence of calcium carbonate (Fig. 7).Fig. 6SEM-EDX and elemental mapping of CO₂-pre-treated marble surfaces from the control group (group 7)Fig. 7SEM-EDX and elemental mapping of CaCO₃ crystals formed by live bacteria on CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (Group 4)
In addition, EDX spectra confirmed the elemental composition of the marble, showing prominent signals corresponding to calcium (Ca), carbon (C), silicon (Si), and oxygen (O). Compared to the control group, bacteria-treated and nutrient-supplemented samples exhibited higher relative Ca signals, while O and Si signals were comparatively reduced.
Quantitative EDX analysis revealed that marble samples treated with live bacteria in CaU medium (Group 4) displayed the highest calcium content (48.88%; Fig. 7), whereas the control group lacking both bacterial activity and nutrient supplementation exhibited a lower calcium content (34.95%; Fig. 6). These compositional differences are consistent with enhanced calcium carbonate deposition in the biocalcified samples.
The production of calcium carbonate crystals by living bacteria was supported by the presence of characteristic elemental peaks detected in the EDX spectra. Additional evidence was obtained from analyses of material scraped from distinct crystalline structures observed within the macropores of the 4 × 4 cm marble surfaces in Group 4. SEM-EDX analysis of these scraped samples confirmed their calcium carbonate composition (Fig. 8).Fig. 8SEM images of CaCO₃ crystals scraped from macropores of CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (group 4, live bacteria) at 5000x magnification. C, calcite; A, aragonite
In addition, SEM-EDX and elemental mapping analyses were performed on the scraped samples. These analyses identified the presence of calcite and aragonite crystal phases within the sampled material. Quantitative EDX analysis indicated a calcium content of 43.46% in these samples (Fig. 9). The corresponding EDX spectra displayed characteristic elemental peaks consistent with calcium carbonate composition.Fig. 9SEM-EDX and elemental mapping of CaCO₃ crystals scraped from macropores of CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (Group 4)
Following the analysis of live-cell treatments, the effects of dead bacterial cells on biocalcification were also examined. SEM analysis of CO₂-pre-treated marble surfaces treated with dead bacteria and immersed in CaCl₂+U medium (Group 6) revealed the presence of calcite, aragonite, and amorphous phases (Fig. 10). Calcium carbonate crystals were observed on the marble surfaces in the dead-cell-treated samples; however, crystal density was markedly lower than that observed in samples treated with live cells under the same conditions. Consistent with these observations, SEM-EDX analysis showed that the Ca content in dead-cell-treated groups was higher than in the control group but lower than in the corresponding live-cell-treated groups (data not shown).Fig. 10SEM images of CaCO₃ crystals formed on CO₂-pre-treated marble surfaces treated with dead bacteria in CaCl₂+U medium (Group 6)
Given the influence of nutrient composition and calcium source on biocalcification, an alternative medium (CaAcetatGYE + U) was also evaluated. In contrast to the more pronounced calcification observed in the CaCl₂+U medium, Groups 3 and 5 maintained in CaAcetatGYE + U exhibited sparse, low-density aragonite and vaterite deposits in SEM analyses. These deposits were limited in extent, suggesting that micropore filling was less developed under these conditions (Fig. 11).Fig. 11SEM images of CaCO₃ crystals formed on marble surfaces treated with live (group 3, top) and dead (group 5, bottom) bacteria and incubated in CaAcetate-GYE + U medium. V, vaterite
To assess the effect of CO₂ pre-treatment on biocalcification, CO₂-untreated marble samples were also examined. Macroscopic, stereomicroscopic, and SEM analyses of CO₂-untreated samples inoculated with live bacteria in CaCl₂+U medium revealed no detectable CaCO₃ polymorphs. In contrast, SEM images of CO₂-untreated samples treated with dead bacteria showed the presence of sparse, needle-like aragonite structures with average lengths of approximately 20 μm, although their density was lower than that observed in live-bacteria-treated samples under CO₂-pre-treated conditions (Fig. 12).Fig. 12SEM images of marble surfaces from the CO₂-untreated control group and CaCO₃ crystals formed on CO₂-untreated marble surfaces treated with dead bacteria and incubated in CaCl₂+U medium (group 13). C, calcite; A, aragonite
SEM-EDX analysis further indicated lower carbon levels in the CO₂-untreated control samples compared to the CO₂-pre-treated controls (data not shown). Similarly, no CaCO₃ crystals were detected in CO₂-untreated samples maintained in CaAcetateGYE medium with live bacteria; instead, predominantly amorphous surface structures resembling the native marble matrix were observed. Overall, these observations indicate that CO₂ pre-treatment is associated with enhanced CaCO₃ formation and more pronounced differences between live- and dead-bacteria-treated samples.
