Genetic Diversity of the Non-Polio Enteroviruses Detected in Samples of Patients with Aseptic Meningitis in the Ural Federal District and Western Siberia
Tarek M. Itani, Vladislav I. Chalapa, Anastasia K. Patrusheva, Evgeniy S. Kuznetsov, Aleksandr V. Semenov

TL;DR
This study identifies the genetic diversity of non-polio enteroviruses causing aseptic meningitis in the Ural and Siberia regions of Russia.
Contribution
The study provides new insights into the genotypic composition and evolution of NPEVs in a specific geographic region using Sanger and NGS sequencing.
Findings
E30, E6, and CVA9 were the most prevalent NPEV strains in the study region.
Thirteen whole NPEV genomes were assembled using next-generation sequencing.
The study highlights the importance of genetic monitoring for public health and outbreak prediction.
Abstract
Human non-polio enteroviruses (NPEVs) cause a plethora of infections in humans, ranging from mild to severe neurological diseases including aseptic meningitis. NPEVs are the leading cause of aseptic meningitis in both children and adults worldwide. In Russia, reports of NPEV infections have surged, especially in the post-COVID era starting in 2022, with elevated infection rates into 2023. A comprehensive examination of the whole genome is crucial for understanding the evolution of NPEV genes and for predicting potential outbreaks. This study focused on identifying the circulating NPEV strains in the Ural Federal District and Western Siberia, using Sanger sequencing and next-generation sequencing (NGS) methodologies. Biological samples were collected from (n = 225) patients diagnosed with aseptic meningitis. Bioinformatics analysis targeted the nucleotide sequences of the major capsid…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
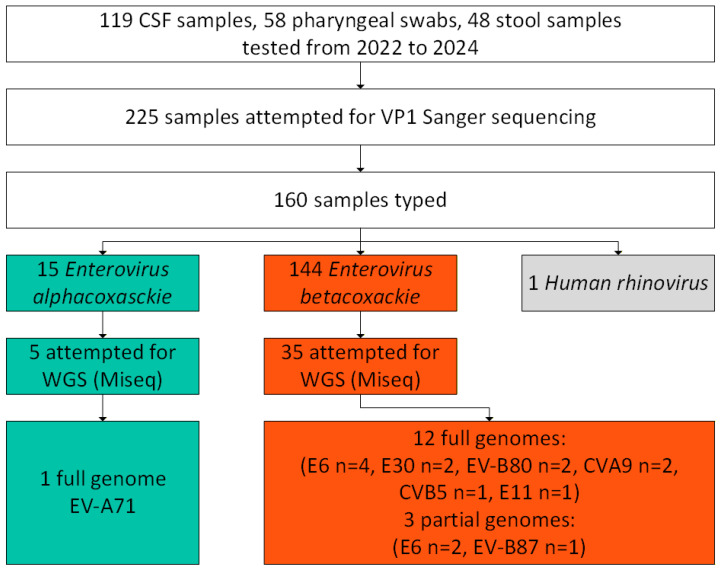 Figure 1
Figure 1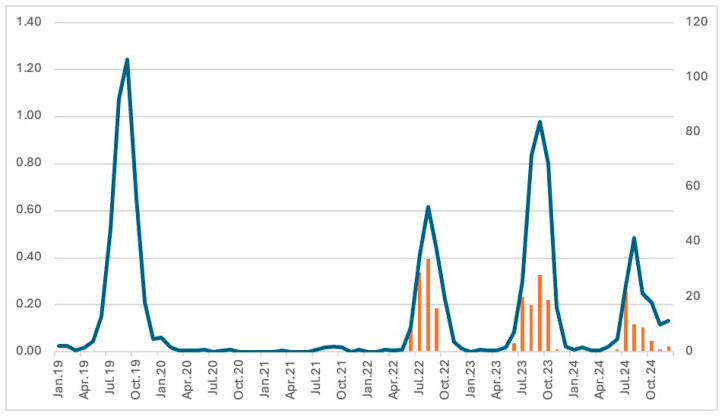 Figure 2
