Unveiling Intestinal Emphysema in Pigs: Morphological Insights and Pathogenetic Implications
Alfonso Rosamilia, Simona Baghini, Chiara Guarnieri, Anastasia Romano, Umberto Tosi, Giuseppe Marruchella, Attilio Corradi

TL;DR
This study investigates intestinal emphysema in pigs, revealing that gas-filled cysts are linked to lymphatic issues and granulomas, similar to human cases.
Contribution
The study provides new morphological evidence linking porcine intestinal emphysema to lymphatic-centered pathology and granuloma formation.
Findings
Gas-filled cysts in pigs are mainly located in the submucosal and mesenteric layers.
Cysts are partially lined by lymphatic endothelial cells and associated with granulomas.
The findings suggest a lymphatic origin for the lesions, similar to human intestinal emphysema.
Abstract
Intestinal emphysema is characterized by gas-filled cysts within the intestinal wall, and it is occasionally observed in slaughtered pigs. The etiology and pathogenesis of intestinal emphysema remain poorly understood. This study aimed to provide further morphological insights into porcine intestinal emphysema through histopathological, histochemical and immunohistochemical methods. Ten slaughtered heavy pigs were examined, showing gross lesions consistent with intestinal emphysema. Gaseous cysts were mainly located in the submucosal and mesenteric layers, at least partially lined by lymphatic endothelial cells, and almost invariably associated with granulomas. Overall, data suggests that porcine intestinal emphysema is a lymphatic-centered disorder of the intestinal wall and mesentery, showing pathological features very similar to those described in human medicine. Intestinal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
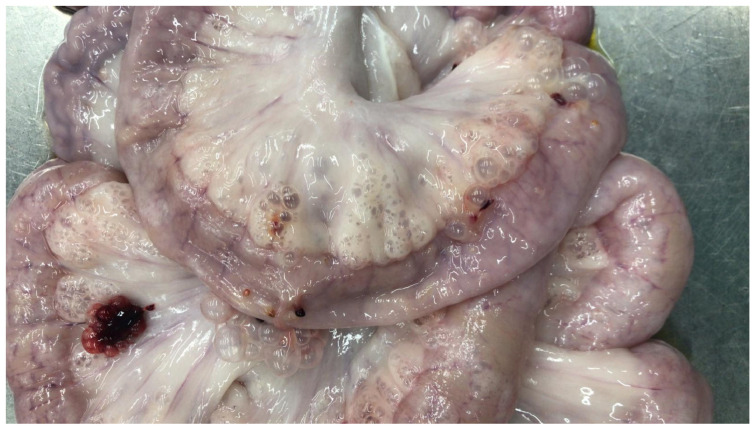 Figure 1
