Structural and Functional Analysis of Porcine CR1-like Proteins in C4b-Mediated Immune Responses
Wei Yin, Nan Wang, Jingze Li, Haoxiang Yao, Qiongyu Li, Hongquan Li, Kuohai Fan, Jia Zhong, Zhenbiao Zhang, Na Sun, Panpan Sun, Huizhen Yang, Jianzhong Wang, Yaogui Sun

TL;DR
This study explores how porcine CR1-like proteins interact with C4b to help the immune system respond to PRRSV, a major swine disease.
Contribution
The study identifies specific CR1-like fragments and key amino acids involved in C4b binding, offering new insights into porcine immune mechanisms.
Findings
PRRSV triggers C4 cleavage, and C4b facilitates adhesion of porcine red blood cells to the virus.
Three CR1-like fragments bind C4b, with 18 key amino acids identified through structural simulations.
The 118I mutation reduces the affinity between CR1-like CCPs 1-3 and C4b, as confirmed by SPR.
Abstract
The complement system is crucial for innate immunity, but its role against Porcine Reproductive and Respiratory Syndrome Virus (PRRSV), a significant swine pathogen, is not well understood. This study investigated the interaction between porcine CR1-like protein and C4b in PRRSV immunity. ELISA showed PRRSV triggers C4 cleavage, and immunoelectron microscopy indicated C4b facilitates porcine red blood cell adhesion to PRRSV. Yeast two-hybrid assays identified three CR1-like fragments binding C4b, structural simulations pinpointed 18 key amino acids, and SPR confirmed that the 118I mutation reduced affinity between CCPs 1-3 and C4b. These findings reveal that porcine CR1-like protein aids immune adhesion to PRRSV through C4b binding, clarify the structural basis, improve understanding of porcine complement defense, and lay the groundwork for PRRSV treatments. The complement system is…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
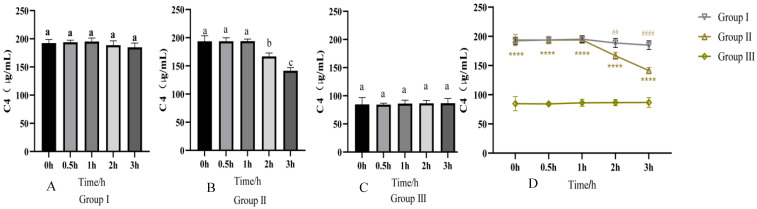 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3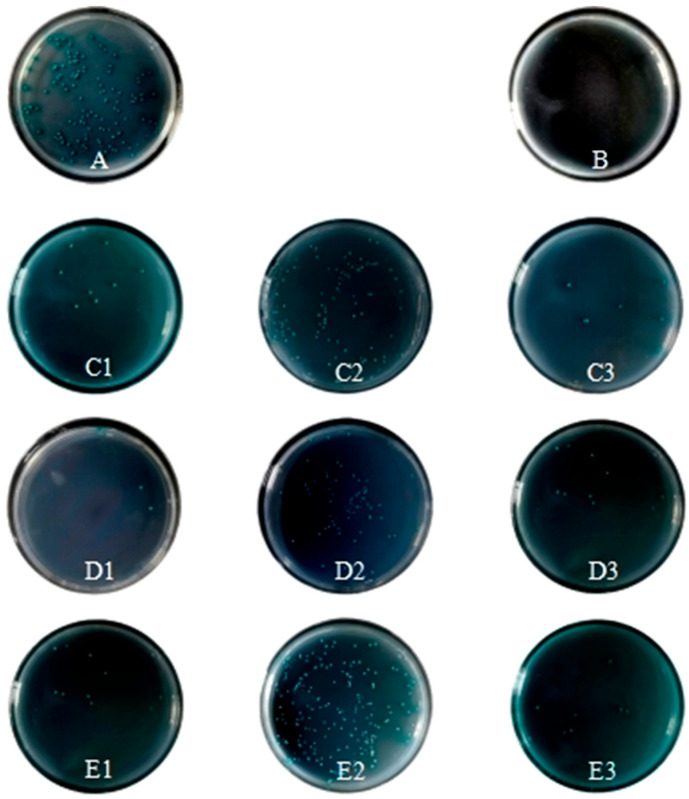 Figure 4
Figure 4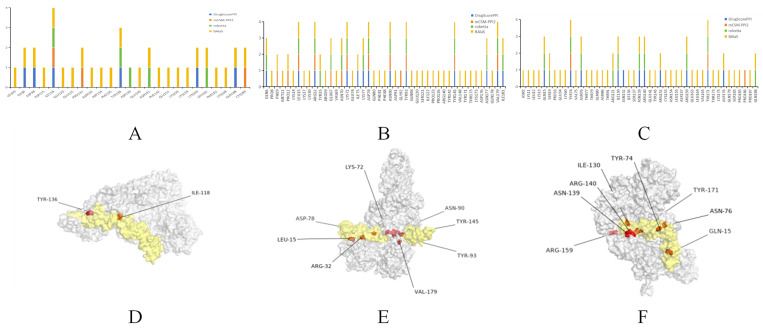 Figure 5
Figure 5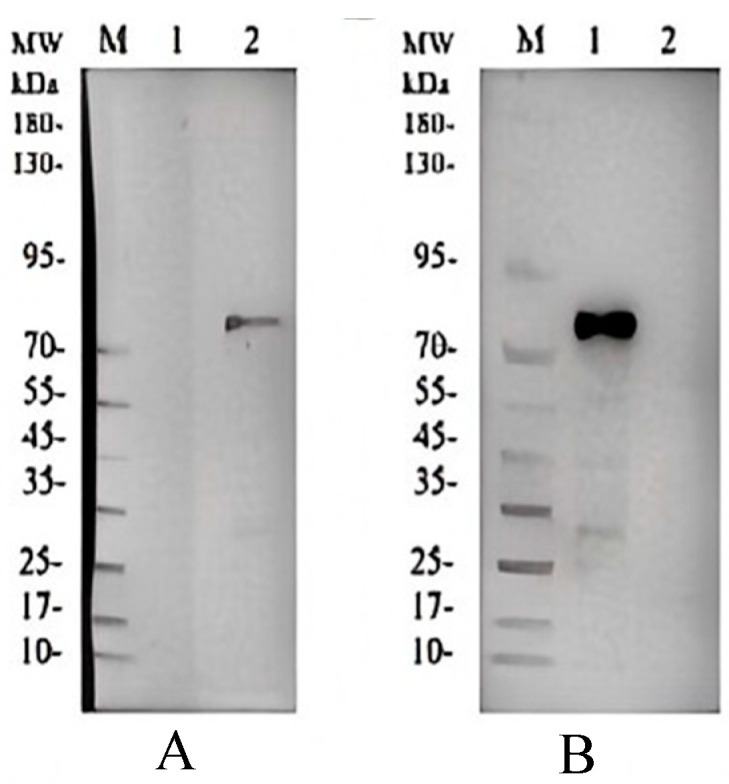 Figure 6
Figure 6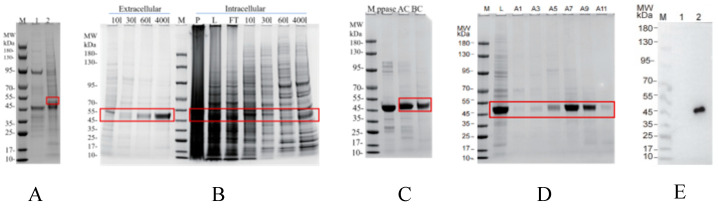 Figure 7
Figure 7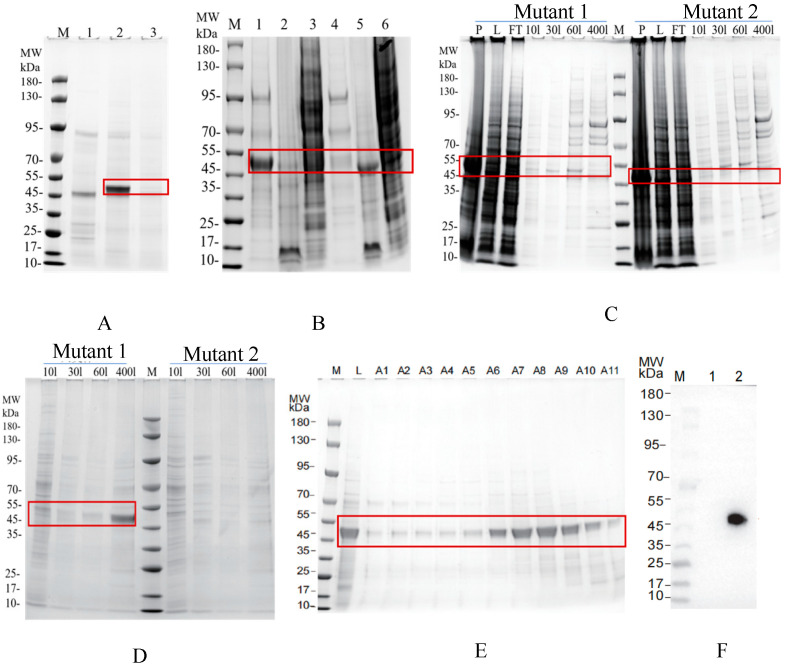 Figure 8
Figure 8 Figure 9
Figure 9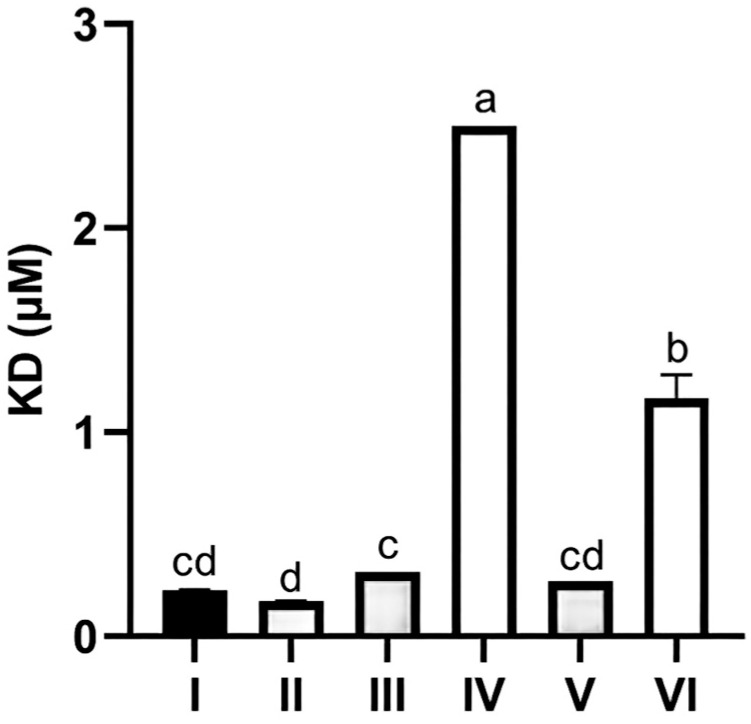 Figure 10
Figure 10- —Shanxi Natural Science Research Project
- —Shanxi Key Laboratory of Modernization of Traditional Chinese Veterinary Medicine
- —Shanxi Agricultural University Doctoral Research Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsComplement system in diseases · Xenotransplantation and immune response · Monoclonal and Polyclonal Antibodies Research
1. Introduction