Figure 2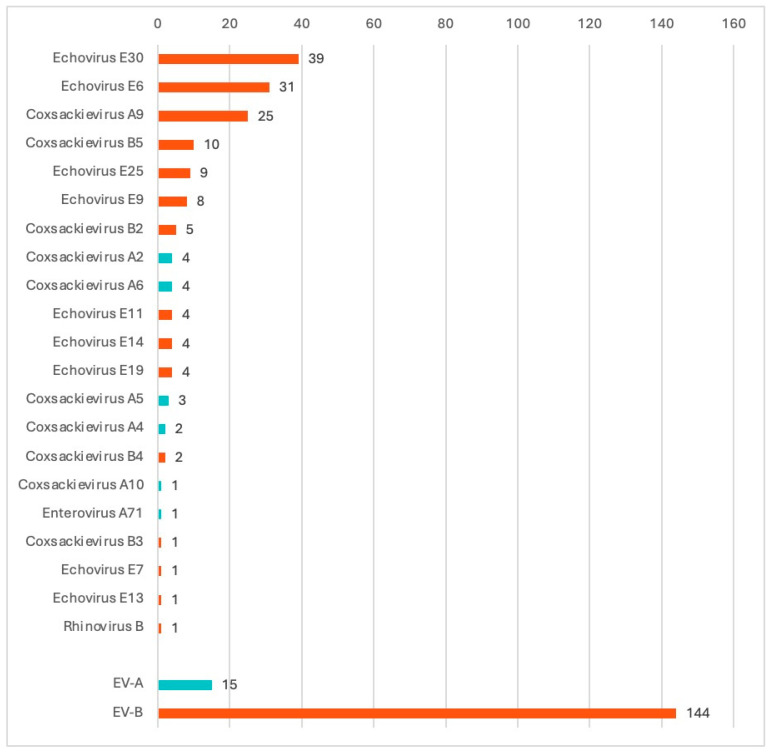 Figure 3
Figure 3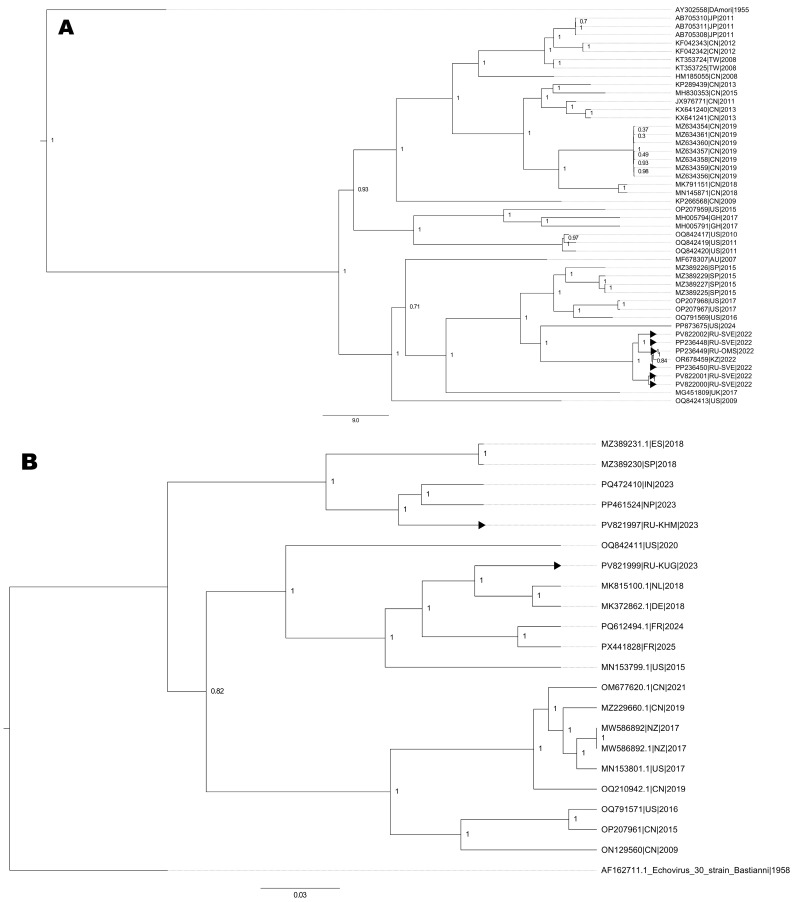 Figure 4
Figure 4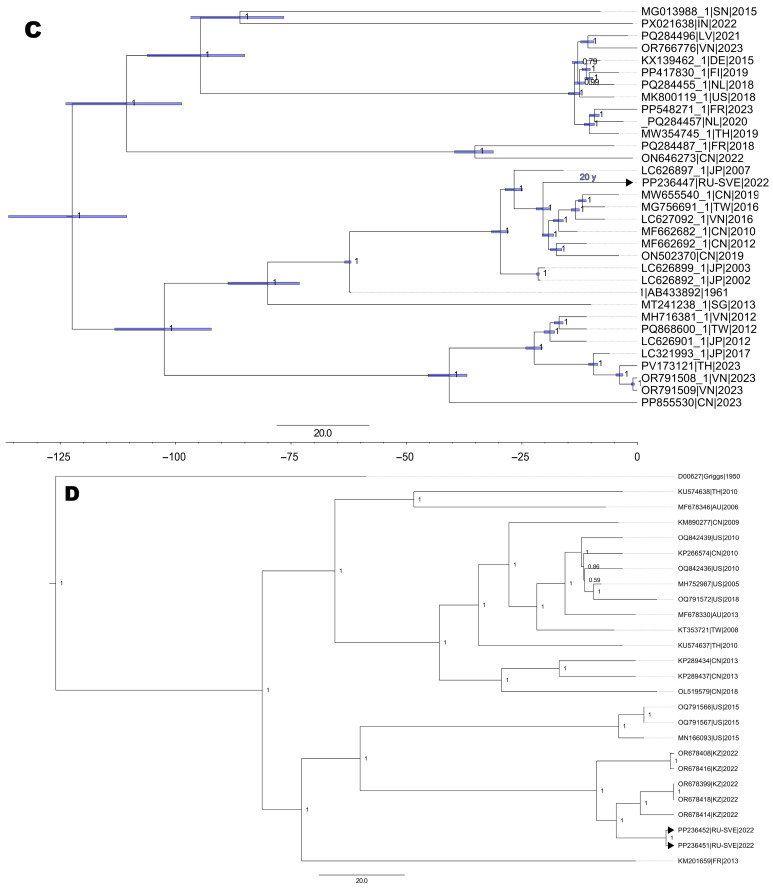 Figure 5
Figure 5- —Russian Federal Service for Surveillance on Consumer Rights Protection and Human Wellbeing
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infections and Immunology Research · Respiratory viral infections research · Immunodeficiency and Autoimmune Disorders
1. Introduction
Non-polio enteroviruses (NPEVs) constitute the largest genus within the Picornaviridae family. These are small naked single-stranded positive RNA viruses of approximately 7500 nucleotides grouped into four species, Enterovirus alphacoxsackie (EV-A), Enterovirus betacoxsackie (EV-B), Enterovirus coxsackiepol (EV-C), and Enterovirus deconjuncti (EV-D), and more than 100 distinct genotypes [1,2]. Most viruses in this genus cause infections in humans, ranging from mild respiratory illnesses to severe neurological diseases [3]. Clinical manifestations may develop after an incubation period of 3 to 21 days. NPEVs are distributed worldwide and have a seasonal incidence pattern in temperate regions during summer and fall, while they occur year-round in tropical regions [4].
NPEVs are associated with a range of central nervous system infections and are the leading cause of aseptic meningitis in adults and children worldwide [5,6]. Aseptic meningitis is a clinical entity characterized by an elevated leukocyte count in the cerebrospinal fluid (CSF), typically manifesting as pleocytosis. This condition is distinguished by the absence of positive findings on Gram stain and bacterial culture. Furthermore, aseptic meningitis is defined by the lack of a discernible parameningeal focus or systemic illness, underscoring its etiological heterogeneity and the challenges in identifying its causative agents. This leads to favorable clinical outcomes [7] and manifests differently depending on the patient’s age and immune health [8]. Nearly 50 NPEV types have been identified as causes of aseptic meningitis globally, distributed across the four species [9]. EV-B types including CV-B3-5, CVA9, and E6, 9, and 30 are the most identified in patients with aseptic meningitis [5] with E30 as the most common cause of viral meningitis and reported in outbreaks worldwide [8,10].
The NPEV genome includes a long single open reading frame (ORF) flanked by 5’UTR and 3’UTR. The ORF encodes a single polyprotein that can be cleaved into mature viral capsid proteins P1 (VP4, VP2, VP3, and the major capsid protein VP1) and non-structural proteins P2 (2A–C) and P3 (3A–D). VP1, VP2, and VP3 are on the surface of the viral capsid and are under immune pressure, whereas VP4 is inside the capsid [11]. VP1 is the most external and immunodominant capsid protein that contains the neutralization epitopes, and can be used for NPEV genotyping [12]. Currently, Sanger sequencing of the VP1 capsid protein gene is the gold standard for NPEV genotyping. But whole-genome sequencing has become increasingly affordable, accessible, and cost-effective, which will likely allow improved typing in the near-future [13].
There is limited information on circulating NPEV genotypes in aseptic meningitis patients from the Ural Federal District (UFD) and Western Siberia (WS). Such knowledge is essential for laboratory diagnostics, patient management, and future outbreak responses. To gain a deeper understanding of the epidemiology of aseptic meningitis for the period spanning 2022 to 2024, this study aimed to identify the NPEV circulating in the Ural Federal District and Western Siberia population using VP1 gene typing and whole-genome sequencing (WGS). Additionally, this research aimed to investigate the correlation between NPEV types and the genomic evolution of the disease.