Figure 1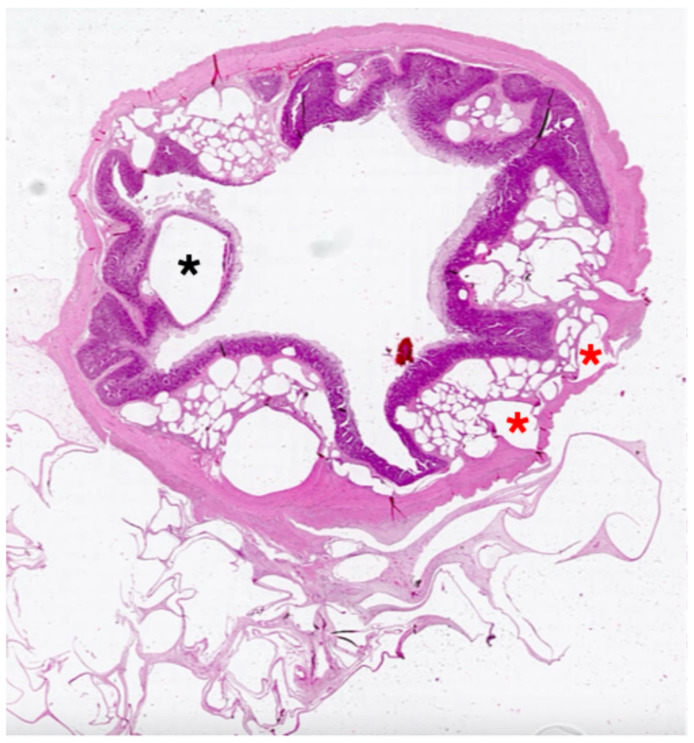 Figure 2
Figure 2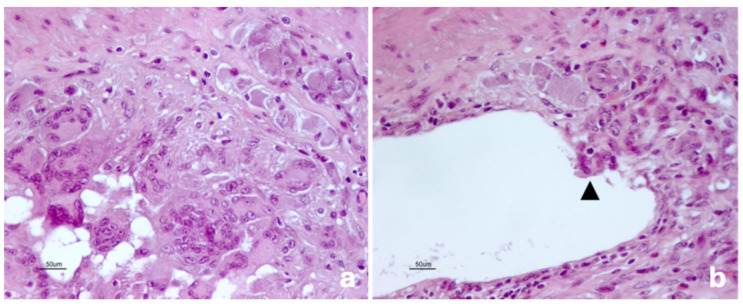 Figure 3
Figure 3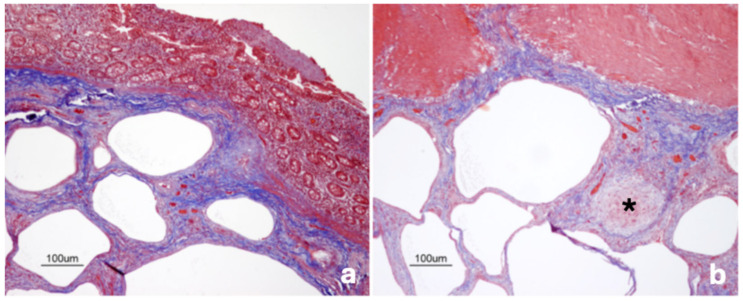 Figure 4
Figure 4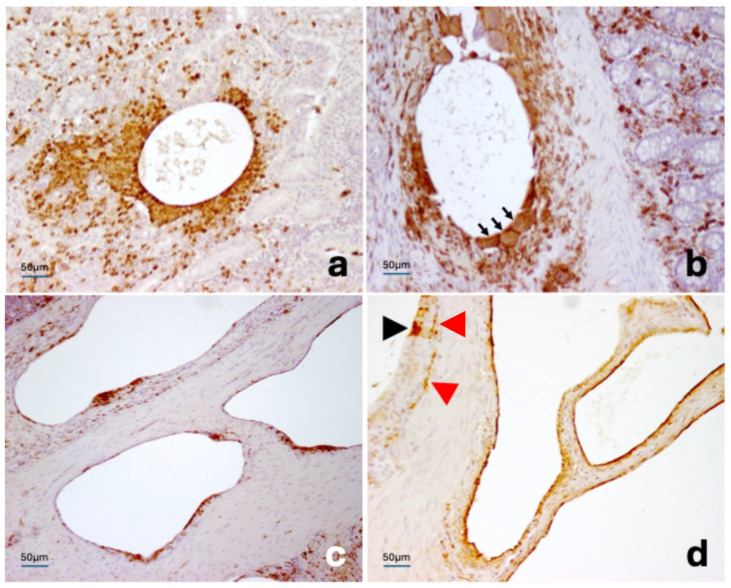 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumothorax, Barotrauma, Emphysema · Pleural and Pulmonary Diseases · Abdominal Surgery and Complications
1. Introduction
Intestinal emphysema (IE) is a disease condition observed in humans and animals, characterized by clusters of gas-filled cysts within the intestinal wall. Over time, this entity has been described using various terms—e.g., pneumatosis cystoides intestinalis, mesenteric emphysema, intestinal pneumatosis, and cystic lymphopneumatosis—reflecting its distinctive morphological features [1].
Intestinal emphysema primarily affects the small intestine and typically arises along the mesenteric border. However, IE may extend transmurally, involve the large intestine, and reach the mesenteric lymph nodes. Microscopically, IE lesions consist of variably sized, empty cystic spaces, which are located predominantly in the submucosa and mesentery, often surrounded by epithelioid macrophages and multinucleated giant cells [2,3,4].
In humans, IE is diagnosed incidentally in asymptomatic individuals or in patients presenting with nonspecific gastrointestinal signs [2,5,6,7,8]. In pigs, IE is occasionally detected at slaughter and appears to have no impact on public health, food safety, carcass suitability for human consumption, or farm profitability [4,9]. A recent survey reported prevalence values ranging from 1.25% to 5.12% in different slaughter batches, suggesting that porcine IE is more common than generally assumed [10]. Even much higher prevalence values have been rarely described under unusual environmental conditions [9].