SEM analysis of CO₂-untreated marble surfaces treated with dead bacteria revealed the presence of vaterite as the dominant calcium carbonate polymorph (Fig. 13). In contrast, SEM examination of CO₂-untreated and bacteria-free marble surfaces maintained in CaCl₂+U medium showed only limited crystal formation, with the detected CaCO₃ polymorphs predominantly corresponding to aragonite (Fig. 14).Fig. 13SEM image of CO₂-untreated marble surfaces treated with dead bacteria in CaAcetate-GYE + U medium (group 12)Fig. 14SEM image of CO₂-untreated, bacteria-free marble surfaces incubated in CaCl₂+U medium (group 9)
As summarized in Table 2, the combination of CO₂ pre-treatment and live bacterial application was associated with the most extensive biohealing features observed on marble surfaces. In this group, crystal sizes ranged from approximately 20 to 100 μm and consisted predominantly of calcite and aragonite. The affected regions were widespread, with both micro- and macropores partially or fully filled, as reflected by measured pore diameters of 0.214 mm and 0.059 mm, pore areas of 0.033 × 0.062 mm² and 0.031 × 0.066 mm², and macropores of approximately 3 × 5 mm exhibiting CaCO₃ sealing.Table 2. Summary of crystal size, polymorph type, and biohealed area characteristics observed on marble surfaces under different bacterial, calcium source, and CO₂ pre-treatment conditionsCO₂ ApplicationCalcium SourceBacterial FormCrystal Size (µm)Crystal TypeBiohealed Area SizePresentCaCl₂+ULive20–100Calcite, Aragonite0.214 mm (diameter); 0.059 mm (diameter); 0.033 × 0.062 mm²; 0.031 × 0.066 mm²; macropores ~ 3 × 5 mmPresentCaCl₂+UDead10–20Calcite, Aragonite0.036 mm (diameter)PresentCaAcetateGYELive1–10Vaterite—PresentCaAcetateGYEDead1–10Vaterite—AbsentCaCl₂+ULive10–20Aragonite—AbsentCaCl₂+UDead10–20Calcite, Aragonite—AbsentCaAcetateGYELive———AbsentCaAcetateGYEDead1–10Vaterite—“—” indicates that no measurable biohealed area was detected under the corresponding experimental conditions. Biohealed area size refers to the dimensions of CaCO₃-filled regions determined from stereomicroscopic and SEM observations.
In contrast, CO₂-untreated samples treated with live bacteria exhibited only smaller aragonite crystals (10–20 μm), and no measurable biohealed areas were detected. Samples treated with dead bacteria showed limited calcification. Under CO₂-pre-treated conditions, crystals with sizes of 10–20 μm (calcite and aragonite) were observed, whereas the biohealed area was restricted to a diameter of approximately 0.036 mm. In the absence of CO₂ pre-treatment, crystal sizes were further reduced (1–10 μm) and were predominantly composed of vaterite. Overall, the data in Table 2 illustrate clear differences in crystal size, polymorph distribution, and pore filling across the experimental groups.