2. Materials and Methods
2.1. Ethical Consideration
This research was performed as part of the Russian state program for NPEV surveillance. Since 2017, national enterovirus surveillance has been collecting laboratory data and biological samples from enteroviral infections in the UFD and WS. The participating centers were asked to report NPEV meningitis/encephalitis detections monthly to the Ural-Siberian regional scientific and methodological center for the study of enteroviral infections. According to national regulations, the use of anonymous samples and data from state epidemiological surveillance does not require informed consent. The research was performed with a waiver of consent under IRB № 4 of the Local Ethics Committee of the State Scientific Center of Virology and Biotechnology “Vector” (date of approval 24 June 2022).
2.2. Clinical Samples and Data Collection
A total of 281 positive samples from 225 patients with NPEV meningitis were received in the Laboratory of Enteric Infections from January 2022 to the end of December 2024. These samples included 119 residual CSF samples positive for NPEV from patients with confirmed aseptic meningitis, which were collected by participating centers, analyzed by real-time PCR amplification using “AmpliSens Enterovirus-FL” (InterLabService Ltd., Moscow, Russia), and transported to our laboratory at approximately 4 °C. Additionally, pharyngeal swabs (n = 89) and stool specimens (n = 73) were included in our study. Pharyngeal swabs (n = 58) and stool samples (n = 48) were also included in our study when CSF samples were not available or untypable (Table 1). All samples were stored at −20 °C until processing. For laboratory studies, stool suspensions were prepared by adding approximately 0.1–0.2 g of each stool sample to polypropylene test tubes, containing 1–1.5 mL of PBS.
2.3. Enterovirus Diagnosis and Molecular Genotyping
First, the samples were centrifuged for 2 min at 10,000× g at room temperature in order to remove the solid particles. Viral RNA genome was isolated from CSF, pharyngeal swabs, and stool suspensions employing the RIBO-prep kit (InterLabService Ltd., Moscow, Russia) according to manufacturer’s protocol. cDNA synthesis was carried out using the REVERTA-L-100 kit (InterLabService Ltd., Moscow, Russia). Next, two rounds of PCR targeting the VP1 gene were performed according to Nix et al. with minor modifications as previously described [14]. The reaction products were separated and visualized on 1.5% agarose gels and extracted by using the Cleanup Standard reagent kit (Evrogen JSC, Moscow, Russia). Purified products were sequenced in both directions (forward and reverse) using the deoxy sequencing Sanger method with the BigDye™ Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Austin, TX, USA) on an automatic genetic analyzer 3130 Genetic Analyzer (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacturers’ protocol.
2.4. Virus Isolation, Preparation of Libraries, Whole-Genome Sequencing
All the samples that were attempted for WGS were inoculated on RD (Rhabdomyosarcoma) for the detection of non-polio enteroviruses as per the WHO non-polio enteroviruses Guidelines (Figure 1). Briefly, a volume of 0.2 mL of treated fecal sample, CSF, or pharyngeal swab suspension was used for the RD inoculation. The inoculated tubes were then incubated at 37 °C for a period of 1–10 days, until a complete cytopathic effect was observed under an ordinary light microscope. Uninfected cells were used as a negative control. NPEV strains isolated on RD cell lines were directed to WGS. Cells were frozen and thawed three times and then subjected to an RNA extraction procedure. A volume of 200 µL of the cell culture supernatant was used for total RNA extraction with ExtractRNA reagent (Evrogen JSC, Moscow, Russia) for further molecular biological analysis. The RNA concentration was determined on a Qubit 4.0 fluorimeter using the Qubit RNA Assay kit (Qubit™ RNA BR Assay, Thermo Fisher Scientific, Waltham, MA, USA). To confirm the isolation of NPEV RNA, real-time PCR amplification was performed using a set of reagents “AmpliSens Enterovirus-FL” (InterLabService Ltd., Moscow, Russia) in a StepOnePlus amplifier (Applied Biosystems, Austin, TX, USA).
