Culture-Based Wastewater Surveillance for the Detection and Monitoring of Antimicrobial Resistance in Staphylococcal Species
Douha Shouqair, Rashed Alghafri, Mohammed Naji, Abdulla Albastaki, Rania Nassar, Lobna Mohamed, Bisola Aloba, Bayan S. Awad, Fatima Al Dhaheri, Dean Everett, Ihab Habib, Mahmood Almashadani, Ahmed A. Shibl, Jorge Rodríguez, Danesh Moradigaravand, Stefan Monecke, Ralf Ehricht

TL;DR
This study used wastewater to track antibiotic-resistant staphylococcal bacteria in Dubai, finding diverse resistant strains and highlighting wastewater as a tool for early detection of antimicrobial resistance.
Contribution
The study provides new insights into the diversity and antibiotic resistance patterns of Staphylococcus species in wastewater from the Arabian Gulf region.
Findings
Wastewater in Dubai contained diverse Staphylococcus species, including multidrug-resistant strains.
Resistance was highest to benzylpenicillin and fusidic acid, but all isolates remained susceptible to glycopeptides, tigecycline, and linezolid.
Wastewater surveillance revealed potential for tracking antimicrobial resistance across human, animal, and environmental interfaces.
Abstract
Antimicrobial resistance, when bacteria change in ways that make antibiotics no longer effective, has become one of the most serious global healthcare challenges of our time. This study examined bacteria found in wastewater in Dubai to better understand how resistant bacteria spread across the community, hospitals and the environment. The focus was on a group of bacteria called Staphylococcus, some of which can cause a wide range of infections in humans and animals, from mild skin infections to serious bloodstream diseases. Over an eight-month period, wastewater samples were collected from community locations, hospitals, and wastewater treatment plants. The bacteria were isolated and tested in the laboratory to determine which antibiotics could stop them and which ones couldn’t. We found a diverse range of Staphylococcus species, including several that showed resistance to multiple…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