The complement system constitutes a fundamental element of innate immunity, playing a crucial role in combating microbial infections, eliminating foreign pathogens, and preserving tissue homeostasis. Activation of the complement system induces an upregulation of cytokines, chemokines, and other molecules integral to the innate immune response [1,2]. The complement component C4, with a molecular weight of 204 kDa, is a crucial part of the complement system and is essential in the classical and lectin pathway cascades. It is a three-chain glycoprotein linked by disulfide bonds, consisting of α chains (95 kDa), β chains (75 kDa), and γ chains (30 kDa) [3,4]. C4b is a product of C4 cleavage and is produced by activation of the C1q, C1r, and C1s complex (classical pathway) and mannan-binding lectin (MBL) [5,6]. Through its reactive thioester, C4b binds to the target surface and can be transformed into the intermediate iC4b form, undergoing proteolysis by serine protease factor I and cofactor CD46 [7,8]. Further cleavage of iC4b yields thioester-linked C4d (45 kDa) and soluble C4c (146 kDa), which can act as biomarkers for complement activation in the classical and lectin pathways. The risk of infections and autoimmune diseases can rise with a complete or partial lack of complement C4. Many studies have indicated that complement C4 is essential for protection against microbial infections [9]. Increased vulnerability to infection is associated with a full or partial deficiency of complement C4 [10]. Additionally, an insufficiency of complement C4 might cause several autoimmune disorders [11]. Decreased C4 protein concentration and decreased serum complement activity are common in systemic lupus erythematosus (SLE) and other active infectious diseases. The change in C4 content in plasma can also be used as an indicator to monitor various diseases [12,13,14,15]. On the surface of Group B Streptococcus (GBS), the complement-interfering protein (CIP) enhances cell adhesion and penetration and disrupts the host’s innate and adaptive immune systems. Findings suggest that CIP can associate with human C4b ligands, impacting the classical and lectin complement pathways [16]. Bordetella pertussis (Gram-negative bacteria) can evade attack by the complement system by releasing the virulence protein Vag8. Recombinant Vag8, when secreted endogenously, can hinder complement deposition on bacterial surfaces at the C4b level [17]. Recent studies have shown that flavivirus non-structural (NS1) proteins of dengue virus (DENV), West Nile virus (WNV), and yellow fever virus (YFV) are capable of attaching to C4, promoting its cleavage, reducing the deposition of C4b, and decreasing the activity of the C3 converting enzyme (C4bC2a), and thus confer a viral immune escape function. A recent study has uncovered a new antiviral mechanism that relies on C4 but not on late-acting complement components. C4 impedes human adenovirus infection by depositing lysed C4b onto the capsid, preventing its decomposition, release from endosomes, and infiltration into the cytoplasm [18]. Increased C4b may exacerbate tissue damage through inflammatory responses, suggesting that C4b could serve as a prognostic biomarker for type 1 diabetes. Severe bacterial infections can cause tissue destruction, cavity formation, and activation of the complement system.