cDNA synthesis was performed using the Mint cDNA synthesis kit (Evrogen JSC, Moscow, Russia). DNA libraries were obtained with the NEB Next Ultra II RNA Library Prep Kit (New England Biolabs, Ipswich, MA, USA) and DNA concentration was measured after library preparation on a Qubit 4.0 fluorimeter using the Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) for the quantitative determination of double-stranded DNA. The quality of each library was assessed by analyzing a volume of 2 µL using a Qsep One Plus fragment analyzer (BioOptic, New Taipei City, Taiwan) with a high-resolution cartridge and by visualizing on a 2% agarose gel (TBE buffer, 180 V for 30 min). Prepared libraries were pooled into a single tube pool of 4 nM by dilution in sterile water. The concentration of the final library pool was measured using the QubitTM dsDNA HS Assay Kit. Genomic libraries were sequenced on the Illumina MiSeq platform (Illumina, San Diego, CA, USA), utilizing the MiSeq v2 cartridge, which generates paired readings of 2 × 250 cycles. All kits were used following the manufacturer’s instructions.
2.5. Bioinformatic Analysis for Sanger and WGS
All VP1 sequences were visualized using Chromas 2.6.6 (Technelysium, Pty, Ltd, South Brisbane, QLD, Australia. version 2.6.6) and NPEV genotyping was carried out by comparing the results of direct genome sequencing of NPEV with the reference sequences presented in the international GenBank database of genetic data and the BLAST service (https://blast.ncbi.nih.gov/, accessed on 19 June 2025). In addition, genotyping was confirmed using the online RIVM program (https://www.rivm.nl/mpf/typingtool/enterovirus/, accessed on 2 July 2025).
For WGS, low-quality reads were filtered, and adaptors were trimmed with Trimmomatic. The quality of the read was visualized using FastQC software V.0.11.9 before and after trimming. De novo assembly was performed using SPAdes genome assembler V3.9 [15] programs with default parameters.
2.6. Phylogenetic and Statistical Analysis
Using MEGA software V11 [16], nucleotide and amino acid sequences were aligned and subjected to preliminary analysis through ClustalW procedures. Next, representative whole-genome sequences—one for each identified genotype—were added to the alignments from the GenBank database. Then, alignments were analyzed in BEAST software V1.10.4 [17] while parsing collection years for each strain and performing calibration of the strict molecular clock for EV-A71 because of the authors’ concern about its origin (a strict clock was used to prevent over-parameterization). The evolutionary models for all trees were GTR+G+I based on the results of ModelTest-NG software V0.1.7 [18]. The obtained phylogenetic trees were inspected using Tracer software V1.7.2 [19], finalized in TreeAnnotator software V1.10.4 (part of BEAST package) and visualized in FigTree software V1.4.4 [20] with minimal graphical adaptations. During the procedure, no formal model selection was conducted.
3. Results
3.1. Incidence of Enterovirus Meningitis in the Ural Federal District and Western Siberia
We analyzed the data on aseptic meningitis that was provided to the Ural-Siberian regional scientific and methodological center for the study of enteroviral infections from January 2019 to December 2024. All these cases were laboratory-confirmed, but not fully genotyped by partial VP1-sequencing. This allowed us to better understand the incidence of enterovirus meningitis in our region, and to better understand the pre- and post-pandemic context. Overall, 2465 cases of NPEV-associated aseptic meningitis were recorded in the UFD and WS (Figure 2). The average monthly incidence rate was 0.19 per 100,000 inhabitants with most cases occurring in the Sverdlovsk oblast (40%; 987/2465), Khanty-Mansi autonomous okrug (13.9%; 342/2465), and Novosibirsk oblast (12%, 295/2465). A notable increase in the number of aseptic meningitis NPEV-positive cases was observed in September–October 2023, declining in November 2023 and peaking again in the summer of 2024. The monthly incidence showed a summer-autumn pattern with 94.85% of the cases (2338/2465) occurring from June until November, with a peak in August–September. The long-term dynamics of the incidence of NPEV aseptic meningitis were characterized by a large decrease in 2020 and an increase in 2022, with a return to pre-pandemic values in 2023.
The most common clinical symptoms were fever, headache, and vomiting. A total of 281 positive samples from 225 patients with NPEV meningitis were received in the Laboratory of Enteric Infections from January 2022 to the end of December 2024. We analyzed and genotyped 225 samples (119 CSF, 58 pharyngeal, and 48 fecal samples). The age of the patients ranged from 1 year to 57 years, with a median of 8 years, and the male-to-female ratio was 1:0.63 (138 males/87 females). Schoolchildren, from 7 to 17 years, constituted 57.3% of the patients (129/225). Preschool children, from 3 to 6 years, constituted the second largest age group at 27.1% (61/225).