Despite investigations, the etiology and pathogenesis of IE remain unclear. Several non-mutually exclusive hypotheses have been proposed, which may operate synergistically:
- (a)Pulmonary theory—Chronic obstructive pulmonary disease (COPD) increases alveolar gas pressure, leading to rupture of the alveolar walls and septa, with air dissecting through the pulmonary interstitium into the mediastinum, the retroperitoneum, and ultimately the mesentery and the intestinal wall [8,11].
- (b)Intestinal–mechanical theory—Increased intraluminal pressure and/or mucosal injury may allow gas to penetrate the intestinal wall, as observed in intestinal obstruction, hyperperistalsis, trauma, endoscopic insufflation, or surgical manipulation. Disruption of mucosal integrity may facilitate the passage of luminal gas into the submucosa or mesentery [5,12,13].
- (c)Bacterial theory—Bacterial pathogens may contribute by damaging the intestinal mucosa and/or producing gas that accumulates within the intestinal wall. Experimental reproduction of IE in rats infected with Clostridium perfringens and in gnotobiotic pigs infected with Escherichia coli supports this hypothesis [14,15,16].
- (d)Chemical–pharmacological theory—Administration of α-glucosidase inhibitors, immunosuppressive or chemotherapeutic agents, as well as occupational exposure to trichloroethylene, may enhance intraluminal gas production or alter mucosal permeability, thereby contributing to the development of IE [17].
This study aims to provide additional insights into the microscopic features of porcine IE, thus hopefully contributing to understand IE pathogenesis.
2. Materials and Methods
Ten heavy pigs (approximate slaughter weight of 160 kg, average age of 9–10 months), slaughtered at a commercial abattoir in Northern Italy and exhibiting gross IE lesions were included in this study (Figure 1). These pigs had been enrolled in a previous investigation [10], showed no relevant clinical sign at the antemortem inspection, and were carefully evaluated at postmortem inspection being admitted for human consumption in accordance with Regulation (UE) 2019/627. In particular, the gastrointestinal tract and pulmonary parenchyma showed no gross evidence of inflammatory lesions.
From each pig, a full-thickness, 2-cm-wide segment of the terminal ileum was collected, including the mesenteric border and the adjacent mesentery. Samples were immediately fixed in 10% neutral-buffered formalin, embedded in paraffin and routinely processed for histopathological evaluation (hematoxylin and eosin staining, H&E). An additional tissue section was subjected to Masson’s trichrome staining (Bio-Optica, Milan, Italy) to visualize fibrous connective tissue.
Given that biomolecular analyses performed on the same samples yielded inconclusive results and did not identify bacterial agents associated with granulomatous inflammation or necrotizing enteritis [10], additional histochemical investigations (e.g., Ziehl–Neelsen and Warthin–Starry staining) were not considered essential within the scope of this study.
Immunohistochemical analyses were performed to identify the cell types lining the cyst surface: lymphatic endothelial cells, macrophages, epithelial cells, and mesenchymal cells (see Table 1 for details). For this purpose, 4-µm-thick sections were mounted on positively charged slices (Bio-Optica, Milan, Italy), deparaffinized in xylene, and rehydrated through a graded ethanol series to distilled water. Heat-induced epitope retrieval was carried out in 0.01 M citrate buffer (pH 6.0) using a microwave oven at 750 W for 3 cycles of 5 min. Tissue sections were then cooled to room temperature and incubated for 20 min with a blocking solution (BLOXALL, Vector Laboratories, Newark, CA, USA) to inhibit endogenous peroxidase activity. Subsequently, the sections were incubated for 30 min with 2.5% normal horse serum (Vector Laboratories, Newark, CA, USA), followed by overnight incubation at 4 °C with the primary antibodies at their optimized working dilutions (Table 1). Immunoreactivity was visualized using a commercially available kit (ImmPRESS^®^ Polymer Detection Kit; Vector Laboratories, Newark, CA, USA) according to the manufacturer’s instructions, and the sections were finally counterstained with Mayer’s hematoxylin (Bio-Optica, Milan, Italy). Negative controls were included in each immunohistochemical run by omitting the primary antibody.
Histological and immunohistochemical slides were examined under a light microscope (Nikon Eclipse E800, Tokyo, Japan) at magnifications ranging from ×40 to ×400. Micrographs were captured using the DXM1200 digital camera (Nikon, Tokyo, Japan).