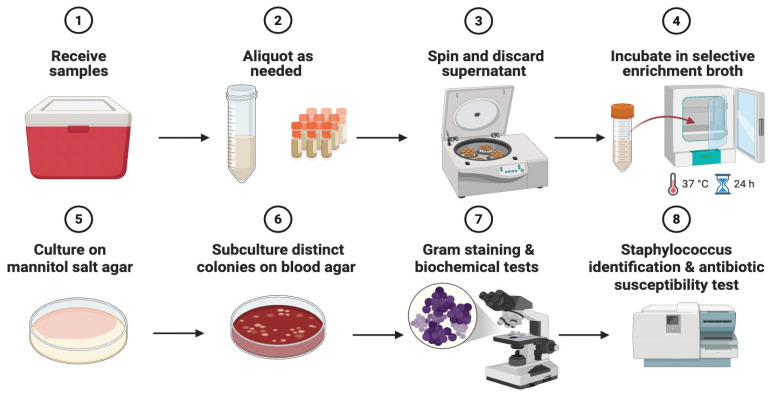 Figure 1
Figure 1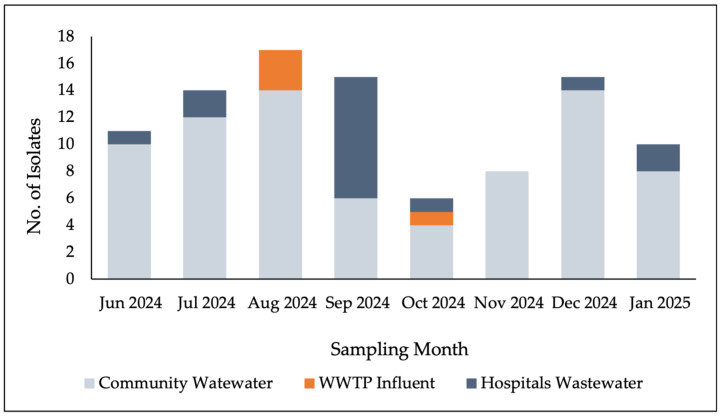 Figure 2
Figure 2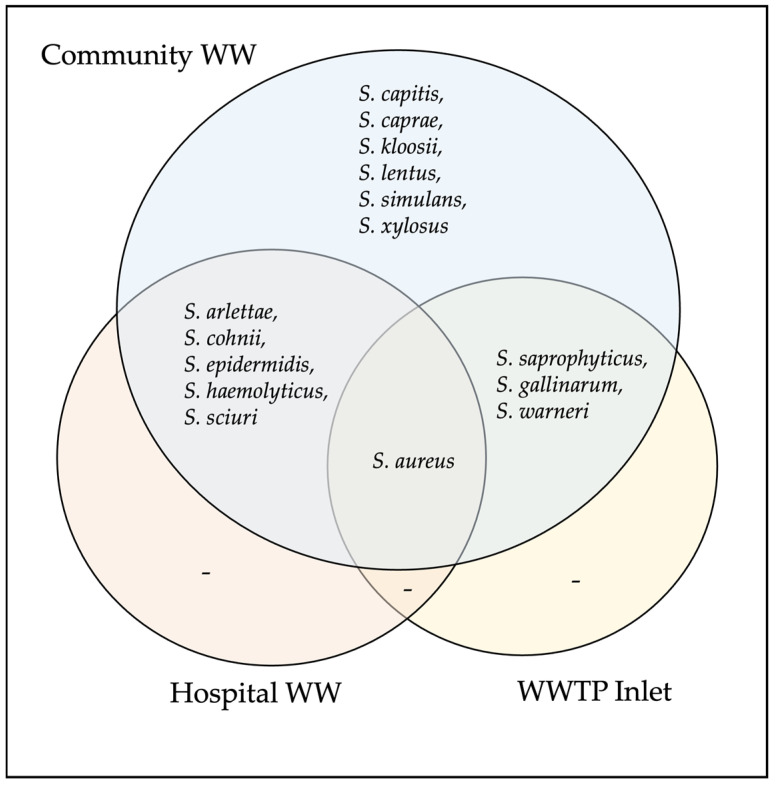 Figure 3
Figure 3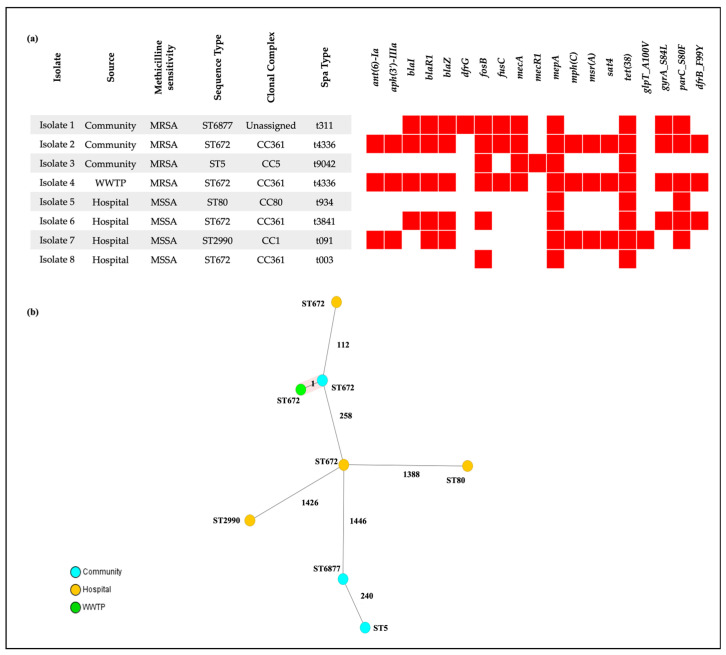 Figure 4
Figure 4- —UAE-NIH Collaborative Research Initiative
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPharmaceutical and Antibiotic Environmental Impacts · Antimicrobial Resistance in Staphylococcus · Fecal contamination and water quality
1. Introduction
Staphylococcus species are clinically significant pathogens that can cause a wide range of infections, including superficial and deep skin, soft tissue, wound [1], blood stream [2] and device related infections [3] primarily due to their high virulence and their pronounced ability to form biofilms [4]. They are increasingly recognized for their ability to acquire and disseminate antimicrobial resistance (AMR) genes [5]. To date, 94 Staphylococcus species have been identified, of which 88 are coagulase-negative Staphylococcus (CoNS) [6]. Although CoNS species such as S. epidermidis and S. haemolyticus were traditionally considered commensals, they are now increasingly being identified as drug-resistant opportunistic pathogens causing infections in humans and animals [7,8]. Coagulase-positive staphylococci (CoPS), such as S. aureus and S. intermedius, also continue to pose well established risks in the human and veterinary sectors [9]. Assessing both CoNS and CoPS in wastewater enables the investigation of these pathogens, which are of clinical relevance across human and animal sectors, as well as exposes the broader resistance-spread dynamics across shared environments. This is crucial as CoNS species, in particular, are hypothesized to function as major hubs of gene exchange and reservoirs of genetic diversity within the genus [10]. Through mobile genetic elements such as plasmids and bacteriophages, they have the potential to facilitate horizontal gene transfer, thus increasing the risk of emergence of multidrug resistance in pathogenic and non-pathogenic staphylococci.