3.2. Partial VP1 Genotyping
From the 225 aseptic meningitis patients, NPEV genotyping was successfully performed in 159 (70.7%), and more specifically 84/119 CSF samples (70.6%), 42/48 stool samples (87.5%), and 33/58 pharyngeal swabs (56.9%), indicating that stool and CSF samples are the most suitable biological samples for meningitis diagnosis. One nasopharyngeal sample typed as rhinovirus B, and was excluded from the analysis. When multiple samples from the same patient were analyzed, we detected the same NPEV genotype in all tested samples. VP1 genotyping revealed twenty-one different NPEV genotypes circulating among patients with aseptic meningitis in the UFD and WS (Figure 2). Most genotypes belonged to the Enterovirus betacoxsackie (EV-B) genus (90%), and less to the Enterovirus alphacosackie (EV-A) genus (9.4%), and only one rhinovirus was detected (0.6%, belonging to a different genus). Some Federal districts reported a low number of registered cases and sent a few samples. Most samples were received and typed from the Sverdlovsk oblast, Khanty-Mansi autonomous okrug, and Novosibirsk oblast (Table 2). Only two samples were successfully typed in the Omsk oblast (from two samples sent to our center) and one case was typed in the Tyumen oblast (from three samples sent). Three Federal districts did not send samples from aseptic meningitis patients (Yamalo-Nenets autonomous okrug, Kemerovo oblast, and Altai krai). The most prevalent genotypes were E30 (24.3%, 39/160), E6 (19.3%, 31/160), CVA9 (15.6%, 25/160), CVB5 (6.25%,10/160), and E25 (5.6%, 9/160) (Table 2 and Figure 3). EV-A species were mainly detected in children less than 6 years (12 out of 15 detected) and from the Sverdlovsk oblast (14 out of 15 detected). E30 was mainly detected in the schoolchildren group from 7 to 17 years (32 out of the 39 detected E30), while E6 was almost exclusively detected in patients from 3 to 18 years (28 out of the 31 detected E6). The sole EV-A71 strain was isolated in a patient with aseptic meningitis from a nasopharyngeal sample.
3.3. Whole-Genome Sequencing (WGS) and Sequence Assembly
Forty NPEV strains (35 EV-B and 5 EV-A) from respective clinical cases were isolated initially by cell culture on VP1 (RD cells). We chose the most predominant NPEV types found by VP1 genotyping: E30, E6, CVA9, CVB5, CVA2, and the potentially neurovirulent EV-A71. Due to the reduced sensitivity that is frequently encountered with conventional cell culture systems, repeated passages were required in most cases until the observation of a cytopathic effect. WGS was attempted with all these NPEV isolates using an Illumina MiSeq and only 16 were successfully sequenced by the NGS method. There were differences between the subsequent culture/NGS typing and the VP1 typing for three samples. Two samples were typed as E6 and one sample as E30 (1 sample) by the VP1 approach, while they were typed by the NGS method as EV-B80 (2 strains) and EV-B87 (1 strain). A list of the sequenced genome is provided in Table 3.
3.4. Phylogenetic Analysis of Full-Length Sequences
Phylogenetic analysis of E6 genovariants (six isolates) showed that most of the identified enteroviruses circulated in the studied regions for some years. As shown in Figure 4A, strains of E6 clustered closely with strains circulating in Baltimore, USA, from 2016 to 2017 [5] and strains that circulated in Spain during 2015 [13], while they were distant from Chinese E9 strains reported between 2013 and 2019.
Concerning the reconstruction of phylogenetic events for nucleotide sequences from two E30 full-length genomes, the topology of the cladogram demonstrates the formation of two distinct nodes (Figure 4B): (i) PV821997.1 clustered with MZ389231, an E30 strain circulating in Spain in 2018 [13], forming a cluster (sub-genogroup II), but was still 10% different; (ii) another cluster consisted of PV821999.1 that formed a monophyletic cluster with strains from Germany and the Netherlands in 2018 (sub-genogroup V).
Phylogenetic reconstruction, involving nucleotide sequences from full-length genomes CVA9 circulating in the Sverdlovsk oblast, show the formation of an internal clade with a hypothetical common ancestor among CVA9 (Figure 4C). These CVA9 strains formed a common cluster with sequences from Kazakhstan, without polyphyletic relationships with sequences from China.
Finally, EV-A71 strains were clustered with strains from the Asian-Pacific region (Figure 4D), but the estimated age of the most recent common ancestor (Time to Most Recent Common Ancestor, tMRCA) was 20 years (95% HPD 18–21) and belonged to subgroup C4.