XRD analysis results
The XRD diffractogram presented in Fig. 15 corresponds to calcium carbonate crystals extracted from the macropores of CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (Group 4). The diffraction pattern was consistent with well-crystallized calcite (CaCO₃), as evidenced by the dominant reflection at approximately 29.4° 2θ corresponding to the (104) plane. Additional characteristic calcite reflections were observed at 23.1°, 36.0°, 39.4°, 43.1°, 47.5–48.5°, 57.5°, 60.7–61.0°, and 65.3° 2θ, in good agreement with standard calcite reference data (Table 3). Calcite was identified as the dominant crystalline phase in the XRD patterns.Fig. 15X-ray diffraction (XRD) pattern of calcium carbonate crystals extracted from the macropores of CO₂-pre-treated marble surfaces incubated in CaCl₂+U medium (group 4)Table 3XRD peak positions, d-spacings, relative intensities, and crystallographic plane assignments for calcite identified in the diffractogram shown in Fig. 15Peak2θ (°)d (Å)Relative intensityPhase(hkl)123.13.8520–25Calcite(012)229.43.04100Calcite(104)331.42.8515–20Calcite(006)436.02.4920–25Calcite(110)539.42.2835–45Calcite(113)643.12.1020–30Calcite(202)747.51.9115–25Calcite(018)848.51.8820–30Calcite(116)957.51.6015–20Calcite(122)1060.71.5310–15Calcite(214)1161.01.528–12Calcite(300)1265.31.4310–15Calcite(125)
AFM analysis results
AFM analysis revealed distinct differences in surface topography and pore/crack geometry between control and biocalcified marble samples. The maximum pore (or crack) depth, determined from AFM height images using line profile analysis, decreased from 35.00 ± 7.07 μm in the control group to 22.50 ± 8.20 μm in biocalcified samples (Fig. 16). This reduction reflects partial filling of surface pores and cracks and a smoother surface morphology associated with CaCO₃ deposition induced by microbial activity.Fig. 16AFM images of CO₂-pre-treated marble surfaces from the control group (group 7, top) and marble surfaces treated with live bacteria (group 4, bottom). Representative line profile analyses were used to determine the maximum pore/crack depth, defined as the vertical height difference between the bottom of surface pores or cracks and the adjacent intact marble surface
The effect of CO₂ pre-treatment on marble surface chemistry was further supported by microstructural and compositional analyses. SEM-EDX and elemental mapping (Figs. 6 and 7) revealed an increased carbon signal on CO₂-pre-treated marble surfaces compared to untreated controls, indicating enhanced surface carbon availability following CO₂ exposure under humid conditions. This chemical preconditioning promoted Ca²⁺–CO₃²⁻ supersaturation at the surface, facilitating nucleation and growth of CaCO₃ crystals during microbial activity. Consistently, XRD analysis of crystals extracted from CO₂-pre-treated samples (Fig. 15) confirmed the formation of well-crystallized calcite, demonstrating that CO₂ pre-treatment played a critical role in stabilizing carbonate mineral phases and enhancing biocalcification efficiency.
Discussion
The present study shows that microbially induced carbonate precipitation (MICP) mediated by Bacillus subtilis ATCC 6633 can contribute to the biohealing of marble surfaces by promoting calcium carbonate deposition within surface pores and cracks. The results indicate that bacterial viability and calcium source play key roles in influencing CaCO₃ polymorphism and surface modification. Live bacterial cells favored the formation of thermodynamically stable calcite and aragonite phases, whereas dead cells and calcium acetate predominantly resulted in metastable vaterite formation, suggesting different nucleation pathways under biologically active and passive conditions. Furthermore, CO₂ pre-treatment acted as a surface-conditioning step that enhanced inorganic carbon availability and promoted more uniform CaCO₃ nucleation, thereby improving biocalcification efficiency. Taken together, these findings support a mechanistic interpretation linking microbial activity, calcium chemistry, and surface preconditioning to marble biohealing. These observations are consistent with established mechanisms governing microbially induced carbonate precipitation and CaCO₃ polymorphism.