3. Results
A few cysts (2–15 per section) were observed within the mucosal layer of three pigs. Submucosal involvement was consistent and characterized by numerous cysts (>50 per section), some of which were large and caused atrophy of the adjacent tissues (Figure 2). The cyst luminal surface was lined by heterogeneous cellular populations—i.e., endothelium-like cells, epithelioid cells, and multinucleated giant cells—which frequently co-occurred within the same lesion (Figure 3a,b). Granulomas and lymphocytic aggregates were common within the submucosa and inside the connective septa separating the cystic structures.
Gas-filled cysts were less frequently observed within the tunica muscularis. The mesentery represented the most severely affected anatomical district, containing an extremely high, often countless burden of cysts. Microscopic features of cysts resembled those observed within the submucosa.
Masson’s trichrome staining showed that septa delineating the cysts—particularly those in the submucosa and mesentery—mainly consisted of dense collagenous stroma. In these same layers (i.e., submucosa and mesentery), a prominent fibrotic reaction also surrounded granulomas enriched in multinucleated giant cells (Figure 4a,b).
Immunohistochemical investigations pointed out that the cysts were variably lined by lymphatic endothelial cells (LYVE-1–immunoreactive), macrophages (immunoreactive for vimentin, MAC387, and IBA1/AIF1), and multinucleated giant cells (immunoreactive for vimentin and IBA1/AIF1, with inconsistent or weak MAC387-immunolabeling). Aggregates of MAC387-positive cells were observed more frequently within the lamina propria, where they tended to cluster around the cysts. LYVE-1-positive macrophages cells were also noted on occasion. In addition, a layer of epithelioid cells was sometimes present between the lymphatic endothelium and the cystic lumen (Figure 5a–d). Intestinal epithelial cells and mesothelial cells showed consistent and intense immunolabeling for cytokeratin. Mesothelial cells were infrequently observed in direct apposition to the surfaces of gas-filled cysts, apparently contributing to the architecture of the cyst wall.
4. Discussion
This study does not resolve the enigmatic nature of IE, as its etiology and pathogenesis remain largely obscure. In addition, some inherent limitations should be acknowledged, primarily related to the limited sample size and the features of the samples, which likely reflect the end stage of a long, complex, and multifactorial process. Nonetheless, it provides a detailed morphological description of porcine IE, allowing some plausible considerations.
Histopathological and immunohistochemical findings support the hypothesis that gas-filled cysts originate from lymphatic vessels. This interpretation is consistent with the topographical distribution of lesions, which mainly affect the submucosa and mesentery, anatomical compartments rich in lymphatic vessels. Moreover, it is further corroborated by the identification of LYVE-1-positive endothelial cells lining the inner surface of the cysts. These findings align closely with those reported in humans by Gui et al. [1], who concluded that “the lymphatic theory is most likely valid… and the lymphatic vessels represent the common pathway for gas transport in various conditions in which the integrity of the intestinal mucosa is compromised”.
Overall, the immunohistochemical patterns of porcine IE mirrored those described in humans [1], even though different markers were employed. Lymphatic endothelial cells inconsistently and/or partially lined the cystic lumina and were frequently replaced by MAC387 and/or IBA1/AIF1-positive cells. Epithelioid macrophages and multinucleated giant cells were often interposed between LYVE-1-positive endothelial cells and the cystic cavity, and occasionally exhibited LYVE-1 immunoreactivity themselves. In this respect, Gui et al. [1], hypothesized that gaseous dilation may damage lymphatic endothelial cells, thereby triggering a chronic inflammatory response. Macrophages may subsequently accumulate around the lymphatics and phagocytose injured endothelial cells, becoming transiently immunoreactive for lymphatic markers (e.g., LYVE-1). We consider this mechanism plausible in humans and reasonably applicable to porcine IE as well.
In our opinion, the “pulmonary theory”—primarily formulated in humans—is unlikely to account for IE in pigs, as the pathogenesis of porcine chronic bronchopneumonia (e.g., caused by Mycoplasma hyopneumoniae) is entirely distinct from that of human COPD, and it does not involve marked emphysematous change [8,11,20]. Similarly, other hypotheses—e.g., post-surgery occurrence of IE or chemical–pharmacological theory—are conceivable in humans or companion animals [12,13,17], but they cannot reasonably be extended to reared pigs.