Expressed on red blood cells and immune cells, CR1 is a type I membrane glycoprotein that centrally regulates the classical pathway, lectin pathway, and replacement pathway of the complement system [19,20,21,22,23]. The structure includes 30 closely related domains known as short consensus repeats (SCRs), complement control protein (CCP), or Sushi domain, in addition to a transmembrane domain and a cytoplasmic tail consisting of 43 amino acids [24]. SCR domains 1–28 are organized into four larger units, each consisting of 7 domains, known as long homologous repeat sequences (LHR), labeled LHR-A to LHR-D [25]. In LHR-A, LHR-B, and LHR-C, the initial three CCPs of each LHR facilitate interactions with either C3b or C4b. SCRs 1-3 mainly bind to C4b and have a weak affinity for C3b. CCPs 8-10 and CCPs 15-17 mainly bind to C3b but also have some affinity for C4b. In the genetic sequence of human CR1, CCPs 8-10 and CCPs 15-17 are more than 99% similar, which means that their structure and function are almost identical [26]. This study analyzed changes in C4 levels in porcine fresh serum at different time points within a co-incubation system using ELISA technology. Immune electron microscopy was employed to observe whether C4b participates in the immune adhesion of red blood cells to Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). The yeast two-hybrid technique was utilized to detect whether CR1-like binds to C4b. By combining homology modeling, molecular docking, and dynamics simulations, an interaction model between porcine CR1-like and C4b was constructed and optimized, further establishing the kinetic parameters of their interaction and elucidating their structural characteristics. Finally, protein expression and surface plasmon resonance (SPR) technology were used to validate the biological activity of the binding region between CR1-like and C4b. This study aims to provide a theoretical foundation for further revealing the mechanism of porcine red blood cell immune adhesion and its relationship with diseases, as well as to expand the in-depth understanding of the complement system’s functions.
2. Materials and Methods
2.1. Animals and Strains
Three healthy female 40-day-old Landrace piglets, each weighing approximately 20 kg, were obtained from Taigu County Guannong Agriculture and Animal Husbandry Technology Co, Ltd., Taigu, China. Our lab cryopreserved the PRRSV isolate JS-1 with a TCID_50_ of 10^−5.55^/mL.
2.2. Detection of Changes in Serum C4 Activity
Three experimental groups were set: group I (fresh pig serum + PBS), group II (fresh pig serum + PRRSV), and group III (inactivated pig serum + PRRSV). All groups were incubated at 37 °C; supernatants were collected via centrifugation at 0, 0.5, 1, 2, and 3 h post-incubation. C4 activity in supernatants was measured using a pig C4 ELISA kit (Shanghai Jianglai Biotechnology Co., Ltd., Shanghai, China), and optical density (OD) values were recorded. Statistical analysis was performed via one-way ANOVA using GraphPad PrismTM 8 (GraphPad Software, San Diego, CA, USA).