Microbially induced carbonate precipitation (MICP) is a mineralization process driven by the metabolic activities of microorganisms. Ureolytic bacterial strains hydrolyze urea via the enzyme urease, generating carbonate species that subsequently react with calcium ions to form calcium carbonate. In addition to urease activity, enzymes such as carbonic anhydrase may also contribute to carbonate generation, and the overall precipitation process is governed by a series of interconnected biochemical reactions (Achal and Pan 2014; Alvarado-Mata et al. 2025). Briefly, urea hydrolysis produces ammonia and carbamate, which further decompose into ammonia and carbonic acid. These products interact in aqueous solution to yield ammonium, hydroxide, and bicarbonate ions, leading to an increase in pH. As a result, bicarbonate ions are converted into carbonate ions, enabling CaCO₃ precipitation in the presence of calcium ions (Hammes et al. 2003).
Bacterial calcium carbonate precipitation can result in the formation of different crystal polymorphs, most commonly calcite, aragonite, and vaterite. Among these, vaterite is a metastable phase that is frequently produced during the early stages of bacterial mineralization and may subsequently transform into the more thermodynamically stable calcite form (Tourney and Ngwenya 2009; Wang and Becker 2009; Bozbeyoğlu et al. 2020; Qafoku et al. 2022). Stereomicroscopic observations of marble macropores at 20× magnification revealed CaCO₃ structures consistent with those reported in previous studies (Wiktor and Jonkers 2011). Correspondingly, SEM analyses confirmed the presence of crystalline deposits whose morphology was comparable to that of CaCO₃ crystals described in the literature. These biogenic mineral phases were clearly distinguishable from the amorphous marble matrix based on their size, morphology, and visual appearance. In contrast, control marble surfaces exhibited no crystalline deposits and were consistent with previously reported observations for untreated marble materials (Colao et al. 2004; Moropoulou et al. 2019).
B. subtilis has been reported to promote calcite formation in vitro when cultured in media containing different calcium sources, including calcium lactate, calcium acetate, and calcium chloride (Shirakawa et al. 2015). Several Bacillus strains are known to mineralize CaCO₃ into distinct polymorphs, most commonly calcite and vaterite (Seifan et al. 2016), although the factors governing polymorph selection remain incompletely understood. In the present study, in addition to calcite, a limited number of aragonite crystals were also identified. Comparable observations have been reported for Sporosarcina pasteurii, in which calcite formation was dominant in calcium chloride media, whereas SEM analyses revealed vaterite structures in media containing calcium lactate, calcium acetate, or calcium gluconate (Gorospe et al. 2013). Polymorph transitions are influenced by incubation conditions such as temperature and time, and less stable phases such as aragonite may nucleate during early stages of mineralization before gradually transforming into the thermodynamically stable calcite phase (Ramakrishna et al. 2016).
Microorganisms possessing negatively charged cell surfaces and urease activity can hydrolyze urea to generate carbonate species and ammonium, thereby facilitating the binding of calcium ions to the cell surface and inducing CaCO₃ precipitation (De Muynck et al. 2010). Under conditions of elevated urea availability, this process is further enhanced, which explains why ureolytic bacteria are commonly selected for MICP applications due to their robustness and adaptability to environmental conditions. Calcium carbonate precipitation typically proceeds through heterogeneous nucleation on the bacterial cell wall once supersaturation is achieved (Siddique and Chahal 2011). For extracellular CaCO₃ precipitation to occur via urease- and carbonic anhydrase–mediated reactions, calcium ions must be present at sufficient concentrations. Consequently, both the type and concentration of the calcium source represent critical factors governing the extent and polymorphic outcome of mineralization (Kart 2021). In agreement with this, the present study showed that different calcium sources resulted in distinct CaCO₃ polymorphs, with calcium chloride favoring the formation of calcite and aragonite, whereas calcium acetate predominantly promoted vaterite formation.