In addition to the challenge posed to human health, CoPS and CoNS also represent important veterinary pathogens with wide-ranging impact across animal species [4,11]. They are frequently implicated in skin, wound, and intramammary infections in livestock, companion animals, and wildlife [4,11]. In cattle, S. aureus is a well-established cause of bovine mastitis, resulting in significant economic losses due to reduced milk yield and increased veterinary expenditures [12]. In horses, species such as S. xylosus and S. sciuri, are associated with skin and wound infections [13], S. lentus, S. sciuri, and S. xylosus have been isolated from camel skin and nasal flora, suggesting they may act as opportunistic pathogens in camels [14]. Domestic birds, including poultry, may harbor S. carnosus, S. xylosus, S. cohnii and S. lentus [15]. In aquaculture, S. xylosus infections in rainbow trout have been linked to high mortality and substantial economic losses [16]. In addition, other Staphylococcus species have been reported in marine animals, including S. delphini in Adélie penguins and S. pseudintermedius in Weddell seals [17]. These diverse staphylococcal species, including S. aureus, S. lentus, and S. sciuri, have also been detected in wastewater systems [18,19], highlighting their persistence in shared environments. Collectively, these observations emphasize the interconnectedness of humans, animals, and the environment, reinforcing the need for a One Health approach.
While clinical surveillance remains central to monitoring infectious pathogens, environmental sources, especially wastewater systems, are increasingly being recognized for their role as major reservoirs for resistant pathogens, AMR genes, and mobile genetic elements [20]. Wastewater integrates inputs from domestic, hospital, and livestock settings, providing a composite view of community-level microbial populations, including CoNS and CoPS, as well as their resistance profiles [21]. Within this complex microbial ecosystem, horizontal gene transfer can occur, promoting the exchange of resistance and virulence determinants among diverse bacterial species. Wastewater-based surveillance (WBS) has recently gained momentum as a practical and cost-effective tool for population-level monitoring of pathogens and AMR [22]. Initially widely utilized during the COVID-19 pandemic, WBS has since been applied to track enteric viruses, and AMR genes in various settings [23,24]. Several studies have shown that staphylococcal species can be consistently isolated from municipal, hospital, and industrial wastewater, largely due to their shedding from skin, mucosal surfaces, infected wounds, and contaminated medical waste [18,21,25,26,27,28].
Despite this increasing global interest in WBS for AMR, there remains a paucity of data from the Arabian Gulf region, including the United Arab Emirates (UAE) [29,30]. Although a recent report from the UAE documented the occurrence of staphylococcal species in wastewater [29], systematic data on the diversity and resistance profiles of staphylococcal species in wastewater from various sources, including community and hospital settings is lacking. To address this gap, our study aimed to determine the prevalence, diversity, and AMR profiles of Staphylococcus species in wastewater collected from multiple sources, including community, hospital, and wastewater treatment plant (WWTP) sites in Dubai, UAE. We employed a culture-based phenotypic approach to isolate and characterize viable Staphylococcus species, thereby supporting the development of WBS as a tool for AMR monitoring in the UAE.