2.3. C4b in EM Detection of RBC-PRRSV Adhesion
Group A served as the cell control group, where porcine red blood cells were co-incubated with PBS. Group B was the virus blank group: 5 mL of PRRSV suspension was mixed with 5 mL of 4% paraformaldehyde (Wuhan Servicebio Technology Co., Ltd., Wuhan, China) and fixed by suspension at 4 °C. Group C was the virus control group, where PRRSV was first co-incubated with PBS and then with porcine red blood cells. Group D was the sensitized virus group, where PRRSV was first co-incubated with fresh porcine serum and then with porcine red blood cells. Group E was the inactivated serum group, where PRRSV was first co-incubated with inactivated porcine serum and then with porcine red blood cells. Group F was the antibody blockade group: 5 mL of porcine red blood cells were incubated with CR1-like antibody (laboratory-made, Patent No.: ZL201410308534.0) at 37 °C for 2 h, while 5 mL of PRRSV suspension was incubated with 250 μL of fresh porcine serum at 37 °C for 1 h and then mixed with 5 mL of porcine red blood cells and incubated at 37 °C for 30 min. The samples were sequentially centrifuged, fixed with glutaraldehyde (0.5%, 4 °C for 3 min; 3%, 4 °C for 10 h, Sigma-Aldrich, St. Louis, MO, USA), rinsed with PBS, dehydrated with an acetone gradient (50–100%,Sigma-Aldrich, St. Louis, MO, USA), and embedded in resin. Ultrathin sections of 80 nm were prepared, etched with hydrogen peroxide (Sigma-Aldrich, St. Louis, MO, USA) for 30 min, and then sequentially washed with PBS, blocked with BSA(Beijing Solarbio Science & Technology Co., Ltd., Beijing, China), incubated with C4b monoclonal antibody (Abcam plc, Cambridge, UK) at 4 °C for 20 h, and incubated with colloidal gold-labeled secondary antibody (Beijing Zhongshan Jinqiao Biotechnology Co., Ltd., Beijing, China) at room temperature for 2 h. Finally, the samples were double-stained with uranyl (Sigma-Aldrich Production GmbH, Buchs, Switzerland) acetate and lead citrate(Sigma-Aldrich Production GmbH, Buchs, Switzerland), dried, and observed under a transmission electron microscope (80 kV).
2.4. Pig CR1-like-C4b Domain Prediction
Amino acid sequences were retrieved from databases: human CR1 (UniProt: P17927), pig CR1-like (GenBank: NC_010451.4), human C4b (UniProt: P0C0L4), and pig C4b (GenBank: NC_010449.5). Domain annotation: Human CR1 and pig CR1-like sequences were uploaded to SMART (Letunic et al., 2021, mode: NORMAL) [27] to identify CCP domains; human CR1 C4b-binding domains (CCPs 1-3) were used for BLAST (National Center for Biotechnology Information, Bethesda, MD, USA) homology analysis against pig CR1-like to screen homologous pig CR1-like fragments. Signal peptide and transmembrane prediction: target sequences (pig CR1-like fragments and C4b) were analyzed via SignalP 6.0 [28] (species: eukaryotes) and DeepTMHMM-1.0 [29] to retain extracellular regions without signal peptides (to reduce computational complexity).
2.5. Construction of Recombinant Plasmid
Recombinant plasmids (pGBKT7-CR1-like CCPs 1-3/12-14/19-21; pGADT7-C4bα/β/γ) were synthesized by Sangon Biotech using pGBKT7/pGADT7 vectors (TaKaRa (Shanghai, China)). Plasmids were verified via double digestion and Sanger sequencing.
2.6. Self-Activation Detection and Toxicity Test
Self-activation and toxicity of recombinant plasmids were tested using the Yeastmaker™ Yeast Conversion System 2 kit (TaKaRa): plasmids included pGBKT7-derived (pGBKT7-CR1-like CCPs 1-3/12-14/19-21, empty pGBKT7) and pGADT7-derived (pGADT7-C4bα/β/γ, empty pGADT7), which were transformed into Y2HGold yeast (Angyu Biotechnology Co., Ltd., Shanghai, China) and cultured on selective plates for 2–4 days (pGBKT7: SD/-Trp, SD/-Trp/X-α-Gal, SD/-Trp/X-α-Gal/AbA; pGADT7: SD/-Leu, SD/-Leu/X-α-Gal, SD/-Leu/X-α-Gal/AbA); non-self-activation was verified by white colonies on SD/-Trp/SD/-Leu and SD/-Trp/X-α-Gal/SD/-Leu/X-α-Gal plus no colonies on SD/-Trp/X-α-Gal/AbA/SD/-Leu/X-α-Gal/AbA, non-toxicity by recombinant plasmid colonies matching empty vector colonies in size, and only non-self-activating, non-toxic plasmids were used for subsequent experiments.
2.7. Identification of CR1-like Active Fragment Combined with C4b Yeast Two-Hybrid
pGBKT7-CR1-like and pGADT7-C4b were transformed into the same yeast strain Y2HGold. The successfully transformed single colony was selected and cultured in 5 mL of YPDA at 30 °C and 250 rmp for 24 h. The bacteria solution was diluted and coated on SD/-Leu, SD/-Leu/-Trp (DDO), SD/-Leu/-Trp/X-α-Gal (DDO/X), and SD/-Leu/-Trp/X-α-gal/Aba (DDO/X/A) plates for 2~4 days at 30 °C to observe the growth of the colony and the growth color of the colony. Y2HGold transformed using pGBKT7-53 vector was mated with Y187 cells containing pGADT7-T and used as a positive control. Y2HGold transformed using pGBKT7-Lam vector was mated with Y187 cells containing pGADT7-T and used as a negative control.
2.8. Homology Modeling
All templates for protein crystal structures in homology modeling came from the PDB database (RCSB Protein Data Bank, https://www.rcsb.org/), and AlphaFold (DeepMind) [30] was employed to forecast the 3D structure of C4b and CR1-like CCPs 1-3. The model quality was assessed using the SAVES v6.0 (RCSB Protein Data Bank) web server and the PROCHECK (EMBL-European Bioinformatics Institute) [31] module of the ProSA (University of Vienna) [32] online inspection tool. CR1-like CCPs 12-14 and CR1-like CCPs 19-21 used the optimized model of Hou [33].