Vaterite nanoparticles have been reported to exhibit advantageous properties as filler materials, including enhanced hydration kinetics, improved pore filling, and increased mechanical performance in cement-based systems (Li et al. 2024). Similarly, Hargis et al. (2014) demonstrated that vaterite reduced setting time compared to calcite, despite differences in particle size. Although vaterite is a metastable and relatively uncommon CaCO₃ polymorph, it can be induced under specific conditions, including biocalcification processes (Mann 2001). The morphology of vaterite is strongly influenced by the calcium source, with calcium acetate promoting lettuce-like lamellar structures, whereas calcium lactate and calcium gluconate favor spherical morphologies (Tai and Chen 1998). In this context, the present study highlights the formation of spherical vaterite associated with dead B. subtilis cells under calcium-rich conditions, which differs from morphologies commonly reported in the existing literature.
XRD, SEM, and FTIR analyses of Bacillus sp. CR2 cultures revealed that calcite was the dominant CaCO₃ polymorph, irrespective of whether calcium chloride, calcium oxide, calcium acetate, or calcium nitrate was used as the calcium source. Notably, in calcium acetate–containing media, minor amounts of vaterite and aragonite were also detected (Achal and Pan 2014). Consistent with these observations, a parallel study on biogenic plaster samples prepared using Sporosarcina pasteurii ATCC 6453 demonstrated that calcium nitrate– and calcium chloride–treated systems predominantly yielded calcite, whereas calcium acetate–based systems additionally exhibited aragonite and vaterite phases (Zhang et al. 2014).
Bacterial carbonate precipitation can occur through both passive and active nucleation (Schultze-Lam and Beveridge 1994; Schultze-Lam et al. 1996; Stocks-Fischer et al. 1999; Warren et al. 2001; Raza and Khushnood 2022). Passive nucleation is primarily driven by metabolically induced changes in the microenvironment surrounding bacterial cells and in the chemistry of the bulk solution, which together increase mineral supersaturation and promote carbonate precipitation (Schultze-Lam et al. 1996). A common indicator of this process is an increase in pH, often resulting from the production of CO₃²⁻ or HCO₃⁻ via bacterial ammonification (Stocks-Fischer et al. 1999) or from sulfate reduction under anoxic conditions (Van Lith et al. 2003). In addition, alkalization may also arise from photosynthetic activity, in which bacteria consume HCO₃⁻ and release OH⁻, further promoting carbonate precipitation (Ferris et al. 1997). In contrast, active nucleation occurs when the negatively charged functional groups on bacterial cell walls or within extracellular polymeric substances (EPS) directly bind calcium ions, providing nucleation sites for CaCO₃ crystal growth (Schultze-Lam and Beveridge 1994; Warren et al. 2001; Hoffmann et al. 2021). This mechanism reflects the direct involvement of microbial cell surfaces in mineral precipitation and complements the indirect biochemical changes associated with passive nucleation. In the present study, SEM-EDX and microscopic observations indicate surface-associated CaCO₃ formation, suggesting that such cell-mediated interactions may contribute to the observed mineralization alongside solution-driven processes.
According to EDX analysis, a higher carbon content was detected on the surface of marble samples from the CO₂-pre-treated control group. This observation suggests that pre-incubation of marble surfaces under CO₂ conditions increases surface-associated inorganic carbon, which may facilitate subsequent CaCO₃ nucleation. This interpretation is consistent with previous studies reporting that CO₂ exposure can modify surface chemistry and influence carbonate precipitation processes (Shirakawa et al. 2015). Similarly, Isar (2020) reported that biocalcite formation is associated with the presence of Ca, O, C, and N in the surrounding environment, while Polat (2019) identified CaCO₃ formation based on elemental compositions including C, O, Na, Al, Si, K, Ca, and Ti.