2. Materials and Methods
2.1. Study Design and Sample Collection
Wastewater samples were collected once every month from June 2024 to January 2025. Samples were obtained from nine community pumping stations, two tertiary healthcare facilities and the two WWTP in the city. Wastewater samples from community sewer nodes and hospital wastewater outlets were collected using the grab sampling method [21] while WWTP influent and effluent samples were collected using 24-h composite sampling [31]. Composite samples were obtained using autosamplers (Hach, Loveland, CO, USA), where samples get automatically accumulated over 24 h in refrigerated containers. During each month, wastewater from all sampling sites was collected within a ±1-day interval to ensure temporal alignment. Between successive months, the sampling date varied by no more than ±2 days, thereby maintaining consistent temporal spacing across the study period. All samples were collected into sterile wide-mouth 1 L bottles (Azlon, Stone, Staffordshire, UK), maintained under a cold chain at 4 °C, and transported to the laboratory. Table 1 shows the sampling strategy, site distribution and frequency that was followed.
2.2. Sample Processing and Primary Isolation
Wastewater samples were received at the laboratory and processed immediately. Each wastewater bottle was mixed briefly, and a 15 mL aliquot [32] was transferred into a sterile conical tube (Corning, NY, USA). Samples were centrifuged at 7500 rpm for 10 min, and the supernatant was discarded. The resulting pellet was resuspended directly in 10 mL of Baird-Parker broth with potassium tellurite (HiMedia, Mumbai, India) and incubated on a Stuart orbital shaker (Stuart, Stone, UK) at 37 °C and 125 rpm for 24 h. After selective enrichment, aliquots were streaked onto mannitol salt agar (Medisynal, Dubai, United Arab Emirates) and incubated at 37 °C for 18–24 h. For each sample, plates were examined for macroscopically distinct colony morphotypes, defined by differences in colony form, elevation, margin, size, texture and color [33]. From each plate, up to 5 representative colonies per distinct morphotype were selected for further work. Streaking and subculturing were carried out under aseptic conditions with sterile disposable loops to minimize the risk of contamination. All morphologically distinct colonies were subcultured onto sheep blood agar (Medisynal, Dubai, United Arab Emirates) and incubated at 37 °C for 18–24 h to promote growth. Plates were then inspected for uniform colony morphology and hemolysis patterns; cultures showing mixed growth were re-streaked until pure.
Representative colonies from each distinct type were subjected to Gram staining and examined microscopically for the presence of Gram-positive cocci (Figure 1). Isolates consistent with staphylococcal species were subjected to biochemical testing (Figure 1), including catalase and coagulase assays. Isolates were preserved in tryptic soy broth (TSB) (Millipore, Darmstadt, Germany) containing 25% glycerol and stored at −80 °C for subsequent analyses.
2.3. Species-Level Identification
Presumptive isolates were further confirmed and identified to the species level using the VITEK^®^ 2 compact automated system with Gram Positive identification (ID-GP) cards (bioMérieux, Marcy L’Etoile, France). Bacteria were first cultured on blood agar plates, and fresh overnight growth was used for identification. Pure colonies were suspended in sterile saline to a standardized turbidity of 0.5–0.6 McFarland on DensiCHEK Plus (bioMérieux, Marcy L’Etoile, France) and loaded into the GP cards, which contains a panel of biochemical assays such as carbohydrate utilization and enzyme activities (e.g., urease, arginine dihydrolase). The VITEK^®^ 2 system automatically incubated and monitored metabolic reactions, after which species-level identification was determined.
2.4. Antimicrobial Susceptibility Testing (AST)
Bacterial identification, confirmation of MRSA and phenotypic antimicrobial susceptibility testing were performed using the VITEK^®^ 2 Compact automated system with the AST-P592 cards (bioMérieux, France), in accordance with manufacturer-provided protocols and Clinical and Laboratory Standards Institute (CLSI) [34]. Antibiotic susceptibility testing with VITEK^®^ 2 AST cards is based on the broth microdilution; the system automatically incubates and monitors growth in the presence of antimicrobials and uses the Advanced Expert System (AES) to generate MIC values and categorical interpretations. Categorical interpretations were assigned according to the CLSI Performance Standards for Antimicrobial Susceptibility Testing [34]. Multidrug resistance (MDR) was defined as non-susceptibility to at least one agent in three or more classes of antibiotics [35].