2.9. Molecular Docking and Kinetic Simulation
The Gramm-X v2.0 (University of Groningen) [34] online server was used for free docking of CR1-like CCPs 1-3, CR1-like CCPs 12-14, and CR1-like CCPs 19-21 with C4b, respectively. The optimal docking results were selected for molecular dynamics simulation. Desmond/Maestro (2022.1, D. E. Shaw Research; Schrödinger, Inc. (New York City, NY, USA)) was selected as the kinetic simulation software, TIP3P was added to the complex system using the TIP3P water model, and 0.15 M of sodium chloride solution (Sigma-Aldrich, St. Louis, MO, USA) was added to the equilibrium system. After minimizing and relaxing the system, 100 ns molecular dynamics simulations were executed in an isothermal isobaric ensemble at 300 K and 1 bar. Every 100 ps, the coordinates of the track were recorded, and Desmond’s simulated interaction diagram was employed for molecular dynamics analysis to review trajectory files and output model files with the least energy. Using PRODIGY v2.0 (University of Warsaw) [35], the affinity of proteins that interacted was calculated.
2.10. Prediction of Key Amino Acids in CR1-like-C4b Interactions
To analyze the interaction interface between CR1-like and C4b, LIGPLOT v4.5.3 (EMBL-European Bioinformatics Institute) was utilized, identifying the residues and types of interactions. DIMPLOT v4.5.3 (EMBL-European Bioinformatics Institute)was selected as the analysis mode for analyzing protein–protein interactions. The complex was further scanned with DrugScorePPI v1.0 (University of Düsseldorf) [36], mCSM-PPI2 v1.1 (King’s College London) [37], Robetta v3.0(University of Washington Baker Laboratory) [38], and BAlaS v1.2 (University of Edinburgh; University of Bristol) [39,40] to mimic alanine mutation. These tools predicted key residues of the complex by simulating alanine mutation scanning and calculated the change value of binding free energy before and after the mutation of residues. The key amino acid binding site was determined when the binding free energy change was more than 1 kcal mol^−1^.
2.11. C4b Is Expressed in the Large Intestine Expression System
The pET28a vector was employed to express C4b in the Escherichia coli expression system, incorporating 6His and Flag-ppase tags at the N-terminus while leaving the C-terminus untagged, thereby constructing the recombinant plasmid. The plasmid was transferred into Escherichia coli for expression, purified by preloaded Ni-NTA column, eluted with different concentrations of imidazole solution, and the fractions were collected and concentrated. Subsequently, further purification was performed using a strong cation exchange column (SP-HP), the fractions were equilibrated with 30 mM MES buffer (pH 6.5), 1 mM TCEP, and 5% glycerol, and the flow-through fraction was collected by continued washing with the same buffer after loading. Finally, fractions were collected and analyzed by SDS-PAGE after 0% to 100% gradient elution with 30 mM MES buffer (pH 6.5), 1 mM TCEP, 5% glycerol, and 1.0 M NaCl.
2.12. CR1-like Fragments and Their Mutants Are Expressed in Insect Cells
Recombinant plasmids were constructed utilizing pFastBac as a vector, incorporating 6His and sumo tags at the N terminus and flag tags at the C terminus. Recombinant baculoviruses were subsequently generated through the Bac-to-Bac system and transfected into SF9 insect cells for expression. Culture supernatants containing CR1-like fragments or their mutants were collected by centrifugation 72 h after infection. Purification was performed using a preloaded Ni-NTA column, and the fractions were collected and concentrated by elution through various concentrations of imidazole solutions. Further purification was carried out using a molecular sieve SEC column P200 10/500. The column was first equilibrated with 50 mM Tris buffer (pH 8.0), 150 mM NaCl, 1 mM TCEP, and 10% glycerol and eluted with the same buffer after sample loading. Fractions were collected according to the time or volume of the elution peak and analyzed by SDS-PAGE to identify fractions containing recombinant proteins.
2.13. Surface Plasmon Resonance (SPR) Detection
Single-cycle interaction analysis was performed on a Biacore 8K instrument (Cytiva (Marlborough, MA, USA)) with C4bα and C4bβ covalently immobilized on a CM5 sensor chip with an amount of approximately 1500 RU. Experiments were performed at 25 °C with a binding time of 60 s, a dissociation time of 300 s, and a flow rate of 30 μL/min. The running buffer used was 10 mM HEPES (pH 7.4), 150 mM NaCl, and 0.05% Tween-20. The size of CR1-like CCPs 1-3, CR1-like CCPs 12-14, and their mutants were 1.56 × 10^−2^ μM, 3.12 × 10^−2^ μM, 6.24 × 10^−2^ μM, 0.125 μM, 0.25 μM, 0.5 μM, 1 μM, 2 μM, and 4 μm, respectively. The concentration of μM was used as the mobile phase to detect the binding of C4bα and C4bβ to CR1-like CCPs 1-3, CR1-like CCPs 12-14, and their mutants, respectively. The apparent equilibrium dissociation constant (KD) was calculated as the ratio of dissociation and binding rate constants (kd/ka), with smaller KD values indicating stronger affinity.
3. Results
3.1. PRRSV Activates C4 and C4b Mediates RBC-PRRSV Immune Adhesion
Regarding the activation of complement C4 by PRRSV, the results showed that group I (fresh serum + PBS) had stable C4 activity in the first hour, with a non-significant slight decrease at 1–3 h (p > 0.05); group II (fresh serum + PRRSV) showed stable C4 activity initially but a gradual decline at 1–3 h; group III (inactivated serum + PRRSV) had nearly unchanged C4 activity. Statistics confirmed significant inter-group differences (p < 0.05), with group II having lower C4 activity than group I (p < 0.05, p < 0.001) and group III (p < 0.001), indicating PRRSV activates C4, leading to its cleavage and reduced activity at 1–3 h (Figure 1).