4. Discussion
Aseptic meningitis, the most frequent infection of the central nervous system, is caused by viruses, bacteria, fungi, and parasites, with viruses being the predominant agents [21]. NPEVs are the primary viral agents responsible for this condition. Hence, rapid NPEV identification and genotyping are critical for patients with aseptic meningitis. NPEV meningitis outbreaks have been frequently reported in the Russian Federation [22].
There is little current data on NPEV circulating in the UFD and WS population with aseptic meningitis. This study describes the NPEV types detected in samples from aseptic meningitis patients (CSF, Feces, NP), as part of the National Surveillance Program from January 2022 to December 2024. A total of 281 NPEV-positive samples were received, and partial VP1 gene sequencing was attempted with 225 samples and WGS was attempted with 40 samples.
The monthly incidence shows a summer-autumn pattern, which is typically observed in temperate regions. The COVID-19 pandemic halted the spread of NPEV, likely due to stringent infection control measures [5]. As these measures relaxed, the circulation and incidence of NPEV meningitis surged in our region. In our cohort, most patients with positive NPEV were schoolchildren (7 and 17 years). Over the three-year study period, partial VP1 genotyping showed that EV-B species E30, E6, and CVA9 were the major identified types in aseptic meningitis patients. Minor genotypes recorded were CVB5, E25, and E9. EV-B species have been globally the most frequent in aseptic meningitis in Europe [23,24,25,26], in Africa [6,27,28,29], in Asia [2,8,12,30,31], in the Middle East [32,33], and in the US [5].
E30 was mainly detected in schoolchildren, which is consistent with a British study by Holmes et al. [23] but differs from previously published reports where E30 was mainly identified in adults [5,27]. E6 was the predominant type in 2024, which is in alignment with previous studies where E6 was the predominant type before the COVID-19 pandemic in 2018 and 2019 [8,30]. Also, CVA9 was reported as a major type in aseptic meningitis cases worldwide including the USA [5]. In addition, the EV-A71 strain subgenogroup C4, mainly circulating in Asia, was identified in this study. This strain is considered highly neurovirulent and accounts for 80% of severe and 93% of fatal HFMD cases in China [34]. The EV-A71 subgroup C4 was reported in several European countries including France [35] and the city of Rostov-on-Don in Russia [22]. Nevertheless, the identified EV-A71 strain has a large evolutionary distance from tMRCA. The authors interpret this result with caution because of possible selection bias during the building of alignment (only whole-genome sequences were included with its linkage to meningitis cases).
Because CSF contains low enteroviral copies, it is recommended to test and type additional respiratory and stool samples [36]. In this study, VP1 genotyping was higher in stool samples (87.5%) compared to CSF (70.6%) and pharyngeal samples (56.9%). In all cases, the NPEV genotype was identical in all tested samples, when applied. This result is in accordance with a Greek study [26]. This finding is important for clinicians as CSF quantity is usually limited for molecular analysis especially in neonates and infants.
Schoolchildren accounted for the highest number of NPEV-positive cases, making up about 60% of all infections. This finding contrasts with a study from Palestine [32], which reported the highest rates among infants under one year old.
The circulation of multiple NPEV genovariants may be affected by various factors such as viral evolution, recombination events, population bottlenecks, virulence, host immunity, and the introduction of novel types [37].
The success rate of direct NPEV typing was high (70.6%), which is significantly higher than in previous studies at 21.2% [8] and 50% [27]. The high detection rate of NPEV infections might be attributed to effective reporting practices, careful sample transportation, and the refined PCR conditions in the Nix nested-PCR protocol [38].
There are some limitations to this study: first, the number of samples is relatively small; second, the epidemic circulation patterns of different types of NPEV, with years of high incidence (e.g., E30), are followed by others with low or very low detection; third, the NGS protocol required the growth of NPEV on cell culture, which is a limiting factor as many aseptic meningitis viruses are characterized by poor growth (EV-A71) or no growth of CVA1-A6 [28].
Limited data are available regarding circulating genotypes, which are associated with aseptic meningitis in Russia and in Europe during this period [39]. E30 was the predominant NPEV genotype in our area in 2023. E30 meningitis outbreaks have been reported to occur worldwide every 3 to 5 years [12,38]. Recent E30 meningitis outbreaks occurred in Germany [25] and in Northern Europe [40].
Other similar studies reported totally different results with a predilection for the infant and adult age group [23,41]. This may be due to variation in the health-seeking behavior between different populations. Outdoor activity is probably the main reason why the male population was more infected by NPEV than the female population, with more aseptic meningitis cases in males. These results are consistent with previous reports [12].