2.5. Whole Genome Sequencing (WGS)
This was carried out at CosmosID Laboratories (Germantown, MD, USA). Briefly, genomic DNA was extracted from all S. aureus wastewater isolates using the DNeasy PowerSoil Pro Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions, and DNA was quantified using a Qubit Flex fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). Libraries were prepared with the Watchmaker DNA Library Prep Kit. Sequencing was performed on the Element AVITI platform using 2 × 150 bp paired-end chemistry (Cloudbreak kit; Element Biosciences, San Diego, CA, USA). WGS data are available on NCBI BioProject ID: 1376508.
2.6. Bioinformatics and Genomic Analyses
Whole-genome sequencing data from all S. aureus isolates were processed using the Bactopia v3.2.0 pipeline [36,37]. Raw paired-end reads underwent quality control and trimming, followed by de novo assembly. Molecular typing included MLST using the S. aureus PubMLST scheme, spa typing, SCCmec typing and core-genome MLST to infer genomic relatedness using RIDOM SeqSphere+ v.10.0.5 EULA (Ridom GmbH, Muenster, Germany), applying a ≤24-allele difference threshold for closely related isolates. AMR genes were identified with AMRFinderPlus v.4.0.19 and Abritamr v.1.0.19.
3. Results
3.1. Detection of Staphylococcal Species
A total of 96 staphylococcal isolates were recovered from three wastewater sources: hospital wastewater, community wastewater, and WWTP influent. The number of captured isolates per month ranged from 6 to 17, with no statistically significant difference across months (p = 0.28). Of these 91.7% (n = 88) were coagulase-negative staphylococci (CoNS) and 8.3% (n = 8) were coagulase-positive S. aureus (CoPS), yielding a CoNS:CoPS ratio of approximately 11:1. No staphylococci were detected in WWTP effluent samples. The highest number of isolates were obtained from community wastewater (n = 67), followed by hospital wastewater (n = 16), while only 4 isolates were recovered from WWTP influent (Figure 2).
The 15 Staphylococcus species were S. aureus, S. arlettae, S. capitis, S. caprae, S. cohnii, S. epidermidis, S. gallinarum, S. haemolyticus, S. kloosii, S. lentus (now Mammaliicoccus lentus), S. saprophyticus, S. sciuri (now Mammaliicoccus sciuri), S. simulans, S. warneri, and S. xylosus. The most frequently identified species were S. cohnii (n = 15), S. saprophyticus (n = 16), S. sciuri (n = 13), S. haemolyticus (n = 9), and S. xylosus (n = 9). The distribution of Staphylococcus species across wastewater sources is shown in Figure 3.
3.2. Antimicrobial Susceptibility
The majority of the isolates, 88% (n/N = 84/96) and 82% (n/N = 79/96), were resistant to benzylpenicillin and fusidic acid, respectively. Fifty-four isolates were resistant to erythromycin, while resistance to clindamycin and folate inhibitors (trimethoprim/sulfamethoxazole) was detected in 22 and 18 isolates, respectively. All isolates were susceptible to glycopeptides (vancomycin and teicoplanin), as well as to tigecycline and linezolid.
The antibiotic resistance profile of the tested Staphylococcus species varied across isolates. On average, each isolate exhibited resistance to approximately four out of the 16 antibiotics on the VITEK panel. However, S. capitis, S. epidermidis and S. simulans isolates showed resistance to only benzylpenicillin. Detailed antibiotic resistance profile for each isolate is shown in Supplementary Table. Resistance to 3 or more classes of antibiotics indicative of MDR phenotype [35] was observed in the majority of staphylococci species identified (n/N = 11/15). Table 2 shows the distribution of MDR isolates across identified species.