The findings in Figure 2A show that porcine red blood cells (RBCs) appeared normal with elliptical or varied shapes under transmission electron microscopy. Figure 2B highlights PRRSV in a red box. Figure 2C shows similar RBC morphology to Figure 2A, without immune adherence. In Figure 2D, immune adherence was observed, with colloidal gold particles indicating binding between the RBC membrane and PRRSV, involving C4b. Figure 2E,F show no RBC immune adherence. These results indicate that C4b plays a role in the immune adherence of porcine RBCs to PRRSV.
3.2. Bioinformatics Prediction of CR1-like-C4b Interaction Domains
BLAST alignment of the C4b-binding domain (CCPs 1-3) of human CR1 with porcine CR1-like identified three homologous fragments: CR1-like CCPs 1-3 (58% identity), CCPs 12-14 (63% identity), and CCPs 19-21 (63% identity). These fragments were selected as potential C4b-binding targets for subsequent experimental validation (Figure 3). Additionally, SignalP 6.0 and DeepTMHMM-1.0 confirmed that C4b and all three CR1-like fragments are extracellular regions without signal peptides, ensuring their spatial accessibility for in vivo interaction. Porcine CR1-like was also found to contain 31 CCP domains, showing structural conservation with human CR1 (37 CCP domains), which supports the hypothesis that porcine CR1-like retains C4b-binding activity.
3.3. Yeast Two-Hybrid Validation of CR1-like-C4b Interaction
Recombinant plasmids for yeast two-hybrid assays were first verified by double digestion and Sanger sequencing, with inserted fragments matching the expected sizes (557 bp for CR1-like CCPs 1-3/12-14, 725 bp for CCPs 19-21, and 1968 bp/2040 bp/885 bp for C4b α/β/γ chains). Toxicity assays confirmed no inhibitory effect of these plasmids on Y2HGold yeast growth, and self-activation assays ruled out non-specific reporter gene activation by either pGBKT7 or pGADT7 recombinant plasmids, ensuring the reliability of the assay system.
Specific binding between CR1-like fragments and C4b chains was confirmed on the stringent DDO/X/A medium (supplemented with AbA to eliminate non-specific binding). Blue colonies were observed in all experimental groups (CR1-like CCPs 1-3/12-14/19-21 paired with C4b α/β/γ chains), while the negative control (pGBKT7-Lam + pGADT7-T) showed no colony growth. Colonies in the CR1-like CCPs 1-3 + C4b α and CR1-like CCPs 12-14 + C4b β groups exhibited more vigorous growth, indicating stronger binding affinity between these pairs (Figure 4).
3.4. Molecular Simulation of CR1-like-C4b Interaction and Key Amino Acid Prediction
Key amino acids critical for CR1-like-C4b binding were predicted using four independent tools (DrugScorePPI, mCSM-PPI2, Robetta, and BAlaS). For CR1-like CCPs 1-3, Ile118 and Tyr136 were validated by at least three tools; for CCPs 12-14, eight residues (including Leu15 and Arg32) were validated by at least four tools; and for CCPs 19-21, eight residues (including Gln15 and Tyr74) were validated by at least three tools. The spatial locations of these key residues further confirmed their direct involvement in the binding interface (Figure 5). Three-dimensional models of porcine CR1-like CCPs 1-3 and C4b were constructed using AlphaFold2 and validated as reliable by ProSA (normal Z-scores) and PROCHECK (core region residues > 89%), and molecular dynamics simulations confirmed the structural stability of C4b-CR1-like complexes.
3.5. Recombinant Protein Expression and SPR Validation of CR1-like-C4b Binding Affinity
Recombinant C4b α/β chains were expressed in Escherichia coli and purified via Ni-NTA and strong cation exchange chromatography (SP-HP). Western blot analysis of 400 mM imidazole eluates confirmed the presence of the Flag tag and expected molecular weights (70–95 kDa) for both C4b α and β chains, verifying high purity and suitability as binding substrates (Figure 6).
Recombinant CR1-like CCPs 1-3 were expressed in insect cells using the Bac-to-Bac system and purified via Ni-NTA and size-exclusion chromatography (SEC). Western blot analysis of concentrated SEC fractions showed a specific Flag tag signal at 45–55 kDa, confirming successful purification and identity as a valid binding receptor (Figure 7).
The 118I mutant of CR1-like CCPs 1-3 was also expressed in insect cells and purified to validate the role of Ile118. SDS-PAGE analysis showed the mutant was soluble in extracellular fractions (45–55 kDa band) and successfully purified via Ni-NTA and SEC; Western blot confirmed the Flag tag and correct molecular weight. In contrast, the 90N93Y mutant formed insoluble intracellular aggregates and could not be purified (Table 1, Figure 8).
Surface plasmon resonance (SPR) assays were performed to measure the binding affinity between CR1-like fragments (or the 118I mutant) and C4b chains. Concentration-dependent binding signals were observed for CR1-like CCPs 1-3 with both the C4b α and β chains, while CR1-like CCPs 12-14 showed binding signals only with the C4b α chain. Importantly, the 118I mutant exhibited significantly reduced binding signals to the C4b β chain compared to wild-type CR1-like CCPs 1-3, while its binding to the C4b α chain remained barely unchanged (Figure 9).
Quantitative analysis of dissociation constants (KD) further confirmed these observations. CR1-like CCPs 1-3 bound C4b α with a KD of 0.226 ± 0.021 μM and C4b β with a KD of 0.173 ± 0.015 μM, indicating high affinity. The 118I mutant showed no significant change in KD for C4b α (0.271 ± 0.019 μM) but a 6.9-fold increase in KD for C4b β (1.2 ± 0.12 μM, p < 0.01), confirming that Ile118 is critical for the binding affinity between CR1-like and the C4b β chain (Figure 10).