Escherichia coli has been shown to induce IE in gnotobiotic pigs [14], and this finding supports the “bacterial theory” as a plausible explanation for porcine IE. Nevertheless, in-depth analyses of the gut microbiota have failed to identify a consistent bacterial pathogen or dysbiosis signature in IE-affected pigs. Speculatively, non-mutually exclusive explanations may account for this observation. For instance, a bacterial pathogen may act in a “hit-and-run” manner, being no longer detectable at the time of slaughter [10]. Alternatively, the mucosal integrity may be compromised by multiple pathogens and/or by additional concurrent factors. In this context, we have recently experienced cases of colonic IE in growing pigs with swine dysentery; these lesions likely result from Brachyspira hyodysenteriae-induced necrosis, they lack granulomatous reactions and may represent an early stage of IE (unpublished data). The potential association between IE and specific intestinal diseases, particularly those characterized by necrotizing lesions (e.g., swine dysentery, proliferative ileitis, salmonellosis), warrants further investigation. If substantiated, IE could be regarded as a sentinel lesion, contributing to offer valuable feedback for herd health monitoring.
Notably, cysts are often surrounded by MAC387-immunoreactive, indicative of recent recruitment [21]. This finding is particularly evident within the mucosal layer, suggesting that IE is a long-standing inflammatory and fibroproliferative process, which continues to progress over time and possibly originates at the mucosal interface.
In conclusion, this study supports the view that porcine IE is a chronic, lymphatic-centered disorder. Considering its pathological features and according to the literature [1], the following scenario may be envisaged: (1) a transient, yet unidentified, insult perturbs the lymphatic function or integrity within the gut wall; (2) impaired lymphatic drainage or structural damage of lymphatic vessels predisposes to the accumulation of gas; (3) gaseous distension further damages the lymphatic endothelium, triggering macrophages recruitment and granuloma formation; (4) persistent, low-grade inflammation leads to fibrosis, partial replacement of the lymphatic endothelium by macrophages, and ultimately the formation of stable gas-filled cysts. The parallels between porcine and human IE suggest shared pathogenetic mechanisms across species and underscore the value of pigs as a comparative model. Deeper and prospective investigations of potential risk factors (e.g., dietary components, microbiota composition, genetic background) are warranted for elucidating the etiology and pathogenesis of IE. Finally, additional insights may be gained from the analysis of the gas composition within the cysts, an approach that is already planned for future investigations. Comparative evaluation of gas composition data with those available from human pathology [22] may contribute to a better understanding of the pathogenesis of porcine IE and further clarify the extent to which it mirrors the human condition.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gui X. Zhou Y. Eidus L. Falck V. Gao Z.H. Qin L. Is pneumatosis cystoides intestinalis gas-distended and ruptured lymphatics? Reappraisal by immunohistochemistry Arch. Pathol. Lab. Med.20141381059106610.5858/arpa.2013-0145-OA 25076294 · doi ↗ · pubmed ↗
- 2Azzaroli F. Turco L. Ceroni L. Galloni S.S. Buonfiglioli F. Calvanese C. Mazzella G. Pneumatosis cystoides intestinalis World J. Gastroenterol.2011174932493610.3748/wjg.v 17.i 44.493222171137 PMC 3235639 · doi ↗ · pubmed ↗
- 3Tharmaradinam S. Kanthan S. Kanthan R. Pneumatosis cystoides intestinalis and hyperganglionosis—Cause or effect? A review Pathol. Res. Pract.202021615287910.1016/j.prp.2020.15287932089409 · doi ↗ · pubmed ↗
- 4Marcato P.S. Patologia Sistematica Veterinaria 1st ed.Edagricole Bologna, Italy 2002678
- 5Voboril R. Pneumatosis cystoides intestinalis—A review Acta Medica 200144899210.14712/18059694.2019.9011811082 · doi ↗ · pubmed ↗
- 6Sánchez-Rodríguez J.J. Utrillas-Martínez A.C. Antón-Beranoaguirre J.S. Moreno-Olivieri A.A. Del Val-Gil J.M. Pneumatosis cystoides intestinalis: A case report Cir. Cir.20188637037310.24875/CIRUE.M 1800005430067713 · doi ↗ · pubmed ↗
- 7Sugihara Y. Harada K. Ogawa H. Otsuka F. Okada H. Pneumatosis Cystoides Intestinalis J. Lung Health Dis.20182242710.29245/2689-999X/2017/1.1121 · doi ↗
- 8Di Pietropaolo M. Trinci M. Giangregorio C. Galluzzo M. Miele V. Pneumatosis cystoides intestinalis: Case report and review of literature Clin. J. Gastroenterol.202013313610.1007/s 12328-019-00999-331161540 · doi ↗ · pubmed ↗