Overall, 60% of the CoNS isolates exhibited an MDR phenotype (n = 53), predominantly represented by S. cohnii (n = 13) and S. saprophyticus (n = 11). As shown in Table 2, MDR isolates were distributed across multiple Staphylococcus species, with 33 identified as methicillin-resistant CoNS (MR-CoNS).
Phenotypic methicillin resistance was detected in 33 CoNS and 4 CoPS. The methicillin resistant CoNS include S. arlettae, S. caprae, S. cohnii, S. epidermidis, S. haemolyticus, S. saprophyticus, S. sciuri and S. warneri isolates. These were detected from community WW, WWTP inlet and hospital WW. Of the 8 S. aureus isolates detected, 4 (50%) were methicillin-resistant S. aureus (MRSA). These MRSA isolates were from WWTP influent (n = 1) and community wastewater (n = 3).
3.3. Whole-Genome Sequencing of S. aureus Isolates
Whole-genome sequencing of the 8 S. aureus isolates revealed that the dominant sequence type (ST) was ST672 (n/N = 4/8; 50%) with the STs identified in MRSA being ST672 (n = 2), ST5 (n = 1), and ST6877 (n = 1). MSSA had isolate each of ST80 and ST2990, and two isolates were ST672. We identified 7 spa types with t4336 associated with ST672 detected in two isolates and the remaining spa types were each represented by a single isolate (Figure 4a). Two MRSA isolates (ST672 and ST6877) harboured SCCmec type V, while the ST5 isolate carried type IV. Phylogenomic analysis showed that the isolates were genetically diverse (Figure 4b). In the cgMLST scheme, all isolate pairs differed by more than 24 alleles, consistent with substantial genomic distance. A single exception was observed: one WWTP influent derived MRSA isolate differed by only one allele from a community derived MRSA isolate (Figure 4b). The MRSA isolates haboured mecA gene and largely exhibited a larger repertoire of resistance genes compared to MSSA isolates although the tet(38) and mepA resistance were universally detected across all isolates (Figure 4a).
4. Discussion
This study applied a culture-based approach to investigate antimicrobial resistance in Staphylococcus isolated from multiple wastewater sources. Sampling locations were selected to reflect diverse urban activities, including community and hospital wastewater, as well as WWTP influent and effluent, thereby allowing for a comprehensive assessment of isolates from wastewater sources. Our findings align with global reports that demonstrate the occurrence of diverse CoNS and CoPS in various wastewater sources [5,19,21,26,27]. The identification of opportunistic pathogens such as S. haemolyticus and S. cohnii, alongside the widespread presence of multidrug-resistant isolates, particularly those exhibiting β-lactam and fusidic acid resistance phenotypes, is noteworthy [38,39]. The predominance of these environmental staphylococci, particularly the widespread detection of CoNS species across multiple wastewater sources, suggests substantial ecological resilience. Moreover, the high levels of resistance to major antibiotic classes may reflect selective pressures exerted by residual antibiotics, disinfectants, and heavy metals within wastewater systems [40,41]. Furthermore, several species recovered from wastewater are known to colonize livestock, companion animals, and wildlife [11].
The consistent presence of animal-associated staphylococci, including S. sciuri and S. cohnii, which were the predominant CoNS identified, suggests that animal-derived contributions influence the wastewater microbiome in our setting. Such overlap between human and animal staphylococcal populations highlights the possibility of interspecies transmission. Wastewater thus provides a One Health interface where microbial and resistance determinants from multiple sectors intersect and where horizontal gene transfer is likely to occur [42,43]. Notably, S. sciuri, which is generally considered a non-pathogenic environmental species, has recently been shown to predominate in the farm environment as well as readily adapt to and persist in health care settings [44]. Additionally, S. sciuri harboring the mecA gene and resistance genes for lincosamide and streptogramin [44], have been reported in the literature, suggesting their potential to serve as reservoirs of AMR genes that may be transferred to pathogenic staphylococci, including S. aureus. In this study, there was high occurrence of S. sciuri, including isolates that showed methicillin resistance and MDR phenotype. Further work to investigate their genomic profiles, characterize associated AMR genes, and assess potential horizontal gene transfer events is warranted.