Crystallization observed on surfaces associated with dead bacterial cells can be attributed to surface-mediated active nucleation mechanisms. The electronegative charges present on bacterial cell surfaces act as effective substrates for nucleation (Warren et al. 2001), although the underlying process differs from CaCO₃ precipitation driven by HCO₃⁻ production in metabolically active cells. In this context, chemically heterogeneous macromolecules on bacterial surfaces generate net negative charges that adsorb Ca²⁺ ions and subsequently attract CO₃²⁻ or HCO₃⁻ ions, thereby initiating CaCO₃ nucleation (Stocks-Fischer et al. 1999; Warren et al. 2001; Hoffmann et al. 2021). This surface-associated pathway is widely recognized as active nucleation (Van Lith et al. 2003). Furthermore, carboxyl groups present on the surfaces of both Gram-positive and Gram-negative bacteria exhibit spatial configurations homologous to those of calcite lattice planes and carbonate ions, facilitating active calcite formation (Braissant et al. 2003). Recent nanoscale studies have further demonstrated that substrate mineralogy, biofilms, and extracellular polymeric substances (EPS) strongly influence bacterially induced carbonate mineralization, highlighting EPS as an additional nucleation matrix beyond cell wall functional groups (Dubey et al. 2025). In contrast, crystallization observed on surfaces completely devoid of bacterial cells is more appropriately explained by abiogenic calcification or by organic matrix-mediated nucleation processes rather than cell-driven active nucleation.
These findings suggest that CO₂ pre-treatment functions as an effective surface-conditioning step by increasing inorganic carbon availability at the marble interface, which may promote local Ca²⁺–CO₃²⁻ supersaturation and facilitate stable CaCO₃ nucleation during microbial activity. This interpretation is consistent with previous studies demonstrating that shifts in carbonate equilibria strongly influence CaCO₃ polymorphism and phase stability. Moreover, the distinct CaCO₃ polymorphs observed in samples treated with live and dead bacterial cells can be attributed to differences in local supersaturation conditions and nucleation pathways, as variations in surface chemistry are known to govern the preferential formation of calcite, aragonite, or vaterite (Liendo et al. 2022).
Overall, the findings of this study indicate that microbially induced carbonate precipitation mediated by B. subtilis ATCC 6633 represents a promising approach for marble biohealing. By modulating bacterial viability, calcium source, and CO₂ pre-treatment conditions, CaCO₃ polymorphism and pore-filling behavior can be influenced, providing valuable insights for future applications in material conservation and durability enhancement; however, further studies are required to assess the long-term durability of the biocalcified layers under outdoor environmental conditions.
Conclusion
This study demonstrates that B. subtilis is capable of inducing effective biocalcification on marble surfaces, resulting in partial filling of both micro- (0.02–0.03 mm) and macro-pores (3–5 mm). The combination of live bacterial cells with CO₂ pre-treatment yielded the most extensive and stable CaCO₃ deposition, predominantly in the form of calcite and aragonite, whereas dead bacterial cells and calcium acetate favored the formation of less stable vaterite. These findings underscore the critical roles of bacterial viability and calcium source in governing CaCO₃ polymorphism and overall biohealing efficiency.
Beyond its mechanistic insights, this eco-friendly and sustainable approach offers a promising alternative for marble conservation and related biomineralization-based applications. By enabling the filling of natural pores and cracks, including regions that are difficult to access using conventional restoration techniques, microbially induced carbonate precipitation (MICP) represents a viable strategy for enhancing the durability of marble surfaces. Moreover, this work contributes to a broader understanding of microbial roles in carbonate mineralization and supports the development of low-impact conservation technologies aligned with contemporary sustainability and reduced carbon-emission goals.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kart NNB (2021) Interaction of ureolytic bacteria and clay in bacterial calcium carbonate mineralization: Paenibacillus favisporus U 3 (Doctoral dissertation). Pamukkale University, Institute of Science, Denizli
- 2Su Y, Liu C, Fang H, Zhang D (2020) Bacillus subtilis: a universal cell factory for industry, agriculture, biomaterials and medicine. Microb Cell Fab 19:173