In this study, a preliminary phylogenetic analysis of E30, CVA9, and E6 strains was performed and indicated a possible correlation with other strains circulating in Spain (E6, E30), Germany and the Netherlands (E30), the USA (E6), and Kazakhstan (CVA9), which could be attributed to migration and population movement.
A large proportion of the NPEV-positive samples originated from preschool/schoolchildren from 3 to 17 years (190 out of the 225 included patients). An interesting observation of this data is that the number of cases in the infant and adult group remained low at 6.7% and 8.9%, respectively. Within the predominant genotype, E30, E6, and CVA9 represented a surprising genetic diversity with full-length sequences like those obtained from Europe and Asia. This provides a reservoir from which novel strains may potentially emerge and perhaps as an outbreak event. Although the E30, E6, and CVA9 strains detected in this study belonged to a single genogroup within each type, they clustered closely as shown by the phylogenetic analysis.
The high mutation rate and the RNA recombination are responsible for the genetic diversity of NPEV. The recombination increases viral pathogenicity, removes lethal mutations, and increases viral fitness. Of the four NPEV species, Enterovirus exhibited the highest rates of recombination particularly between members of the same species. The continuous diversity of the VP1 coding region, which is involved in virus–cell interactions and antibody neutralization, might explain the high endemicity and infection of NPEV [32].
5. Conclusions
In conclusion, this study documents for the first time the WGS sequences of E6, CVA9, E30, and EV-A71 circulating in parts of the Russian Federation. The public health impact of enteroviruses has been highlighted by outbreaks of severe neurological disease that mimicked polio but was linked to non-polio enteroviruses. Considering this, we hypothesize that NPEVs may predominate in their ability to cause paralysis after polio is completely eradicated. Thus, investigating non-polio enteroviruses linked to neurological effects becomes essential.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tapparel C. Siegrist F. Petty T.J. Kaiser L. Picornavirus and Enterovirus Diversity with Associated Human Diseases Infect. Genet. Evol.20131428229310.1016/j.meegid.2012.10.01623201849 · doi ↗ · pubmed ↗
- 2Farshadpour F. Taherkhani R. Molecular Epidemiology of Enteroviruses and Predominance of Echovirus 30 in an Iranian Population with Aseptic Meningitis J. Neuro Virology 20212744445110.1007/s 13365-021-00973-133788142 · doi ↗ · pubmed ↗
- 3Zhou F. Kong F. Wang B. Mc Phie K. Gilbert G.L. Dwyer D.E. Molecular Characterization of Enterovirus 71 and Coxsackievirus A 16 Using the 5′ Untranslated Region and VP 1 Region J. Med. Microbiol.20116034935810.1099/jmm.0.025056-021148280 · doi ↗ · pubmed ↗
- 4Pons-Salort M. Parker E.P.K. Grassly N.C. The Epidemiology of Non-Polio Enteroviruses: Recent Advances and Outstanding Questions Curr. Opin. Infect. Dis.20152847948710.1097/QCO.000000000000018726203854 PMC 6624138 · doi ↗ · pubmed ↗
- 5Fall A. Forman M. Morris C.P. Gniazdowski V. Luo C.H. Hanlon A. Miller H. Bergman Y. Mostafa H.H. Enterovirus Characterized from Cerebrospinal Fluid in a Cohort from the Eastern United States J. Clin. Virol.202316110540110.1016/j.jcv.2023.10540136805602 · doi ↗ · pubmed ↗
- 6Nkosi N. Preiser W. Van Zyl G. Claassen M. Cronje N. Maritz J. Newman H. Mc Carthy K. Ntshoe G. Essel V. Molecular Characterisation and Epidemiology of Enterovirus-Associated Aseptic Meningitis in the Western and Eastern Cape Provinces, South Africa 2018–2019 J. Clin. Virol.202113910484510.1016/j.jcv.2021.10484533962182 · doi ↗ · pubmed ↗
- 7Shukla B. Aguilera E.A. Salazar L. Wootton S.H. Kaewpoowat Q. Hasbun R. Aseptic Meningitis in Adults and Children: Diagnostic and Management Challenges J. Clin. Virol.20179411011410.1016/j.jcv.2017.07.01628806629 PMC 5581214 · doi ↗ · pubmed ↗
- 8Wang J. Meng M. Xu H. Wang T. Liu Y. Yan H. Liu P. Qin D. Yang Q. Analysis of Enterovirus Genotypes in the Cerebrospinal Fluid of Children Associated with Aseptic Meningitis in Liaocheng, China, from 2018 to 2019 BMC Infect. Dis.20212140510.1186/s 12879-021-06112-933933008 PMC 8088645 · doi ↗ · pubmed ↗