The findings demonstrate widespread antibiotic resistance among the staphylococci isolates identified, with the majority (82%) exhibiting resistance to fusidic acid, particularly among CoNS, such as S. haemolyticus, S. cohnii, and S. saprophyticus. These findings provide the first insight into the occurrence of fusidic acid resistance among CoNS and non-clinical CoPS isolates from our setting. In addition, whilst fusidic acid resistance predominated in isolates from community wastewater samples, β-lactam resistance was more frequently observed in isolates from hospital wastewater. This distribution likely reflects the differential antibiotic usage patterns, with β-lactams being more commonly administered in clinical settings compared to topical fusidic acid in community settings. Previous reports from the UAE and the broader Arabian Gulf region have documented a high prevalence of fusidic acid resistance in clinical S. aureus isolates, which often carry the SCCmec+SCCfusC composite resistance genes. Further work is needed to determine the molecular makeup of these environmental CoNS with fusidic acid resistance and explore relationships with clinical S. aureus isolates harboring SCCmec+SCCfus composite genes.
Interestingly, the sole CoPS identified in our study was S. aureus, and it was the only species found across all three wastewater sources. Notably, MRSA has been commonly associated with hospital wastewater in reported studies [25,45]. Thus, the detection of MSSA in hospital wastewater, with MRSA being found in community wastewater and WWTP influent, differs from the patterns reported in the literature [25,46]. This detection of multidrug-resistant MRSA in community wastewater and WWTP influent suggests that highly pathogenic strains are circulating more broadly in our setting. Such divergence from reported literature may reflect variations in wastewater inputs, or differences in AMR epidemiology such as the growing prevalence of community-associated MRSA lineages in our setting [47,48]. This underscores the value of regionally tailored WBS studies, as reliance on global patterns risks overlooking locally relevant dynamics.
Molecular characterization of food and clinical S. aureus isolates from the UAE has revealed a predominance of community-associated MRSA lineages [47,49], although WGS data for wastewater-derived isolates remain limited. The eight S. aureus genomes analysed here therefore provide new insight into this environmental reservoir, demonstrating the presence of genetically distinct MRSA and MSSA isolates. In the cgMLST analysis, all isolate pairs differed by more than 24 alleles, with the exception of two ST672 isolates, indicating an overall lack of close relatedness.
ST672 belongs to CC361, thus making CC361, which has been described as an emerging community-associated lineage [50,51] the predominant CC in this study. Its detection in retail meat products [52], alongside reports from cattle and monkeys [53], and the identification of ST672 in fish and fishery environments [54] have all been documented in the literature, underscoring its relevance across multiple ecosystems. Resistance determinants associated with reduced susceptibility to tetracyclines and fluoroquinolones were present in all isolates, raising concern for treatment options should animals or humans be exposed to such strains. Furthermore, the identification of plasmid replicons rep16 and rep5a suggests a potential for horizontal gene exchange by these isolates.
In the UAE, treated wastewater effluent from WWTPs is commonly reused for irrigation due to limited freshwater resources. Staphylococci were not detected in any WWTP effluent, which suggests that the wastewater treatment likely removed viable cells, although the presence of viable but non-culturable Staphylococci in WWTP effluent remains a possibility. In addition, staphylococci or their associated AMR determinants may have partitioned into the sludge fraction, which was not assessed in this study. As sewage sludge is frequently applied to agricultural land as fertilizer, this represents a potential transmission route that merits further investigation. Moreover, resistance genes may persist in treated effluent and, when reused for irrigation, could re-enter the environment, expanding the reservoir of resistant staphylococci, promoting horizontal gene transfer to other bacteria, and increasing the potential for human and animal exposure through contaminated soil, water, or agricultural produce. Future work employing shotgun metagenomic sequencing is recommended to identify and determine the abundance of AMR genes and mobile genetic elements in WWTP effluent.