4. Discussion
4.1. Functional Mechanisms of CR1-like-C4b Interaction in Complement Regulation and Immune Adhesion
The complement system is a core component of innate immunity, mediating microbial clearance and tissue homeostasis via cytokine/chemokine upregulation. Complement C4, a key molecule in classical/lectin pathways, is cleaved into C4b to initiate opsonization and pathogen clearance. Human CR1 CCPs 1-3 are known to bind C4b [24], and this study explored the functional homology of porcine CR1-like.
We confirmed that PRRSV activates porcine complement C4 in fresh serum, leading to C4b generation—evidenced by reduced C4 activity and C4b-mediated RBC-PRRSV immune adhesion (immunogold-labeled colloidal gold particles observed via electron microscopy). BLAST analysis identified three porcine CR1-like fragments (CCPs 1-3/12-14/19-21) with 58–63% identity to the human CR1 C4b-binding domain, and yeast two-hybrid assays validated their interaction with C4b α/β/γ chains. Due to C4b’s large molecular weight, we expressed it as separate chains, which is a study limitation.
AlphaFold 2-generated 3D models of CR1-like CCPs 1-3 and C4b (validated by SAVES v6.0 and proSA) were used for molecular docking and dynamics simulations. Four tools predicted hot spot residues, but sample exhaustion prevented direct functional validation—we inferred mechanisms via experimental data and the literature.
First, complement activation regulation: SPR showed Ile118 mutation in CR1-like CCPs 1-3 increased KD for the C4b β chain from 0.173 μM to 1.2 μM, with molecular simulations confirming Ile118 forms hydrophobic interactions with C4b β chain’s Leu68/Asn70. This suggests CR1-like competes with CD46 for C4b binding [24,26], restricting C3 convertase (C4bC2a) formation to prevent excessive complement activation and inflammatory damage [33].
Second, immune adhesion efficiency: Electron microscopy and yeast two-hybrid/SPR confirmed C4b mediates RBC-PRRSV adhesion, with CR1-like as the C4b receptor. CR1-like-C4b binding enhances RBC avidity for C4b-opsonized PRRSV, and reduced binding of the Ile118 mutant highlights key residues’ role in adhesion efficiency—analogous to human CR1’s regulation of RBC-mediated pathogen clearance [19,24].
4.2. Roles of RBC-PRRSV Adhesion in PRRSV Pathogenesis and Immune Evasion
4.2.1. Facilitation of PRRSV Pathogenesis via RBC-Mediated Viral Dissemination
The activity-dependent CR1-like-C4b adhesion (SPR-verified KD = 0.173 μM for CCPs 1-3/C4b β chain; 6.9-fold KD increase after Ile118 mutation) promotes PRRSV pathogenesis. As the most abundant blood cells, RBCs act as “mobile carriers” for C4b-opsonized PRRSV, efficiently transporting it to target organs (lungs and lymph nodes) via circulation. This enhances PRRSV’s tropism for alveolar macrophages, exacerbating interstitial pneumonia and systemic inflammation—consistent with PRRSV’s respiratory tropism and rapid dissemination. The Ile118 mutation-induced adhesion reduction suggests targeting this axis may impair viral spread, providing a candidate for pathogenicity validation in mutant animal models.
4.2.2. Contribution to PRRSV Immune Evasion via Antigen Masking and Complement Modulation
Beyond known PRRSV escape strategies (interferon inhibition and MHC downregulation), RBC-PRRSV adhesion represents a novel pathway. C4b-opsonized PRRSV binds RBC CR1-like to mask antigen epitopes, evading macrophage/dendritic cell recognition. Additionally, CR1-like’s complement regulatory activity inhibits local complement overactivation, reducing inflammatory cell (neutrophil) recruitment and creating an “immune sanctuary” for viral survival. This aligns with flaviviruses’ NS1-mediated C4 binding and escape [18], but our study is the first to report RBC-mediated complement-dependent evasion.
Collectively, the CR1-like-C4b axis dual-regulates PRRSV pathogenesis and evasion. Targeting this interaction may simultaneously inhibit viral dissemination and escape, offering a promising anti-PRRSV translational direction.
4.3. Technical Challenges and Optimization Strategies in Experimental Processes
C4b α/β chains were expressed in E. coli and CR1-like CCPs 1-3/118I mutant in insect cells. Tag removal caused protein instability and diffuse bands, so tagged proteins were used—with ExPASy-Prot Param predicting ~40 kDa molecular weight (larger apparent size via SDS-PAGE due to glycosylation). The 90N93Y mutant formed insoluble aggregates and was unobtainable.
Recombinant protein yield/purity variability (common in multi-domain complement proteins [33,40]) stemmed from the following: (1) E. coli’s tendency for C4b β chain (≈75 kDa) misfolding/inclusion bodies and insect cell batch-to-batch glycosylation differences [33]; (2) minor induction condition variations (±2 °C, 0.1–0.5 mM IPTG, MOI) causing 15–20% yield fluctuations [40]; (3) partial proteolysis during Ni-NTA purification (faint 10–15 kDa bands), despite protease inhibitors.
This variability only slightly interfered with results: SPR KD values for CCPs 1-3/C4b α/β showed <10% variation (0.226 ± 0.021 μM/0.173 ± 0.015 μM), and degradation products (<5% of total protein) did not bind C4b [35]. Key limitations included C4b’s separate-chain expression (α/β only) and CR1-like’s partial fragment expression (CCPs 1-3/12-14; CCPs 12-14 from our lab’s pre-validated stock [33]). SPR assays only analyzed individual C4b chains (full-length ≈ 204 kDa is difficult to express solubly [33]), with CCPs 1-3 binding C4b α/β (KD = 0.226/0.173 μM), CCPs 12-14 binding only C4b α (KD = 0.317 μM), and Ile118 mutant showing a 6.9-fold higher KD for C4b β (1.2 μM, p < 0.01). Future full-length C4b-CR1-like interaction studies could use (1) baculovirus–insect cell co-expression [40]; (2) fragment complementation; and (3) BLI (more suitable for large molecules [35]).
An additional challenge was CR1-like CCPs 1-3/118I instability after tag removal, forcing tagged protein use for SPR—this may have minorly interfered with physiological interaction validation, though core binding trends remained unaltered.
Optimization strategies for future studies: (1) porcine protein-adapted vectors (pET-32a, 6 × His-SUMO) to reduce tag interference [33]; (2) optimized induction (16 °C, 0.1–0.5 mM IPTG, 16–20 h) to minimize inclusion bodies [40]; (3) SEC post Ni-NTA purification + PMSF to reduce aggregation/degradation.
4.4. Future Research Directions to Validate In Vivo Functions
To validate functional inferences, CRISPR-Cas9-generated porcine CR1-like mutant cell lines (Ile118Ala, Asn90Ala) or transgenic pigs should assess the following: (1) mutant effects on complement activation markers (C3a and C5a); (2) in vitro/in vivo PRRSV clearance via RBC immune adhesion; (3) lung inflammation and pathological changes in infected mutants. These will clarify CR1-like-C4b’s in vivo role in anti-PRRSV immunity.
Two additional in vitro-derived hypotheses require investigation: (1) full-length C4b (α/β/γ assembled) may have higher affinity for CR1-like than individual chains (CCPs 1-3 bind α/β strongly), requiring soluble full-length C4b expression [4.3]; (2) functional roles of predicted key residues (Tyr136 in CCPs 1-3 and Leu15 in CCPs 12-14) remain unvalidated—Ala-scanning mutagenesis should clarify their contributions.
4.5. Translational Implications: Therapeutic Potential Targeting the CR1-like-C4b Axis
Based on CR1-like-C4b interaction data, two anti-PRRSV strategies are proposed: (1) Small-molecule inhibitors: Virtual screening (ZINC database) of compounds blocking CR1-like-C4b binding, using interaction models and key residues (Ile118, Tyr136)—analogous to human CR1-C3b inhibitor design [26]; (2) anti-CR1-like monoclonal antibodies targeting CCPs 1-3/12-14, which may block PRRSV-C4b-RBC adhesion and reduce lung inflammation—specificity is critical to avoid cross-reactivity with other porcine complement receptors [33].
Key challenges and solutions: (1) Poor drug delivery: Lipid nanoparticles enhance inhibitor accumulation in PRRSV-infected lungs [40]; (2) Antibody cross-reactivity: Humanized/camelid single-domain antibodies reduce off-target binding [35]. These strategies require further validation but provide a translational framework for PRRSV control.
5. Conclusions
In vitro, fresh serum mediates C4b deposition on PRRSV via complement activation. Porcine erythrocyte CR1-like molecules specifically bind C4b through CCPs 1-3, 12-14, and 19-21, facilitating immune adhesion to PRRSV. Mutation of isoleucine 118 (118I) in CR1-like CCPs 1-3 impairs C4b binding, highlighting its critical role in complement-mediated erythrocyte immune adhesion.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Coss S.L. Zhou D. Chua G.T. Aziz R.A. Hoffman R.P. Wu Y.L. Ardoin S.P. Atkinson J.P. Yu C.Y. The complement system and human autoimmune diseases J. Autoimmun.202313710297910.1016/j.jaut.2022.10297936535812 PMC 10276174 · doi ↗ · pubmed ↗
- 2KallionpääH. Elo L.L. Laajala E. Mykkänen J. Ricaño-Ponce I. Vaarma M. Laajala T.D. Hyöty H. Ilonen J. Veijola R. Innate immune activity is detected prior to seroconversion in children with HLA-conferred type 1 diabetes susceptibility Diabetes 2014632402241410.2337/db 13-177524550192 · doi ↗ · pubmed ↗
- 3Law S.K. Dodds A.W. The internal thioester and the covalent binding properties of the complement proteins C 3 and C 4Protein Sci.1997626327410.1002/pro.55600602019041627 PMC 2143658 · doi ↗ · pubmed ↗
- 4Sim R.B. Sim E. Autolytic fragmentation of complement components C 3 and C 4 and its relationship to covalent binding activity Ann. N. Y. Acad. Sci.198342125927610.1111/j.1749-6632.1983.tb 18114.x 6202197 · doi ↗ · pubmed ↗
- 5Wallis R. Mitchell D.A. Schmid R. Schwaeble W.J. Keeble A.H. Paths reunited: Initiation of the classical and lectin pathways of complement activation Immunobiology 201021511110.1016/j.imbio.2009.08.00619783065 PMC 2824237 · doi ↗ · pubmed ↗
- 6Pilely K. Skjoedt M.O. Nielsen C. Andersen T.E. Louise Aabom A. Vitved L. Koch C. Skjødt K. Palarasah Y. A specific assay for quantification of human C 4c by use of an anti-C 4c monoclonal antibody J. Immunol. Methods 2014405879610.1016/j.jim.2014.01.01124472768 · doi ↗ · pubmed ↗
- 7Barilla-La Barca M.L. Liszewski M.K. Lambris J.D. Hourcade D. Atkinson J.P. Role of membrane cofactor protein (CD 46) in regulation of C 4b and C 3b deposited on cells J. Immunol.20021686298630410.4049/jimmunol.168.12.629812055245 · doi ↗ · pubmed ↗
- 8Wieser M. Francisci T. Lackner D. Buerckstuemmer T. Wasner K. Eilenberg W. Stift A. Wahrmann M. Böhmig G.A. Grillari J. CD 46 knock-out using CRISPR/Cas 9 editing of h TERT immortalized human cells modulates complement activation P Lo S ONE 201914 e 021451410.1371/journal.pone.021451430958843 PMC 6453361 · doi ↗ · pubmed ↗