Culture-based approaches remain a critical component of wastewater surveillance, enabling the isolation of viable bacteria for antimicrobial susceptibility testing and thereby revealing resistant phenotypes of clinical and veterinary relevance. As wastewater contains a complex and often low-abundance microbial community, culture methods also facilitate the enrichment of rare organisms, including staphylococcal species that may be underrepresented or missed entirely by metagenomic analyses. Importantly, cultured isolates provide isolates for downstream bacterial whole-genome sequencing work. However, applying culture-based methods to wastewater is inherently challenging. Competition from diverse microbial populations and exposure to environmental stressors, such as residual antibiotics, biocides, toxic compounds, and lytic phages, can inhibit the growth of more fastidious or low-abundance taxa [43,55,56,57]. In addition, some staphylococcal species may enter a viable but non-culturable state, in which cells remain metabolically active and retain their resistance determinants but fail to grow on routine laboratory media. Differences in isolate recovery between community sites and WWTP samples suggest that environmental pressures in centralized systems may impact culturability, underscoring the value of sampling across multiple wastewater sources.
5. Conclusions
In conclusion, this study demonstrates the utility of culture-based methods for detecting viable staphylococci across multiple wastewater sources. While wastewater treatment reduces the prevalence of staphylococci, the consistent detection of MDR isolates in upstream sources underscores the role of wastewater as a reservoir and potential convergence for both human- and animal-associated microbes. Within a One Health framework, these findings highlight cross-sector transmission risks and provide a foundation for implementing AMR WBS to protect human and animal health.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mariani F. Galvan E.M. Staphylococcus aureus in Polymicrobial Skinand Soft Tissue Infections: Impact of Inter-Species Interactionsin Disease Outcome Antibiotics 202312116410.3390/antibiotics 1207116437508260 PMC 10376372 · doi ↗ · pubmed ↗
- 2Serra N. Di Carlo P. Andriolo M. Mazzola G. Diprima E. Rea T. Anastasia A. Fasciana T.M.A. Pipito L. Capra G. Staphylococcus aureus and Coagulase-Negative Staphylococci from Bloodstream Infections: Frequency of Occurrence and Antimicrobial Resistance, 2018–2021 Life 202313135610.3390/life 1306135637374138 PMC 10305610 · doi ↗ · pubmed ↗
- 3Albavera-Gutierrez R.R. Espinosa-Ramos M.A. Rebolledo-Bello E. Paredes-Herrera F.J. Carballo-Lucero D. Valencia-Ledezma O.E. Castro-Fuentes C.A. Prevalence of Staphylococcus aureus Infections in the Implantation of Orthopedic Devices in a Third-Level Hospital: An Observational Cohort Study Pathogens 20241362010.3390/pathogens 1308062039204221 PMC 11357249 · doi ↗ · pubmed ↗
- 4Silva-Santana G. Staphylococcus aureus: Dynamics of pathogenicity and antimicrobial-resistance in hospital and community environments—Comprehensive overview Res. Microbiol.202517610426710.1016/j.resmic.2025.10426739805330 · doi ↗ · pubmed ↗
- 5Adesoji T.O. Somda N.S. Tetteh-Quarcoo P. Shittu A.O. Donkor E.S. Prevalence of methicillin-resistant coagulase-negative staphylococci in Africa: A systematic review and meta-analysis BMC Infect. Dis.20252590610.1186/s 12879-025-11149-140640717 PMC 12243303 · doi ↗ · pubmed ↗
- 6Leibniz Institute DSMZ List of Prokaryotic Names with Standing in Nomenclature (LPSN). Genus Staphylococcus Available online: https://lpsn.dsmz.de/search?word=staphylococcus(accessed on 1 October 2025)
- 7Wildsmith C. Barratt S. Kerridge F. Thomas J. Negus D. Genomic and phenotypic characterization of staphylococci isolated from the skin of non-human primates Microbiology 202517100154610.1099/mic.0.00154640131783 PMC 11936348 · doi ↗ · pubmed ↗
- 8Argemi X. Hansmann Y. Prola K. Prevost G. Coagulase-Negative Staphylococci Pathogenomics Int. J. Mol. Sci.201920121510.3390/ijms 2005121530862021 PMC 6429511 · doi ↗ · pubmed ↗